
Dynamic In Vivo Profiling of DNA Damage and Repair after Radiotherapy Using Canine Patients as a Model

Abstract
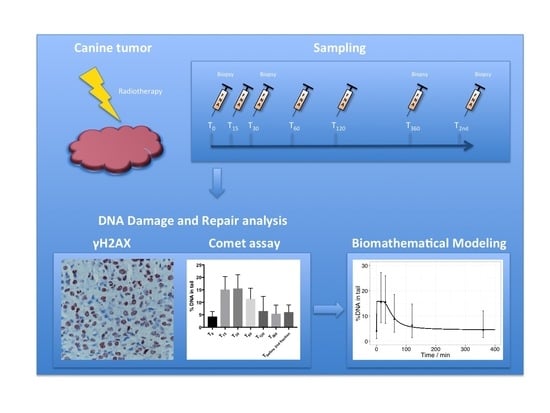
:
1. Introduction
2. Results
2.1. Patient, Tumor, and Radiation Dose Characteristics
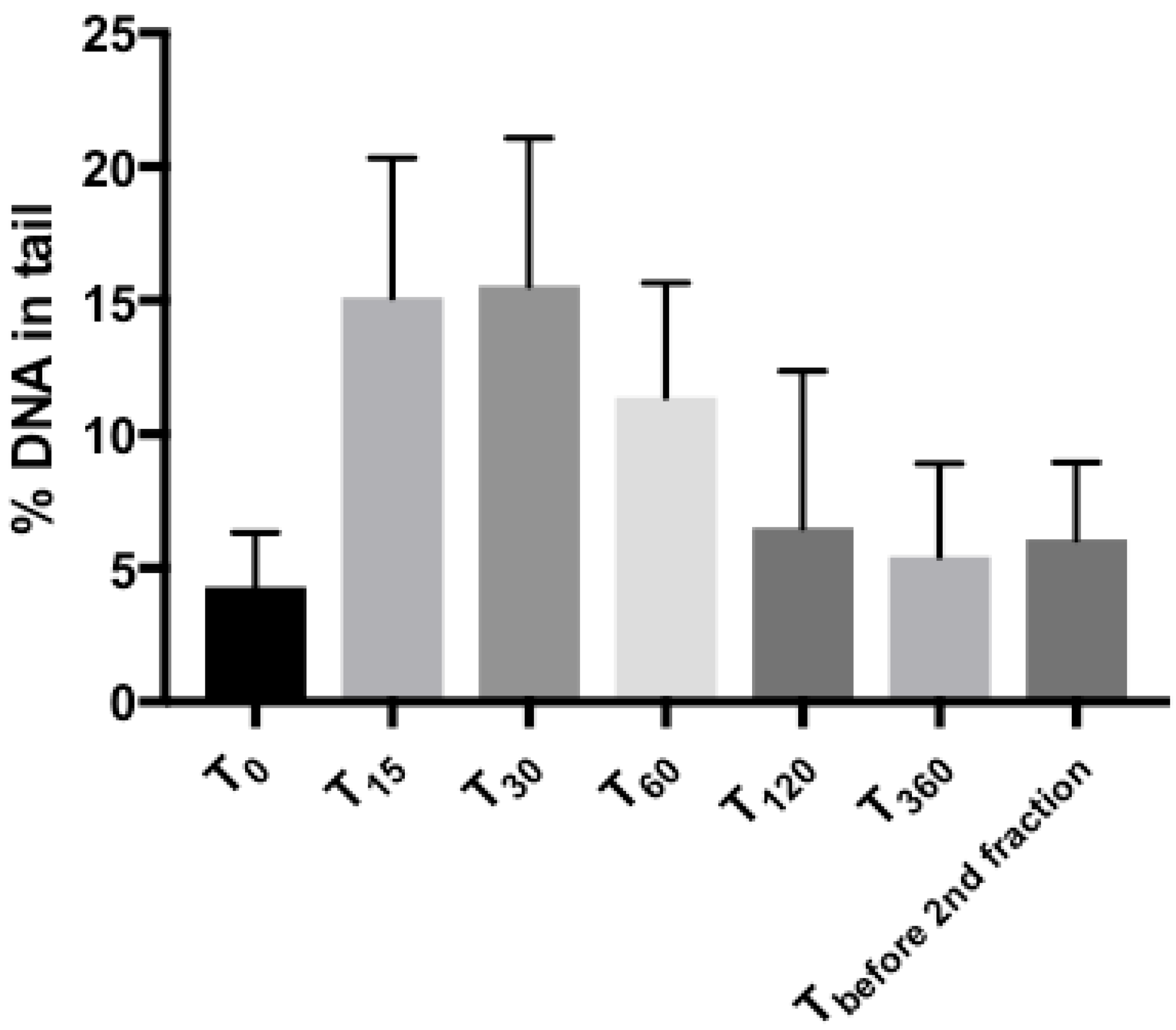
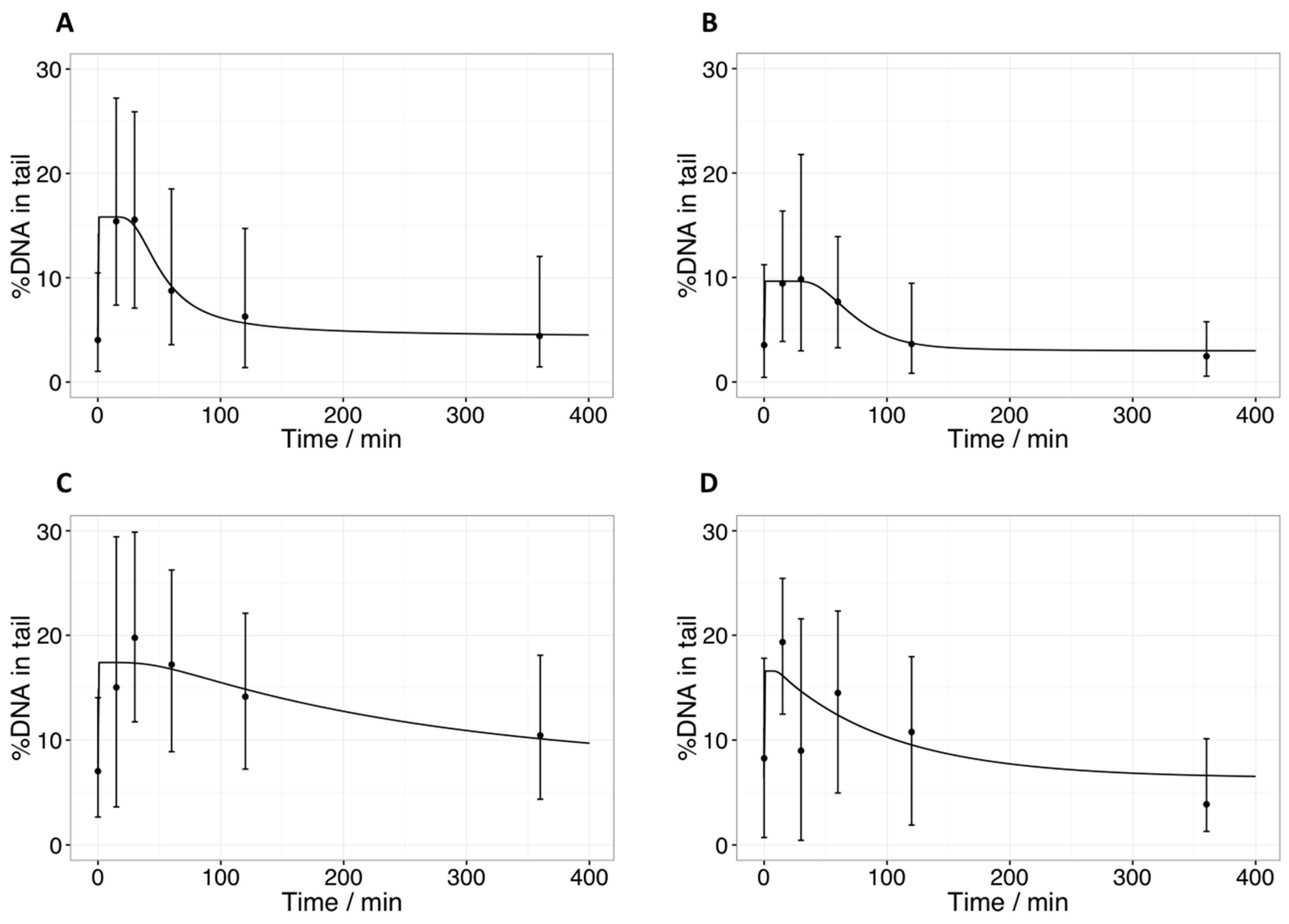
2.2. Comet Assay: Tail Intensities (%DNA in Tail) after Radiotherapy in Tumor Samples
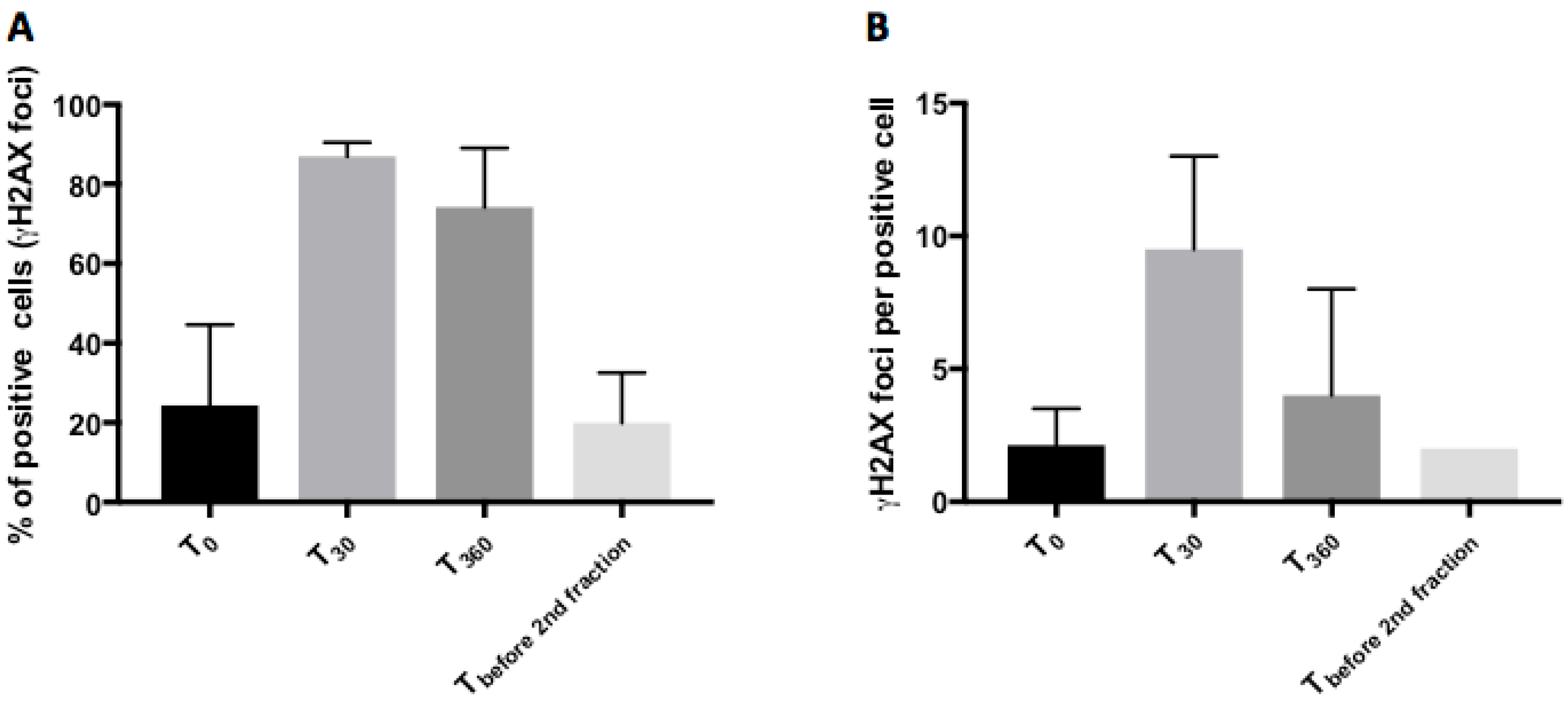
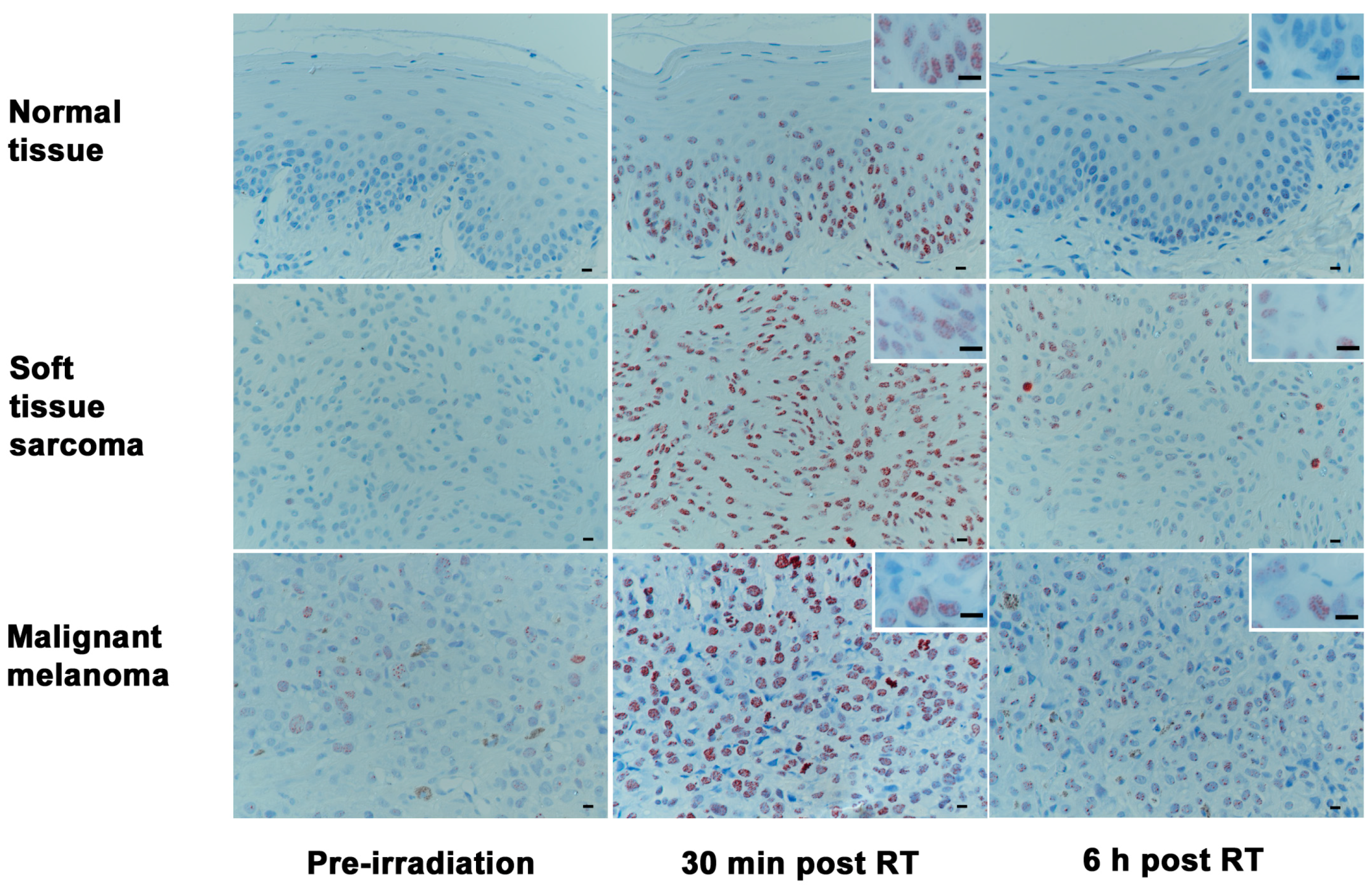
2.3. Phosphorylated H2AX (γH2AX): Number of Positive Cells and Foci per Cell after Radiotherapy in Tumor Samples
2.4. Covariate Analysis
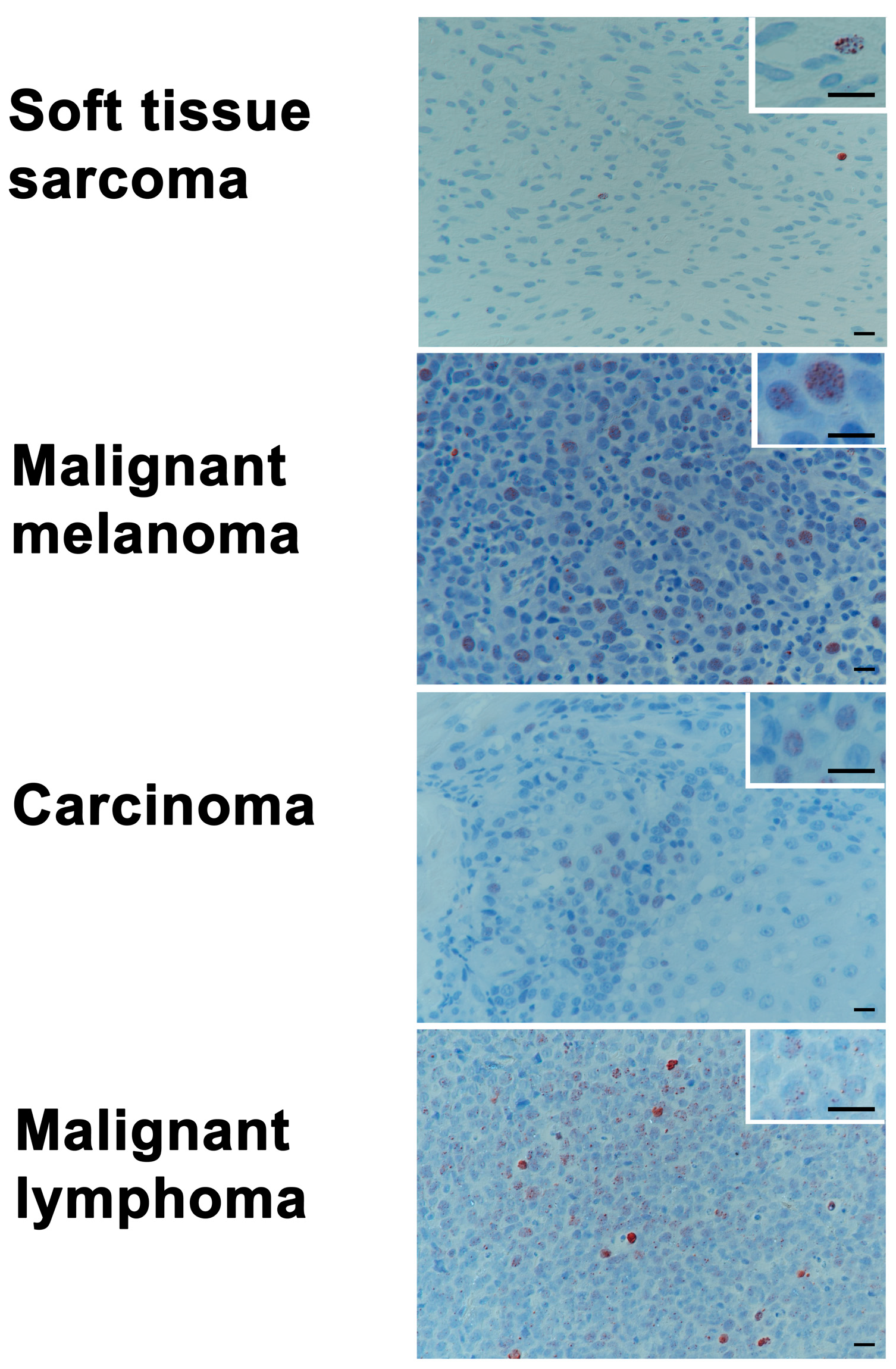
2.5. Baseline Levels of γH2AX in Tumor Biopsy Samples of Various Histologies
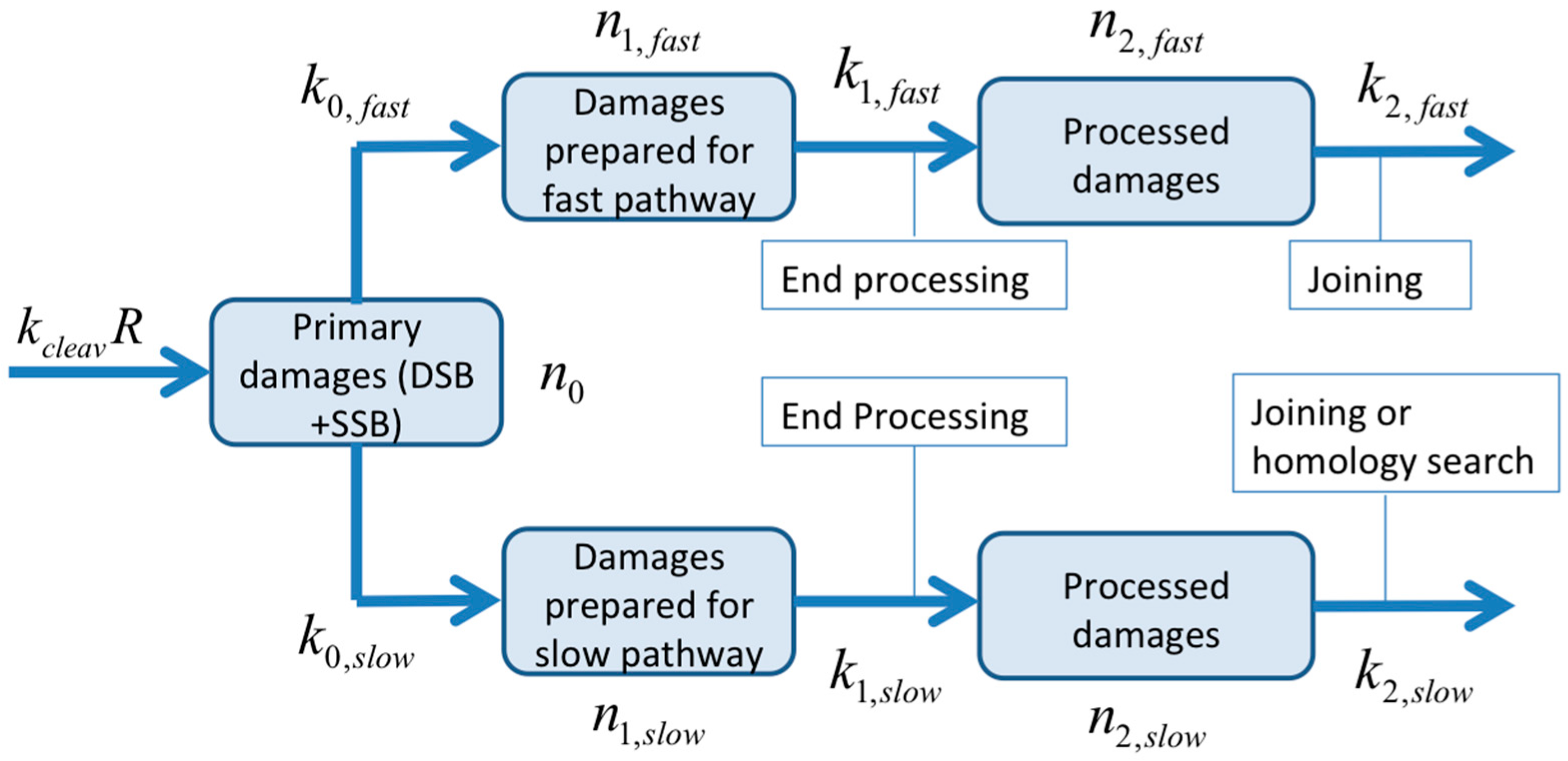
2.6. Quantification of the Time-Course of DNA Damage and Repair Using Novel Mathematical Modeling
3. Discussion
4. Materials and Methods
4.1. Patients and Sampling Procedures
4.2. Treatment
4.3. Alkaline Comet Assay
4.4. γH2AX Immunohistochemistry
4.5. Bio-Mathematical Modeling
4.6. Statistical Methods
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koch, U.; Hohne, K.; von Neubeck, C.; Thames, H.D.; Yaromina, A.; Dahm-Daphi, J.; Baumann, M.; Krause, M. Residual γH2AX foci predict local tumour control after radiotherapy. Radiother. Oncol. 2013, 108, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Menegakis, A.; de Colle, C.; Yaromina, A.; Hennenlotter, J.; Stenzl, A.; Scharpf, M.; Fend, F.; Noell, S.; Tatagiba, M.; Brucker, S.; et al. Residual γH2AX foci after ex vivo irradiation of patient samples with known tumour-type specific differences in radio-responsiveness. Radiother. Oncol. 2015, 116, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Menegakis, A.; Yaromina, A.; Eicheler, W.; Dorfler, A.; Beuthien-Baumann, B.; Thames, H.D.; Baumann, M.; Krause, M. Prediction of clonogenic cell survival curves based on the number of residual DNA double strand breaks measured by γH2AX staining. Int. J. Radiat. Biol. 2009, 85, 1032–1041. [Google Scholar] [CrossRef] [PubMed]
- Wouters, B.G.; Begg, A.C. Irradiation-induced damage and the DNA damage response. In Basic Clinical Radiobiology, 4th ed.; Joiner, M., Van der Kogel, A., Eds.; CRC Press: Boca Raton, FL, USA, 2009; pp. 11–26. [Google Scholar]
- Menegakis, A.; von Neubeck, C.; Yaromina, A.; Thames, H.; Hering, S.; Hennenlotter, J.; Scharpf, M.; Noell, S.; Krause, M.; Zips, D.; et al. γH2AX assay in ex vivo irradiated tumour specimens: A novel method to determine tumour radiation sensitivity in patient-derived material. Radiother. Oncol. 2015, 116, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Bowman, K.J.; Al-Moneef, M.M.; Sherwood, B.T.; Colquhoun, A.J.; Goddard, J.C.; Griffiths, T.R.; Payne, D.; Singh, S.; Butterworth, P.C.; Khan, M.A.; et al. Comet assay measures of DNA damage are predictive of bladder cancer cell treatment sensitivity in vitro and outcome in vivo. Int. J. Cancer 2014, 134, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.J.; Ashworth, A. Bringing DNA repair in tumors into focus. Clin. Cancer Res. 2009, 15, 3241–3243. [Google Scholar] [CrossRef] [PubMed]
- Graeser, M.; McCarthy, A.; Lord, C.J.; Savage, K.; Hills, M.; Salter, J.; Orr, N.; Parton, M.; Smith, I.E.; Reis-Filho, J.S.; et al. A marker of homologous recombination predicts pathologic complete response to neoadjuvant chemotherapy in primary breast cancer. Clin. Cancer Res. 2010, 16, 6159–6168. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, F.; Huang, Q.; Shen, J.; Wolf, F.; He, Y.; Liu, X.; Hu, Y.A.; Bedford, J.S.; Li, C.Y. Quantitative, noninvasive imaging of radiation-induced DNA double-strand breaks in vivo. Cancer Res. 2011, 71, 4130–4137. [Google Scholar] [CrossRef] [PubMed]
- Somaiah, N.; Yarnold, J.; Daley, F.; Pearson, A.; Gothard, L.; Rothkamm, K.; Helleday, T. The relationship between homologous recombination repair and the sensitivity of human epidermis to the size of daily doses over a 5-week course of breast radiotherapy. Clin. Cancer Res. 2012, 18, 5479–5488. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications, and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Fikrova, P.; Stetina, R.; Hronek, M.; Hyspler, R.; Ticha, A.; Zadak, Z. Application of the comet assay method in clinical studies. Wien. Klin. Wochenschr. 2011, 123, 693–699. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; McKeown, S.R.; McKelvey-Martin, V.J. Potential use of the comet assay in the clinical management of cancer. Mutagenesis 2008, 23, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Banath, J.P.; Olive, P.L. Expression of phosphorylated histone H2AX as a surrogate of cell killing by drugs that create DNA double-strand breaks. Cancer Res. 2003, 63, 4347–4350. [Google Scholar] [PubMed]
- Grosse, N.; van Loon, B.; Rohrer Bley, C. DNA damage response and DNA repair-dog as a model? BMC Cancer 2014, 14, 203. [Google Scholar] [CrossRef] [PubMed]
- Khanna, C.; Lindblad-Toh, K.; Vail, D.; London, C.; Bergman, P.; Barber, L.; Breen, M.; Kitchell, B.; McNeil, E.; Modiano, J.F.; et al. The dog as a cancer model. Nat. Biotechnol. 2006, 24, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Kang, H.J.; Kim, H.S.; Kim, M.J.; Heo, J.I.; Kim, J.H.; Kho, Y.J.; Kim, S.C.; Kim, J.; Park, J.B.; et al. Higher DNA repair activity is related with longer replicative life span in mammalian embryonic fibroblast cells. Biogerontology 2011, 12, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Strom, T.; Hoffe, S.E.; Fulp, W.; Frakes, J.; Coppola, D.; Springett, G.M.; Malafa, M.P.; Harris, C.L.; Eschrich, S.A.; Torres-Roca, J.F.; et al. Radiosensitivity index predicts for survival with adjuvant radiation in resectable pancreatic cancer. Radiother. Oncol. 2015, 117, 159–164. [Google Scholar] [CrossRef] [PubMed]
- McKeown, S.R.; Robson, T.; Price, M.E.; Ho, E.T.; Hirst, D.G.; McKelvey-Martin, V.J. Potential use of the alkaline comet assay as a predictor of bladder tumour response to radiation. Br. J. Cancer 2003, 89, 2264–2270. [Google Scholar] [CrossRef] [PubMed]
- Moneef, M.A.; Sherwood, B.T.; Bowman, K.J.; Kockelbergh, R.C.; Symonds, R.P.; Steward, W.P.; Mellon, J.K.; Jones, G.D. Measurements using the alkaline comet assay predict bladder cancer cell radiosensitivity. Br. J. Cancer 2003, 89, 2271–2276. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R.; Dobson, V.L.; Dusinska, M.; Kennedy, G.; Stetina, R. The comet assay: What can it really tell us? Mutat. Res. 1997, 375, 183–193. [Google Scholar] [CrossRef]
- Klaude, M.; Eriksson, S.; Nygren, J.; Ahnstrom, G. The comet assay: Mechanisms and technical considerations. Mutat. Res. 1996, 363, 89–96. [Google Scholar] [CrossRef]
- Olive, P.L. The role of DNA single- and double-strand breaks in cell killing by ionizing radiation. Radiat. Res. 1998, 150 (Suppl. S5), S42–S51. [Google Scholar] [CrossRef] [PubMed]
- Dunne, A.L.; Price, M.E.; Mothersill, C.; McKeown, S.R.; Robson, T.; Hirst, D.G. Relationship between clonogenic radiosensitivity, radiation-induced apoptosis and DNA damage/repair in human colon cancer cells. Br. J. Cancer 2003, 89, 2277–2283. [Google Scholar] [CrossRef] [PubMed]
- Bonner, W.M.; Redon, C.E.; Dickey, J.S.; Nakamura, A.J.; Sedelnikova, O.A.; Solier, S.; Pommier, Y. γH2AX and cancer. Nat. Rev. Cancer 2008, 8, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Boon, C.; Redon, C.; Bonner, W.M. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J. Cell Biol. 1999, 146, 905–916. [Google Scholar] [CrossRef] [PubMed]
- Mirzayans, R.; Severin, D.; Murray, D. Relationship between DNA double-strand break rejoining and cell survival after exposure to ionizing radiation in human fibroblast strains with differing ATM/p53 status: Implications for evaluation of clinical radiosensitivity. Int. J. Radiat. Oncol. Biol. Phys. 2006, 66, 1498–1505. [Google Scholar] [CrossRef] [PubMed]
- Derenzini, E.; Agostinelli, C.; Imbrogno, E.; Iacobucci, I.; Casadei, B.; Brighenti, E.; Righi, S.; Fuligni, F.; Di Rora, A.G.L.; Ferrari, A.; et al. Constitutive activation of the DNA damage response pathway as a novel therapeutic target in diffuse large B-cell lymphoma. Oncotarget 2015, 6, 6553–6569. [Google Scholar] [CrossRef] [PubMed]
- Fontana, A.O.; Augsburger, M.A.; Grosse, N.; Guckenberger, M.; Lomax, A.J.; Sartori, A.A.; Pruschy, M.N. Differential DNA repair pathway choice in cancer cells after proton- and photon-irradiation. Radiother. Oncol. 2015, 116, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Maeda, J.; Froning, C.E.; Brents, C.A.; Rose, B.J.; Thamm, D.H.; Kato, T.A. Intrinsic Radiosensitivity and Cellular Characterization of 27 Canine Cancer Cell Lines. PLoS ONE 2016, 11, e0156689. [Google Scholar] [CrossRef] [PubMed]
- Mariotti, L.G.; Pirovano, G.; Savage, K.I.; Ghita, M.; Ottolenghi, A.; Prise, K.M.; Schettino, G. Use of the γ-H2AX assay to investigate DNA repair dynamics following multiple radiation exposures. PLoS ONE 2013, 8, e79541. [Google Scholar] [CrossRef] [PubMed]
- Menegakis, A.; Eicheler, W.; Yaromina, A.; Thames, H.D.; Krause, M.; Baumann, M. Residual DNA double strand breaks in perfused but not in unperfused areas determine different radiosensitivity of tumours. Radiother. Oncol. 2011, 100, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Paris, L.; Cordelli, E.; Eleuteri, P.; Grollino, M.G.; Pasquali, E.; Ranaldi, R.; Meschini, R.; Pacchierotti, F. Kinetics of γ-H2AX induction and removal in bone marrow and testicular cells of mice after X-ray irradiation. Mutagenesis 2011, 26, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Olive, P.L.; Banath, J.P. Phosphorylation of histone H2AX as a measure of radiosensitivity. Int. J. Radiat. Oncol. Biol. Phys. 2004, 58, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Rube, C.E.; Grudzenski, S.; Kuhne, M.; Dong, X.; Rief, N.; Lobrich, M.; Rube, C. DNA double-strand break repair of blood lymphocytes and normal tissues analysed in a preclinical mouse model: Implications for radiosensitivity testing. Clin. Cancer Res. 2008, 14, 6546–6555. [Google Scholar] [CrossRef] [PubMed]
- Sedelnikova, O.A.; Bonner, W.M. γH2AX in cancer cells: A potential biomarker for cancer diagnostics, prediction and recurrence. Cell Cycle 2006, 5, 2909–2913. [Google Scholar] [PubMed]
- Novik, K.L.; Spinelli, J.J.; Macarthur, A.C.; Shumansky, K.; Sipahimalani, P.; Leach, S.; Lai, A.; Connors, J.M.; Gascoyne, R.D.; Gallagher, R.P.; et al. Genetic variation in H2AFX contributes to risk of non-Hodgkin lymphoma. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1098–1106. [Google Scholar] [CrossRef] [PubMed]
- Bodgi, L.; Foray, N. The nucleo-shuttling of the ATM protein as a basis for a novel theory of radiation response: Resolution of the linear-quadratic model. Int. J. Radiat. Biol. 2016, 92, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Scheidegger, S.; Fuchs, H.U.; Zaugg, K.; Bodis, S.; Fuchslin, R.M. Using state variables to model the response of tumour cells to radiation and heat: A novel multi-hit-repair approach. Comput. Math. Methods Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Zhu, W.; Diao, H.; Zhou, C.; Chen, F.F.; Yang, J. A comparative study of using comet assay and γH2AX foci formation in the detection of N-methyl-N′-nitro-N-nitrosoguanidine-induced DNA damage. Toxicol. In Vitro 2006, 20, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Begg, A.C. Predicting response to radiotherapy: Evolutions and revolutions. Int. J. Radiat. Biol. 2009, 85, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Torres-Roca, J.F.; Stevens, C.W. Predicting response to clinical radiotherapy: Past, present, and future directions. Cancer Control. 2008, 15, 151–156. [Google Scholar] [PubMed]
- Olive, P.L.; Banath, J.P. The comet assay: A method to measure DNA damage in individual cells. Nat. Protoc. 2006, 1, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Qvarnstrom, O.F.; Simonsson, M.; Johansson, K.A.; Nyman, J.; Turesson, I. DNA double strand break quantification in skin biopsies. Radiother. Oncol. 2004, 72, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [PubMed]
- MacNeill, A.L. Cytology of canine and feline cutaneous and subcutaneous lesions and lymph nodes. Top. Companion Anim. Med. 2011, 26, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Cancedda, S.; Marconato, L.; Meier, V.; Laganga, P.; Roos, M.; Leone, V.F.; Rossi, F.; Rohrer Bley, C. Hypofractionated radiotherapy for macroscopic canine soft tissue sarcoma: A retrospective study of 50 cases treated with a 5 × 6 Gy protocol with or without metronomic chemotherapy. Vet. Radiol. Ultrasound 2015, 57, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Keyerleber, M.A.; McEntee, M.C.; Farrelly, J.; Podgorsak, M. Completeness of reporting of radiation therapy planning, dose, and delivery in veterinary radiation oncology manuscripts from 2005 to 2010. Vet. Radiol. Ultrasound 2012, 53, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Rohrer Bley, C.; Blattmann, H.; Roos, M.; Sumova, A.; Kaser-Hotz, B. Assessment of a radiotherapy patient immobilization device using single plane port radiographs and a remote computed tomography scanner. Vet. Radiol. Ultrasound 2003, 44, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Iliakis, G.; Wu, W.; Wang, M. DNA double strand break repair inhibition as a cause of heat radiosensitization: Re-evaluation considering backup pathways of NHEJ. Int. J. Hyperther. 2008, 24, 17–29. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time-Points | Difference of %DNA in Tail (95% CI) | p-Value |
|---|---|---|
| T0 → T15 | 10.80 (6.56; 15.04) | p = 0.001 |
| T0 → T30 | 11.24 (6.38; 16.10) | p = 0.001 |
| T0 → T60 | 7.11 (4.52; 9.71) | p < 0.001 |
| T0 → T120 | 2.47 (−1.41; 6.36) | p = 0.172 |
| T0 → T360 | 1.24 (−0.86; 3.33) | p = 0.185 |
| T0 → Tbefore 2nd fraction | 1.56 (−0.87; 3.99) | p = 0.168 |
| (a) | ||
| Time-Points | Median γH2AX Positive Cells (%; 95% CI) | p-Value, Compared to T0 |
| T0 | 20.17 (3.89; 62.36) | |
| T30 | 87.18 (81.75; 91.78) | p < 0.001 |
| T360 | 78.48 (41.88; 85.67) | p < 0.001 |
| Tbefore 2nd fraction | 15.61 (9.80; 41.44) | p = 0.764 |
| (b) | ||
| Time-Points | Median γH2AX-Foci Per Nucleus (95% CI) | p-Value, Compared to T0 |
| T0 | 2.13 (2.00; 3.50) | |
| T30 | 8.00 (6.00; 15.00) | p < 0.001 |
| T360 | 4.00 (3.00; 8.00) | p = 0.002 |
| Tbefore 2nd fraction | 2.00 (2.00; 2.00) | p = 0.163 |
| Tumor Type | Age (Years) Mean (SD; Range) | Weight (kg) Mean (SD; Range) | Gender | Median Positive Cells (%) | 95% CI | p-Value | Median Foci/Tumor | 95% CI | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Overall (n = 83) | 9.89 (±2.92; 1.00–15.00) | 23.70 (±10.90; 3.60–55.00) | f = 14 fs = 27 m = 19 mn = 23 | 20.72 | 21.88; 24.29 | 0.014 | 2.50 | 2.40; 2.90 | <0.001 |
| Soft tissue sarcoma (n = 16) | 9.74 (±1.80; 5.92–12.83 | 30.40 (±9.88; 14.20–50.70) | f = 3 fs = 7 m = 3 mn = 3 | 13.41 | 7.42; 20.59 | 2.00 | 1.50; 2.47 | ||
| Malignant melanoma (n = 19) | 11.71 (±2.59; 6.00–15.00) | 19.24 (±8.78; 3.60–30.50) | f = 2 fs = 4 m = 7 mn = 6 | 15.35 | 12.25; 28.22 | 2.50 | 2.14; 2.99 | ||
| Carcinoma (n = 25) | 9.48 (±3.33; 1.00–15.00) | 20.79 (±10.23; 4.50–35.50) | f = 6 fs = 6 m = 8 mn = 5 | 18.12 | 14.55; 28.67 | 2.00 | 2.03; 2.81 | ||
| Malignant lymphoma (n = 23) | 8.94 (±2.67; 3.58–13.75) | 25.88 (±11.04; 9.00–55.00) | f = 3 fs = 10 m = 1 mn = 9 | 31.30 | 24.72; 34.93 | 3.00 | 2.87; 4.04 |
| Parameter | Sarcoma, All Patients and Fractions Pooled | Patient 1, Fraction 1 | Patient 2, Fraction 1 | Patient 2, Fraction 2 |
|---|---|---|---|---|
| /min−1 | 1.93 (1.96) | 1.12 | 1.71 | 1.71 |
| /min−1 | 3.93·10−1 (1.59 × 10−1) | 3.76 × 10−2 | 4.35 × 10−3 | 5.24 × 10−2 |
| /min−1 | 2.92·10−2 (3.44 × 10−2) | 2.55 × 10−2 | 5.24 × 10−2 | 1.39 × 10−1 |
| /min−1 | 1.00∙10−2 (9.01 × 10−3) | 8.45 × 10−2 | 2.65 × 10−2 | 1.01 |
| /min−1 | - (3.22 × 10−3) | - | - | - |
| /min−1 | - (1.01 × 10−3) | - | - | - |
| /min−1 | - (1.23 × 102) | - | - | - |
| /min | - (7.07) | - | - | - |
| /min | 1.28·101 (1.09 × 101) | 1.93 × 101 | 8.51 × 10−1 | 3.77 |
| /% DNA | 4.23 (4.09) | 2.94 | 7.12 | 6.35 |
| Error of fit | 1.20 (1.03) | 7.31 × 10−1 | 1.23 × 101 | 6.97 × 101 |
| Antigen | Vendor | Antibody Type | Catalogue No./Clone | Dilution, Incubation Conditions | Pre-Treatment | Visualization Method | Positive Control |
|---|---|---|---|---|---|---|---|
| γH2AX Ser 139 | Millipore Temecula, CA, USA | mouse mAb, IgG1 | 05-0636/clone JBW301 | 1:200, 1.3 h, room temperature | HIER *, 20 min 98 °C, citrate buffer pH 6.0 | Envision Kit (Dako) | Irradiated canine tumor tissue |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulz, N.; Chaachouay, H.; Nytko, K.J.; Weyland, M.S.; Roos, M.; Füchslin, R.M.; Guscetti, F.; Scheidegger, S.; Rohrer Bley, C. Dynamic In Vivo Profiling of DNA Damage and Repair after Radiotherapy Using Canine Patients as a Model. Int. J. Mol. Sci. 2017, 18, 1176. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061176
Schulz N, Chaachouay H, Nytko KJ, Weyland MS, Roos M, Füchslin RM, Guscetti F, Scheidegger S, Rohrer Bley C. Dynamic In Vivo Profiling of DNA Damage and Repair after Radiotherapy Using Canine Patients as a Model. International Journal of Molecular Sciences. 2017; 18(6):1176. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061176
Chicago/Turabian StyleSchulz, Nadine, Hassan Chaachouay, Katarzyna J. Nytko, Mathias S. Weyland, Malgorzata Roos, Rudolf M. Füchslin, Franco Guscetti, Stephan Scheidegger, and Carla Rohrer Bley. 2017. "Dynamic In Vivo Profiling of DNA Damage and Repair after Radiotherapy Using Canine Patients as a Model" International Journal of Molecular Sciences 18, no. 6: 1176. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061176