Novel Molecular Insights about Lactobacillar Sortase-Dependent Piliation


Department of Veterinary Biosciences, Faculty of Veterinary Medicine, University of Helsinki, Helsinki FIN-00014, Finland
Int. J. Mol. Sci. 2017, 18(7), 1551; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071551
Submission received: 14 June 2017
/
Revised: 11 July 2017
/
Accepted: 14 July 2017
/
Published: 18 July 2017
(This article belongs to the Special Issue Host-Microbe Interaction 2018)
Abstract
:One of the more conspicuous structural features that punctuate the outer cell surface of certain bacterial Gram-positive genera and species is the sortase-dependent pilus. As these adhesive and variable-length protrusions jut outward from the cell, they provide a physically expedient and useful means for the initial contact between a bacterium and its ecological milieu. The sortase-dependent pilus displays an elongated macromolecular architecture consisting of two to three types of monomeric protein subunits (pilins), each with their own specific function and location, and that are joined together covalently by the transpeptidyl activity of a pilus-specific C-type sortase enzyme. Sortase-dependent pili were first detected among the Gram-positive pathogens and subsequently categorized as an essential virulence factor for host colonization and tissue invasion by these harmful bacteria. However, the sortase-dependent pilus was rebranded as also a niche-adaptation factor after it was revealed that “friendly” Gram-positive commensals exhibit the same kind of pilus structures, which includes two contrasting gut-adapted species from the Lactobacillus genus, allochthonous Lactobacillus rhamnosus and autochthonous Lactobacillus ruminis. This review will highlight and discuss what has been learned from the latest research carried out and published on these lactobacillar pilus types.
1. Introduction
For humans, the extended arm and hand is a useful tool that lets us navigate certain social settings, often as a friendly feeling gesture epitomized by a welcoming handshake, but then sometimes as a protective-aggressive response that comes by wielding a clenched fist. In a mechanistic sense for certain microscopic bacteria, they operate in their surrounding conditions somewhat similarly through the use of long proteinaceous limb-like protrusions that reach out from the periphery of the cell wall envelope. These multi-subunit appendages are commonly known as pili (sing., pilus) or fimbriae (sing., fimbria) and can be detected in both a number of Gram-negative and Gram-positive genera and species, with each type having an archetypal polymeric structure held together by either non-covalent or covalent forces, respectively (for detailed review, see [1,2]). As a function, surface pili mediate the opening contact between the bacterial cell and its physical environment, this accomplished by virtue of their variable lengths and then by being imparted with a characteristic adhesiveness for targeted substrates. Historically, the Gram-negative pili were discovered about sixty years ago [3], and subsequent to extensive characterization, they have come to be known by their complexity, both from a functional and structural perspective (for detailed review, see [4]). On the other hand, it was about fifteen years later when pilus-like formations were first recognized among Gram-positive pathogens [5,6], and as these surface features play an essential and early role during the invasive colonization of various body regions, they were soon deemed a virulence factor of such damaging bacteria.
Gram-negative pili come in different forms and depending on which kind (i.e., chaperone-usher, curli, type IV, and the type III and IV secretion), they have diameters and lengths within the ranges of 4–14 nm and 1–20 µm, respectively [4]. Typically, they have characteristic quaternary macromolecular structures that can appear as a helical rod-like assemblage of several different pilin-called protein subunits [4]. By comparison, the architectural arrangement of the Gram-positive pilus is more simplified than its counterpart in Gram-negative bacteria, and although two varieties can be distinguished by electron microscopy (EM), one short and the other long (reaching upwards of 0.5 or 3 µm), a good deal of recent scientific interest has been drawn to the lengthier pilus structures [2]. Typically, these pili are comprised of just two to three types of pilin subunits, each with its own distinct role and location in the pilus structure. For this, a transpeptidase enzyme called the pilus-specific C-type sortase catalyzes the head-to-tail assembly of the pilins into the final polymerized form, which in most instances has the thickness of a single subunit molecule (~2–3 nm) [7] and lacks a complex quaternary organization [8]. Surface piliation such as this is generally described as being “sortase-dependent”, the occurrence of which can be found only amongst the Gram-positive bacteria. Largely stemming from their prospective use as a vaccine candidate against disease-causing Gram-positive bacteria [9], the ensuing years have seen much effort to characterize the function and structure of sortase-dependent pili from various differently piliated pathogens. As a consequence, many molecular aspects about the pathogen-derived pilus types in Gram-positive bacteria have already emerged and continue to do so. However, the universal moniker as a trait for virulence no longer seemed to apply when, more recently, it had been revealed that sortase-dependent pili could assemble themselves on the cell surface of non-pathogenic bacteria. Here, it was visually shown that the gut-friendly commensals Lactobacillus rhamnosus [10,11,12,13], Lactobacillus ruminis [14], and Bifidobacterium bifidum [15] are also surface-piliated natively, at which point the sortase-dependent pilus began to carry the new name of a niche-adaptation factor.
This present review will concentrate on the newly found lactobacillar pilus types, whereupon an attempt is made to chronicle some of their molecular characteristics and properties that have been reported in the literature during the past few years. For this, an emphasis is placed on what new information has been learned concerning the biology of sortase-dependent pili, and as well on how these macromolecular surface features can contribute to gut colonization behavior.
2. Sortase-Dependent Pilus: An Overview
As mentioned above, the various sortase-dependent pili that originate from Gram-positive pathogens have already become well characterized in many respects, these including their genetic organization, assembly and structural composition, and as well molecular and cellular functionality. In this section, a description will be given on what is now understood as an overall consensus about these pathogen-derived pili after about a dozen years of intensive research and study. For more in-depth information on this specific topic the reader is referred to several recent reviews (e.g., [2,16,17,18,19]).
2.1. Gene Expression
As a characteristic genome feature of the piliated Gram-positive pathogens, the genes that encode the proteins for assembling a sortase-dependent pilus are clustered close together in a genetic island or operon [20]. Such a tandem arrangement of genes ensures the harmonized expression of the protein products that are involved with constructing a macromolecular pilus structure (Figure 1A). This genomic attribute is rather simply organized and includes the genes for the different pilin subunits, the one major type that comprises the polymerized length of the pilus backbone, and those deemed minor or ancillary that are positioned at the base and tip of the pilus. Also found within this gene cluster is the locus for a pilus-specific C-type sortase enzyme. However, some exceptions to this description also exist, e.g., where in the genes for the basal pilin-proteins are missing (e.g., [21,22]) and with additional sortase genes being present (e.g., [23,24]). Moreover, depending on the particular Gram-positive genus and species, the genome can encode a fimbrial operon for more than just one type of pilus. As an example of this, certain species of Streptococcus [23] and Corynebacterium [24] are known to contain the operon loci for expressing three structurally distinct pilus types.
For the most part, the basic genetic principles that underlie the regulation of sortase-dependent pilus expression are not yet fully deciphered and understood, although some studies are now beginning to reveal more about the mechanisms and factors involved with these controlling processes. While in many instances the cellular production of pili can be constitutive, one interesting phenomenon that has been noticed is the heterogeneity in pilus expression for a number of piliated Gram-positive pathogens, such as Enterococcus faecalis [25,26], Streptococcus pyogenes [27], Streptococcus pneumoniae [28], Streptococcus gallolyticus [22], Corynebacterium pilosum [29], and Corynebacterium renale [29], in which case not all of the cells are observed to display surface piliation. Additional studies indicate that for this bistable pattern of pilus expression there can also exist a mixed population of highly and lowly piliated cells [30,31,32]. Gene expression from the sortase-dependent pilus operons is controlled mainly at the transcriptional level, with the regulatory loci and elements being located in upstream regions and the regulatory proteins ordinarily acting as an activator [33]. For instance, the manifestation of bistability as found with S. pneumoniae PI-1 pili involves the RlrA positive regulator that not only activates the expression of the corresponding pilus operon [32], but as well, uses a positive feedback mechanism to regulate its own production [34]. Interestingly, an entirely different regulatory feature was described to account for the heterogeneous pilus expression of S. gallolyticus, in which the control of its pil1 fimbrial operon includes a phase variation mechanism that combines the length modification of an upstream leader peptide with ribosome-mediated transcriptional attenuation [35]. As a reason for the lack of homogeneity in the piliated cell population, it has been proposed to represent an adaptive survival strategy that allows bacterial cells to not only adhere and colonize the host environment via their pili, but, by being either non-piliated or less piliated, also able to evade those defenses of the host immune system that are directed against surface piliation [16,36].
2.2. Assembly and Anchoring
In a manner consistent with its namesake, the assembly and anchoring of the sortase-dependent pilus entails the covalent cross-linking action by two transpeptidase sortases: (1) the pilus-specific C-type for initiating and polymerizing together the various pilin-proteins and (2) the housekeeping A-type for attaching the fully assembled structure to the cell wall (for detailed review, see [37,38,39]) (Figure 1). As the site of pilus assembly is restricted to the cell surface, the various protein components are exported through the cell membrane via the Sec pathway [40]. This also necessitates that each of the sortases and the pilin subunits contain hydrophobic stretches to enable their membrane attachment, but also so that the bulk of each protein is within the cell wall region. For the two types of sortases, this occurs through transmembrane domains in their N-terminal regions [41,42]. On the other hand, for the different pilin subunits (tip, basal, and backbone), and similarly for various sortase-dependent surface proteins, they possess a characteristic C-terminal region that contains hydrophobic aliphatic amino acids and a tail end of positively charged lysine and/or arginine residues. In addition, extended out from this stretch of membrane-bound protein there are five amino acids (LPXTG) that serve as an important recognition and cleavage motif for both sortase enzymes [40]. Normally, when the sortases catalyze a break in this pentapeptide region, it occurs at the bond between the threonine and glycine residues. To accomplish this, each sortase enzyme has a large open cleft that contains an active site cysteine, which itself is part of a conserved catalytic motif (TLXTC) and during cleavage will form a thioacyl intermediate with the threonine of the LPXTG motif [43].
A prototypical model for describing sortase-dependent pilus assembly originates from a study of the surface piliation in Corynebacterium diphtheriae [24] (Figure 1B). To initiate the pilus assembly process, two C-type sortases, one coupled to a tip pilin and another carrying a backbone pilin, will first connect together their accompanying protein subunits [37]. Here, the lysine (K) of the so-called pilin motif (WXXXVXVYPKN) near the N-terminal of the backbone pilin and the LPXTG-motif threonine in the tip pilin become cross-linked via a covalent isopeptide bond, such that the pilin subunits are orientated head-to-tail. Following this, and done in the same manner, a succession of backbone pilins is added sequentially to the lengthening pilus structure. After a sufficient number of protein subunits have been incorporated, pilus biogenesis undergoes termination, which, depending on the pilus type, can be facilitated in one of two ways. For those pili with an archetypal three-subunit structure that also includes another ancillary pilin (in addition to the one at the pilus tip), pilus assembly proceeds to terminate via the presence of a housekeeping A-type sortase carrying the basal pilin, with C-type sortase-catalyzed K-T isopeptide bond formation then following and linking this particular subunit to the last backbone pilin of the pilus structure [44,45]. While now attached to the A-type sortase, a covalent bond forms between the basal pilin of the assembled pilus and the peptidoglycan layer, a process involving the lipid II moiety and leading to cell wall anchoring. For the pilus types not having the basal pilin, the housekeeping sortase simply catalyzes the same series of reactions as above, but instead with a backbone-pilin subunit, although less optimally [46]. Intriguingly, while the basal pilin is proposed as a likely candidate for signaling the end to pilus assembly [47], what then actually triggers this termination process in its absence is less clear and still remains to be established.
Much of the work to identify and confirm the relative positioning of the various subunit types within an assembled sortase-dependent pilus was done using EM in combination with immunogold labeling of pilin-proteins (i.e., immuno-EM) [24]. This technique served especially well, and derived from the findings representative structural models could be postulated, with each having in common the predicted location of the tip, basal, and backbone pilin subunits. However, as a peculiarity, the ancillary basal pilin was in some instances also detected along the length of the polymerized pilus backbone [24,48,49]. Because the primary structure of many basal subunits often tend to also exhibit what resembles a canonical pilin motif, a likely reason for its additional location in the pilus is that it was merely a random occurrence and involves being mistakenly recognized and incorporated by the C-type sortase. Since VYPK-like pilin motifs are seldom found in tip pilins, the critical lysine needed for K-T isopeptide bond formation would be missing, which then precludes any possible insertion of the tip subunit between adjacent backbone pilins. Alternatively, it was proposed that another amino acid motif of YXLXETXAPXGY (the so-called E-box) near the C-terminal region is involved in mediating the decorative outer attachment of basal pilins along the pilus backbone [50]. Although subsequent to this, it was reported that the E-box glutamate primarily plays a part in maintaining the proper fold of the pilin-proteins [51], and as well in the formation of intramolecular isopeptide bonds [52,53]. Yet another study using the more advanced cryo-EM technique appeared to refute the notion that the ancillary subunits are attached to the pilus in an atypical manner [8]. Here, it was established that for the S. pneumoniae TIGR4 pilus its structure consists of a repeating length of RrgB major pilins, with the RrgA and RrgC minor pilins confined to only the pilus tip and base, respectively. Significantly, this study also highlighted the limits of interpreting results obtained from conventional immuno-EM when undertaking to structurally characterize sortase-dependent pili, as there exists the tendency for antibody-related artifacts to sometimes occur [8].
2.3. Structural Composition
Apart from the tip, basal, and backbone pilins having a number of basic features in common, such as signaling regions at the N- and C-termini and certain consensus motifs, it is notable as well that their corresponding tertiary structures consist of protein domains with similar folds. Three-dimensional (3D) structures of these pilin subunits are comprised of the CnaA and CnaB domains (for detailed review, see [17,18]), both of which represent variant folds of the immunoglobulin (Ig)-like domain that was originally observed in the staphylococcal collagen adhesin (Cna) protein [54,55]. The structural fold of the CnaA and CnaB domains is dominated by a core of nine and seven β-sheets, respectively [56], which, depending on the pilin type, also contains characteristic regions of loops and α-helices, but as well, extra β-strands and other domains. Intriguingly, Gram-positive pilins are distinguished by their spontaneous formation of intramolecular (or internal) isopeptide bonds, with these occurring either between lysine and aspartate (K-D) or lysine and asparagine (K-N), and thus unlike the intermolecular K-T isopeptide bond that holds together different pilin subunits [52,53]. While perceived as a substitute for disulfide bridges [57], this intramolecular covalent bonding is formed autocatalytically, in which the spontaneity of the reactions involved is set forth when the CnaA and CnaB domains are folded together such that each creates a hydrophobic environment and wherein the key residues are positioned in optimal geometric proximity [7]. Moreover, here it seems that a particular residue environment is tailored to either K-D or K-N isopeptide bond formation [7].
To date, representative crystal structures of the tip, basal, and backbone pilin subunits have been solved from a variety of pilus types and pathogen sources, and with these results it is observed they have various structural features in common, but as well, those that distinguish them well-apart from one another (Figure 2). For instance, the tip pilin is by far the largest subunit in the sortase-dependent pilus, typically consisting of four separate subdomains, three of which having the CnaA/CnaB folds and that together resemble a stalk-like structure upon which is situated a larger fourth domain [58,59,60,61]. In contrast to the two Ig-like domains, this fourth domain is globular and contains recognized-binding subdomains, and it is these characteristics that make the tip pilins suitable for adhesive interactions. On the other hand, the basal pilins are the smallest in size of the pilus proteins and normally encompass one to three CnaB domains, which themselves may or may not display intramolecular isopeptide bond formation [62,63,64]. Of some interest, a universal structural attribute of the basal pilins is the presence of a C-terminal extension rich in hydrophobic proline residues. Because a similar tail-end region is not found in the tip and backbone pilins, its function in the basal subunit might be related to the events leading up to the covalent attachment of the pilus to the outer cell surface [63]. Typically for the backbone pilins, their structural makeup can be double- to quadruple-domained, and as well including both CnaA and CnaB folds [18]. In most cases, intramolecular isopeptide bonds are produced in these domains, which are assumed important in providing the structural rigidity and stability necessary for withstanding the severe shearing conditions encountered by sortase-dependent pili in natural environs [57]. A key feature of backbone (and basal) pilins is the so-called “linking lysine” that forms an isopeptidyl connection with the LPXTG-motif threonine of a nearby adjacent subunit. More often than not, this lysine is within the VYPK peptide region (pilin motif), but invariably this particular residue is found in the “head” N-terminal domain somewhere near the C-terminus [18]. Comparatively, the N-terminal domain of the backbone pilin has the most flexible structure, as its isopeptide bond tends to form slowly or then only during pilus assembly [65]. It is for this reason that when backbone pilins are produced recombinantly, the N-terminal domain will readily succumb to proteases. Here, it is thought that the structurally supple nature of the N-terminal domain will make the lysine residue more accessible for docking to an adjoining pilin via the C-terminal threonine, but as well, then allow for sealing and merging at the head-to-tail interface of two backbone subunits as they become polymerized into a pilus fiber [66].
2.4. Functional Attributes
From a physical perspective, piliated Gram-positive pathogens are considered more competitive than their non-piliated counterparts by being able to adhere to available target sites from a distance. This capacity for first contact with the host is due to the exceptional length of pilus structures, but as well, to the exploitation of their characteristic adhesiveness. For instance, numerous studies have shown that the virulence of piliated Gram-positive pathogens can stem from pilus-mediated adhesion, and it has become widely recognized that the tip pilins are the main adhesive component of pili. Primary structure examination of tip pilins has revealed that several contain sequence for a von Willebrand adhesin (VWA) domain, e.g., such as in C. diphtheriae SpaC [20], E. faecalis EbpA [67], S. pneumoniae RrgA [58], S. agalactiae PilA [68], and S. pyogenes AP-1 [69]. The VWA domain can be found in many other bacterial proteins, but as well, various archaeal and eukaryotic proteins, and as a functional property it is known to interact with the epithelial ECM [70]. Accordingly, an adherence to ECM substrates, such as collagen, fibronectin, fibrinogen, and laminin, has been attributed to various Gram-positive pathogen-sourced pili and tip subunits [22,71,72,73]. Moreover, for some piliated pathogens (e.g., C. diphtheriae, S. pneumoniae, and S. pyogenes [74,75,76]) their adhesion to epithelial cells is also pilus mediated. As host cell invasiveness by most pathogens first requires an attachment to the epithelium, and in particular to any exposed ECM proteins, those genera and species with such substrate-adhesive pili are functionally specialized for this purpose and have a distinct advantage for successful colonization and dissemination.
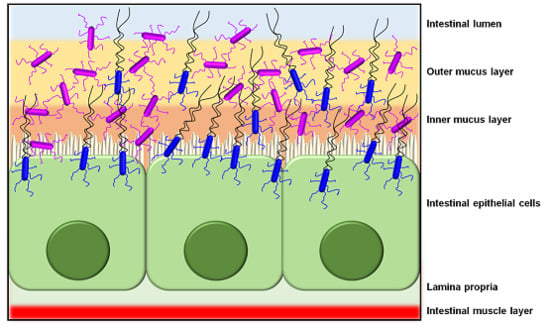
Then again, while most functional analyses of pathogen-derived sortase-dependent pili point to ECM components as adhesion targets, a recent study had reported that intestinal mucus can also act as a binding substrate [77,78]. Here, the Pil3 pilus of the opportunistic pathogen S. gallolyticus was shown to mediate adherence to colonic mucus and was required for the colonization of the murine distal colon, all primarily through the action of its tip pilin (Pil3A). This offers the possibility that the pili of Gram-positive pathogens can as well promote bacterial colonization of the gut mucosa. In addition, sortase-dependent pili have an important role in the development of pathogenic biofilm communities, with a good example being those oral species of Streptococcus and Actinomyces involved with dental plaques and caries [79,80,81]. For these, the adhesive pilus contributes to the early stages of biofilm growth by participating in the primary attachment of bacterial cells to saliva-coated tooth surfaces. Further, as various types of infections are caused by enterococcal and streptococcal pathogens, their pilus-driven accumulation as biofilm (e.g., [22,26,68,69,75]) can often lead up to hindering or delaying wound healing [82]. As a final point, other evidence has begun to emerge that suggests the sortase-dependent pili of pathogenic Gram-positives behave much like many other surface proteins and are targeted by the host innate immune system, e.g., as demonstrated by their ability to elicit certain inflammatory [28,83] and phagocytic [84,85,86] responses.
3. Sortase-Dependent Pili in Non-Pathogenic Lactobacilli
3.1. History of Discovery
After the discoveries that some pathogenic Gram-positive bacteria can be piliated on their outer surface, it was not until 2009 when sortase-dependent pili were also found present in the less harmful commensals. As it happens, there were actually two publications that year, both giving an account of surface piliation in Lactobacillus rhamnosus GG, a gut-adapted strain whose commercial reputation is that of a much advocated and used probiotic (for detailed review, see [87]). In June, a Belgian group published a study [13], in which, by using EM and negative staining, they observed emanating from the cell poles of an extracellular polysaccharide-deficient mutant of L. rhamnosus GG the presence of what was discerned to be pilus-like formations. Later on in October, another report was published by a multinational consortium of scientists that then confirmed these surface structures on L. rhamnosus GG cells as being sortase-dependent pili [11]. Findings for this were from a study that was largely a genomic comparison between the GG and LC705 strains of L. rhamnosus. Here, it was found that the L. rhamnosus GG genome contains loci for two different sortase-dependent fimbrial operons (so-called spaCBA and spaFED), each of which encoding for the predicted tip (SpaC and SpaF), basal (SpaB and SpaE), and backbone (SpaA and SpaD) pilin-proteins, and as well a C-type sortase enzyme (SrtC1 and SrtC2). On the other hand, the genome of the L. rhamnosus LC705 dairy starter strain was observed to contain the genes for only the spaFED pilus operon. Moreover, an examination of the deduced primary structure from the corresponding pilus operon genes had confirmed that each of the SpaCBA and SpaFED pilins contains the characteristic conserved sequence motifs and domains of a typical Gram-positive pilin subunit (see Table 1). Incidentally, the “Spa” prefix given to the names of the L. rhamnosus pili was adapted from the nomenclature used for designating each of the pilus types in C. diphtheriae and this corresponds to sortase-mediated pilus assembly [88].
Typically, the tried-and-tested means for determining the surface localization of sortase-dependent pili in Gram-positive bacteria has included the electrophoretic immunoblotting technique and the previously mentioned use of immuno-EM, with both necessitating the need of anti-pilin serum [89]. For the immunoblotting analysis of cells, an observed ladder-like pattern with compressed high-molecular-weight (HMW) protein bands corresponds to the various fragmented lengths of accumulated pilus fibers, and this is normally taken to indicate the presence and production of sortase-dependent pili. In the Kankainen et al. (2009) study, the above methods with SpaC pilin antiserum were used to establish the active expression of the L. rhamnosus GG spaCBA-encoded loci, and ultimately the assemblage of the produced pilin-proteins into what is now normally known as the SpaCBA pilus [11]. At this point, the results from the immuno-EM experiments had indicated the existence of the SpaC pilin subunit along the entire length of the SpaCBA pilus structure [11], and with its apparent predicted location at the pilus tip only confirmed afterward in a follow-up study some three years later [90].
Significantly though, a number of “firsts” were established for sortase-dependent piliation from the study by Kankainen and colleagues. These included the first descriptions of a piliated Gram-positive non-pathogen, a piliated Lactobacillus species, and functionally of a sortase-dependent pilus having an intestinal mucus-binding capacity [11]. For this latter finding, adherence to human intestinal mucus was revealed for the SpaC subunit itself and then as well in the context of the SpaCBA pilus, which was shown by blocking its binding ability through the use of anti-SpaC serum and a spaC-inactivated mutant of L. rhamnosus GG [11]. It is on such a basis that the authors of this study had concluded the mucoadhesive SpaC pilus protein is an important host colonization determinant in L. rhamnosus GG and likely a main contributing factor in the more pronounced persistence of this gut transient (allochthonous) strain. At the time, the unearthing of mucus-binding surface piliation in L. rhamnosus GG had represented an entirely new concept, and as a novel molecular mechanism it ushered in a new impetus to examine and understand what physiological and functional roles might be played in intestinal microecology and probiosis. Curiously as well, not only did these research findings draw some attention and interest within the related scientific community [91], but there also was an element of sensational newspaper and social media coverage aimed at the general public.
3.2. Other Lactobacillar Pilus Operons
Following close on the heels of the above-mentioned findings came the expected further scrutiny of other Lactobacillus species, and from this it appeared that the genetic potential for sortase-dependent surface piliation is not too prevalent among other members of this genus. For instance, only the phylogenetically close “casei group” of lactobacilli (Lactobacillus casei, Lactobacillus paracasei, and L. rhamnosus) are containing the representative fimbrial spaCBA and spaFED operons [92,93,94], and where expectedly a reasonably high degree of sequence identity exists among the primary structures of the various predicted proteins at a species and strain level. Added to this, genomic evidence was brought forth that the sortase-dependent pilus genes are present in Lactobacillus ruminis, a gut-indigenous flagellated species taxonomically distant from the members of the casei group [95]. Here, it was found that the fimbrial operon (eventually named as lrpCBA; [14]) encodes a canonical set of three pilus proteins and single sortase that are distinguishable from those associated with the SpaCBA and SpaFED pili, thus appearing to represent another type of lactobacillar pilus. It was later confirmed in a published report that L. ruminis is a natively surface-piliated bacterium [14]. Further, based on a study that carried out in silico sequence predictions to estimate the genomic presence of sortase-dependent pilus operons, it was inferred from the data output that other potentially piliated lactobacilli are distributed in clades containing the Lactobacillus composti, Lactobacillus brevis, and Lactobacillus parabrevis species [96]. However, as of yet, any number of these pilus-producing lactobacilli still remains to be corroborated experimentally in further studies.
4. SpaCBA Piliation
Thus far among the three types of sortase-dependent pili in lactobacilli, the SpaCBA pilus from L. rhamnosus GG has become the best studied, which in large part is due to it being the first one discovered and made known to be produced by cells, but as well, stemming from a strong commercial interest in the science behind the advocated probiosis of its host strain. Up to now, any other published reports indicating the active expression of the spaCBA operon are limited to a just a few L. rhamnosus strains [10,12], but otherwise nearly all molecular and functional characterization studies have involved the L. rhamnosus GG SpaCBA pilus.
4.1. Genetics
As introduced beforehand, the genomic distribution of the spaCBA operon (i.e., spaC-spaB-spaA-srtC1) appears to be restricted to only the L. casei, L. paracasei, and L. rhamnosus species of the so-called casei group [12,92,93,94]. However, while so far the spaCBA loci are a near-common attribute of the L. casei and L. paracasei genomes, surface localization of the SpaCBA pilus is still to be established in these two species. On the other hand, the presence of the fimbrial spaCBA operon in L. rhamnosus seems to be proportionally more widespread in gut-sourced strains than those originating from dairy products [10,12]. This suggests that whereas a strong evolutionary pressure had prevailed to maintain mucoadhesive SpaCBA pili in the L. rhamnosus strains whose ecological niche is the mucosal gut, it was not sufficiently manifested for those strains isolated from a milk-based environment, since presumably an adaptive advantage would not have been similarly gained by this molecular trait [10,12]. Still, although mucus itself might be perceived as a unique selection determinant for acquiring the SpaCBA pilus-related loci, it is not a very dominant driving force, as there are also intestinal strains without the spaCBA operon [10,12]. Moreover, the spaCBA genes seem not to be overly prevalent in any L. rhamnosus strains that originate from other mucus-lined regions in the body, such as the vaginal cavity and the respiratory airways, including the mouth [10,12]. Further, because a pan-genomic appraisal of L. rhamnosus found that the spaCBA operon is not part of the essential core genome, and instead is included among the dispensable accessory loci, the SpaCBA pilus genes can be regarded as a genomic rarity in this species [12]. Thus, as it is clear the spaCBA operon is a genetic element not shared by all L. rhamnosus strains, this might suggest that the corresponding genes had evolved recently in the genome, potentially through the lateral transfer of sortase-dependent pilus loci from the closely related L. casei and L. paracasei species, or perhaps even more ancestrally from piliated gut bacteria like Enterococcus faecium and Enterococcus faecalis [12]. While purely speculative, the premise for such a lateral gene acquisition is biologically conceivable, given that a transposon-like insertion sequence (IS) is identifiable within a region at both flanking ends of the L. rhamnosus spaCBA operon [97], and as this type of genomic feature is known for helping expedite the gene movement process.
To date, much of what is known regarding the regulation of SpaCBA pilus production is based on a comparative in silico analysis of DNA segments bordering the spaCBA operon in the genome of the L. rhamnosus GG strain. In the study by Douillard and colleagues, a possible regulatory region that might serve to control constitutive expression of the spaCBA loci was found within a specific DNA sequence immediately upstream from the spaC gene, and which may have arisen in the genome as an iso-IS30 element [97]. Here, putative -10 and -35 promoter DNA elements were identified and include the pentameric 5′-TTGAA-3′ and hexameric 5′-TGGTCT-3′ sequences, respectively. Interestingly, in spite of their divergence from canonical consensus promoter sequences for RNA polymerase binding (i.e., -10 TATAAT and -35 TTGACA), the involvement of an alternative sigma (σ) factor was not being promoted as a possibility. However, in another study [98], it was found that the spaCBA operon from the L. rhamnosus E800 strain, which also produces SpaCBA pili [12], might instead be regulated by a promoter region that more closely resembles a typical -10 and -35 consensus sequence (i.e., TATAAT and TTGTTA, respectively), and thus better suited to being recognized by a standard σ70 factor. Moreover, similarly matching -10 and -35 recognition sites were also detected in the genome of L. rhamnosus GG, and as well for the LMS2-1 strain, though whose SpaCBA pilus production is not yet proven. Expectedly, these promoter sequences were not at all obvious for the spaCBA genes of the non-SpaCBA-piliated L. casei BL23 strain.
Owing to the juxtaposed presence of IS elements at both ends of the L. rhamnosus GG spaCBA operon, and therein the chance for increased genomic instability, Douillard and colleagues had also conducted an intriguing investigation to ascertain what impact evolutionary forces have on the permanency of the spaCBA loci in the genome [99]. In an examination of L. rhamnosus GG cells that had been propagated for at least 1000 generations under normal or highly stressed environmental conditions, the authors found that for the former there was no related gene loss from the genome, but for the latter when bile salt was used as the stressing agent, the IS elements became activated and appeared to cause some genetic recombination involving the spaCBA operon. In the context of the SpaCBA pilus as an evolving molecular trait, these findings clearly revealed the genomic plasticity of the L. rhamnosus GG strain and its adaptive prowess for change when confronted by a different environment.
4.2. Structure
In terms of overall subunit architecture, lactobacillar SpaCBA piliation adopts the typical structure and molecular arrangement of a sortase-dependent pilus (Figure 3), wherein each of the three types of pilin-proteins is found situated in its predicted location. For instance, immuno-EM evidence from the Reunanen et al. (2012) study indicates that for the SpaCBA pilus, a repeating number of major SpaA pilins (~30 kDa) are polymerized linearly into the backbone structure, with the ancillary SpaB (~20 kDa) and SpaC (~90 kDa) subunits located at the anchored base and outwardly tip sites, respectively [90]. However, because a conventional EM technique was used to pinpoint the positioning of the individual pilin subunits, the experiments also tended to show the sporadic presence of the SpaB and SpaC pilins along the pilus structure. Notwithstanding the possibility that the added location for the ancillary pilins may well represent an artifact due to the use of polyclonal antiserum [8], this other site was nonetheless included into the schematic model being proposed for the SpaCBA pilus [90]. Here, the basal SpaB subunit is sandwiched between two SpaA pilins and as well appears on the pilus exterior, whereas the SpaC tip pilin only has the outer decorative location along the pilus backbone. However, as the mucoadhesive SpaC subunit is considered a key molecular determinant for gut colonization by L. rhamnosus GG, its extra presence would give SpaCBA pili a distinctively heightened ability to extend binding to the intestinal mucosa. Yet, in the years following this particular investigation there has been no further data or research to substantiate the presumptive structural uniqueness of the lactobacillar SpaCBA pilus, and in hindsight its suggested architectural divergence from an accepted norm of other sortase-dependent pili [8] might have been based on circumstantial observations and at the time simply a practical interpretation. Thus, in retrospect, it is quite plausible that in the case of SpaC, since the stalk-like domain regions of the tip pilin-protein structure could bear a topological resemblance to the backbone subunit, a certain portion of antibodies in the polyclonal serum raised against SpaC might as well have an affinity for SpaA pilins when they coalesce to form the structural backbone of the pilus. Alternatively, and more significantly, the recently solved crystal structure of L. rhamnosus GG SpaA did not reveal recognizable structural topologies for a possible mechanism that would lead to a peripheral attachment of SpaC along the pilus backbone [100]. Even so, in the context of competing for the binding sites available throughout the intestinal mucosa, any L. rhamnosus GG cells having SpaCBA pili with just a single SpaC adhesin per fiber would presumably retain the capacity to outlast other strains not piliated in a similar way.
As it stands, there have been no characterization studies examining the molecular aspects of how the various SpaCBA pilins are polymerized together into a pilus structure, although it is largely assumed that this will conform to the pilin assembly process established for other types of sortase-dependent pili. However, there is one related report in which the authors had tried to uncover what controlling role sortase specificity has on the assembly and anchoring of the L. rhamnosus GG SpaCBA pilus [101]. Previously, with another piliated Gram-positive host [102], it was revealed that a structural difference existing between the pilus-specific C-type and housekeeping A-type sortases is responsible for regulating their respective activity and specificity on the LPXTG peptide of different pilin subunits. For this, the C-type sortase has an extra N-terminal region that folds into a flexible helical loop (a so-called lid), wherein certain conserved residues mimic themselves as a pseudo-substrate that can interact with and cover the active site. While it was found that eliminating the lid from the sortase does not impede catalysis, the shifting or movement of the lid from the open cleft that contains the active site is viewed necessary for making the LPXTG-substrate accessible to the catalytic pocket. Interestingly, the authors of this particular study went on to suggest that the pilin-specificity of C-type sortases is not just dependent on recognizing the limited stretch of residues in the LPXTG motif, and instead most probably involves other molecular determinants [102]. Much along these lines, the study by Douillard et al. (2014) had found that by extending the amino acid sequence of the LPXTG motif at the C-terminus, this would allow differentiation between those substrates being recognized by a C-type or A-type sortase [101]. Presumably for the specificity of the SrtC1 and SrtC2 C-type sortases, among the backbone (SpaA and SpaD) and tip (SpaC and SpaF) pilins of L. rhamnosus GG, there occurs a conserved Gly-Xaa-Gly pattern proximately downstream of the LPXTG residues that is not present in the L. rhamnosus GG basal pilins (SpaB and SpaE) and various other sortase-dependent surface proteins. Having been designated the “triple glycine motif”, it was revealed their residue-substitution in the SpaA and SpaC pilins effects the normal specificity of the SrtC1 sortase, with a perturbed SpaCBA pilus polymerization as the observed outcome. With the added presence of non-polar glycines, the rationale is that the backbone of this lengthier peptide region (LPXTGGXG) would then have more conformational freedom and increased flexibility, and thus better catering to the structural nuances of C-type sortase specificity and binding affinity. Further, as a conserved version of this new motif was also identified in the backbone and tip pilins of other piliated Gram-positive genera and species [101], it might well be considered a universally shared recognition site of pilus-specific C-type sortases. Alternatively proposed for the housekeeping A-type sortase, whose structure lacks the lid feature, its specificity for LPXTG-like proteins does not require the existence of a triple glycine motif [101].
So far, among the three different SpaCBA pilins, the backbone SpaA subunit from L. rhamnosus GG has been crystallized [103] and its tertiary structure solved by X-ray crystallography [100]. As the first crystal structure of a pilin-protein derived from a non-pathogenic source, SpaA was shown to be structurally representative of other Gram-positive pilins, e.g., by both being multi-domained and adopting Ig-like folds. More specifically, the SpaA structure is made up of two CnaB subdomains, each identified with an internal isopeptide bond and an E-box motif [100]. However, the chemistry of the isopeptide bonds is of particular interest, since whereas this covalent bond in the N-terminal domain is typical and formed between lysine and asparagine residues (K-N), in the C-terminal domain region, the bond formation is uncharacteristic by involving an aspartate instead (K-D) [100]. It is worth mentioning that from the structural work on SpaA, this marked the first time that the crystal structure for the N-terminal domain of a Gram-positive pilin was determined, since generating good X-ray-diffracting crystals has normally been quite difficult due to the intrinsic flexibility of this protein region. Concerning the functional role of the E-box, despite showing that residue-substitution has a negative impact on the proteolytic and thermal stability of SpaA pilin-protein, polymerization of the SpaCBA pilus was unaffected and remained normal [100]. Interestingly, because in the study by Chaurasia and colleagues a structural likeness to a native-like pilus was found in the molecular packing of SpaA crystals, this led the authors to propose a macromolecular model of the SpaCBA pilus that shows the way in which the SpaA pilins are arranged together. Here it was observed that a head-to-tail orientation of SpaA proteins places the C-terminal LPXTG peptide region of each subunit in the vicinity of an open cavity next to the N-terminal pilin motif residues of the adjoining subunit. With the added contribution of an inter-domain hinge angle of 152° in each SpaA pilin, the polymerized repeats of SpaA subunits can be modeled as having an extended spiraled staircase shape, which should give both rigidity and flexibility to the pilus backbone structure, and thereby explaining how the SpaCBA pili are designed to withstand the harshness of the highly dynamic intestinal environment [100].
4.3. Adhesion
As previously mentioned [11], the SpaCBA pilus is inherently mucoadhesive, with the large-sized SpaC pilin being viewed as the main determinant for such binding behavior. This was further supported by the results of mucus-binding experiments performed with a recombinant SpaCBA-piliated lactococcal clone that had the SpaC subunit deleted from the pilus structure [104]. Interestingly, as also found with some pathogen-derived tip pilins, the primary structure of SpaC shows the presence of a VWA domain, wherein additionally there is included a short region that is partially homologous to the domain for a fucose-binding lectin [11]. Based on this, it is speculated that substrate binding by SpaC might be attributed to a lectin-type recognition mechanism, and then particularly since one of the prime constituents of mucus is glycosylated mucin protein [11]. This type of SpaC-glycan interaction is also supported by a recent study that indicates SpaC pilin can bind to the carbohydrate chains of mucin, with an essential role being played by the β-galactoside moiety at the non-reducing end [105].
In addition to having an ability to bind mucus, SpaCBA pili appear to mediate the adherence of L. rhamnosus GG to epithelial cells. For this, two different studies using mutated strains of L. rhamnosus GG, a spaCBA knockout mutant [106] and an isogenic spaC mutant [107], were able to show that the SpaCBA pilus, and explicitly the SpaC pilin [107], have a functional role in bacterial adhesion to the human intestinal Caco-2 cell line. Moreover, while a dedicated domain for binding ECM collagen cannot be detected within the SpaC primary structure, there is one report that indicates a binding interaction between SpaC and collagen can occur [108]. Here, it was found that the SpaC-collagen interaction is also subject to rapid dissociation and from this it had been speculated that this could allow for a quick detachment and attachment between the SpaCBA pilus and the host environment, such as in those disrupted regions of the gut epithelium where ECM proteins are exposed and available for binding.
While the 3D structure of SpaC is yet to be solved, its crystallization as a near full-length protein has been recently reported [109], and arising from this work were some interesting observations about the molecular determinants for substrate binding by this pilin subunit. For instance, conserved domain prediction via InterPro had revealed the VWA domain is located within the N-terminal region of the SpaC protein, whereas the domain for the CnaB-type fold is localized at the C-terminal end. Moreover, it was further revealed that SpaC contains the motif for a metal ion-dependent adhesion site (MIDAS; D-x-S-x-S T D), and for which many VWA domains use to bind metal ions [70]. Further, based on predictive homology comparison with the RrgA and GBS104 tip pilins, the MIDAS-containing VWA domain can be found at the tip and surrounded by two inserted arms, and functionally it has been implicated with collagen binding.
In a trio of published studies [108,110,111], atomic force microscopy (AFM) was used to examine and characterize in detail the subunit organization and binding mechanics of SpaCBA pili, and here some notable findings were revealed. For instance, in the first study it was observed that the SpaCBA pili can appear bundled together as a single structural unit, which was thought to originate from glycan-lectin or hydrophobic interactions involving the SpaC subunit [110]. This was further expanded upon by the second study where it was found that SpaCBA pili have a propensity for self-adhesion that is driven by homophilic interactions between SpaC pilins, and which potentially also contributes to bacterial cell aggregation [108]. Rather interestingly, when L. rhamnosus GG cells were experimentally subjected to being pulled apart, and depending on the amount of force used, the observed mechanical retort of the pili could be categorized in two possible ways: (1) as a zipper-like mechanism involving SpaC-SpaC binding at low applied force and (2) as a molecular nanospring at higher forces, in which pili have a springiness and thus able to withstand a sustained stress or shock. Finally, a third study went further to reveal that the mechanical forces behind SpaCBA pilus-mediated adhesion can vary with the type of target-binding substrate [111]. Here, the coiling and extension behavior (nanospring) of SpaCBA pili are what help mediate adhesion to solid surfaces, as was observed with hydrophobic or glycan (i.e., mucin) substrates. Oddly, it was found that the binding of L. rhamnosus GG cells to an intestinal epithelial cell line (Caco-2) was less dependent on the nanospring properties of the SpaCBA pilus, but instead had relied on a surface tethering between the corresponding cellular membranes (i.e., membranous nanotethers). Although laboratory-cultured Caco-2 cells lack mucus on their outer surface, it was hypothesized that the mechanical forces promoting L. rhamnosus GG adhesion to the gut mucosal epithelium will likely involve both the SpaCBA pilus nanosprings and the membranous nanotethers [111].
Interestingly from another study [112], it emerged that the recombinant form of the basal SpaB pilin-protein has adhesiveness for intestinal mucus, and for which the binding is about seven-fold more pronounced than that by SpaC. The peculiarity of this finding is that the SpaB primary structure shares no homology to any of the known (or otherwise) mucus-binding domains, and thus the ability of this basal pilin to adhere strongly to mucus substrates was unpredicted and rather surprising. By comparison, the backbone SpaA subunit also lacks any homology matches to recognizable mucus adhesion domains, but more predictably it showed no such affinity for mucus [112]. However, in the case of SpaB, its mucoadhesiveness seems not to rely on certain residues of a dedicated binding domain, but rather it might be explainable through a less specific type of adhesion mechanism. Stemming from the observation that SpaB has a higher isoelectric point than the SpaC and SpaA pilins (pI 8 vs. ~5), evidence was presented to show that the positively charged SpaB pilin might use electrostatic contacts to interact with negatively charged mucus glycans [112]. Still, while such a mucosal interaction was revealed for the recombinant SpaB protein, any such contribution to the affinity of the SpaCBA pilus for adherence to intestinal mucus could not be established, either through the use of SpaB-specific antiserum on L. rhamnosus GG cells [112], or then by using a recombinant lactococcal clone that produces SpaB-deleted SpaCBA pili [104]. Consequently, in terms of actual function, the purpose for which the basal SpaB subunit is strongly mucoadhesive remains rather puzzling and intriguing.
As a host colonization strategy, the overall adhesive properties offered by SpaCBA piliation likely enables L. rhamnosus GG cells to extend their transient occupation of the intestinal niche (Figure 4), or at least more so than those strains without such pili [11]. Presumably, not only would the SpaCBA pilus facilitate the initial cell attachment to host binding sites, but once fastened, it also acts like the sortase-dependent pili from certain other Gram-positive bacteria by helping L. rhamnosus GG undergo self-aggregation to form biofilm, which could further enhance its colonization of the gut. For instance, from the analysis of knockout phenotypes for the spaCBA operon [106], and as well from the behavior of recombinant SpaCBA-piliated lactococci [14], SpaCBA piliation was shown to contribute to the growth of L. rhamnosus GG biofilm colonies, which appears to include a strong reliance on the adhesive SpaC tip pilin as cells aggregate together [14]. In this particular situation, the SpaCBA pilus is likely to be an adaptive advantage to L. rhamnosus GG and by its characteristic adhesiveness will help make this strain a less stringent allochthonous gut inhabitant [112].
The adhesive role of the SpaCBA pilus as a colonization factor is further demonstrated by its capacity to compete for binding sites in the gut mucosal epithelium. For instance, a recent study by Tytgat and colleagues had examined whether SpaCBA-piliated L. rhamnosus GG cells can supplant the binding of a vancomycin-resistant enterococcal species to mucus [113]. Here, the Gram-positive E. faecium E1165 strain is known to display sortase-dependent pili, which also happen to be quite homologous to the SpaCBA pili, reaching upwards to at least 50% sequence identity. By using polyclonal antibodies with specificity toward L. rhamnosus GG cells and the SpaC pilin, the authors were able to show that these immune sera can obstruct the mucus-binding ability of E. faecium, which presumably is due to its own surface pili. This binding interference was further linked to L. rhamnosus GG SpaCBA piliation by the observation that purified SpaC protein can also prevent E. faecium cells from adhering to mucus. As inferred from these findings, the authors suggested that the apparent outcompeting effect exerted by the SpaCBA pilus exemplifies some of the mechanistic rationale behind the advocated probiosis of the L. rhamnosus GG strain.
4.4. Immunogenicity
Although the published reports describing L. rhamnosus GG as a piliated strain were on hand during 2009 [11,13], it was only until three years later when a study came out that provided some first insights about the immunomodulating properties of the SpaCBA pilus [106]. Here, Lebeer and colleagues examined the phenotypic variances of different L. rhamnosus GG knockout mutants, including of the spaCBA operon, and found that the SpaCBA pilus is able to dampen the lipoteichoic acid (LTA)-induced mRNA levels of the inflammatory cytokine interleukin (IL)-8, for which it was suggested the related interactive mechanism might also involve the activation of the innate Toll-like receptor-2 (TLR2). Further to this, the authors of another study had concluded that, on the basis of a comparison between L. rhamnosus GG and two non-piliated strains of L. casei, the SpaCBA pilus should likely participate in TLR2-dependent NF-κB signaling [97]. Lending further support to the above findings, the added evidence from the study by von Ossowski and colleagues had suggested the SpaCBA pilus plays, in all likelihood, some sort of contributory role as a TLR2 agonist or activator [104]. Here, it was established that recombinant SpaCBA-piliated lactococci could activate NF-κB responses in HEK-TLR2 cells, which in itself was taken as an indication of stimulated TLR2-dependent activity. However, by using mutated lactococcal constructs, in which the SpaB and SpaC subunits had been removed from recombinantly expressed SpaCBA pili, it was shown that neither of these two ancillary pilins is specifically recognized by TLR2 [104]. Instead, it was reasoned that TLR2 recognition of SpaCBA pili might have more to do with the topology and overall protein fold of the polymerized pilin subunits [104]. This line of logic was consistent with the findings of another study, wherein the RrgA tip-pilin of the S. pneumoniae type 1 pilus could only activate TLR2-dependent responses while as an aggregated macromolecular form, and not in its monomeric state [83]. Yet, beyond its activation of TLR2-related signaling, the von Ossowski et al. (2013) study had revealed that the SpaCBA pilus is also involved with modulating the dendritic cell (DC) production of various inflammatory cytokines, such as IL-6, IL-10, and IL-12, and as well tumor necrosis factor-alpha (TNF-α). Noticeably, though, what emerged from this work was to be a recurrent theme involving the adhesive and surface-localized nature of SpaCBA piliation. Because innate immune signals have a short half-life, they require being released nearby to function effectively. For this, the SpaCBA pilus and, in particular, its SpaC tip-pilin adhesin would seem to provide the needed cell-to-cell contact, such that the closer proximity between immune-related cells and bacteria allows the pili (or perhaps some other types of surface features) to then promote the release of, e.g., TLR2-dependent responses or DC inflammatory cytokines [104]. Additionally (and going out on a speculative limb), it was suggested that the SpaCBA pilus might represent a novel microbe-associated molecular pattern (MAMP)-like modulator of innate immunity, and thus, in the case of L. rhamnosus GG, it would be one of several other cell surface structures that have been branded as immunomodulatory [104].
Further highlighting the immuno-adhesiveness of the SpaCBA pilus were the results from several studies examining other immunogenic responses and signaling pathways from a mix of immune-related cells. For instance, Ardita and colleagues showed through the use of a spaC-negative isogenic L. rhamnosus GG mutant that the SpaC subunit has a discernible role in stimulating certain types of cellular responses from human intestinal epithelial cell lines and the murine small intestine [107]. Here in this study, these included stimulating reactive oxygen species (ROS) production, pathway activation involving extracellular signal-regulated kinase (ERK) and mitogen-activated protein kinase (MAPK), and protection from intestinal damage induced by radiation. In the work by Ganguli et al. (2015), the focus was on human fetal gut epithelial cells, and where purified recombinant L. rhamnosus GG SpaC pilin was shown to cause a reduction in IL-6 levels when exposed to IL-1β, but as well, a modulation of immune gene expression by down-regulating TLR3, TLR4, and TIRAP (a TLR-associated intracellular adapter molecule) mRNA levels, yet up-regulating the levels for IL-1RA (IL-1 receptor antagonist) mRNA [114]. On the other hand, it was seen from the findings reported by Vargas García and colleagues that the SpaCBA pilus was also involved with modulating anti-inflammatory cytokine mRNA levels in macrophages, in this case by up-regulating IL-10 and down-regulating IL-6 in the RAW 264.7 murine cell line [115]. However, SpaCBA piliation was itself found not directly responsible for these immunomodulatory effects, but more likely the means used for maintaining contact between macrophage cells and other L. rhamnosus GG cell-surface attributes. Interestingly, an entirely different premise for the immunomodulating behavior of the SpaCBA pilus was proposed in the study by Tytgat and colleagues [116]. Here, the authors had identified the presence of mannose and fucose moieties on L. rhamnosus GG SpaCBA pili, presumably in the form of glycosylation, which were shown to be recognized by primary DCs through an interaction with DC-SIGN (Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin), a C-type lectin receptor found on the surface of these cells and macrophages. Significantly, in addition to functioning as an adhesin, DC-SIGN is also known to initiate innate immune responses via TLR-dependent mechanisms. Related to this, Tytgat and colleagues had shown the elevated gene expression for certain DC cytokines (IL-6, IL-10, IL-12A, and IL-12B) that was roused by the mannosylated/fucosylated SpaCBA pili could be partly reduced by the blocking action of DC-SIGN antibody [116].
However, in spite of its immuno-characteristics, the results reported on the L. rhamnosus GG SpaCBA pilus should be interpreted in a broader context. For instance, the magnitude of innate responses from the host gut immune system toward commensals (and probiotics) is much more subdued than to any opportunistic pathogens, and then what can be best inferred as only basal-level immune activities between various MAMPs present on the outer bacterial surface and the pattern recognition receptors (e.g., TLRs) of host-immune cells [117,118,119]. Despite this, over the past few years considerable research effort has been geared at understanding the immunogenic basis for the advocated health benefits of certain gut-residing (and transient) commensals, for which much of the focus is on a variety of outer surface features, both proteinaceous and carbohydrous. Regarding the SpaCBA pilus, its own recognition by the host innate immune defenses is to be expected, but while the various response strengths are more or less at low stimulatory levels, this would be consistent with the borderline immuno-intrusiveness of L. rhamnosus GG as a harmless gut-adapted commensal. Still, as an envisaged benefit to the host, one can theorize that this commensal-derived pilus is possibly correlated with sensitizing the intestinal epithelium to an immune-alerted readiness state, which itself could conceivably be one of the protective mechanisms that specifically thwarts gut-invading piliated pathogens.
5. SpaFED Piliation
The first published account of the spaFED operon, which encodes for the second type of lactobacillar pilus called SpaFED, comes from the study by Kankainen and colleagues, wherein the related pilin (spaF, spaE, and spaD) and sortase (srtC2) genes are found in the genomes of the L. rhamnosus GG and LC705 strains [11]. Surprisingly, since this original finding, along with the observed incidence of the fimbrial spaFED operon in additional L. rhamnosus strains [10,12] and in two other Lactobacillus species [92,93,94], there have still been no reports of a natively produced SpaFED pilus structure. Thus, by comparison to the published studies on SpaCBA piliation, the extent of characterization available for the SpaFED pilus is less comprehensive, with most findings about its functional and structural properties relating to a recombinantly expressed form in Lactococcus lactis [98]. However, just as a caveat, beyond the phenotypic revelations about the SpaFED pilus, any applicability in vivo in the original host species or strain is quite notional and should be viewed with some circumspect.
5.1. Genetics
While the fimbrial spaFED operon (i.e., spaF-spaE-spaD-srtC2) can only be detected in the genomes of the three Lactobacillus species from the casei group, i.e., L. casei, L. paracasei, and L. rhamnosus [92,93,94], it appears to be a nearly common feature within all the related strains. Moreover, shared among these species and strains is an absence of experimental evidence for any active gene expression from the spaFED operon, which in effect makes the SpaFED pilus a conjectural structure with no genuine role to play in these lactobacilli. Thus, in contrast to those spaCBA operons in a few L. rhamnosus strains that can be constitutively expressed, such as through an upstream activating iso-IS30 element [97], the spaFED genes lack this capacity and instead lay dormant. On the other hand, whether the expression of the spaFED operon is inducibly controlled via some sort of stimulus-responsive promoter is not yet certain [12]. For instance, although varying the conditions of growth (e.g., nutrients and temperature) did not trigger the activation of the spaFED operon in L. rhamnosus GG [90], gene expression might still be induced by other environmental stimuli, or then in situ by the intestine as reported for the Tad (tight adherence) pili of the Bifidobacterium breve UCC2003 [120].
Nonetheless, in the Rintahaka et al. (2014) study, the near-upstream DNA region of the spaFED operon from 13 different L. rhamnosus strains was aligned and examined in silico for the presence of any canonical regulatory elements [98]. For this, sequence found directly upstream of the coding region for the spaF gene had corresponded to a ribosomal binding site (RBS), which itself was also identified for the spaE, spaD, and srtC2 genes. As it was also found that encoded for each of these pilus genes is an open reading frame without any premature stop codons, the spaFED operon appears to be a complete translational unit, and thus at this level there is no reason preventing a native (or recombinant) pilus from ultimately being assembled together. Added scrutiny of DNA sequence further upstream had not identified a symmetric operator site for binding a repressor protein, which seemed to rule out inducible gene expression requiring this type of regulatory element [98]. However, while this study did locate the consensus sequence for the -10 and -35 promoter recognition sites, along with possible nucleotides for a transcriptional start site, additional sequence elements for either inducible or constitutive gene expression were not evident. This upstream region had also lacked the DNA elements associated with the attenuation mechanism for controlling S. gallolyticus pilus expression [35]. Moreover, since a stem-loop DNA motif for a possible transcriptional terminator was not identified anywhere between the -10 and -35 promoter region and the start of the spaF gene, a non-expressible spaFED operon is less likely to occur for this reason, but which alternatively might be explained by regulatory sequence that has become somehow corrupted.
Although results from a pan-genome study of L. rhamnosus [12], and similarly of L. casei and L. paracasei [92,93], suggest that the spaFED-related genes are a commonality in various strains and part of the core genome, it remains puzzling why the spaFED operon would still persist in the genome, particularly as without any pilus gene products this would not offer cells an adaptive advantage or fitness benefit. Yet, as it seems carrying the spaFED operon does not impose an additional genetic load or liability on the casei group of lactobacilli, the corresponding genes have resisted the evolutionary forces of loss or decay, and thus continue to remain a constant genomic attribute, though for what cellular purpose or specific function is uncertain.
5.2. Structure
The undocumented status of the SpaFED pilus in the L. rhamnosus, L. casei, and L. paracasei species has precluded the structural characterization of its native form. However, as the spaFED operon was predicted to be an intact translational unit [98], it was undertaken to clone and express the spaFED-related genes of L. rhamnosus GG in the recombinant host of L. lactis. Based on results from electrophoretic immunoblotting and immuno-EM, it was found that the spaF, spaE, spaD, and srtC2 genes of the spaFED operon were readily transcribed and translated recombinantly in lactococcal cells to produce a surface-assembled pilus [98]. From single and double immunogold-labeling experiments, it was revealed that the spaFED operon encodes a prototypical sortase-dependent pilus, for which the backbone SpaD (~51 kDa), basal SpaE (~45 kDa), and tip SpaF (~104 kDa) pilin subunits are structurally located in their expected sites. However, while it appeared that the ancillary SpaF and SpaE pilins are also present along the length of the SpaFED pilus backbone, they were interpreted to be a gold-particle immunolabeling artifact [98].
To date, there are no reports in the literature that any of the SpaFED pilins has had its tertiary structure solved by X-ray crystallographic methods. However, there are two published studies describing the successful crystallization of the backbone SpaD [121] and basal SpaE [122] subunits, along with their preliminary X-ray diffraction analysis. For SpaD, sodium iodide-soaked orthorhombic crystals yielded diffraction data to a resolution of 2.2 Å from which an interpretable electron-density map was generated by single-wavelength anomalous diffraction (SAD) phasing [121]. For SpaE and its SAD phasing, selenium-substituted orthorhombic crystals were able to diffract to 1.8 Å, with a reasonably strong anomalous signal observed in the X-ray data [122]. Correspondingly, model building and refinement are said to be in progress for both types of pilin structures.
5.3. Adhesion
Aside from the presence of domains for the CnaA or CnaB Ig-like fold as a structural stalk, none of the primary structures for the L. rhamnosus SpaFED pilins are predicted to have homology to a familiar binding domain [11]. However, based on its molecular size and thus likely location at the pilus tip, the SpaF pilin would otherwise be expected to have binding properties. This in fact was established in the study by von Ossowski et al. (2010), with the finding that recombinant SpaF protein is able to adhere to intestinal mucus and at a level similar to that of SpaC pilin [102]. Moreover, as neither basal SpaE nor backbone SpaD was shown to have the same adhesion capacity, it would appear that the SpaF subunit is solely responsible for the mucus-binding property of the SpaFED pilus. This latter point was subsequently confirmed in the Rintahaka et al. (2014) study through the use of lactococcal clones expressing WT and SpaF-deleted SpaFED pili [98]. Here, it was revealed that the lactococcal cells with WT SpaFED pili had bound readily to mucus, which was related to the SpaF pilin since the lactococci having SpaF-deleted pili could no longer display a similar level of substrate specificity. Moreover, by using the same two SpaFED-piliated lactococcal clones, it was also shown that the SpaFED pilus can adhere to ECM proteins (i.e., fibronectin and collagens I and IV) and human intestinal Caco-2 and HT-29 epithelial cells, where in all cases the main determinant of binding was judged to be the SpaF tip pilin. Surprisingly though, in another study [14], it was found that the binding capacity of the SpaFED pilus did not lend itself to helping form biofilm, as this ability was not detectable with the SpaFED-piliated lactococci. Apart from this latter finding, in terms of adhesive interactions with mucus, ECM proteins, and gut epithelial cells, the SpaFED pilus can still be seen as functionally akin to its SpaCBA counterpart.
5.4. Immunogenicity
Again, through the use of WT and SpaF-deleted SpaFED-piliated lactococcal clones [98], SpaFED piliation was examined for any innate immunogenic properties, much like was done previously for the SpaCBA pilus [104]. For the immuno-characterization of the SpaFED pilus, it was examined whether the two SpaFED-piliated lactococcal constructs can prompt a TLR2 response, which was inferred from the measured NF-κB activation in HEK-TLR2 cells. In addition, these SpaFED-piliated lactococci were tested for their effect on the endogenic levels of IL-8 in Caco-2 cells. Surprisingly, the WT SpaFED pilus did not produce the same stimulatory effects as the SpaCBA pilus or as the bifidobacterial [15] and streptococcal [83] sortase-dependent pili, but rather instead it behaved in an opposite manner by dampening the levels of TLR2-induced activation and IL-8 production [98]. Somewhat intriguingly, this dampening influence was not observed with the SpaF-deleted pilus, which seemed to suggest the SpaF subunit might have an adhesive-driven role in lowering the tested immune responses [98]. In the speculative scenario of the gut, a natively produced SpaFED pilus would thus offer an opposing innate immunogenic role than that of the SpaCBA pilus, but granted if these two pilus types were to be expressed at the same time, e.g., by L. rhamnosus GG cells, it is conceivable that they could contribute to a state of immune tolerance by providing a localized counterbalance to the induced high-and-low immuno-responsiveness of the host [98].
6. LrpCBA Piliation
The third type of sortase-dependent pilus in lactobacilli is associated with the L. ruminis species and called LrpCBA, where its prefix name is derived from “L. ruminis pilus” [14]. In comparative terms, L. ruminis as a surface-piliated host is quite unlike the L. rhamnosus species since it is one of the few lactobacilli that is motile by flagella [95,123,124,125], strictly anaerobic [126], and confined to only the gut environment [124], where in fact its colonization of this ecological niche is considered autochthonous (indigenous) [127,128]. Although the related pilus loci had been identified in the L. ruminis genome as early as 2009 (unpublished observations; see [14]) and then later in 2011 in the published work by Forde et al. (2011) [95], it was only until 2015 when Yu and colleagues produced results confirming that LrpCBA pilus structures were actively produced by this lactobacillar species [14]. Thus far, the majority of work done to characterize the LrpCBA pilus comes from this particular study on L. ruminis and, for this reason, the various properties determined for it are limited and not as far-reaching as for SpaCBA piliation.
6.1. Genetics
Genetically, the LrpCBA pilus follows suit with other sortase-dependent pili by having its genes clustered into an operon [14,129]. Here, the loci for the predicted tip (LrpC), basal (LrpB), and backbone (LrpA) pilins and C-type sortase (SrtC) are arranged in tandem on the L. ruminis genome as the fimbrial lrpCBA operon, i.e., lrpC-lrpB-lrpA-srtC [14,129]. Almost invariably, every strain of L. ruminis that has been examined so far is containing the lrpCBA loci, and based on a pan-genome survey of strains from various host sources (human, bovine, porcine, and equine) all of them but the lrpB gene are found to be part of the core genome. Of note, lrpB was excluded from the core genome because of certain nucleotide inconsistencies in the bovine- and equine-sourced versions that caused a reading-frameshift change to be present [129]. However, as it remains uncertain if these represent an authentic indel mutation or a DNA sequencing error, lrpB might itself be put among the L. ruminis core genes [129].
Results from the Yu et al. (2015) study [14] indicated that the primary structures of the lrpC, lrpB, and lrpA genes from the human-derived L. ruminis ATCC 25644 strain exhibit the typical sequence characteristics of a Gram-positive pilin subunit, such as an E-box and pilin motif, along with the established domains for both N-terminal secretion signaling and C-terminal sortase recognition (Table 1). Further, while an amino acid sequence alignment of the predicted lrpCBA-encoded proteins from human, bovine, porcine, and equine isolates of L. ruminis showed a high level of shared identity (e.g., ranging from 89.7% to 100%), this was unobserved with alignments of the L. rhamnosus GG spaCBA- and spaFED-encoded pilus proteins [14]. This finding lends support to LrpCBA piliation being a third lactobacillar pilus type. Of added interest, the lrpCBA operon appears to be genome-specific to only L. ruminis, as a BlastP search of the NCBI database using the LrpCBA pilin-proteins revealed no counterparts in any other Lactobacillus species [14]. This observation contrasts with that of the spaCBA and spaFED operons, both of which can be found in the genomes of more than one species, i.e., L. rhamnosus, L. casei, and L. paracasei [12,92,93,94].
As the microarray analysis of L. ruminis cells had already indicated the lrpCBA genes are actively transcribed, for which their expression is more up-regulated in a human-isolated strain than one originating from a bovine source [95], this was followed up by a study that examined the DNA region that precedes the lrpC gene for the presence of any possible regulatory elements [14]. Based on an aligned 106-bp DNA segment from the genomes of various host strains of L. ruminis (human, bovine, porcine, and equine), a putative RBS motif of 5′-GGAGAG-3′ and a pair of hexanucleotide sequences resembling the -10 and -35 consensus promoter regions were identified [14]. In addition, a candidate purine nucleotide was proposed as a possible starting point for transcribing the genes of the lrpCBA operon [14]. Based on these findings it was reasonably concluded that such sequence elements are what would presumably regulate constitutive lrpCBA gene expression in L. ruminis.
6.2. Structure
Given that active expression of the lrpCBA genes had been reported to occur in L. ruminis cells [95], there was all likelihood that the corresponding transcripts would be translatable into the specific protein products needed for building up the LrpCBA pilus. This in fact was the outcome as the constitutive production of “native” LrpCBA pili was observed on the cell surface of L. ruminis [14] (Figure 3). Convincing evidence of the LrpCBA pilus as a fully assembled surface-anchored structure was initially demonstrated for the human-isolated ATCC 25644 strain [14] and subsequently for the porcine GRL1172 strain [130] by using the customary immunoblotting and immuno-EM approaches employed for the same purpose with the SpaCBA and SpaFED pili [11,90,98]. Here, through single and double immunogold-labeling of pilin-proteins, EM had helped visualize LrpCBA piliation in the ATCC 25644 strain as being typical of other sortase-dependent pili, with the tip LrpC (~123 kDa), basal LrpB (~39 kDa), and backbone LrpA (~49 kDa) pilins assuming their predicted roles in the assembled structure [14]. Once again, though, the immuno-labeling artifact of ancillary pilins (LrpC and LrpB) along the pilus backbone seemed apparent. Additionally, the electron microscopic examination of the overall L. ruminis cell population revealed that the outer surface of most cells is piliated, with an average of one to five pili per bacterium [14], which is on the order of ten-fold less than estimated for L. rhamnosus GG SpaCBA piliation [11]. Further, by comparison, the EM-deduced structure of the recombinantly produced LrpCBA pilus in L. lactis is remarkably authentic looking, notwithstanding its tendency to be somewhat lengthier than the native version [14].
When a subunit size comparison is made between the various types of LrpCBA pilins and their counterparts in the SpaCBA and SpaFED pili, LrpC, SpaE, and SpaD are the largest (123, 45, and 51 kDa, respectively), whereas SpaC, SpaB, and SpaA are the smallest (90, 20, and 30 kDa, respectively) [11,14,90,98]. Put in the context of the polymerized backbone pilins, the SpaCBA pilus can be projected as retaining the narrowest structural width, with the LrpCBA and SpaFED pili both then being wider and of somewhat similar thickness. Thus, at the molecular level, for the two pilus types that are known to be natively produced by cells of L. ruminis and L. rhamnosus, one might interpret their potential difference in backbone girth (i.e., inferred from the subunit sizes 49 kDa for LrpA and 30 kDa for SpaA) most likely points to the LrpCBA pilus having the structural characteristics and ability to better tolerate the environmental stress of the intestinal habitat, e.g., by being thicker and stronger, and thus less susceptible to breakage or shearing at longer lengths. Consequently, it is more than tempting to speculate that from the autochthonous perspective of host colonization, this particular aspect of the LrpCBA pilus structure can confer an adaptive advantage or gain to gut-dwelling L. ruminis cells.
6.3. Adhesion
Among the three types of LrpCBA pilins, only the LrpC primary structure is homologous to a protein domain that is predicted to be substrate binding. Here, the tip LrpC pilin has homology to a collagen-binding domain, and one which differs from those CnaA or CnaB domains that serve as a stalk-like component of most Gram-positive pilins [14]. This predicted homology contrasts with the L. rhamnosus SpaC and SpaF tip pilins, as neither shares any sequence similarity with collagen-binding proteins [11,112], albeit that both were ultimately shown to adhere to collagen substrates [98,108]. For the LrpCBA pilus, its affinity for collagen protein was demonstrated by using recombinant lactococcal clones that produce WT and LrpC-deleted LrpCBA pili [14]. Here, it was observed that the WT LrpCBA pilus can readily adhere to collagen, binding more to type I than type IV [14]. As adhesion to these collagen types is substantially diminished for the LrpC-deleted pili, this suggested the LrpC subunit has a requisite adhesive role in the binding of collagen by LrpCBA pili [14]. In the context of L. ruminis itself, this adhesion capacity of the LrpCBA pilus is in keeping with the collagen-binding ability found with the ATCC 25644 strain [14] and other host-sourced isolates [130]. As L. ruminis cells can also adhere to fibronectin [14,130], the ability to do so was partially attributed to the LrpCBA pilus, in which LrpC acts as the main source of binding [14], and in spite of its lack of homology to any other fibronectin-binding proteins [14,129]. Likewise, a similar trend was observed for the binding of the LrpCBA pilus to human Caco-2 and HT-29 gut epithelial cells [14]. However, unlike found for the SpaCBA and SpaFED piliation [11,98,112], the LrpCBA pilus lacks the ability to be mucoadhesive [14]. Although this nonbinding ability toward mucus is a clear departure from the other lactobacillar pili, it remains consistent with the fact that there are no encoded proteins in the L. ruminis genome that display any similarity to proven mucus-binding domains [129] and that L. ruminis (e.g., the ATCC 25644 strain) is itself an extremely poor binder of mucus [14]. Interestingly, the functionality of LrpCBA piliation also departs from that of the SpaCBA pilus [14,106] and other types of Gram-positive pili [131,132] as evidence suggests it has no adhesive role in the self-aggregation of cells to produce biofilm, even though L. ruminis demonstrates a strong biofilm-forming phenotype [14]. Thus it seems that while the formation into biofilm communities by some piliated bacteria is seen as adaptively advantageous during niche development and expansion, for L. ruminis cells their LrpCBA piliation would have no particular contribution to such ecological processes.
Then again, for the ecological aspects underlying the gut autochthony of the L. ruminis species, the bi-functional binding specificity of the LrpCBA pilus toward collagen and fibronectin proteins represents an effective colonization strategy for bacterial attachment to the intestinal epithelium, and overtly to any exposed location along the ECM layer (Figure 4). Together with its other characteristics (e.g., motility, poor mucoadhesiveness, and strict anaerobiosis), LrpCBA-piliated L. ruminis would be well-equipped to avoid and get past the mucosal barrier and then establish a stable cell population in the specific micro-environment of folded and deoxygenated crevices found throughout the epithelium lining of the gut. Undoubtedly, the first-contact ability of an ECM-adhesive LrpCBA pilus can contribute to helping promote and support the indigenous lifestyle of L. ruminis in the GI tract.
6.4. Immunogenicity
While there are some published accounts characterizing the innate immunogenic potential of L. ruminis cells [14,130,133], only one study thus far has specifically scrutinized the related role played by LrpCBA piliation [14]. This investigation was performed similarly to what had been done for the L. rhamnosus SpaCBA and SpaFED pili [98,104] and involved the use of recombinant LrpCBA-piliated Lactococcus constructs, one WT and another LrpC-deleted. Interestingly, although live cells of the L. ruminis ATCC 25644 strain were able to potentiate increased TLR2-dependent NF-κB signaling and endogenic production of IL-8 in the recombinant HEK-TLR2 cell line, the WT LrpCBA pilus aroused the opposite effect and had a dampening influence instead [14]. A similar result occurred for the endogenous IL-8 secretion in human intestinal Caco-2 cells, as some moderate lowering of the levels was observed with WT LrpCBA pili [14]. However, whereas these findings were in accordance with the immuno-dampening exerted by the SpaFED pilus [98], there was no causal role for the tip LrpC subunit since its removal from the LrpCBA pilus had no counter influence. This contrasts with the tip SpaC and SpaF pilins of the SpaCBA and SpaFED pili, respectively, as their presence and intrinsic adhesiveness appeared necessary for any immunomodulatory activities [98,104]. Seemingly, as the LrpC constituent of the LrpCBA pilus bears no direct responsibility for the dampened immune responses from intestinal or immune-related cells, it was suggested this diminishing effect could be based on the recognition of the overall topological fold for the pilus structure [14]. On the other hand, although there appears to be an LrpCBA pilus-mediated dampening of IL-8 levels, another study reported that the interaction between TLR5 and the flagella of L. ruminis stimulates increased IL-8 secretion from Caco-2 cells [123]. In practice, this would suggest that via the immunostimulatory capacity of their pili and flagella, L. ruminis cells are able to simultaneously fluctuate the production of IL-8 from host intestinal cells, which in itself might represent a type of homeostatic counterbalancing mechanism that allows for host immune tolerance and, in doing so, an autochthonous presence in the gut [14].
7. Conclusions
Irrespective of its Gram-positive host, the sortase-dependent pilus can be viewed as a molecular engineering coup for those bacteria whose outer surfaces display this particularly long and adhesive protrusion. Undoubtedly, the building-block simplicity of piecing together various role-defined pilin subunits represents a successful genetic and evolutionary adaptive strategy for such bacteria to gain their initial foothold in a biological niche or habitat. For the three types of lactobacillar piliation presented in this review (i.e., SpaCBA, SpaFED and LrpCBA), each has been described as having the basic characteristics common to all sortase-dependent pili, but as well, certain unique properties and associated actions that are inherent to them. As for this latter point, it is likely these pilus types exhibit some of the phenotypic and physiological variation that conveys an ecological advantage to their respective bacterial hosts. However, still remaining unresolved is what native role might be played by the SpaFED piliation of the casei group of lactobacilli as well as what additional number of Lactobacillus species predicted as containing sortase-dependent pilus genes are in fact surface-piliated. In all likelihood, by tackling and solving these challenges this will open up new chapters of scientific discovery to the continuing and unfolding story of sortase-dependent pili in Gram-positive commensal bacteria.
Acknowledgments
The author acknowledges the funding support from an Academy of Finland research grant (No. 277362) awarded to Emeritus Airi Palva (Department of Veterinary Biosciences, Faculty of Veterinary Medicine, University of Helsinki). The author also acknowledges Mervi Lindman at the Institute of Biotechnology Electron Microscopy Unit (University of Helsinki) for her helpful advice and assistance.
Author Contributions
This review was conceived and written by the author alone. The author has also approved the final version for publication.
Conflicts of Interest
The author declares no conflict of interest.
References
- Kline, K.A.; Dodson, K.W.; Caparon, M.G.; Hultgren, S.J. A tale of two pili: Assembly and function of pili in bacteria. Trends Microbiol. 2010, 18, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Proft, T.; Baker, E.N. Pili in Gram-negative and Gram-positive bacteria—Structure, assembly and their role in disease. Cell. Mol. Life Sci. 2009, 66, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Duguid, J.P.; Smith, I.W.; Dempster, G.; Edmunds, P.N. Non-flagellar filamentous appendages (fimbriae) and haemagglutinating activity in Bacterium coli. J. Pathol. Bacteriol. 1955, 70, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Fronzes, R.; Remaut, H.; Waksman, G. Architectures and biogenesis of non-flagellar protein appendages in Gram-negative bacteria. EMBO J. 2008, 27, 2271–2280. [Google Scholar] [CrossRef] [PubMed]
- Yanagawa, R.; Otsuki, K. Some properties of the pili of Corynebacterium renale. J. Bacteriol. 1970, 101, 1063–1069. [Google Scholar] [PubMed]
- Yanagawa, R.; Otsuki, K.; Tokui, T. Electron microscopy of fine structure of Corynebacterium renale with special reference to pili. Jpn. J. Vet. Res. 1968, 16, 31–37. [Google Scholar] [PubMed]
- Baker, E.N.; Squire, C.J.; Young, P.G. Self-generated covalent cross-links in the cell-surface adhesins of Gram-positive bacteria. Biochem. Soc. Trans. 2015, 43, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Hilleringmann, M.; Ringler, P.; Müller, S.A.; de Angelis, G.; Rappuoli, R.; Ferlenghi, I.; Engel, A. Molecular architecture of Streptococcus pneumoniae TIGR4 pili. EMBO J. 2009, 28, 3921–3930. [Google Scholar] [CrossRef] [PubMed]
- Soriani, M.; Telford, J.L. Relevance of pili in pathogenic streptococci pathogenesis and vaccine development. Future Microbiol. 2010, 5, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Douillard, F.P.; Ribbera, A.; Järvinen, H.M.; Kant, R.; Pietilä, T.E.; Randazzo, C.; Paulin, L.; Laine, P.K.; Caggia, C.; von Ossowski, I.; et al. Comparative genomic and functional analysis of Lactobacillus casei and Lactobacillus rhamnosus strains marketed as probiotics. Appl. Environ. Microbiol. 2013, 79, 1923–1933. [Google Scholar] [CrossRef] [PubMed]
- Kankainen, M.; Paulin, L.; Tynkkynen, S.; von Ossowski, I.; Reunanen, J.; Partanen, P.; Satokari, R.; Vesterlund, S.; Hendrickx, A.P.; Lebeer, S.; et al. Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc. Natl. Acad. Sci. USA 2009, 106, 17193–17198. [Google Scholar] [CrossRef] [PubMed]
- Kant, R.; Rintahaka, J.; Yu, X.; Sigvart-Mattila, P.; Paulin, L.; Mecklin, J.P.; Saarela, M.; Palva, A.; von Ossowski, I. A comparative pan-genome perspective of niche-adaptable cell-surface protein phenotypes in Lactobacillus rhamnosus. PLoS ONE 2014, 9, e102762. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Verhoeven, T.L.; Francius, G.; Schoofs, G.; Lambrichts, I.; Dufrêne, Y.; Vanderleyden, J.; De Keersmaecker, S.C. Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional analysis of the priming glycosyltransferase. Appl. Environ. Microbiol. 2009, 75, 3554–3563. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jaatinen, A.; Rintahaka, J.; Hynönen, U.; Lyytinen, O.; Kant, R.; Åvall-Jääskeläinen, S.; von Ossowski, I.; Palva, A. Human gut-commensalic Lactobacillus ruminis ATCC 25644 displays sortase-assembled surface piliation: Phenotypic characterization of its fimbrial operon through in silico predictive analysis and recombinant expression in Lactococcus lactis. PLoS ONE 2015, 10, e0145718. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Serafini, F.; Foroni, E.; Duranti, S.; O’Connell Motherway, M.; Taverniti, V.; Mangifesta, M.; Milani, C.; Viappiani, A.; Roversi, T.; et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions. Proc. Natl. Acad. Sci. USA 2013, 110, 11151–11156. [Google Scholar] [CrossRef] [PubMed]
- Danne, C.; Dramsi, S. Pili of Gram-positive bacteria: Roles in host colonization. Res. Microbiol. 2012, 163, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Baker, E.N. Structure and assembly of Gram-positive bacterial pili: Unique covalent polymers. Curr. Opin. Struct. Biol. 2012, 22, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V. Pilins in Gram-positive bacteria: A structural perspective. IUBMB Life 2015, 67, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Pansegrau, W.; Bagnoli, F. Pilus assembly in Gram-positive bacteria. Curr. Top. Microbiol. Immunol. 2017, 404, 203–233. [Google Scholar] [PubMed]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Pili in Gram-positive bacteria: Assembly, involvement in colonization and biofilm development. Trends Microbiol. 2008, 16, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Budzik, J.M.; Marraffini, L.A.; Schneewind, O. Assembly of pili on the surface of Bacillus cereus vegetative cells. Mol. Microbiol. 2007, 66, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Danne, C.; Entenza, J.M.; Mallet, A.; Briandet, R.; Débarbouillé, M.; Nato, F.; Glaser, P.; Jouvion, G.; Moreillon, P.; Trieu-Cuot, P.; et al. Molecular characterization of a Streptococcus gallolyticus genomic island encoding a pilus involved in endocarditis. J. Infect. Dis. 2011, 204, 1960–1970. [Google Scholar] [CrossRef] [PubMed]
- Rosini, R.; Rinaudo, C.D.; Soriani, M.; Lauer, P.; Mora, M.; Maione, D.; Taddei, A.; Santi, I.; Ghezzo, C.; Brettoni, C.; et al. Identification of novel genomic islands coding for antigenic pilus-like structures in Streptococcus agalactiae. Mol. Microbiol. 2006, 61, 126–141. [Google Scholar] [CrossRef] [PubMed]
- Ton-That, H.; Schneewind, O. Assembly of pili on the surface of Corynebacterium diphtheriae. Mol. Microbiol. 2003, 50, 1429–1438. [Google Scholar] [CrossRef] [PubMed]
- Bourgogne, A.; Singh, K.V.; Fox, K.A.; Pflughoeft, K.J.; Murray, B.E.; Garsin, D.A. EbpR is important for biofilm formation by activating expression of the endocarditis and biofilm-associated pilus operon (ebpABC) of Enterococcus faecalis OG1RF. J. Bacteriol. 2007, 189, 6490–6493. [Google Scholar] [CrossRef] [PubMed]
- Nallapareddy, S.R.; Singh, K.V.; Sillanpää, J.; Garsin, D.A.; Höök, M.; Erlandsen, S.L.; Murray, B.E. Endocarditis and biofilm-associated pili of Enterococcus faecalis. J. Clin. Investig. 2006, 116, 2799–2807. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Köller, T.; Moritz, K.; Ribardo, D.; Jonas, L.; McIver, K.S.; Sumitomo, T.; Terao, Y.; Kawabata, S.; Podbielski, A.; et al. Mode of expression and functional characterization of FCT-3 pilus region-encoded proteins in Streptococcus pyogenes serotype M49. Infect. Immun. 2009, 77, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Barocchi, M.A.; Ries, J.; Zogaj, X.; Hemsley, C.; Albiger, B.; Kanth, A.; Dahlberg, S.; Fernebro, J.; Moschioni, M.; Masignani, V.; et al. A pneumococcal pilus influences virulence and host inflammatory responses. Proc. Natl. Acad. Sci. USA 2006, 103, 2857–2862. [Google Scholar] [CrossRef] [PubMed]
- Ito, H.; Ono, E.; Yanagawa, R. Comparison of surface hydrophobicity of piliated and non-piliated clones of Corynebacterium renale and Corynebacterium pilosum. Vet. Microbiol. 1987, 14, 165–171. [Google Scholar] [CrossRef]
- Basset, A.; Turner, K.H.; Boush, E.; Sayeed, S.; Dove, S.L.; Malley, R. Expression of the type 1 pneumococcal pilus is bistable and negatively regulated by the structural component RrgA. Infect. Immun. 2011, 79, 2974–2983. [Google Scholar] [CrossRef] [PubMed]
- Basset, A.; Turner, K.H.; Boush, E.; Sayeed, S.; Dove, S.L.; Malley, R. An epigenetic switch mediates bistable expression of the type 1 pilus genes in Streptococcus pneumoniae. J. Bacteriol. 2012, 194, 1088–1091. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, G.; Moschioni, M.; Muzzi, A.; Pezzicoli, A.; Censini, S.; Delany, I.; Lo Sapio, M.; Sinisi, A.; Donati, C.; Masignani, V.; et al. The Streptococcus pneumoniae pilus-1 displays a biphasic expression pattern. PLoS ONE 2011, 6, e21269. [Google Scholar] [CrossRef] [PubMed]
- Kreikemeyer, B.; Gámez, G.; Margarit, I.; Giard, J.C.; Hammerschmidt, S.; Hartke, A.; Podbielski, A. Genomic organization, structure, regulation and pathogenic role of pilus constituents in major pathogenic Streptococci and Enterococci. Int. J. Med. Microbiol. 2011, 301, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Hava, D.L.; Hemsley, C.J.; Camilli, A. Transcriptional regulation in the Streptococcus pneumoniae rlrA pathogenicity islet by RlrA. J. Bacteriol. 2003, 185, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Danne, C.; Dubrac, S.; Trieu-Cuot, P.; Dramsi, S. Single cell stochastic regulation of pilus phase variation by an attenuation-like mechanism. PLoS Pathog. 2014, 10, e1003860. [Google Scholar] [CrossRef] [PubMed]
- Pancotto, L.; de Angelis, G.; Bizzarri, E.; Barocchi, M.A.; del Giudice, G.; Moschioni, M.; Ruggiero, P. Expression of the Streptococcus pneumoniae pilus-1 undergoes on and off switching during colonization in mice. Sci. Rep. 2013, 3, 2040. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, A.P.; Budzik, J.M.; Oh, S.Y.; Schneewind, O. Architects at the bacterial surface—Sortases and the assembly of pili with isopeptide bonds. Nat. Rev. Microbiol. 2011, 9, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Marraffini, L.A.; Dedent, A.C.; Schneewind, O. Sortases and the art of anchoring proteins to the envelopes of Gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2006, 70, 192–221. [Google Scholar] [CrossRef] [PubMed]
- Spirig, T.; Weiner, E.M.; Clubb, R.T. Sortase enzymes in Gram-positive bacteria. Mol. Microbiol. 2011, 82, 1044–1059. [Google Scholar] [CrossRef] [PubMed]
- Schneewind, O.; Missiakas, D. Sec-secretion and sortase-mediated anchoring of proteins in Gram-positive bacteria. Biochim. Biophys. Acta 2014, 1843, 1687–1697. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, R.; Malito, E.; Nuccitelli, A.; D’Onofrio, M.; Martinelli, M.; Ferlenghi, I.; Grandi, G.; Telford, J.L.; Maione, D.; Rinaudo, C.D. Structure analysis and site-directed mutagenesis of defined key residues and motives for pilus-related sortase C1 in group B Streptococcus. FASEB J. 2011, 25, 1874–1886. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, G.; Ton-That, H.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Khare, B.; Krishnan, V.; Rajashankar, K.R.; I-Hsiu, H.; Xin, M.; Ton-That, H.; Narayana, S.V. Structural differences between the Streptococcus agalactiae housekeeping and pilus-specific sortases: SrtA and SrtC1. PLoS ONE 2011, 6, e22995. [Google Scholar] [CrossRef] [PubMed]
- Necchi, F.; Nardi-Dei, V.; Biagini, M.; Assfalg, M.; Nuccitelli, A.; Cozzi, R.; Norais, N.; Telford, J.L.; Rinaudo, C.D.; Grandi, G.; et al. Sortase a substrate specificity in GBS pilus 2a cell wall anchoring. PLoS ONE 2011, 6, e25300. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, A.; Mandlik, A.; Swierczynski, A.; Gaspar, A.; Das, A.; Ton-That, H. Housekeeping sortase facilitates the cell wall anchoring of pilus polymers in Corynebacterium diphtheriae. Mol. Microbiol. 2007, 66, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Budzik, J.M.; Oh, S.Y.; Schneewind, O. Cell wall anchor structure of BcpA pili in Bacillus anthracis. J. Biol. Chem. 2008, 283, 36676–36686. [Google Scholar] [CrossRef] [PubMed]
- Mandlik, A.; Das, A.; Ton-That, H. The molecular switch that activates the cell wall anchoring step of pilus assembly in Gram-positive bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 14147–14152. [Google Scholar] [CrossRef] [PubMed]
- Dramsi, S.; Caliot, E.; Bonne, I.; Guadagnini, S.; Prévost, M.C.; Kojadinovic, M.; Lalioui, L.; Poyart, C.; Trieu-Cuot, P. Assembly and role of pili in group B streptococci. Mol. Microbiol. 2006, 60, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, A.H.; Ton-That, H. Assembly of distinct pilus structures on the surface of Corynebacterium diphtheriae. J. Bacteriol. 2006, 188, 1526–1533. [Google Scholar] [CrossRef] [PubMed]
- Ton-That, H.; Marraffini, L.A.; Schneewind, O. Sortases and pilin elements involved in pilus assembly of Corynebacterium diphtheriae. Mol. Microbiol. 2004, 53, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, R.; Nuccitelli, A.; D’Onofrio, M.; Necchi, F.; Rosini, R.; Zerbini, F.; Biagini, M.; Norais, N.; Beier, C.; Telford, J.L.; et al. New insights into the role of the glutamic acid of the E-box motif in group B Streptococcus pilus 2a assembly. FASEB J. 2012, 26, 2008–2018. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Baker, E.N. Intramolecular isopeptide bonds give thermodynamic and proteolytic stability to the major pilin protein of Streptococcus pyogenes. J. Biol. Chem. 2009, 284, 20729–20737. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Coulibaly, F.; Clow, F.; Proft, T.; Baker, E.N. Stabilizing isopeptide bonds revealed in Gram-positive bacterial pilus structure. Science 2007, 318, 1625–1628. [Google Scholar] [CrossRef] [PubMed]
- Deivanayagam, C.C.; Rich, R.L.; Carson, M.; Owens, R.T.; Danthuluri, S.; Bice, T.; Höök, M.; Narayana, S.V. Novel fold and assembly of the repetitive B region of the Staphylococcus aureus collagen-binding surface protein. Structure 2000, 8, 67–78. [Google Scholar] [CrossRef]
- Symersky, J.; Patti, J.M.; Carson, M.; House-Pompeo, K.; Teale, M.; Moore, D.; Jin, L.; Schneider, A.; DeLucas, L.J.; Höök, M.; et al. Structure of the collagen-binding domain from a Staphylococcus aureus adhesin. Nat. Struct. Biol. 1997, 4, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Paterson, N.G.; Gaspar, A.H.; Ton-That, H.; Baker, E.N. The Corynebacterium diphtheriae shaft pilin SpaA is built of tandem Ig-like modules with stabilizing isopeptide and disulfide bonds. Proc. Natl. Acad. Sci. USA 2009, 106, 16967–16971. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Baker, E.N. Intramolecular isopeptide bonds: Protein crosslinks built for stress? Trends Biochem. Sci. 2011, 36, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Izoré, T.; Contreras-Martel, C.; El Mortaji, L.; Manzano, C.; Terrasse, R.; Vernet, T.; Di Guilmi, A.M.; Dessen, A. Structural basis of host cell recognition by the pilus adhesin from Streptococcus pneumoniae. Structure 2010, 18, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Dwivedi, P.; Kim, B.J.; Samal, A.; Macon, K.; Ma, X.; Mishra, A.; Doran, K.S.; Ton-That, H.; Narayana, S.V. Structure of Streptococcus agalactiae tip pilin GBS104: A model for GBS pili assembly and host interactions. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1073–1089. [Google Scholar] [CrossRef] [PubMed]
- Linke-Winnebeck, C.; Paterson, N.G.; Young, P.G.; Middleditch, M.J.; Greenwood, D.R.; Witte, G.; Baker, E.N. Structural model for covalent adhesion of the Streptococcus pyogenes pilus through a thioester bond. J. Biol. Chem. 2014, 289, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Pointon, J.A.; Smith, W.D.; Saalbach, G.; Crow, A.; Kehoe, M.A.; Banfield, M.J. A highly unusual thioester bond in a pilus adhesin is required for efficient host cell interaction. J. Biol. Chem. 2010, 285, 33858–33866. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Gaspar, A.H.; Ye, N.; Mandlik, A.; Ton-That, H.; Narayana, S.V. An IgG-like domain in the minor pilin GBS52 of Streptococcus agalactiae mediates lung epithelial cell adhesion. Structure 2007, 15, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Linke, C.; Young, P.G.; Kang, H.J.; Bunker, R.D.; Middleditch, M.J.; Caradoc-Davies, T.T.; Proft, T.; Baker, E.N. Crystal structure of the minor pilin FctB reveals determinants of group A streptococcal pilus anchoring. J. Biol. Chem. 2010, 285, 20381–20389. [Google Scholar] [CrossRef] [PubMed]
- Shaik, M.M.; Maccagni, A.; Tourcier, G.; Di Guilmi, A.M.; Dessen, A. Structural basis of pilus anchoring by the ancillary pilin RrgC of Streptococcus pneumoniae. J. Biol. Chem. 2014, 289, 16988–16997. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Paterson, N.G.; Kim, C.U.; Middleditch, M.; Chang, C.; Ton-That, H.; Baker, E.N. A slow-forming isopeptide bond in the structure of the major pilin SpaD from Corynebacterium diphtheriae has implications for pilus assembly. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1190–1201. [Google Scholar] [CrossRef] [PubMed]
- Vengadesan, K.; Narayana, S.V. Structural biology of Gram-positive bacterial adhesins. Protein Sci. 2011, 20, 759–772. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.V.; Guiton, P.S.; Kline, K.A.; Port, G.C.; Pinkner, J.S.; Neiers, F.; Normark, S.; Henriques-Normark, B.; Caparon, M.G.; Hultgren, S.J. The metal ion-dependent adhesion site motif of the Enterococcus faecalis EbpA pilin mediates pilus function in catheter-associated urinary tract infection. MBio 2012, 3, e00177-12. [Google Scholar] [CrossRef] [PubMed]
- Konto-Ghiorghi, Y.; Mairey, E.; Mallet, A.; Duménil, G.; Caliot, E.; Trieu-Cuot, P.; Dramsi, S. Dual role for pilus in adherence to epithelial cells and biofilm formation in Streptococcus agalactiae. PLoS Pathog. 2009, 5, e1000422. [Google Scholar] [CrossRef] [PubMed]
- Becherelli, M.; Manetti, A.G.; Buccato, S.; Viciani, E.; Ciucchi, L.; Mollica, G.; Grandi, G.; Margarit, I. The ancillary protein 1 of Streptococcus pyogenes FCT-1 pili mediates cell adhesion and biofilm formation through heterophilic as well as homophilic interactions. Mol. Microbiol. 2012, 83, 1035–1047. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, C.A.; Hynes, R.O. Distribution and evolution of von Willebrand/integrin a domains: Widely dispersed domains with roles in cell adhesion and elsewhere. Mol. Biol. Cell 2002, 13, 3369–3387. [Google Scholar] [CrossRef] [PubMed]
- Hilleringmann, M.; Giusti, F.; Baudner, B.C.; Masignani, V.; Covacci, A.; Rappuoli, R.; Barocchi, M.A.; Ferlenghi, I. Pneumococcal pili are composed of protofilaments exposing adhesive clusters of RrgA. PLoS Pathog. 2008, 4, e1000026. [Google Scholar] [CrossRef] [PubMed]
- Nallapareddy, S.R.; Sillanpää, J.; Mitchell, J.; Singh, K.V.; Chowdhury, S.A.; Weinstock, G.M.; Sullam, P.M.; Murray, B.E. Conservation of Ebp-type pilus genes among Enterococci and demonstration of their role in adherence of Enterococcus faecalis to human platelets. Infect. Immun. 2011, 79, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Sillanpää, J.; Nallapareddy, S.R.; Qin, X.; Singh, K.V.; Muzny, D.M.; Kovar, C.L.; Nazareth, L.V.; Gibbs, R.A.; Ferraro, M.J.; Steckelberg, J.M.; et al. A collagen-binding adhesin, Acb, and ten other putative MSCRAMM and pilus family proteins of Streptococcus gallolyticus subsp. gallolyticus (Streptococcus bovis group, biotype I). J. Bacteriol. 2009, 191, 6643–6653. [Google Scholar]
- Abbot, E.L.; Smith, W.D.; Siou, G.P.; Chiriboga, C.; Smith, R.J.; Wilson, J.A.; Hirst, B.H.; Kehoe, M.A. Pili mediate specific adhesion of Streptococcus pyogenes to human tonsil and skin. Cell. Microbiol. 2007, 9, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Corynebacterium diphtheriae employs specific minor pilins to target human pharyngeal epithelial cells. Mol. Microbiol. 2007, 64, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Manetti, A.G.; Zingaretti, C.; Falugi, F.; Capo, S.; Bombaci, M.; Bagnoli, F.; Gambellini, G.; Bensi, G.; Mora, M.; Edwards, A.M.; et al. Streptococcus pyogenes pili promote pharyngeal cell adhesion and biofilm formation. Mol. Microbiol. 2007, 64, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Aymeric, L.; du Merle, L.; Danne, C.; Robbe-Masselot, C.; Trieu-Cuot, P.; Sansonetti, P.; Dramsi, S. Streptococcus gallolyticus Pil3 pilus is required for adhesion to colonic mucus and for colonization of mouse distal colon. J. Infect. Dis. 2015, 212, 1646–1655. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Porrini, C.; du Merle, L.; Danne, C.; Robbe-Masselot, C.; Trieu-Cuot, P.; Dramsi, S. The pil3 pilus of Streptococcus gallolyticus binds to intestinal mucins and to fibrinogen. Gut Microbes 2016, 7, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Devarajan, B.; Reardon, M.E.; Dwivedi, P.; Krishnan, V.; Cisar, J.O.; Das, A.; Narayana, S.V.; Ton-That, H. Two autonomous structural modules in the fimbrial shaft adhesin FimA mediate Actinomyces interactions with streptococci and host cells during oral biofilm development. Mol. Microbiol. 2011, 81, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Wu, C.; Yang, J.; Cisar, J.O.; Das, A.; Ton-That, H. The Actinomyces oris type 2 fimbrial shaft FimA mediates co-aggregation with oral streptococci, adherence to red blood cells and biofilm development. Mol. Microbiol. 2010, 77, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Okahashi, N.; Nakata, M.; Terao, Y.; Isoda, R.; Sakurai, A.; Sumitomo, T.; Yamaguchi, M.; Kimura, R.K.; Oiki, E.; Kawabata, S.; et al. Pili of oral Streptococcus sanguinis bind to salivary amylase and promote the biofilm formation. Microb. Pathog. 2011, 50, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, D.G.; Bowler, P.G. Biofilm delays wound healing: A review of the evidence. Burns Trauma 2013, 1, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Basset, A.; Zhang, F.; Benes, C.; Sayeed, S.; Herd, M.; Thompson, C.; Golenbock, D.T.; Camilli, A.; Malley, R. Toll-like receptor (TLR) 2 mediates inflammatory responses to oligomerized RrgA pneumococcal pilus type 1 protein. J. Biol. Chem. 2013, 288, 2665–2675. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, D.; Carey, A.J.; Caliot, E.; Webb, R.I.; Layton, J.R.; Wang, Y.; Bohnsack, J.F.; Adderson, E.E.; Ulett, G.C. Phylogenetic lineage and pilus protein Spb1/SAN1518 affect opsonin-independent phagocytosis and intracellular survival of group B Streptococcus. Microbes Infect. 2011, 13, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Maisey, H.C.; Quach, D.; Hensler, M.E.; Liu, G.Y.; Gallo, R.L.; Nizet, V.; Doran, K.S. A group B streptococcal pilus protein promotes phagocyte resistance and systemic virulence. FASEB J. 2008, 22, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Papasergi, S.; Brega, S.; Mistou, M.Y.; Firon, A.; Oxaran, V.; Dover, R.; Teti, G.; Shai, Y.; Trieu-Cuot, P.; Dramsi, S. The GBS PI-2a pilus is required for virulence in mice neonates. PLoS ONE 2011, 6, e18747. [Google Scholar] [CrossRef] [PubMed]
- Pace, F.; Pace, M.; Quartarone, G. Probiotics in digestive diseases: Focus on Lactobacillus GG. Minerva Gastroenterol. Dietol. 2015, 61, 273–292. [Google Scholar] [PubMed]
- Ton-That, H.; Schneewind, O. Assembly of pili in Gram-positive bacteria. Trends Microbiol. 2004, 12, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Huang, I.H.; Hendrickx, A.P.; Ton-That, H. Visualization of Gram-positive bacterial pili. Methods Mol. Biol. 2013, 966, 77–95. [Google Scholar] [PubMed]
- Reunanen, J.; von Ossowski, I.; Hendrickx, A.P.; Palva, A.; de Vos, W.M. Characterization of the SpaCBA pilus fibers in the probiotic Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2012, 78, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Walker, A. Genome watch: Probiotics stick it to the man. Nat. Rev. Microbiol. 2009, 7, 843. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, J.R.; Neeno-Eckwall, E.C.; Stahl, B.; Tandee, K.; Cai, H.; Morovic, W.; Horvath, P.; Heidenreich, J.; Perna, N.T.; Barrangou, R.; et al. Analysis of the Lactobacillus casei supragenome and its influence in species evolution and lifestyle adaptation. BMC Genom. 2012, 13, 533. [Google Scholar] [CrossRef] [PubMed]
- Smokvina, T.; Wels, M.; Polka, J.; Chervaux, C.; Brisse, S.; Boekhorst, J.; van Hylckama Vlieg, J.E.; Siezen, R.J. Lactobacillus paracasei comparative genomics: Towards species pan-genome definition and exploitation of diversity. PLoS ONE 2013, 8, e68731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toh, H.; Oshima, K.; Nakano, A.; Takahata, M.; Murakami, M.; Takaki, T.; Nishiyama, H.; Igimi, S.; Hattori, M.; Morita, H. Genomic adaptation of the Lactobacillus casei group. PLoS ONE 2013, 8, e75073. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.M.; Neville, B.A.; O’Donnell, M.M.; Riboulet-Bisson, E.; Claesson, M.J.; Coghlan, A.; Ross, R.P.; O’Toole, P.W. Genome sequences and comparative genomics of two Lactobacillus ruminis strains from the bovine and human intestinal tracts. Microb. Cell. Fact. 2011, 10, S13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Harris, H.M.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef] [PubMed]
- Douillard, F.P.; Ribbera, A.; Kant, R.; Pietilä, T.E.; Järvinen, H.M.; Messing, M.; Randazzo, C.L.; Paulin, L.; Laine, P.; Ritari, J.; et al. Comparative genomic and functional analysis of 100 Lactobacillus rhamnosus strains and their comparison with strain GG. PLoS Genet. 2013, 9, e1003683. [Google Scholar] [CrossRef] [PubMed]
- Rintahaka, J.; Yu, X.; Kant, R.; Palva, A.; von Ossowski, I. Phenotypical analysis of the Lactobacillus rhamnosus GG fimbrial spaFED operon: Surface expression and functional characterization of recombinant SpaFED pili in Lactococcus lactis. PLoS ONE 2014, 9, e113922. [Google Scholar] [CrossRef] [PubMed]
- Douillard, F.P.; Ribbera, A.; Xiao, K.; Ritari, J.; Rasinkangas, P.; Paulin, L.; Palva, A.; Hao, Y.; de Vos, W.M. Polymorphisms, chromosomal rearrangements, and mutator phenotype development during experimental evolution of Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 2016, 82, 3783–3792. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, P.; Pratap, S.; von Ossowski, I.; Palva, A.; Krishnan, V. New insights about pilus formation in gut-adapted Lactobacillus rhamnosus GG from the crystal structure of the SpaA backbone-pilin subunit. Sci. Rep. 2016, 6, 28664. [Google Scholar] [CrossRef] [PubMed]
- Douillard, F.P.; Rasinkangas, P.; von Ossowski, I.; Reunanen, J.; Palva, A.; de Vos, W.M. Functional identification of conserved residues involved in Lactobacillus rhamnosus strain GG sortase specificity and pilus biogenesis. J. Biol. Chem. 2014, 289, 15764–15775. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, R.; Prigozhin, D.; Rosini, R.; Abate, F.; Bottomley, M.J.; Grandi, G.; Telford, J.L.; Rinaudo, C.D.; Maione, D.; Alber, T. Structural basis for group B Streptococcus pilus 1 sortases C regulation and specificity. PLoS ONE 2012, 7, e49048. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; von Ossowski, I.; Palva, A.; Krishnan, V. Purification, crystallization and preliminary crystallographic analysis of the SpaA backbone-pilin subunit from probiotic Lactobacillus rhamnosus GG. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2013, 69, 1182–1185. [Google Scholar] [CrossRef] [PubMed]
- Von Ossowski, I.; Pietilä, T.E.; Rintahaka, J.; Nummenmaa, E.; Mäkinen, V.M.; Reunanen, J.; Satokari, R.; de Vos, W.M.; Palva, I.; Palva, A. Using recombinant lactococci as an approach to dissect the immunomodulating capacity of surface piliation in probiotic Lactobacillus rhamnosus GG. PLoS ONE 2013, 8, e64416. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, K.; Ueno, S.; Sugiyama, M.; Yamamoto, Y.; Mukai, T. Lactobacillus rhamnosus GG SpaC pilin subunit binds to the carbohydrate moieties of intestinal glycoconjugates. Anim. Sci. J. 2016, 87, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Lebeer, S.; Claes, I.; Tytgat, H.L.; Verhoeven, T.L.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; Vos, W.M.; Keersmaecker, S.C.; et al. Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ardita, C.S.; Mercante, J.W.; Kwon, Y.M.; Luo, L.; Crawford, M.E.; Powell, D.N.; Jones, R.M.; Neish, A.S. Epithelial adhesion mediated by pilin SpaC is required for Lactobacillus rhamnosus GG-induced cellular responses. Appl. Environ. Microbiol. 2014, 80, 5068–5077. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Beaussart, A.; Alsteens, D.; Dupres, V.; Claes, I.; von Ossowski, I.; de Vos, W.M.; Palva, A.; Lebeer, S.; Vanderleyden, J.; et al. Adhesion and nanomechanics of pili from the probiotic Lactobacillus rhamnosus GG. ACS Nano 2013, 7, 3685–3697. [Google Scholar] [CrossRef] [PubMed]
- Kant, A.; von Ossowski, I.; Palva, A.; Krishnan, V. Crystallization and X-ray crystallographic analysis of the adhesive SpaC pilin subunit in the SpaCBA pilus of gut-adapted Lactobacillus rhamnosus GG. Protein Pept. Lett. 2016, 23, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Dupres, V.; Beaussart, A.; Lebeer, S.; Claes, I.J.; Vanderleyden, J.; Dufrêne, Y.F. Deciphering the nanometer-scale organization and assembly of Lactobacillus rhamnosus GG pili using atomic force microscopy. Langmuir 2012, 28, 2211–2216. [Google Scholar] [CrossRef] [PubMed]
- Sullan, R.M.; Beaussart, A.; Tripathi, P.; Derclaye, S.; El-Kirat-Chatel, S.; Li, J.K.; Schneider, Y.J.; Vanderleyden, J.; Lebeer, S.; Dufrêne, Y.F. Single-cell force spectroscopy of pili-mediated adhesion. Nanoscale 2014, 6, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Von Ossowski, I.; Reunanen, J.; Satokari, R.; Vesterlund, S.; Kankainen, M.; Huhtinen, H.; Tynkkynen, S.; Salminen, S.; de Vos, W.M.; Palva, A. Mucosal adhesion properties of the probiotic Lactobacillus rhamnosus GG SpaCBA and SpaFED pilin subunits. Appl. Environ. Microbiol. 2010, 76, 2049–2057. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, H.L.; Douillard, F.P.; Reunanen, J.; Rasinkangas, P.; Hendrickx, A.P.; Laine, P.K.; Paulin, L.; Satokari, R.; de Vos, W.M. Lactobacillus rhamnosus GG outcompetes Enterococcus faecium via mucus-binding pili: Evidence for a novel and heterospecific probiotic mechanism. Appl. Environ. Microbiol. 2016, 82, 5756–5762. [Google Scholar] [CrossRef] [PubMed]
- Ganguli, K.; Collado, M.C.; Rautava, J.; Lu, L.; Satokari, R.; von Ossowski, I.; Reunanen, J.; de Vos, W.M.; Palva, A.; Isolauri, E.; et al. Lactobacillus rhamnosus GG and its SpaC pilus adhesin modulate inflammatory responsiveness and TLR-related gene expression in the fetal human gut. Pediatr. Res. 2015, 77, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Vargas García, C.E.; Petrova, M.; Claes, I.J.; De Boeck, I.; Verhoeven, T.L.; Dilissen, E.; von Ossowski, I.; Palva, A.; Bullens, D.M.; Vanderleyden, J.; et al. Piliation of Lactobacillus rhamnosus GG promotes adhesion, phagocytosis, and cytokine modulation in macrophages. Appl. Environ. Microbiol. 2015, 81, 2050–2062. [Google Scholar] [CrossRef] [PubMed]
- Tytgat, H.L.; van Teijlingen, N.H.; Sullan, R.M.; Douillard, F.P.; Rasinkangas, P.; Messing, M.; Reunanen, J.; Satokari, R.; Vanderleyden, J.; Dufrêne, Y.F.; et al. Probiotic gut microbiota isolate interacts with dendritic cells via glycosylated heterotrimeric pili. PLoS ONE 2016, 11, e0151824. [Google Scholar] [CrossRef] [PubMed]
- Cheesman, S.E.; Guillemin, K. We know you are in there: Conversing with the indigenous gut microbiota. Res. Microbiol. 2007, 158, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Medzhitov, R. Role of the innate immune system and host-commensal mutualism. Curr. Top. Microbiol. Immunol. 2006, 308, 1–18. [Google Scholar] [PubMed]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [PubMed]
- O’Connell Motherway, M.; Zomer, A.; Leahy, S.C.; Reunanen, J.; Bottacini, F.; Claesson, M.J.; O’Brien, F.; Flynn, K.; Casey, P.G.; Munoz, J.A.; et al. Functional genome analysis of Bifidobacterium breve UCC2003 reveals type IVb tight adherence (Tad) pili as an essential and conserved host-colonization factor. Proc. Natl. Acad. Sci. USA 2011, 108, 11217–11222. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, P.; von Ossowski, I.; Palva, A.; Krishnan, V. Purification, crystallization and preliminary X-ray diffraction analysis of SpaD, a backbone-pilin subunit encoded by the fimbrial spaFED operon in Lactobacillus rhamnosus GG. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Megta, A.K.; Palva, A.; von Ossowski, I.; Krishnan, V. Crystallization and X-ray diffraction analysis of SpaE, a basal pilus protein from the gut-adapted Lactobacillus rhamnosus GG. Acta Crystallogr. F Struct. Biol. Commun. 2017, 73, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Neville, B.A.; Forde, B.M.; Claesson, M.J.; Darby, T.; Coghlan, A.; Nally, K.; Ross, R.P.; O’Toole, P.W. Characterization of pro-inflammatory flagellin proteins produced by Lactobacillus ruminis and related motile lactobacilli. PLoS ONE 2012, 7, e40592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Donnell, M.M.; Harris, H.M.; Lynch, D.B.; Ross, R.P.; O’Toole, P.W. Lactobacillus ruminis strains cluster according to their mammalian gut source. BMC Microbiol. 2015, 15, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvetti, E.; Torriani, S.; Felis, G.E. The genus Lactobacillus: A taxonomic update. Probiotics Antimicrob. Proteins 2012, 4, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, M.E.; Latham, M.J.; Garvie, E.I.; Zirngibl, J.; Kandler, O. Two new species of Lactobacillus isolated from the bovine rumen, Lactobacillus ruminis sp.nov. And Lactobacillus vitulinus sp.nov. J. Gen. Microbiol. 1973, 77, 37–49. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, J.; O’Toole, P.W. Lactobacillus: Host-microbe relationships. Curr. Top. Microbiol. Immunol. 2013, 358, 119–154. [Google Scholar] [PubMed]
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar] [PubMed]
- Kant, R.; Palva, A.; von Ossowski, I. An in silico pan-genomic probe for the molecular traits behind Lactobacillus ruminis gut autochthony. PLoS ONE 2017, 12, e0175541. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Åvall-Jääskeläinen, S.; Koort, J.; Lindholm, A.; Rintahaka, J.; von Ossowski, I.; Palva, A.; Hynönen, U. A comparative characterization of different host-sourced Lactobacillus ruminis strains and their adhesive, inhibitory, and immunomodulating functions. Front. Microbiol. 2017, 8, 657. [Google Scholar] [CrossRef] [PubMed]
- Manetti, A.G.O.; Spadafina, T. The role of pili in the formation of biofilm and bacterial communities. In Bacterial Pili: Structure, Synthesis and Role in Disease, 1st ed.; Barocchi, M.A., Telford, J.L., Eds.; CAB International: Wallingford, UK, 2013; Volume 27, pp. 151–164. [Google Scholar]
- Rosini, R.; Margarit, I. Biofilm formation by Streptococcus agalactiae: Influence of environmental conditions and implicated virulence factors. Front. Cell. Infect. Microbiol. 2015, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Taweechotipatr, M.; Iyer, C.; Spinler, J.K.; Versalovic, J.; Tumwasorn, S. Lactobacillus saerimneri and Lactobacillus ruminis: Novel human-derived probiotic strains with immunomodulatory activities. FEMS Microbiol. Lett. 2009, 293, 65–72. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Genetics and mechanism behind the pilin assembly and cell wall anchoring of sortase-dependent pili. (A) Depicted is a schematic representation of a characteristic sortase-dependent pilus operon, which includes the genes for the tip, basal, and backbone pilin subunits and the pilus-specific C-type sortase enzyme. The housekeeping A-type sortase gene is invariably found elsewhere along the genome. Examples of the various pilin gene products are illustrated, and for each of these pilins, the location of the N-terminally located pilin motif (VYPK) and the LPXTG motif at the C-terminus are indicated correspondingly. Sortase gene products are shown with the conserved catalytic motif (TLXTC) in the vicinity of the active-site cleft. Key residues of each amino acid motif are highlighted by color. (B) Schematically depicted from left to right are the sequential steps leading up to the assembly and anchoring of a typical three-subunit sortase-dependent pilus. See (A) for the identities of the various pilins and sortases. (Refer to main text for details).
Figure 1.
Genetics and mechanism behind the pilin assembly and cell wall anchoring of sortase-dependent pili. (A) Depicted is a schematic representation of a characteristic sortase-dependent pilus operon, which includes the genes for the tip, basal, and backbone pilin subunits and the pilus-specific C-type sortase enzyme. The housekeeping A-type sortase gene is invariably found elsewhere along the genome. Examples of the various pilin gene products are illustrated, and for each of these pilins, the location of the N-terminally located pilin motif (VYPK) and the LPXTG motif at the C-terminus are indicated correspondingly. Sortase gene products are shown with the conserved catalytic motif (TLXTC) in the vicinity of the active-site cleft. Key residues of each amino acid motif are highlighted by color. (B) Schematically depicted from left to right are the sequential steps leading up to the assembly and anchoring of a typical three-subunit sortase-dependent pilus. See (A) for the identities of the various pilins and sortases. (Refer to main text for details).
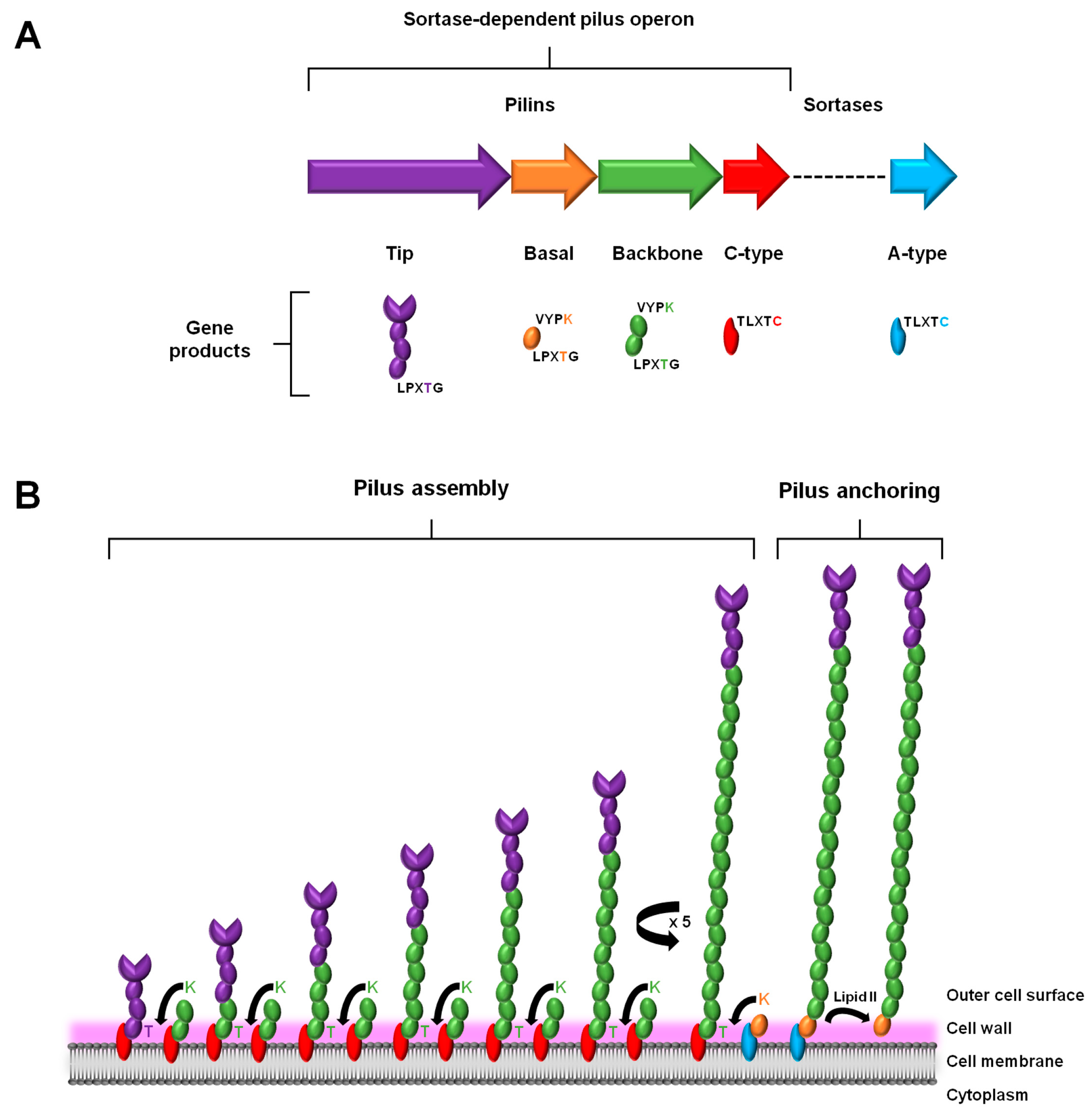
Figure 2.
Three-dimensional view of the tip, backbone, and basal pilin structures within a typical sortase-dependent pilus. Shown are representative tertiary structures of the three types of pilin subunits. The structure of the tip pilin is comprised of a stalk-like region consisting of three Ig-fold domains (e.g., CnaA or CnaB), upon which rests a globular head region that contains the substrate-binding domain. The C-terminal LPXTG peptide is normally situated within the bottom subdomain. The backbone pilin structure is shown having two Ig-fold domains, with the linking lysine and the LPXTG peptide located within the upper and lower domains, respectively. The basal pilin structure is depicted with a single Ig-fold domain that includes a proline-rich (blue) extended C-terminal tail. All domain regions are highlighted by circular background shading. Structural visualization of the tip, backbone, and basal pilins was rendered by PyMol (http://www.pymol.org/) using protein atomic coordinates (2WW8, 3B2M, and 3LKQ, respectively) retrieved from the RCSB Protein Data Bank (http:// http://www.rcsb.org/pdb/home/home.do).
Figure 2.
Three-dimensional view of the tip, backbone, and basal pilin structures within a typical sortase-dependent pilus. Shown are representative tertiary structures of the three types of pilin subunits. The structure of the tip pilin is comprised of a stalk-like region consisting of three Ig-fold domains (e.g., CnaA or CnaB), upon which rests a globular head region that contains the substrate-binding domain. The C-terminal LPXTG peptide is normally situated within the bottom subdomain. The backbone pilin structure is shown having two Ig-fold domains, with the linking lysine and the LPXTG peptide located within the upper and lower domains, respectively. The basal pilin structure is depicted with a single Ig-fold domain that includes a proline-rich (blue) extended C-terminal tail. All domain regions are highlighted by circular background shading. Structural visualization of the tip, backbone, and basal pilins was rendered by PyMol (http://www.pymol.org/) using protein atomic coordinates (2WW8, 3B2M, and 3LKQ, respectively) retrieved from the RCSB Protein Data Bank (http:// http://www.rcsb.org/pdb/home/home.do).

Figure 3.
Visualization of native sortase-dependent piliation in lactobacilli by immuno-electron microscopy. Immunogold pilin-protein labeling and electron microscopy (EM) of bacterial cells were done using established techniques [89]. Long protruding structures representing the SpaCBA pili in L. rhamnosus GG (A) and LrpCBA pili in L. ruminis ATCC 25644 (B) are identified by immuno-labeling first with polyclonal antibody specific for the backbone-pilin subunit (SpaA and LrpA, respectively) and then protein A-conjugated gold particles (10 nm in diameter; black dots), followed by negative staining and electron microscopy (EM). Scale bars for each electron micrograph image are found at the bottom-left corner.
Figure 3.
Visualization of native sortase-dependent piliation in lactobacilli by immuno-electron microscopy. Immunogold pilin-protein labeling and electron microscopy (EM) of bacterial cells were done using established techniques [89]. Long protruding structures representing the SpaCBA pili in L. rhamnosus GG (A) and LrpCBA pili in L. ruminis ATCC 25644 (B) are identified by immuno-labeling first with polyclonal antibody specific for the backbone-pilin subunit (SpaA and LrpA, respectively) and then protein A-conjugated gold particles (10 nm in diameter; black dots), followed by negative staining and electron microscopy (EM). Scale bars for each electron micrograph image are found at the bottom-left corner.
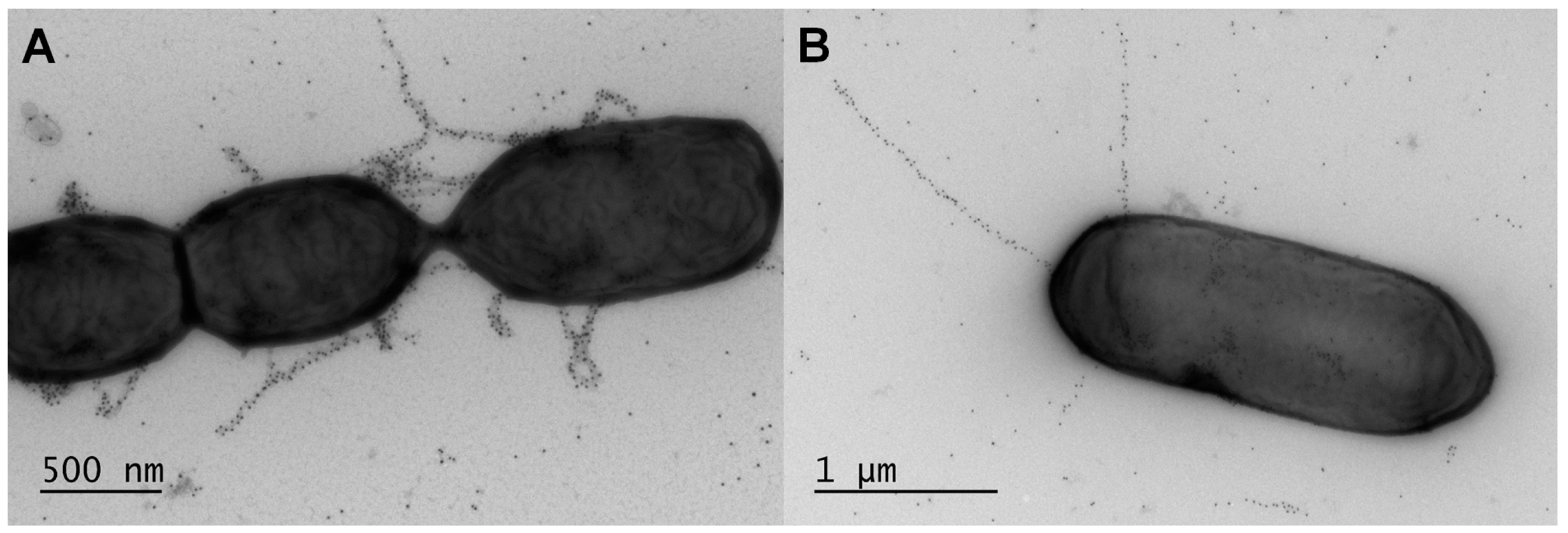
Figure 4.
Schematic depiction showing the presumptive binding of piliated lactobacilli to the intestinal mucosa epithelium. Based on substrate-binding capacities, L. rhamnosus cells (purple) with their mucoadhesive SpaCBA pili would be predominantly found within the outer and inner mucus layers, whereas flagellated L. ruminis cells (blue) would be able to maneuver through this mucosal barrier and then, via their ECM-binding LrpCBA pili, attach themselves to the single cell layer of intestinal epithelial tissue. This cartoon image is representative only, and includes various aspects and features that are not drawn to scale.
Figure 4.
Schematic depiction showing the presumptive binding of piliated lactobacilli to the intestinal mucosa epithelium. Based on substrate-binding capacities, L. rhamnosus cells (purple) with their mucoadhesive SpaCBA pili would be predominantly found within the outer and inner mucus layers, whereas flagellated L. ruminis cells (blue) would be able to maneuver through this mucosal barrier and then, via their ECM-binding LrpCBA pili, attach themselves to the single cell layer of intestinal epithelial tissue. This cartoon image is representative only, and includes various aspects and features that are not drawn to scale.


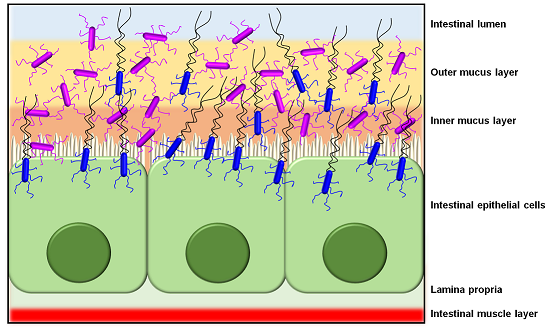{kind=link}
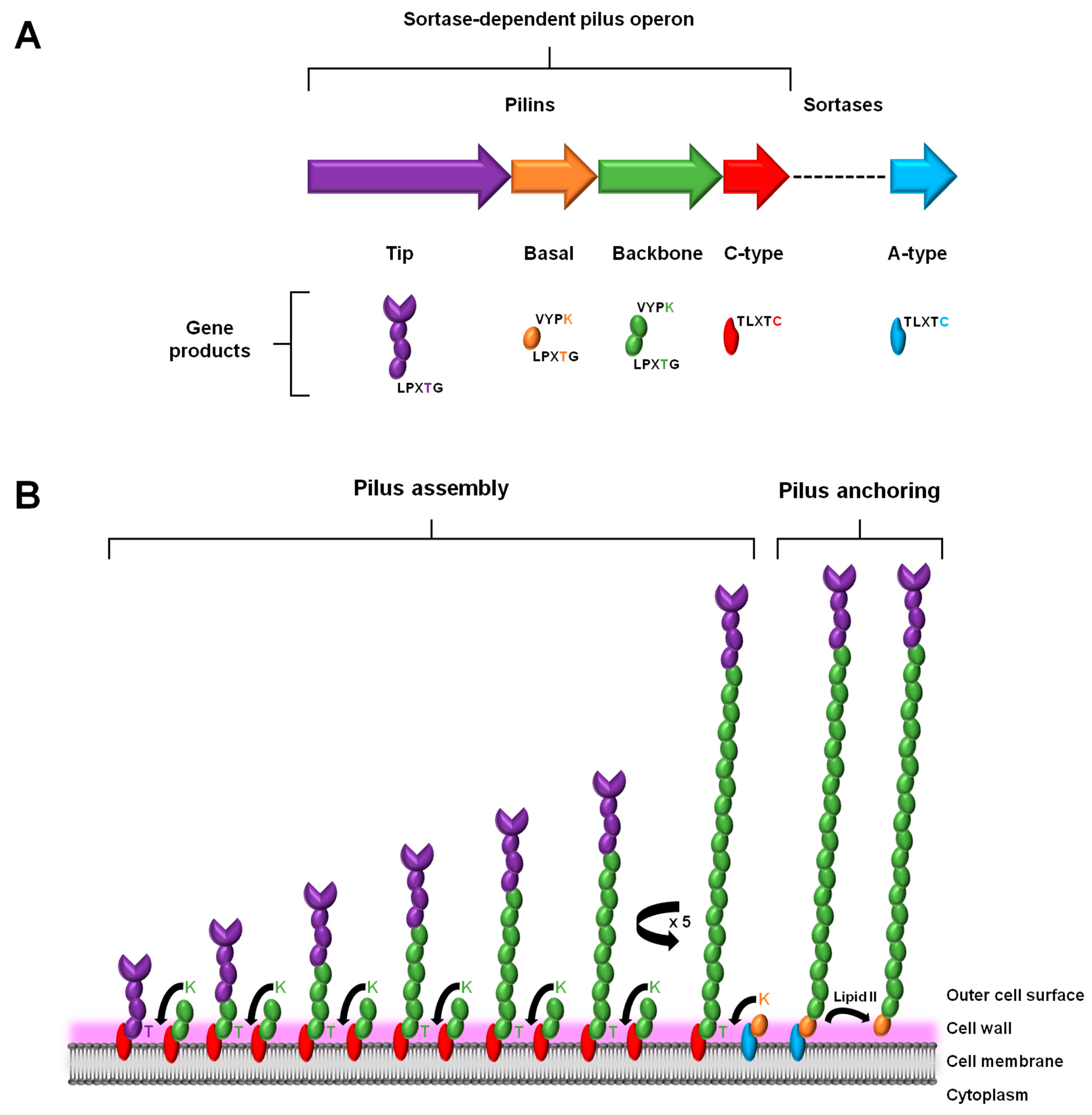{kind=link}
{kind=link}
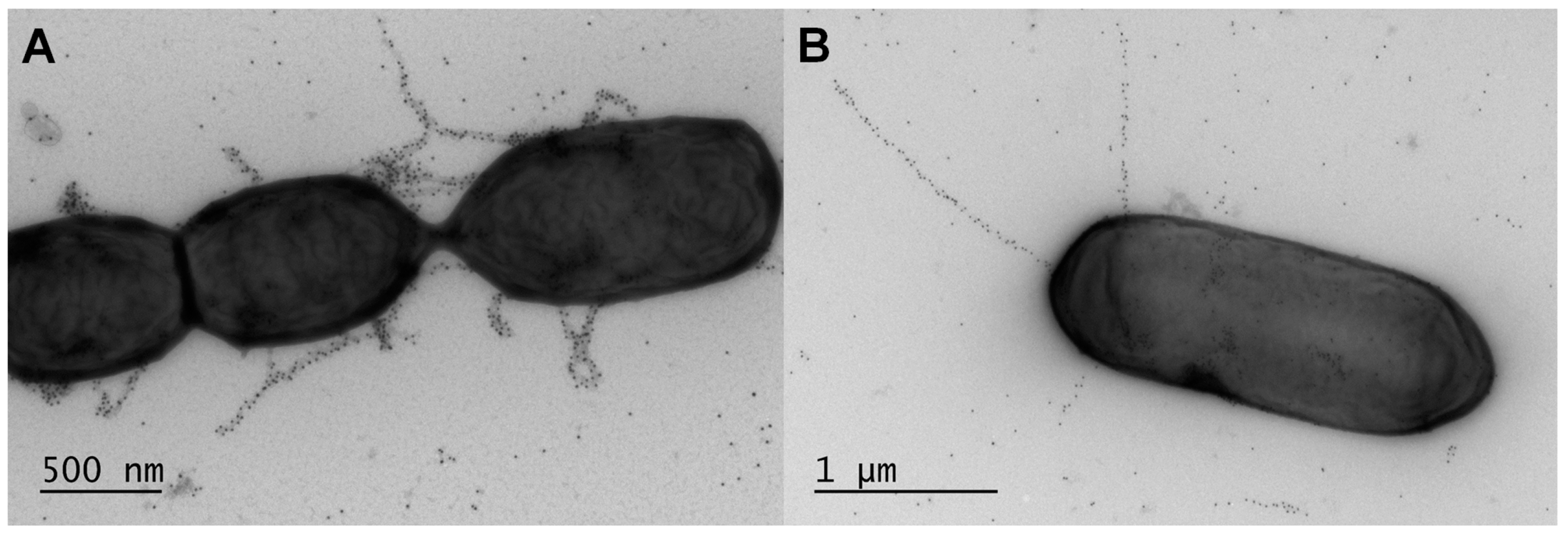{kind=link}
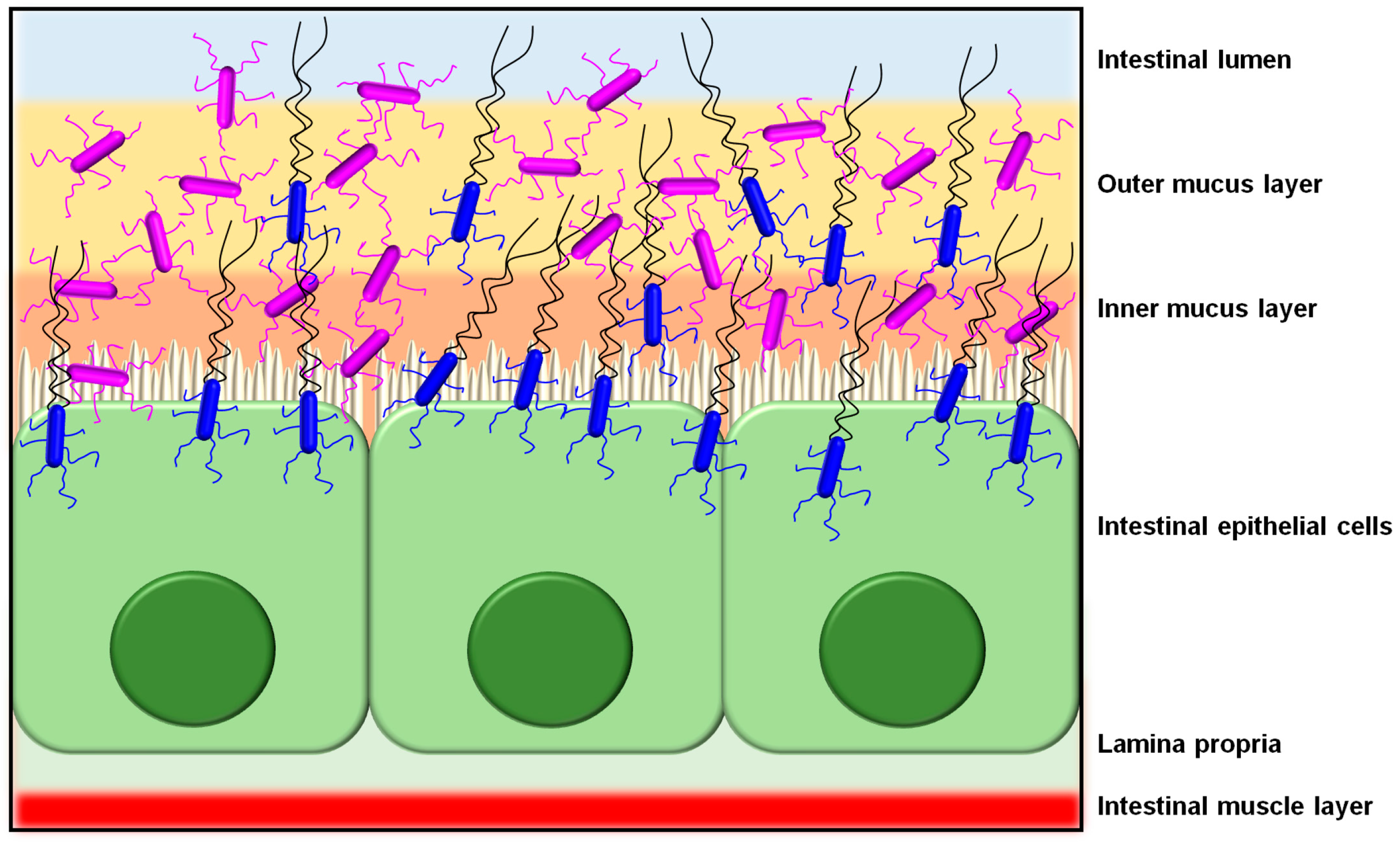{kind=link}
Table 1.
Overview of conserved amino acid sequence motifs and domains in the lactobacillar SpaCBA, SpaFED, and LrpCBA pilin subunits. *
Table 1.
Overview of conserved amino acid sequence motifs and domains in the lactobacillar SpaCBA, SpaFED, and LrpCBA pilin subunits. *
| Primary Structure Feature | Tip Pilin Subunit |
|---|---|
| N-terminal secretory domain | MTAKVARTGHLFAVLLILMSMLTGLVTSGSSVVT (SpaC) LPRKWIHMLMLLLMLVTQIGSA (SpaF) MERNKIFKKLLCILGAVATVFAIVFAMGKFDGEKANA (LrpC) |
| C-terminal LPXTG domain | LPHTGGQGYQRLLGIALGLISAAFLLLLVVLIKRRVVKQHD (SpaC) LPKTGGSGILLFLMVAISACGGGWLLYLYLKRKEAR (SpaF) LPQTGGPGRLLFEALGSLLIVVACALTEVLIWRRIRSSKGV (LrpC) |
| Pilin motif | not detected (SpaC) not detected (SpaF) not detected (LrpC) |
| E-box | YGIQEAAAPTGY and YTMSETKAPDGY (SpaC) YRLTETKAPAGF (SpaF) YKLVETRTQSGY (LrpC) |
| Primary Structure Feature | Basal Pilin Subunit |
| N-terminal secretory domain | MTKSFRPLVILTFCLALLVSLATTTLQQTQA (SpaB) MRRFYWWLVPLLLLIGIVLGNTPHWVHA (SpaE) MKRVLKLLFMIVAFMTAVFAGSGQASA (LrpB) |
| C-terminal LPXTG domain | LPQTGDTVAAWLSVLGLIIFATVLAFNIKNQKINKWER (SpaB) LPAMSDWRNLRFVLLGSLLLLLATYFFIKNKKARHHACK (SpaE) LPQTGEAKSIMALLGIGIICLVVLVSVGRRNYKEEH (LrpB) |
| Pilin motif | TADFWQLVSKN (SpaB) PLQTIHLYPKN (SpaE) FPLGGQSYAKN (LrpB) |
| E-box | YLFKETAAPKNI (SpaB) YFFEELQGVPGY (SpaE) YYFSEVQAPKGY (LrpB) |
| Primary Structure Feature | Backbone Pilin Subunit |
| N-terminal secretory domain | MKKTIAKKVLTLTSTILMTLLMVLGFNGTRVQA (SpaA) MQVTFKKIGHSLLAALMLMSFLLPLLSAGKPVHA (SpaD) MKNHKKLRNALATLLLALPLALQGAVGVKTAQA (LrpA) |
| C-terminal LPXTG domain | LPHTGGTGTVIFAILGVALIAFGAVAYRKRRNGF (SpaA) LPMTGGIGLFAFLMIGAILMGGGHLMKKKTSKKV (SpaD) LPSTGGMGIVLFIAAGVVVMAGAAGTMIVRRNRRENI (LrpA) |
| Pilin motif | ADGNVYVYPKN (SpaA) DLTNIHLYPKD (SpaD) VQKSINIYPKN (LrpA) |
| E-box | YLFHETNPRAGY and YTAVETNVPDGY (SpaA) YAFHEAVTPQPY and YTLVETAAPEGY (SpaD) YLFAETDAPANI and YAVKEVKAPTGY (LrpA) |
* Sequence motifs and domains for the pilin subunits are based on the deduced primary structure from the pilus operon genes in the L. rhamnosus GG (spaCBA and spaFED) and L. ruminis ATCC 25644 (lrpCBA) strains.
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Von Ossowski, I. Novel Molecular Insights about Lactobacillar Sortase-Dependent Piliation. Int. J. Mol. Sci. 2017, 18, 1551. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071551
AMA Style
Von Ossowski I. Novel Molecular Insights about Lactobacillar Sortase-Dependent Piliation. International Journal of Molecular Sciences. 2017; 18(7):1551. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071551
Chicago/Turabian StyleVon Ossowski, Ingemar. 2017. "Novel Molecular Insights about Lactobacillar Sortase-Dependent Piliation" International Journal of Molecular Sciences 18, no. 7: 1551. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071551
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.