Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii

Abstract
:1. Introduction
2. Results
2.1. The Yield of Extracted Glucans from Seven Pleurotus Strains
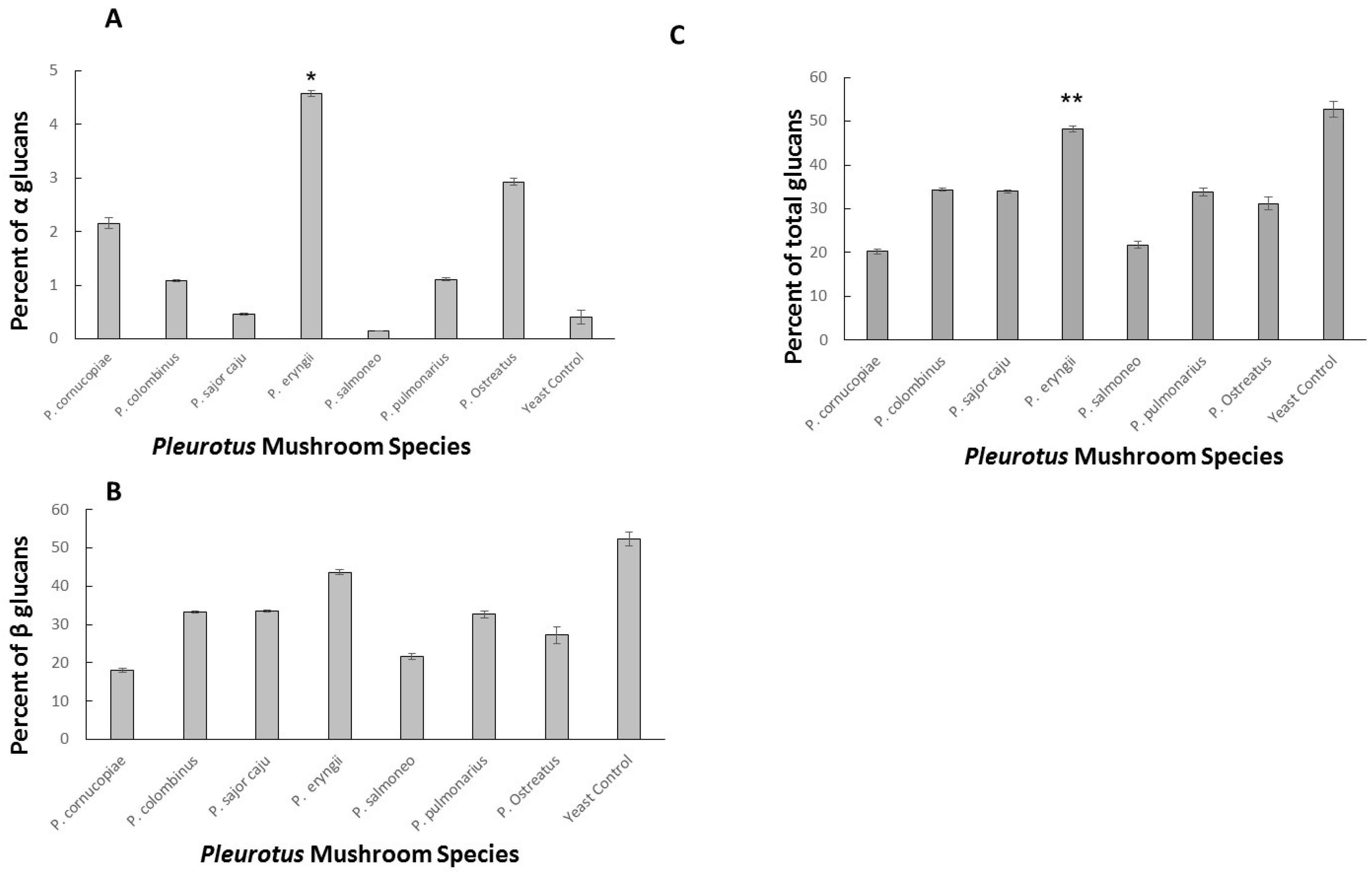
2.2. α-Glucan and Total Glucan Content among Seven Pleurotus Strains (Five Species)
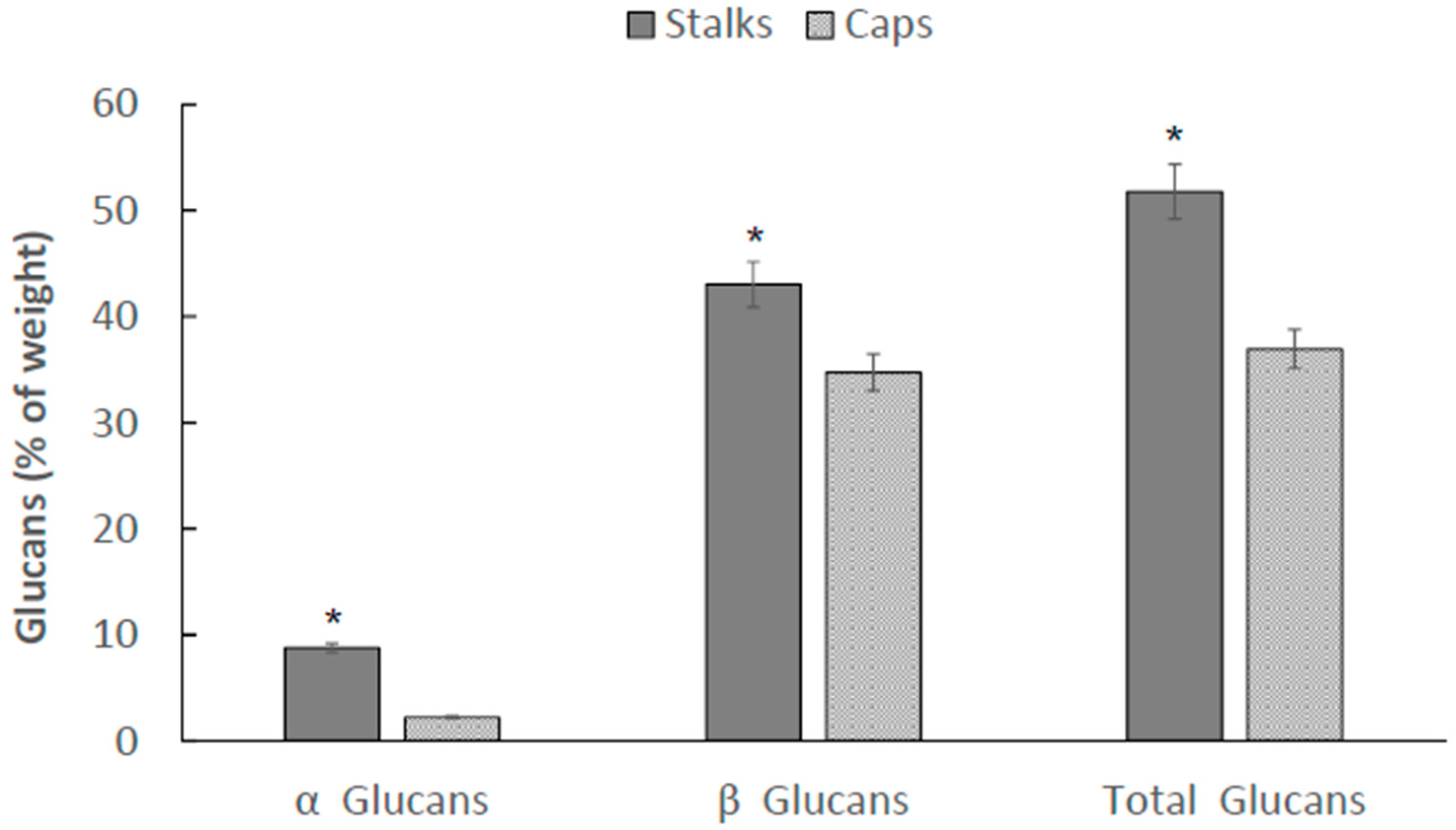
2.3. Total, α-Glucan, and β-Glucan Distribution in P. eryngii between Caps (C) and Stalks (S)
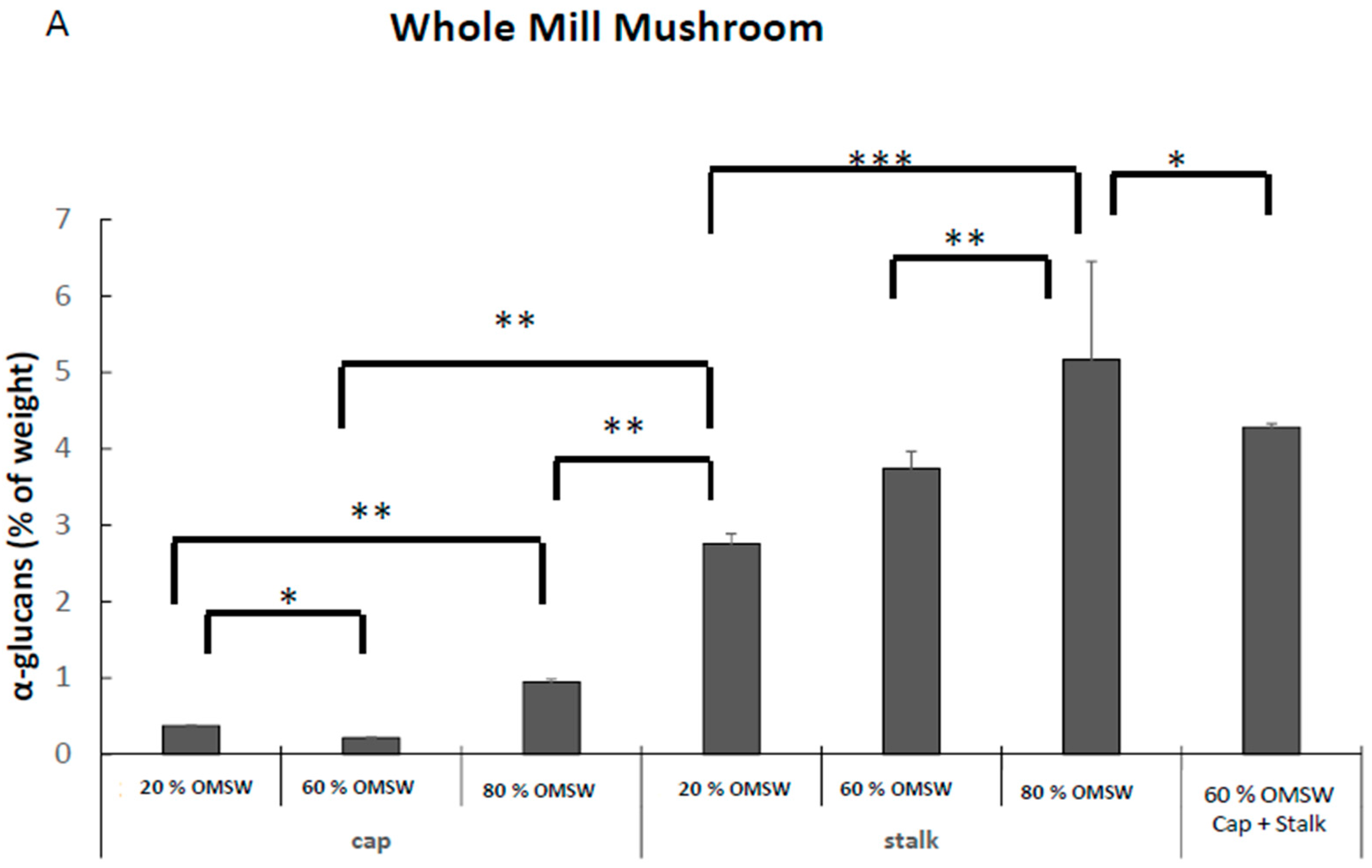
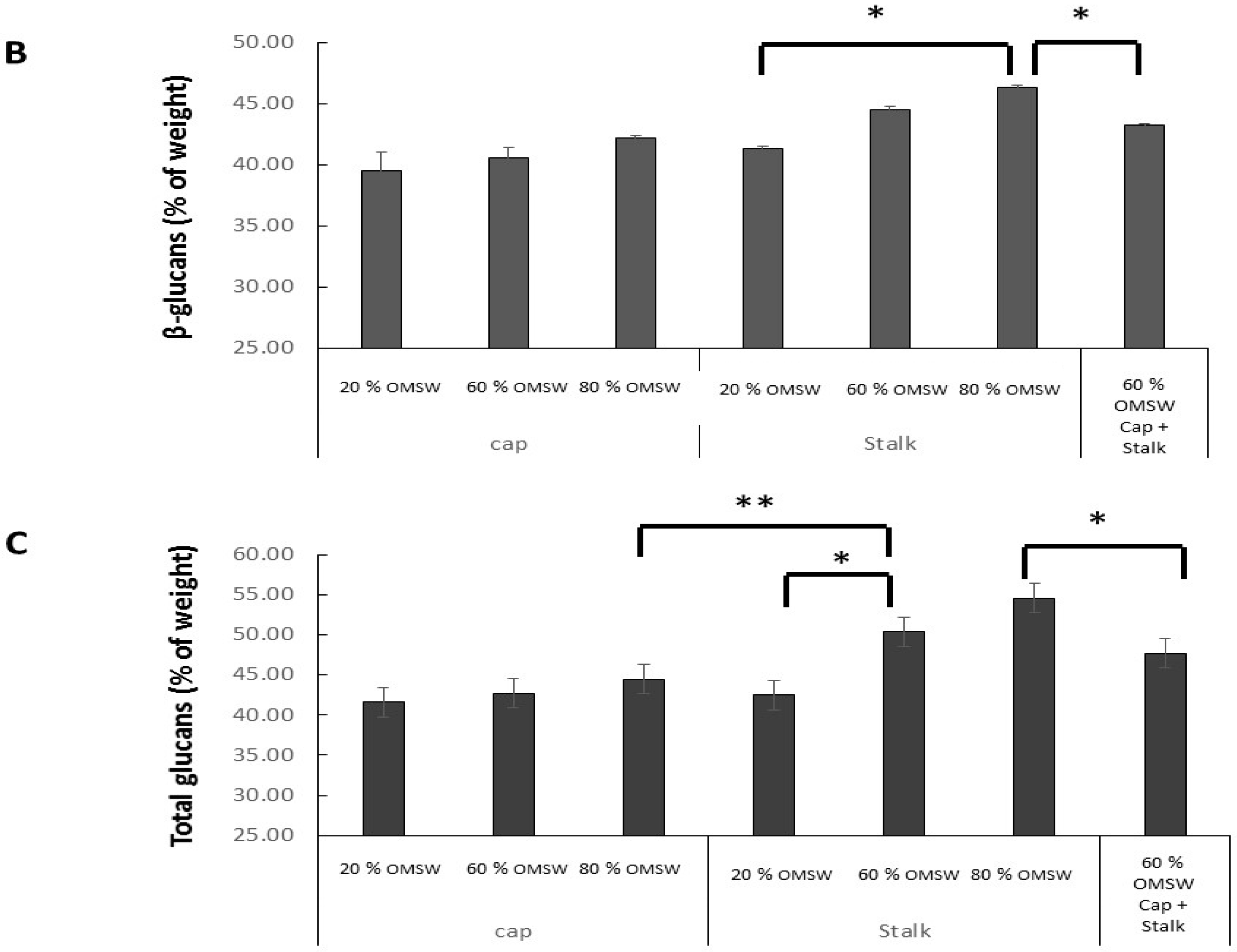
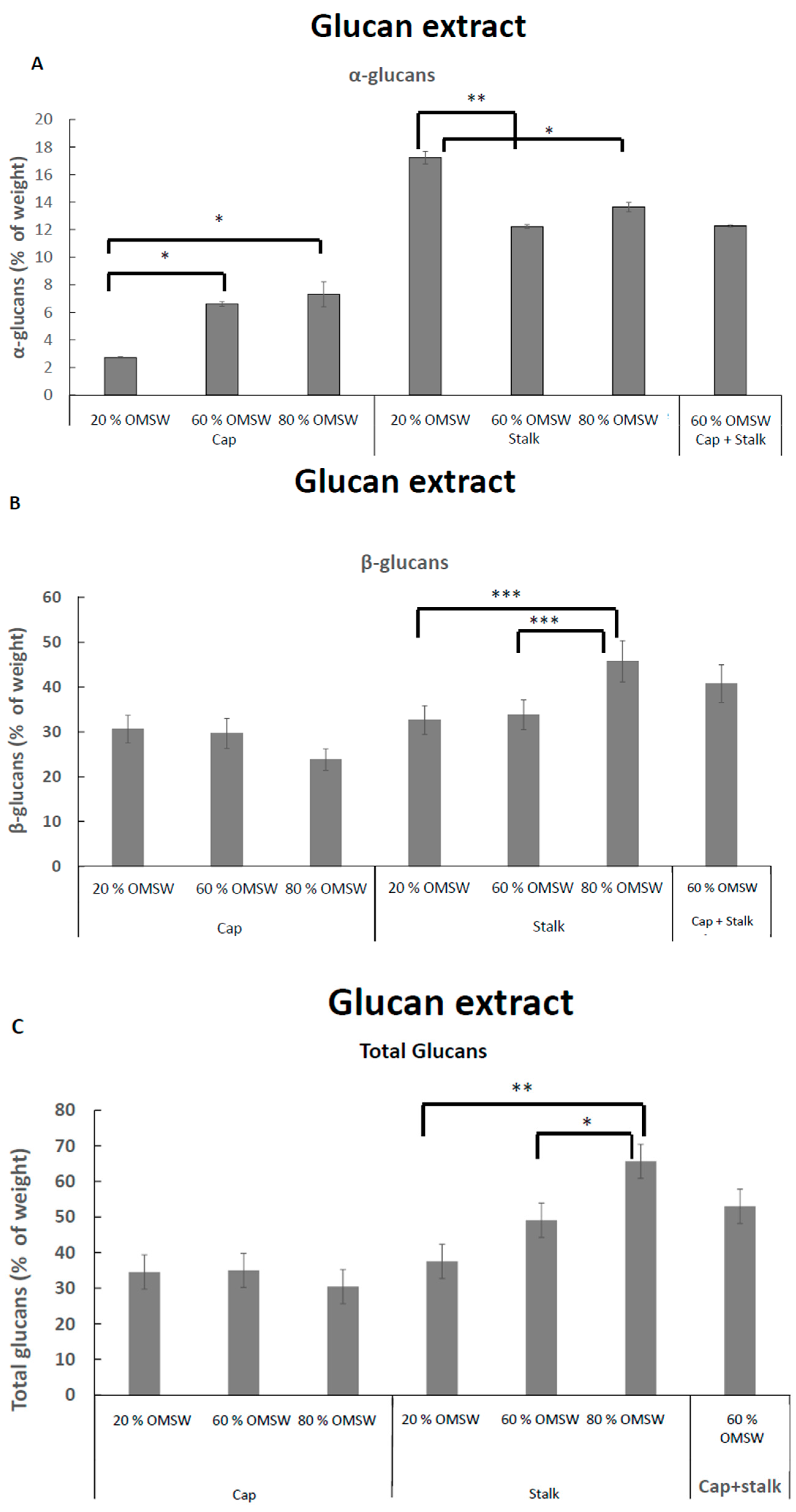
2.4. The Effect of Cultivation of Pleurotus Eryngii in Various Relative Amounts of Eucalyptus Sawdust and Olive Mill Solid Waste (OMSW) on Concentrations of Total Glucans, α-Glucans, and β-Glucans
3. Discussion
4. Materials and Methods
4.1. Mushroom Growth Conditions
4.2. Mushroom Sample Collection and Preparation
4.3. Preparation and Extraction of Glucans from Different Powdered Dried Mushroom Strains
4.4. Glucans Analysis
4.5. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhang, Y.; Li, S.; Wang, X.; Zhang, L.; Cheung, P.C. Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocoll. 2011, 25, 196–206. [Google Scholar] [CrossRef]
- Carrasco-Gonzalez, J.A.; Serna-Saldívar, S.O.; Gutierrez-Uribe, J.A. Mycochemical changes induced by selenium enrichment in P. ostreatus fruiting bodies. J. Agric. Food Chem. 2017, 65, 4074–4082. [Google Scholar] [CrossRef] [PubMed]
- Wasser, S.P. Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl. Microbiol. Biotechnol. 2011, 89, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.J.; Lin, C.P.; Tsai, S.Y. Vitamin D 2 content and antioxidant properties of fruit body and mycelia of edible mushrooms by UV-B irradiation. J. Food Compos. Anal. 2015, 42, 38–45. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Stathopoulos, P.; Kaliora, A.C.; Zervakis, G.I. Bioactive compounds and antioxidant activity exhibit high intraspecific variability in Pleurotus ostreatus mushrooms and correlate well with cultivation performance parameters. World J. Microbiol. Biotechnol. 2017, 33, 98. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Novak, M. Structural analysis of glucans. Ann. Transl. Med. 2014, 2, 17. [Google Scholar] [PubMed]
- Rop, O.; Mlcek, J.; Jurikova, T. B-glucans in higher fungi and their health effects. Nutr. Rev. 2009, 67, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Sobieralski, K.; Siwulski, M.; Lisiecka, J.; Jędryczka, M.; Sas-Golak, I.; Frużyńska-Jóźwiak, D. Fungi-derived β-glucans as a component of functional food. Acta Sci. Pol. Hortorum Cult. 2012, 11, 111–128. [Google Scholar]
- Wiater, A.; Paduch, R.; Pleszczyńska, M.; Próchniak, K.; Choma, A.; Kandefer-Szerszeń, M.; Szczodrak, J. α-(1→3)-d-Glucans from fruiting bodies of selected macromycetes fungi and the biological activity of their carboxymethylated products. Biotechnol. Lett. 2011, 33, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Shida, M.; Uchida, T.; Matsuda, K. A (1/ar3)-α-d-glucan isolated from the fruit bodies of Lentinus edodes. Carbohydr. Res. 1978, 60, 117–127. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spěváček, J.; Erban, V.; Kovárˇíková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Synytsya, A.; Míčková, K.; Jablonsky, I.; Siuková, M.; Copikova, J. Mushrooms of genus Pleurotus as a source of dietary fibers and glucans for food supplements. Czech J. Food Sci. 2008, 26, 441–446. [Google Scholar]
- Chan, G.C.F.; Chan, W.K.; Sze, D.M.Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [PubMed]
- Zhang, M.; Cui, S.W.; Cheung, P.C.K.; Wang, Q. Antitumor polysaccharides from mushrooms: A review on their isolation process, structural characteristics and antitumor activity. Trends Food Sci. Technol. 2007, 18, 4–19. [Google Scholar] [CrossRef]
- Ishibashi, K.I.; Miura, N.N.; Adachi, Y.; Naohito, O.H.N.O.; Yadomae, T. Relationship between solubility of grifolan, a fungal 1, 3-β-d-glucan, and production of tumor necrosis factor by macrophages in vitro. Biosci. Biotechnol. Biochem. 2001, 65, 1993–2000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Cheung, P.C. Evaluation of sulfated Lentinus edodes α-(1→3)-d-glucan as a potential antitumor agent. Biosci. Biotechnol. Biochem. 2002, 66, 1052–1056. [Google Scholar] [CrossRef] [PubMed]
- Lavi, I.; Friesem, D.; Geresh, S.; Hadar, Y.; Schwartz, B. An aqueous polysaccharide extract from the edible mushroom Pleurotus ostreatus induces anti-proliferative and pro-apoptotic effects on HT-29 colon cancer cells. Cancer Lett. 2006, 244, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Lavi, I.; Levinson, D.; Peri, I.; Nimri, L.; Hadar, Y.; Schwartz, B. Orally administered glucans from the edible mushroom Pleurotus pulmonarius reduce acute inflammation in dextran sulfate sodium-induced experimental colitis. Br. J. Nutr. 2010, 103, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Lavi, I.; Nimri, L.; Levinson, D.; Peri, I.; Hadar, Y.; Schwartz, B. Glucans from the edible mushroom Pleurotus pulmonarius inhibit colitis-associated colon carcinogenesis in mice. J. Gastroenterol. 2012, 47, 504–518. [Google Scholar] [CrossRef] [PubMed]
- Descroix, K.; Větvička, V.; Laurent, I.; Jamois, F.; Yvin, J.C.; Ferrières, V. New oligo-β-(1,3)-glucan derivatives as immunostimulating agents. Bioorg. Med. Chem. 2010, 18, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Shabtay, A.; Hadar, Y.; Eitam, H.; Brosh, A.; Orlov, A.; Tadmor, Y.; Izhaki, I.; Kerem, Z. The potential of Pleurotus-treated olive mill solid waste as cattle feed. Bioresour. Technol. 2009, 100, 6457–6464. [Google Scholar] [CrossRef] [PubMed]
- Koutrotsios, G.; Larou, E.; Mountzouris, K.C.; Zervakis, G.I. Detoxification of olive mill wastewater and bioconversion of olive crop residues into high-value-added biomass by the choice edible mushroom hericium erinaceus. Appl. Biochem. Biotechnol. 2016, 180, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Koutrotsios, G.; Mountzouris, K.C.; Chatzipavlidis, I.; Zervakis, G.I. Bioconversion of lignocellulosic residues by Agrocybe cylindracea and Pleurotus ostreatus mushroom fungi—Assessment of their effect on the final product and spent substrate properties. Food Chem. 2014, 161, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Zervakis, G.I.; Koutrotsios, G.; Katsaris, P. Composted versus raw olive mill waste as substrates for the production of medicinal mushrooms: An assessment of selected cultivation and quality parameters. Biomed. Res. Int. 2013, 2013, 546830. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Rodriguez, A.; Soler-Rivas, C.; Polonia, I.; Wichers, H.J. Effect of olive mill waste (OMW) supplementation to Oyster mushrooms substrates on the cultivation parameters and fruiting bodies quality. Int. Biodeterior. Biodegrad. 2010, 64, 538–645. [Google Scholar] [CrossRef] [Green Version]
- Lavi, I.; Levinson, D.; Peri, I.; Tekoah, Y.; Hadar, Y.; Schwartz, B. Chemical characterization, antiproliferative and antiadhesive properties of polysaccharides extracted from Pleurotus pulmonarius mycelium and fruiting bodies. Appl. Microbiol. Biotechnol. 2010, 85, 1977–1990. [Google Scholar] [CrossRef] [PubMed]
- Megazyme, Mushroom and Yeast Β-Glucan Assay Procedure. 2014. Available online: www.megazyme.com (accessed on 18 July 2017).
- McCleary, B.V.; Draga, A. Measurement of β-glucan in mushrooms and mycelial products. J. AOAC Int. 2016, 99, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Reverberi, M.; di Mario, F.; Tomati, U. β-Glucan synthase induction in mushrooms grown on olive mill wastewaters. Appl. Microbiol. Biotechnol. 2004, 66, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Hernandez, M.; Kramer, J.K.; Rinker, D.L.; Tsao, R. Ergosterol profiles, fatty acid composition, and antioxidant activities of button mushrooms as affected by tissue part and developmental stage. J. Agric. Food Chem. 2010, 58, 11616–11625. [Google Scholar] [CrossRef] [PubMed]
- Manzi, P.; Aguzzi, A.; Pizzoferrato, L. Nutritional value of mushrooms widely consumed in Italy. Food Chem. 2001, 73, 321–325. [Google Scholar] [CrossRef]
- Manzi, P.; Pizzoferrato, L. β-Glucans in edible mushrooms. Food Chem. 2000, 68, 315–318. [Google Scholar] [CrossRef]
- Dawoud, M.E.A. Participation of different γ irradiation techniques, doses and radioprotective compounds upon carbon metabolism, polypeptide upregulation and subsequent 1,3-β-glucans over-production by Pleurotus ostreatus mushrooms. J. Food Agric. Environ. 2012, 10, 235–244. [Google Scholar]
- Elisashvili, V.I.; Kachlishvili, E.T.; Wasser, S.P. Carbon and nitrogen source effects on basidiomycetes exopolysaccharide production. Appl. Biochem. Microbiol. 2009, 45, 531–535. [Google Scholar] [CrossRef]
- Smits, G.J.; van den Ende, H.; Klis, F.M. Differential regulation of cell wall biogenesis during growth and development in yeast. Microbiology 2001, 147, 781–794. [Google Scholar] [CrossRef] [PubMed]
- De Nobel, H.; Ruiz, C.; Martin, H.; Morris, W.; Brul, S.; Molina, M.; Klis, F.M. Cell wall perturbation in yeast results in dual phosphorylation of the Slt2/Mpk1 MAP kinase and in an Slt2-mediated increase in FKS2–lacZ expression, glucanase resistance and thermotolerance. Microbiology 2000, 146, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Hong, L.; Xun, M.; Wutong, W. Anti-diabetic effect of an α-glucan from fruit body of maitake (Grifola frondosa) on KK-Ay mice. J. Pharm. Pharmacol. 2007, 59, 575–582. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mushroom Species | Total Dried Weight (g) | Glucans Extracted (g) | Yield % |
|---|---|---|---|
| Pleurotus ostreatus | 66 | 2.64 | 4 |
| Pleurotus ostreatus var. colombinus | 58 | 1.74 | 3 |
| Pleurotus pulmonarius | 70 | 2.80 | 4 |
| Pleurotus sajor caju | 74 | 1.48 | 2 |
| Pleurotus eryngii | 55 | 3.30 | 6 |
| Pleurotus cornucopiae | 64 | 1.92 | 3 |
| Pleurotus salmoneostramineus | 86 | 2.58 | 3 |
| Treatment (% OMSW) | Yield (%) |
|---|---|
| Control (100% eucalyptus sawdust) | 6.2 ± 0.39 |
| 20 | 12.56 ± 1.39 |
| 60 | 24.6 ± 4.23 |
| 80 | 29.24 ± 6.65 |
| Mushroom Name | Commercial Strain |
|---|---|
| Pleurotus ostreatus | Fungisem K-12 |
| Pleurotus ostreatus var. colombinus | Sylvan 3030 |
| Pleurotus pulmonarius | Sylvan 3014 |
| Pleurotus pulmonarius var. sajor caju | Jamaica 503 |
| Pleurotus eryngii | Mycelia 2600 |
| Pleurotus cornucopiae | Sylvan 3040 |
| Pleurotus salmoneostramineus | Mycelia 2708 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avni, S.; Ezove, N.; Hanani, H.; Yadid, I.; Karpovsky, M.; Hayby, H.; Gover, O.; Hadar, Y.; Schwartz, B.; Danay, O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. Int. J. Mol. Sci. 2017, 18, 1564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071564
Avni S, Ezove N, Hanani H, Yadid I, Karpovsky M, Hayby H, Gover O, Hadar Y, Schwartz B, Danay O. Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii. International Journal of Molecular Sciences. 2017; 18(7):1564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071564
Chicago/Turabian StyleAvni, Sharon, Nirit Ezove, Hilla Hanani, Itamar Yadid, Michal Karpovsky, Hilla Hayby, Ofer Gover, Yitzhak Hadar, Betty Schwartz, and Ofer Danay. 2017. "Olive Mill Waste Enhances α-Glucan Content in the Edible Mushroom Pleurotus eryngii" International Journal of Molecular Sciences 18, no. 7: 1564. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071564