Taxifolin Activates the Nrf2 Anti-Oxidative Stress Pathway in Mouse Skin Epidermal JB6 P+ Cells through Epigenetic Modifications

Abstract
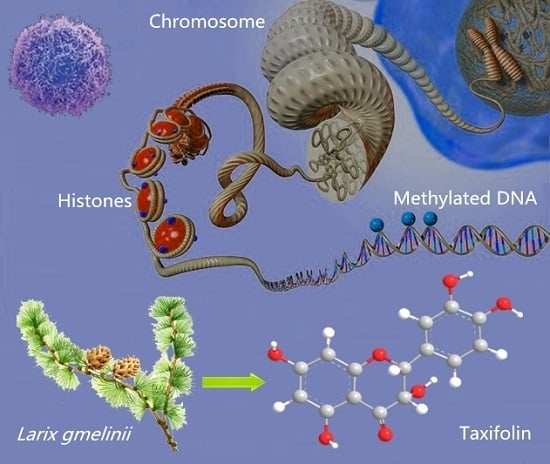
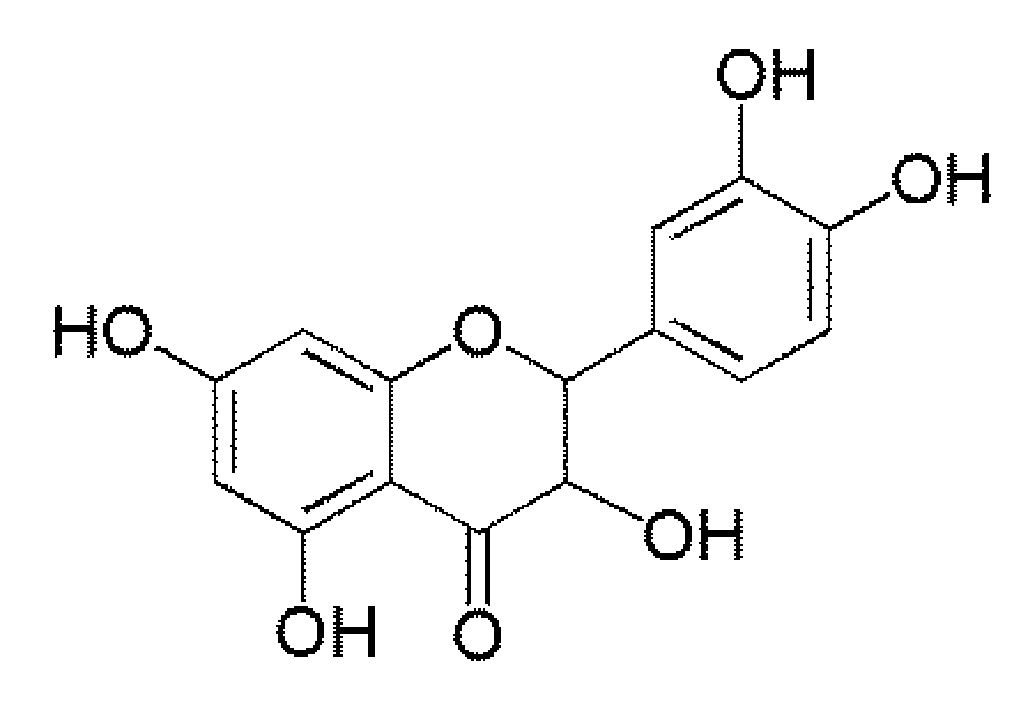
:
1. Introduction
2. Results
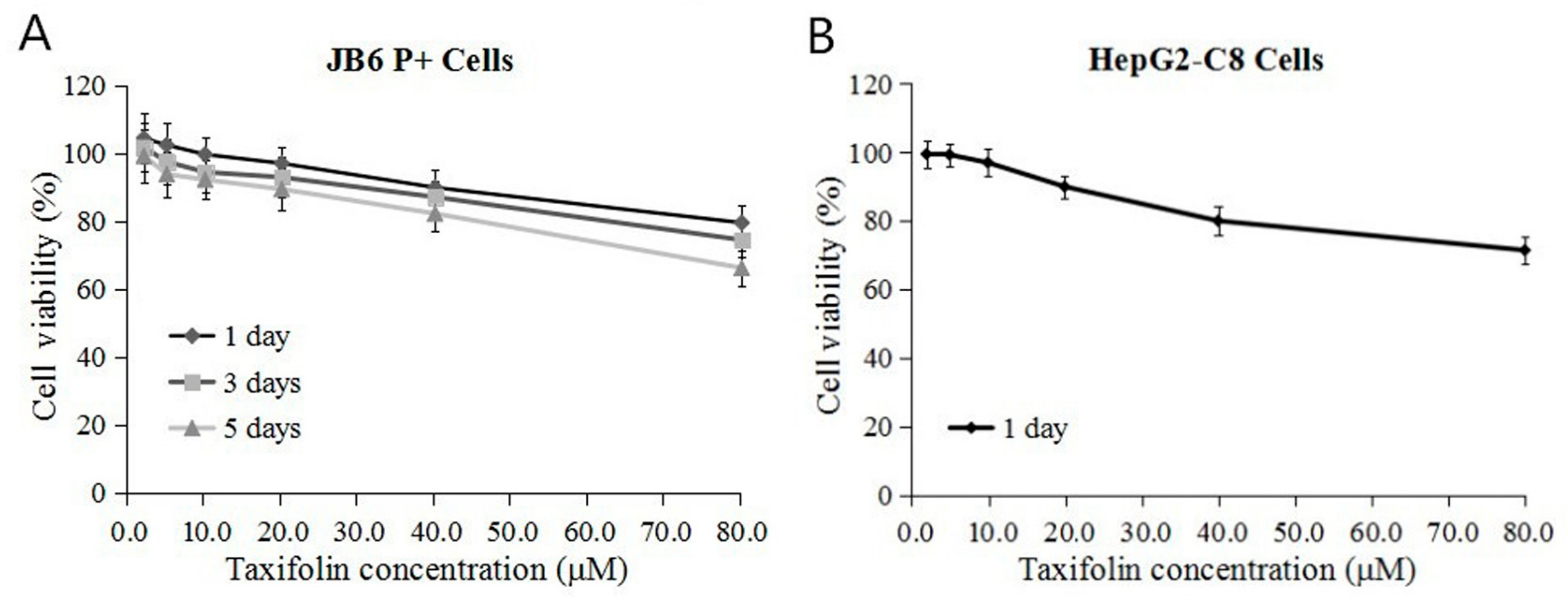
2.1. Cytotoxicity of TAX in JB6 P+ Cells and HepG2-C8 Cells
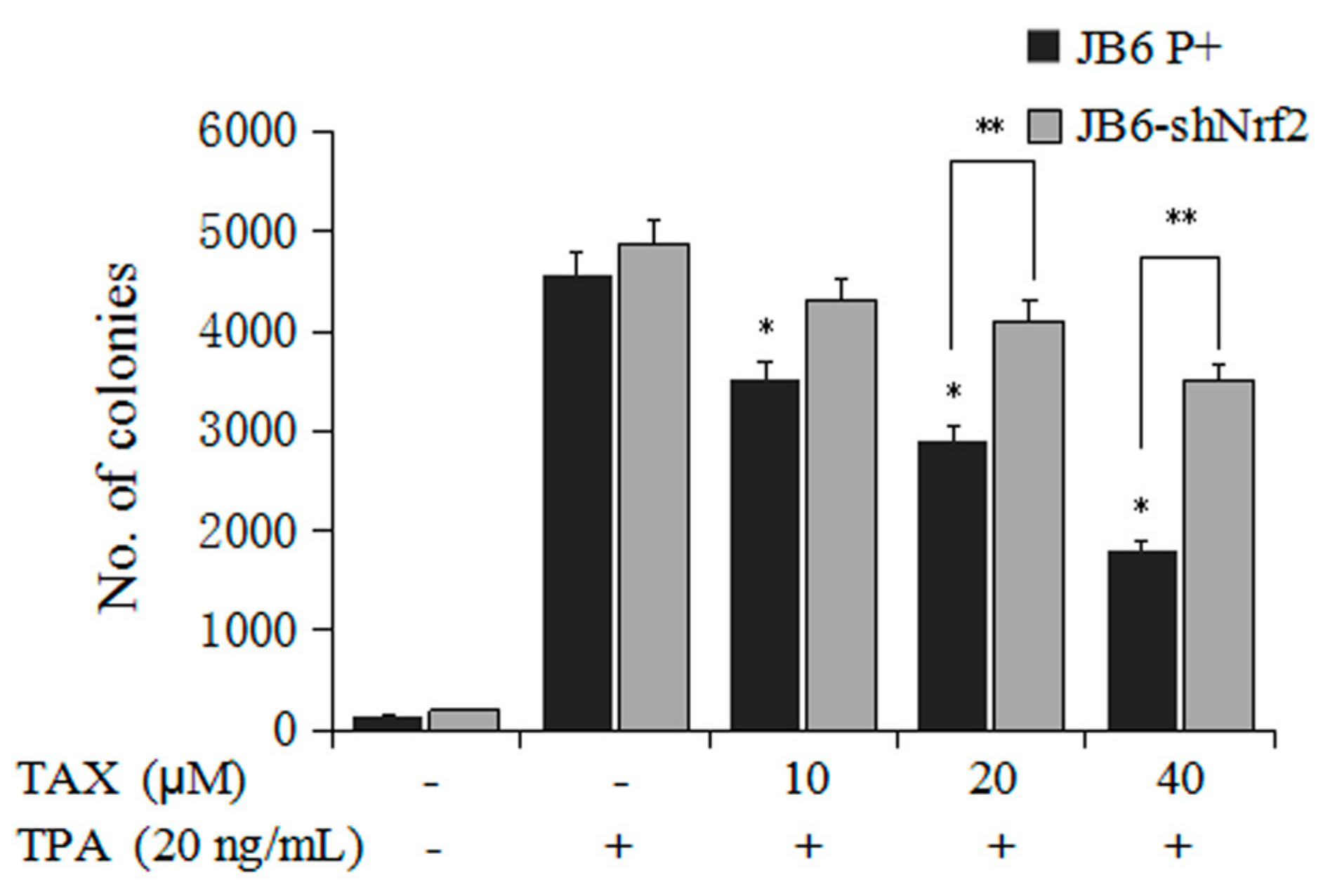
2.2. TAX Inhibits TPA-Induced JB6 P+ Cells and JB6-shNrf2 Cells Transformation
2.3. TAX Induces ARE-Luciferase Reporter Activity
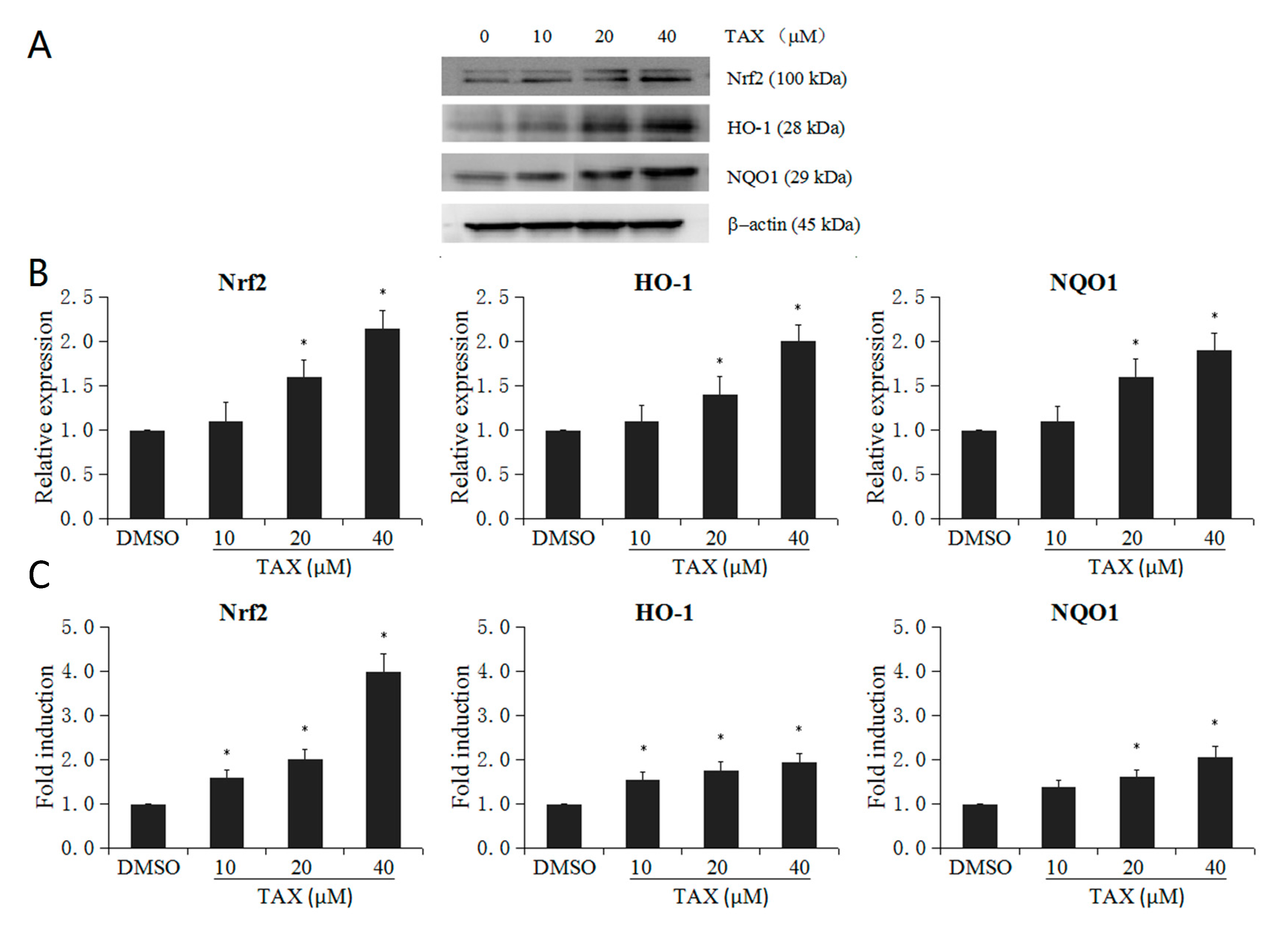
2.4. TAX Upregulates the mRNA and Protein Levels of Nrf2 Target Enzymes in JB6 P+ Cells
2.5. TAX Decreases the Proportion of Methylated CpG in the Nrf2 Gene Promoter Region
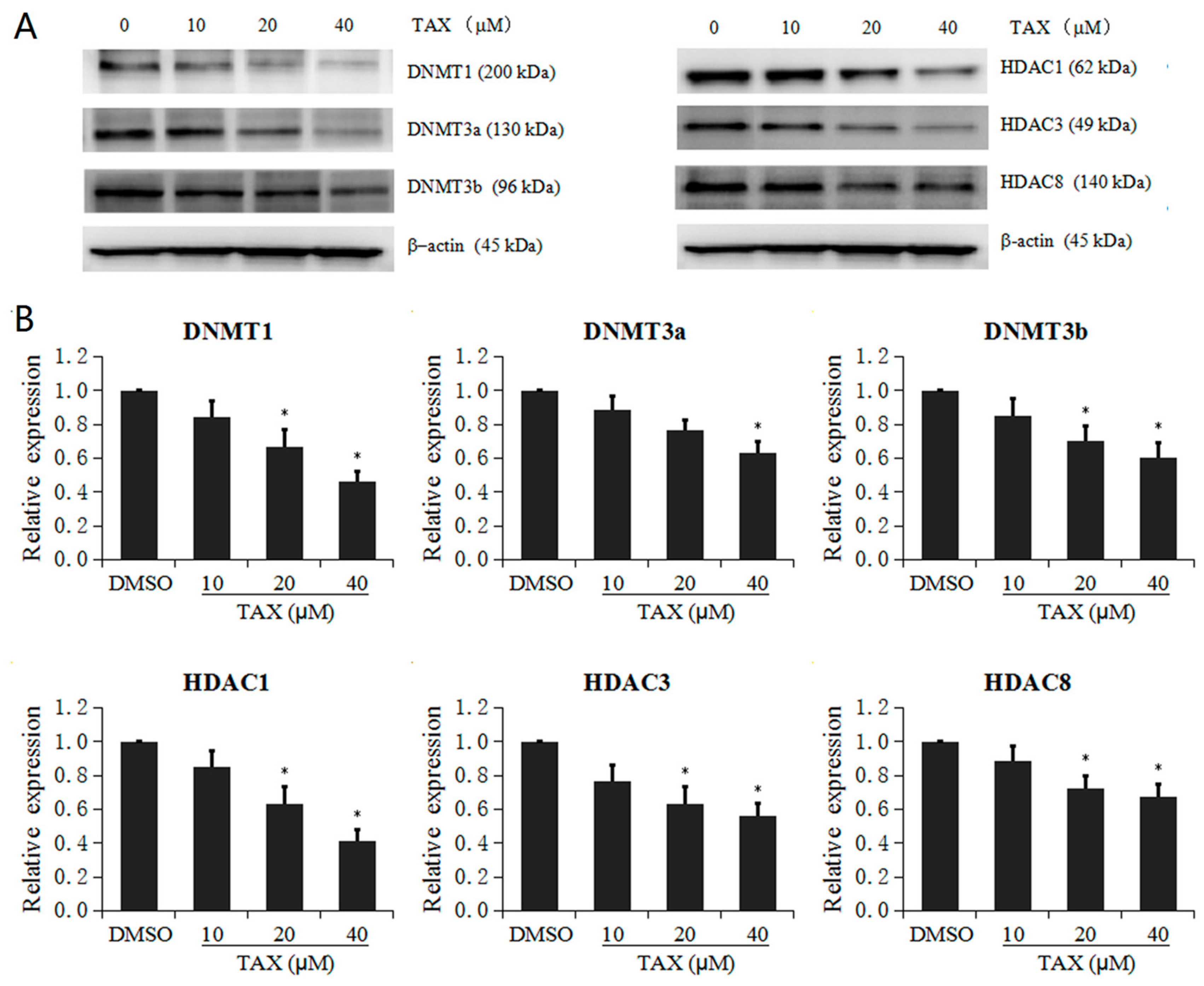
2.6. TAX Inhibits the Protein Expression of Epigenetic Modification Enzymes in JB6 P+ Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals and Biochemicals
4.2. Cell Culture and Treatment
4.3. Cell Viability Tests
4.4. Luciferase Reporter Activity Assay
4.5. Anchorage-Independent Cell Neoplastic Transformation Assay
4.6. Protein Lysate Preparation and Western Blotting
4.7. RNA Extraction and qRT-PCR
4.8. DNA Isolation and Bisulfite Genomic Sequencing
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ARE | Antioxidant response element |
| ATCC | American type culture collection |
| DMEM | Dulbecco’s modification of Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| DNMT | DNA methyltransferase |
| BME | Eagle’s basal medium |
| FBS | Fetal bovine serun |
| HDAC | Histone deacetylase |
| Keap1 | Kelch-like ECH-associating protein 1 |
| HO-1 | Heme oxygenase-1 |
| MEM | Minimum essential medium |
| MTS | 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium |
| NQO1 | NAD(P)H quinone oxidoreductase 1 |
| Nrf2 | Nuclear factor erythroid-2 related factor 2 |
| ROS | Reactive oxygen species |
| SD | Standard deviation |
| TAX | Taxifolin |
| TPA | 12-O-tetradecanoylphorbol-13-acetate |
| TSA | Trichostatin A |
| UV | Ultraviolet |
References
- Reczek, C.R.; Chandel, N.S. ROS-dependent signal transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Dhalla, N.S.; Temsha, R.M.; Netticadan, T. Role of oxidative stress in cardiovascular diseases. J. Hypertens. 2000, 18, 655–673. [Google Scholar] [CrossRef] [PubMed]
- Narendhirakannan, R.T.; Hannah, M.A. Oxidative stress and skin cancer: An overview. Ind. J. Clin. Biochem. 2013, 28, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Godic, A.; Poljšak, B.; Adamic, M.; Dahmane, R. The role of antioxidants in skin cancer prevention and treatment. Oxid. Med. Cell Longev. 2014, 2014, 443–479. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A. Skin cancer as occupational disease. Hautarzt 2016, 67, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, J.K.; Srivastava, D.; Moy, R.L.; Lin, H.J.; Kouba, D.J. The molecular genetics underlying basal cell carcinoma pathogenesis and links to targeted therapeutics. J. Am. Acad. Dermatol. 2012, 66, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Rogers, H.W.; Weinstock, M.A.; Harris, A.R.; Hinckley, M.R.; Feldman, S.R.; Fleischer, A.B.; Coldiron, B.M. Incidence estimate of nonmelanoma skinv cancer in the United States. Arch. Dermatol. 2010, 146, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.C.; McDonald, P.R.; Liu, J.; Klaassen, C.D. Screening of natural compounds as activators of the Keap1-Nrf2 pathway. Planta Med. 2014, 80, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Gordan, J.D.; Jin, J.; Harper, J.W.; Diehl, J.A. The Keap1-BTB protein is an adaptor that bridges Nrf2 to a Cul3-based E3 ligase: Oxidative stress sensing by a Cul3-Keap1 ligase. Mol. Cell Biol. 2004, 4, 8477–8486. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Li, L.; Iwamoto, N.; Nakajima-Takagi, Y.; Kaneko, H.; Nakayama, Y.; Eguchi, M.; Wada, Y.; Kumagai, Y.; Yamamoto, M. The antioxidant defense system Keap1-Nrf2 comprises a multiple sensing mechanism for responding to a wide range of chemical compounds. Mol. Cell Biol. 2009, 29, 493–502. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Lamont, D.J.; Beattie, K.A.; Hayes, J.D. Keap1 perceives stress via three sensors for the endogenous signaling molecules nitric oxide, zinc, and alkenals. Proc. Natl. Acad. Sci. USA 2010, 107, 18838–18843. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Y.; Zhang, C.; Lee, J.H.; Shu, L.; Wu, T.Y.; Khor, T.O.; Conney, A.H.; Lu, Y.P.; Kong, A.N. Requirement and epigenetics re-programming of Nrf2 in suppression of tumor promoter TPA-induced mouse skin cell transformation by sulforaphane. Cancer Prev. Res. 2014, 7, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef] [PubMed]
- Iida, K.; Itoh, K.; Kumagai, Y.; Oyasu, R.; Hattori, K. Nrf2 is essential for the chemopreventive efficacy of oltipraz against urinary bladder carcinogenesis. Cancer Res. 2004, 64, 6424–6431. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Khor, T.O.; Lee, J.H.; Boyanapalli, S.S.; Huang, Y.; Wu, T.Y.; Saw, C.L.; Cheung, K.L.; Kong, A.N. Epigenetic CpG demethylation of the promoter and reactivation of the expression of Neurog1 by curcumin in prostate LNCaP cells. AAPS J. 2011, 13, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.A.; Baylin, S.B. The fundamental role of epigenetic events in cancer. Nat. Rev. Genet. 2002, 3, 415–428. [Google Scholar] [PubMed]
- Yu, S.; Khor, T.O.; Cheung, K.L.; Li, W.; Wu, T.Y.; Huang, Y.; Foster, B.A.; Kan, Y.W.; Kong, A.N. Nrf2 expression is regulated by epigenetic mechanisms in prostate cancer of TRAMP mice. PLoS ONE 2010, 5, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Kong, A.N. Dietary phytochemicals and cancer prevention: Nrf2 signaling, epigenetics, and cell death mechanisms in blocking cancer initiation and progression. Pharmacol. Ther. 2013, 137, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Y.; Shu, L.; Khor, T.O.; Lee, J.H.; Fuentes, F.; Kong, A.N. A perspective on dietary phytochemicals and cancer chemoprevention: Oxidative stress, Nrf2, and epigenomics. Top. Curr. Chem. 2013, 329, 133–162. [Google Scholar] [PubMed]
- Li, J.X.; Mottamal, M.; Li, H.T.; Liu, K.D.; Zhu, F.; Cho, Y.Y.; Sosa, C.P.; Zhou, K.Y.; Bowden, G.T.; Bode, A.M.; et al. Quercetin-3-methyl ether suppresses proliferation of mouse epidermal JB6 P+ cells by targeting ERKs. Carcinoqenesis 2012, 33, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, A.E. Dihydroquercetin: More than just an impurity? Eur. J. Pharmacol. 2012, 684, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Willfor, S.; Ali, M.; Karonen, M.; Reunanen, M.; Arfan, M.; Harlamow, R. Extractives in bark of different conifer species growing in Pakistan. Holzforschung 2009, 63, 551–558. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vagen, I.M. Onions: A source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.P.; Zhang, X.; Cui, T.Q.; Zhou, H. Taxifolin protects against cardiac hypertrophy and fibrosis during biomechanical stress of pressure overload. Toxicol. Appl. Pharmacol. 2015, 287, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, H.; Mochida, Y.; Sakai, S.; Masuda, H.; Tamura, Y.; Mizutani, K. Protection against oxidative damage by dihydroflavonols in Engelhardtia chrysolepis. Biosci. Biotechnol. Biochem. 1996, 60, 945–948. [Google Scholar] [CrossRef] [PubMed]
- Kostyuk, V.A.; Kraemer, T.; Sies, H.; Schewe, T. Myeloperoxidase /nitrite-mediated lipid peroxidation of low-density lipoprotein as modulated by flavonoids. FEBS Lett. 2003, 537, 146–150. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wang, W.Y.; Chang, C.C.; Liou, K.T.; Sung, Y.J.; Liao, J.F. Taxifolin ameliorates cerebral ischemia-reperfusion injury in rats through its anti-oxidative effect and modulation of NF-κB activation. J. Biomed. Sci. 2006, 13, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Manigandan, K.; Manimaran, D.; Jayaraj, R.L.; Elangovan, N.; Dhivya, V.; Kaphle, A. Taxifolin curbs NF-κB-mediated Wnt/β-catenin signaling via up- regulating Nrf2 pathway in experimental colon carcinogenesis. Biochimie 2015, 119, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Gao, C.; Luo, M.; Wang, W. Dihydroquercetin (DHQ) induced HO-1 and NQO1 expression against oxidative stress through the Nrf2-dependent antioxidant pathway. J. Agric. Food Chem. 2013, 61, 2755–2761. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.Y.; Chen, J.J.; Zhu, P. Masayuki Fujinoa Dihydroquercetin (DHQ) ameliorated concanavalin A-induced mouse experimental fulminant hepatitis and enhanced HO-1 expression through MAPK/Nrf2 antioxidant pathway in RAW cells. Int. Immunopharmacol. 2015, 28, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Oi, N.; Chen, H.E.; Ok, K.M.; Lubet, R.A.; Bode, A.M.; Dong, Z.A. Taxifolin suppresses UV-induced skin carcinogenesis by targeting EGFR and PI3-K. Cancer Prev. Res. 2012, 5, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.A. Skin photoprotection by natural polyphenols (jb, ERK) anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.W.; Ohnishi, K.; Murakami, A.; Lee, J.S.; Kundu, J.K.; Na, H.K. Zerumbone induces heme oxygenase-1 expression in mouse skin and cultured murine epidermal cells through activation of Nrf2. Cancer Prev. Res. 2011, 4, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Kaneki, M.; Kharbanda, S.; Pandey, P.; Yoshida, K.; Takekawa, M.; Liou, J.R.; Stone, R.; Kufe, D. Functional role for protein kinase Cb as a regulator of stress-activated protein kinase activation and monocytic differentiation of myeloid leukemia cells. Mol. Cell Biol. 1999, 19, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.; Young, M.R.; Colburn, N.H. The role of AP-1, NF-κB and ROS/NOS in skin carcinogenesis: The JB6 model is predictive. Mol. Cell Biochem. 2002, 234, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Li, X.; Tang, X.W. The role of Nrf2 in carcinogenesis. Prog. Chem. 2013, 25, 1544–1552. [Google Scholar]
- Kang, K.W.; Lee, S.J.; Kim, S.G. Molecular mechanism of Nrf2 activation by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1664–1673. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Gomez, M.; Kwak, M.K.; Dolan, P.M.; Itoh, K.; Yamamoto, M.; Talalay, P.; Kensler, T.W. Sensitivity to carcinogenesis is increased and chemoprotective efficacy of enzyme inducers is lost in Nrf2 transcription factor-deficient mice. Proc. Natl. Acad. Sci. USA 2001, 98, 3410–3415. [Google Scholar] [CrossRef] [PubMed]
- Khor, T.O.; Huang, M.T.; Prawan, A.; Liu, Y.; Hao, X.; Yu, S.; Cheung, W.K.; Chan, J.Y.; Reddy, B.S.; Yang, C.S. Increased susceptibility of Nrf2 knockout mice to colitisassociated colorectal cancer. Cancer Prev. Res. 2008, 1, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Cui, X.X.; Khor, T.O.; Huang, Y.; Dipaola, R.S.; Goodin, S.; Lee, M.J.; Yang, C.S.; Kong, A.N.; Conney, A.H. Inhibitory effect of a γ-tocopherol-Rich mixture of tocopherols on the formation and growth of LNCaP prostate tumors in immunodeficient mice. Cancers 2011, 3, 3762–3772. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.Y.; Khor, T.O.; Shu, L.; Lee, J.H.; Saw, C.L.L.; Wu, T.Y. Epigenetic reactivation of Nrf2 in murine prostate cancer TRAMP C1 cells by natural phytochemicals Z-ligustilide and radix Angelica sinensis via promoter CpG demethylation. Chem. Res. Toxicol. 2013, 26, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Paredes-Gonzalez, X.; Fuentes, F.; Su, Z.Y.; Kong, A.N. Apigenin reactivates Nrf2 anti-oxidative stress signaling in mouse skin epidermal JB6 P+ cells through epigenetics modifications. AAPS J. 2014, 16, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Hong, B.; Su, Z.Y.; Zhang, C.Y.; Yang, Y.Q.; Guo, Y.; Li, W.J.; Kong, A.N. Reserpine inhibit the JB6 P+ cell transformation through epigenetic reactivation of Nrf2-mediated anti-oxidative stress pathway. AAPS J. 2016, 18, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, C.Y.; Guo, Y.; Su, Z.Y.; Yang, Y.; Shu, L.; Kong, A.N. Blocking of JB6 cell transformation by tanshinone IIA: Epigenetic reactivation of Nrf2 anti-oxidative stress pathway. AAPS J. 2014, 16, 1214–1215. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Wang, Z.B.; Mi, Y.Y.; Gao, M.J.; Lv, J.N.; Meng, Y.H.; Yang, B.Y.; Kuang, H.X. UHPLC-MS/MS determination, pharmacokinetic, and bioavailability study of taxifolin in rat plasma after oral administration of its nanodispersion. Molecules 2016, 21, 494. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Direction | Oligonucleotide Sequence 5′–3′ |
|---|---|---|
| Nrf2 | sense | 5′-AGCAGGACTGGAGAAGTT-3′ |
| antisense | 5′-TTCTTTTTCCAGCGAGGAGA-3′ | |
| HO-1 | sense | 5′-CCTCACTGGCAGGAAATCATC-3′ |
| antisense | 5′-CCTCGTGGAGACGCTTTACATA-3′ | |
| NQO1 | sense | 5′-AGCCCAGATATTGTGGCCG-3′ |
| antisense | 5′-CCTTTCAGAATGGCTGGCAC-3′ | |
| β-actin | sense | 5′-CGTTCAATACCCCAGCCATG-3′ |
| antisense | 5′-GACCCCGTCACCAGAGTCC-3′ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, H.; Tang, Z.; Zhang, C.; Wang, Z.; Li, W.; Yang, C.; Wang, Q.; Yang, B.; Kong, A.-N. Taxifolin Activates the Nrf2 Anti-Oxidative Stress Pathway in Mouse Skin Epidermal JB6 P+ Cells through Epigenetic Modifications. Int. J. Mol. Sci. 2017, 18, 1546. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071546
Kuang H, Tang Z, Zhang C, Wang Z, Li W, Yang C, Wang Q, Yang B, Kong A-N. Taxifolin Activates the Nrf2 Anti-Oxidative Stress Pathway in Mouse Skin Epidermal JB6 P+ Cells through Epigenetic Modifications. International Journal of Molecular Sciences. 2017; 18(7):1546. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071546
Chicago/Turabian StyleKuang, Haixue, Zhenqiu Tang, Chengyue Zhang, Zhibin Wang, Wenji Li, Chunjuan Yang, Qiuhong Wang, Bingyou Yang, and Ah-Ng Kong. 2017. "Taxifolin Activates the Nrf2 Anti-Oxidative Stress Pathway in Mouse Skin Epidermal JB6 P+ Cells through Epigenetic Modifications" International Journal of Molecular Sciences 18, no. 7: 1546. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071546