Unravelling the Role of Metallothionein on Development, Reproduction and Detoxification in the Wall Lizard Podarcis sicula


Abstract
:1. Introduction
2. P. sicula Metallothionein
3. Metallothionein in P. sicula Adult Tissues
4. Metallothionein in P. sicula Reproductive Organs
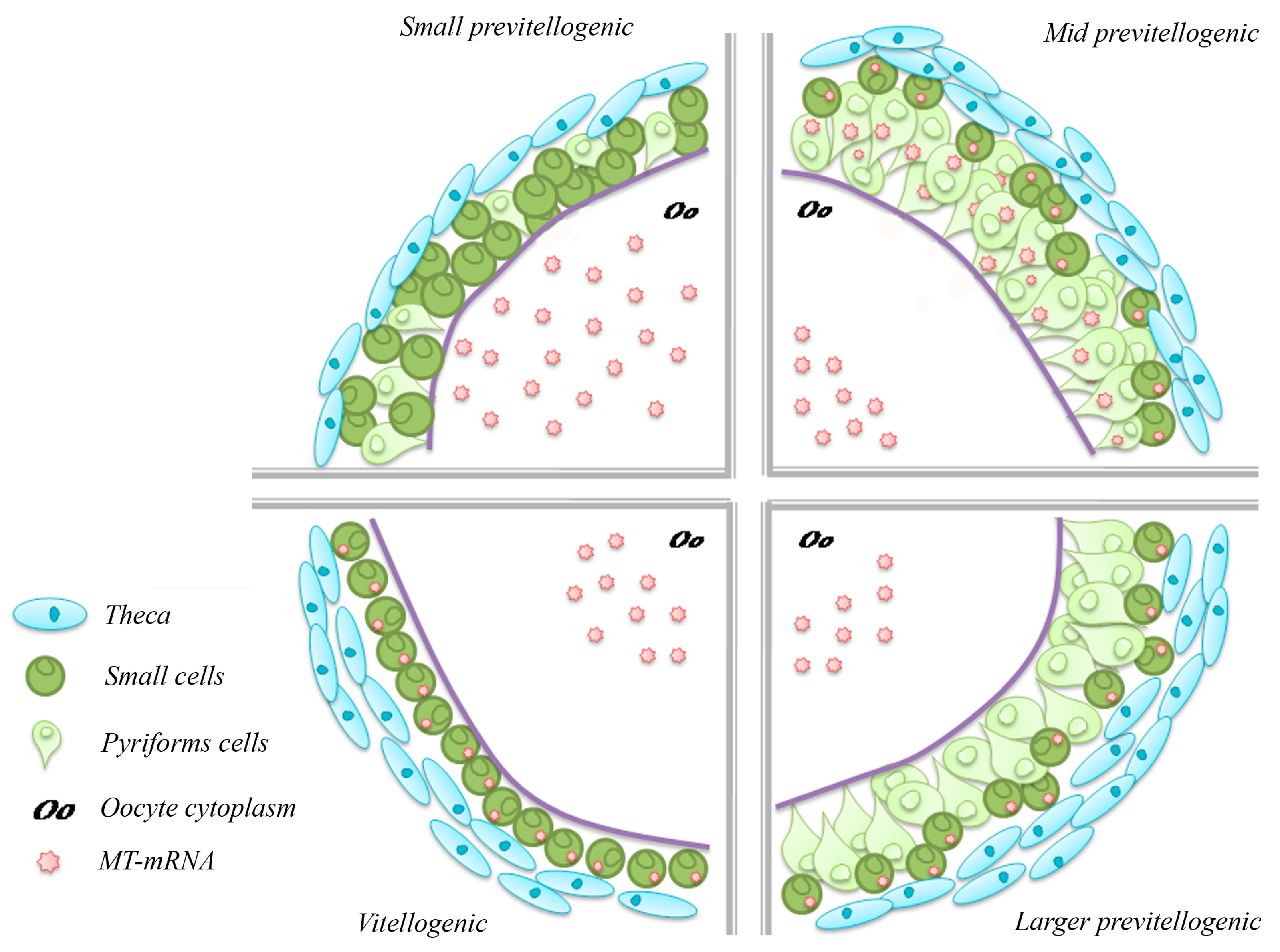
4.1. Metallothionein in the Ovary
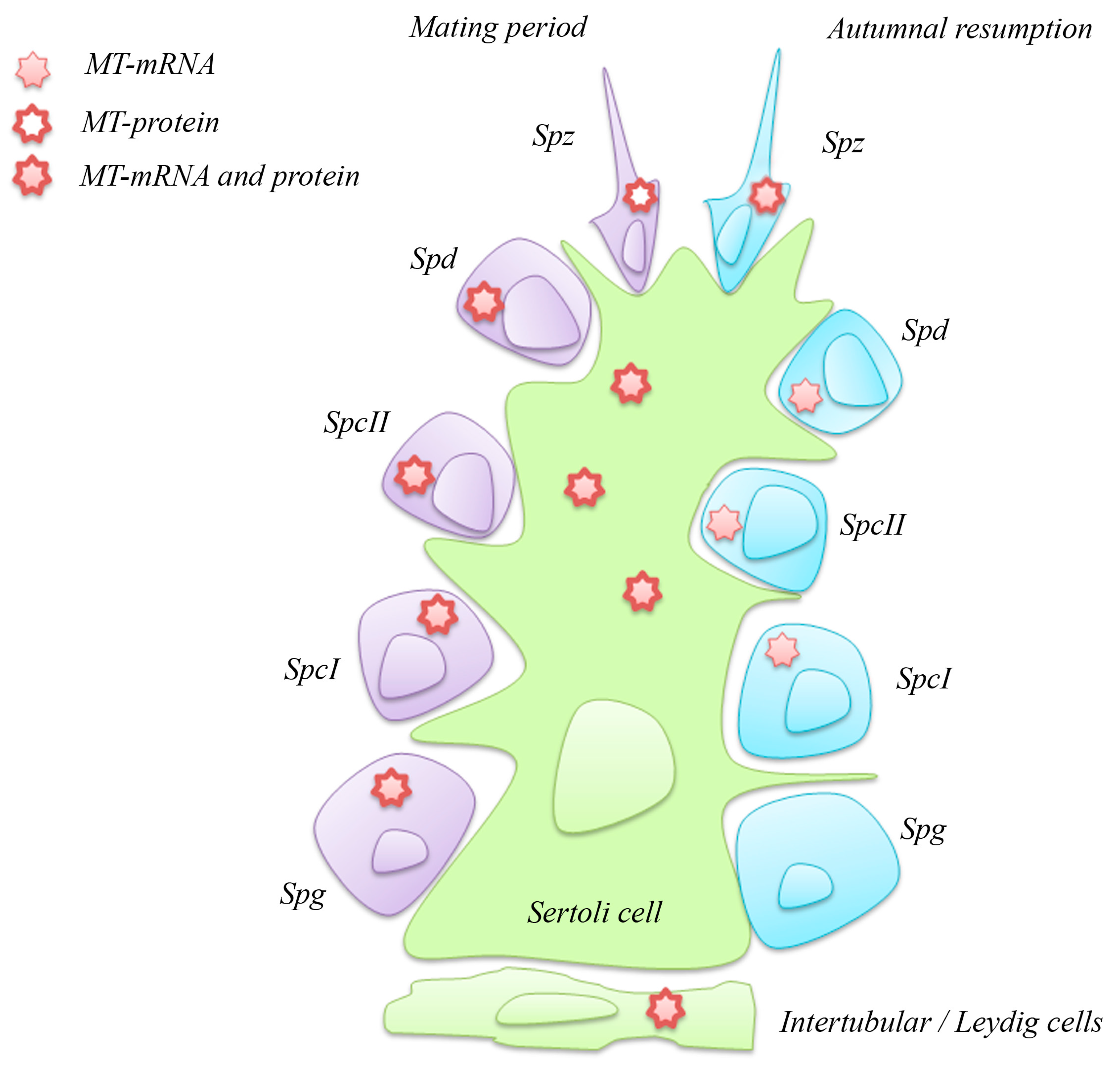
4.2. Metallothionein in the Testis
5. Metallothionein in P. sicula Development
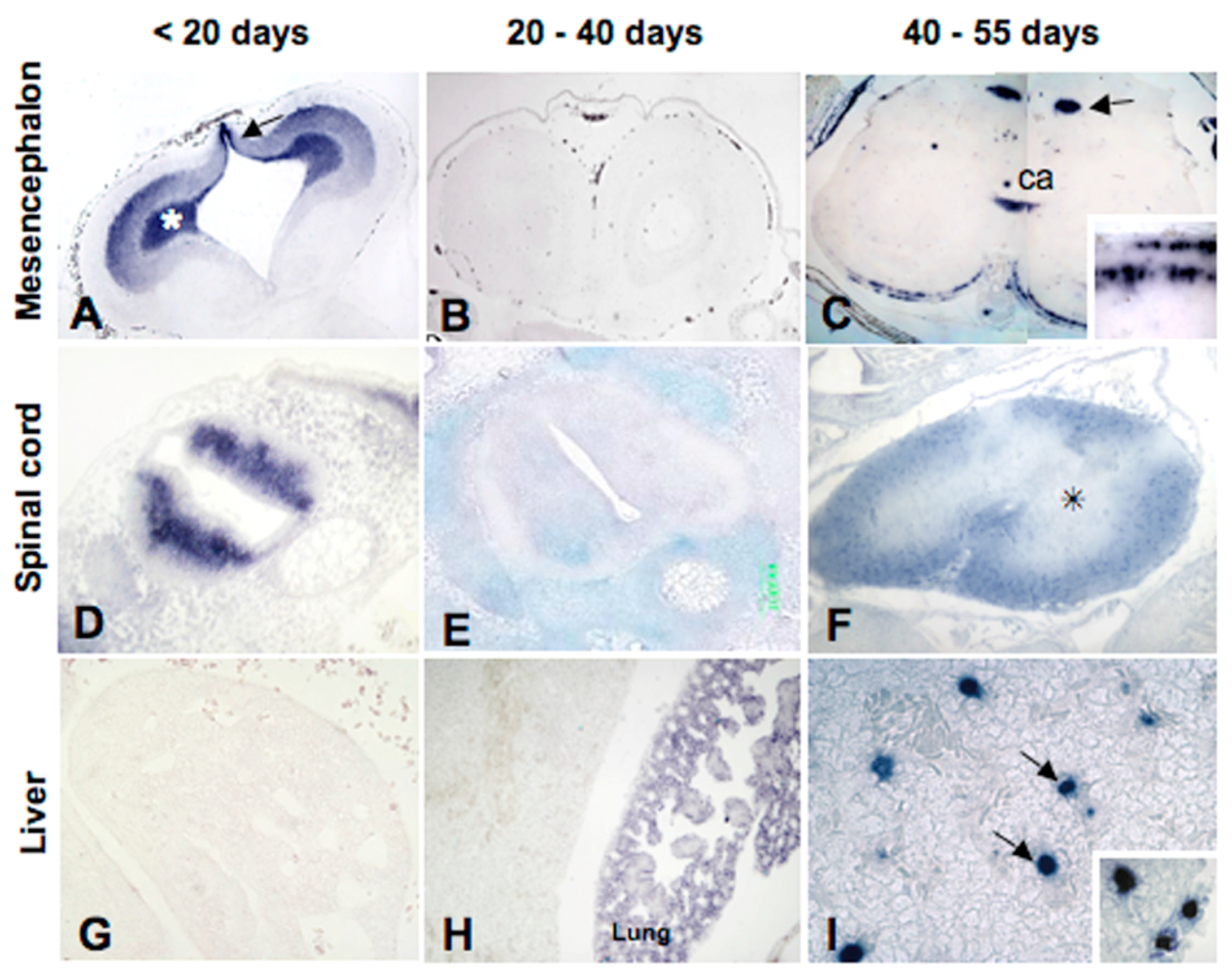
5.1. Metallothionein Expression in P. sicula Embryos under Natural Conditions
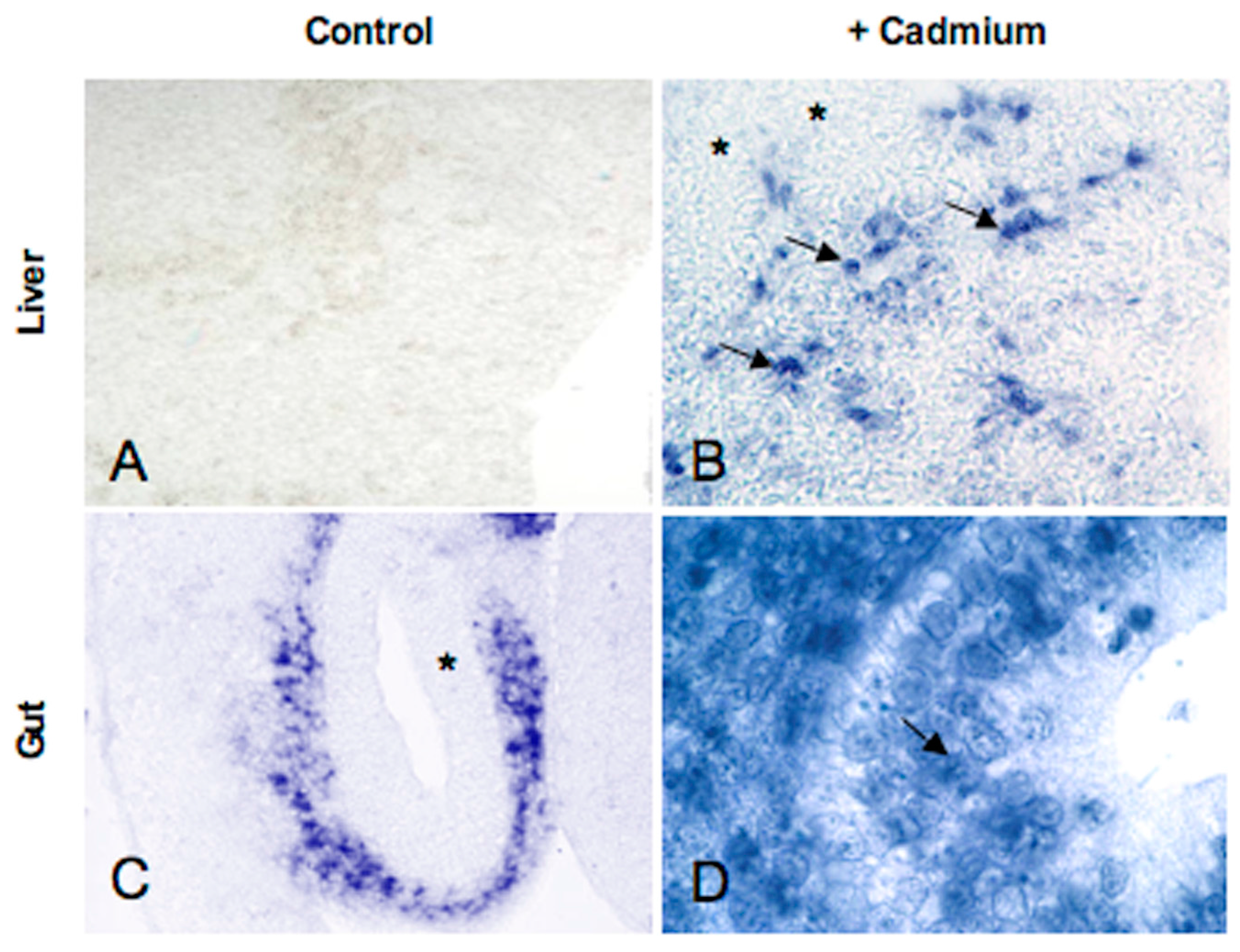
5.2. Metallothionein Expression in P. sicula Embryos Incubated in Cadmium-Contaminated Soil
6. Conclusions
Conflicts of Interest
References
- Margoshes, M.; Vallee, B.L. A cadmium protein from equine kidney cortex. J. Am. Chem. Soc. 1957, 79, 4813–4814. [Google Scholar] [CrossRef]
- Bremner, I. Interactions between metallothionein and trace elements. Prog. Food Nutr. Sci. 1987, 11, 1–37. [Google Scholar] [PubMed]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef] [PubMed]
- Palmiter, R.D. The elusive function of metallothioneins. Proc. Natl. Acad. Sci. USA 1998, 95, 8428–8430. [Google Scholar] [CrossRef] [PubMed]
- Hamer, D.H. Metallothionein. Annu. Rev. Biochem. 1986, 55, 913–951. [Google Scholar] [CrossRef] [PubMed]
- Isani, G.; Carpenè, E. Metallothioneins, unconventional proteins from unconventional animals: A long journey from nematodes to mammals. Biomolecules 2014, 4, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Baurand, P.E.; Pedrini-Martha, V.; de Vaufleury, A.; Niederwanger, M.; Capelli, N.; Scheifler, R.; Dallinger, R. Differential expression of metallothionein isoforms in terrestrial snail embryos reflects early life stage adaptation to metal stress. PLoS ONE 2015, 10, e0116004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacios, O.; Pérez-Rafael, S.; Pagani, A.; Dallinger, R.; Atrian, S.; Capdevila, M. Cognate and noncognate metal ion coordination in metal-specific metallothioneins: The Helix pomatia system as a model. J. Biol. Inorg. Chem. 2014, 19, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Höckner, M.; Dallinger, R.; Stürzenbaum, S.R. Nematode and snail metallothioneins. J. Biol. Inorg. Chem. 2011, 16, 1057–1065. [Google Scholar] [CrossRef] [PubMed]
- Palacios, O.; Pagani, A.; Pérez-Rafael, S.; Egg, M.; Höckner, M.; Brandstätter, A.; Capdevila, M.; Atrian, S.; Dallinger, R. Shaping mechanisms of metal specificity in a family of metazoan metallothioneins: Evolutionary differentiation of mollusc metallothioneins. BMC Biol. 2011, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherland, D.E.; Stillman, M.J. The “magic numbers” of metallothionein. Metallomics 2011, 3, 444–463. [Google Scholar] [CrossRef] [PubMed]
- Hogstrand, C.; Haux, C. Binding and detoxification of heavy metals in lower vertebrates with reference to metallothionein. Comp. Biochem. Physiol. C 1991, 100, 137–141. [Google Scholar] [CrossRef]
- Uchida, Y.; Takio, K.; Titani, K.; Ihara, Y.; Tomonaga, M. The growth inhibitory factor that is deficient in the Alzheimer’s disease brain is a 68 amino acid metallothionein-like protein. Neuron 1991, 7, 337–347. [Google Scholar] [CrossRef]
- Quaife, C.J.; Findley, S.D.; Erickson, J.C.; Froelick, G.J.; Kelly, E.J.; Zambrowicz, B.P.; Palmiter, R.D. Induction of a new metallothionein isoform (MT-IV) occurs during differentiation of stratified squamous epithelia. Biochemistry 1994, 33, 7250–7259. [Google Scholar] [CrossRef] [PubMed]
- Moleirinho, A.; Carneiro, J.; Matthiesen, R.; Silva, R.M.; Amorim, A.; Azevedo, L. Gains, losses and changes of function after gene duplication: Study of the metallothionein family. PLoS ONE 2011, 6, e18487. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, R.; Filosa, S.; Trinchella, F. Metallothionein gene evolution in vertebrates: Events of gene duplication and loss during squamates diversification. In Advances in Medicine and Biology; Berhardt, L.V., Ed.; Nova Science Publishers: Happauge, NY, USA, 2011; Volume 24, pp. 321–335. [Google Scholar]
- Serén, N.; Glaberman, S.; Carretero, M.A.; Chiari, Y. Molecular evolution and functional divergence of the metallothionein gene family in vertebrates. J. Mol. Evol. 2014, 78, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, R. Unexpected diversity of metallothionein primary structure in Amphibians: Evolutionary implications for vertebrate metallothioneins. In Amphibians: Anatomy, Ecological Significance and Conservation Strategies; Lombardi, M.P., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2014; pp. 27–38. ISBN 9781633214347. [Google Scholar]
- Scudiero, R.; Tussellino, M.; Carotenuto, R. Identification and expression of an atypical isoform of metallothionein in the African clawed frog Xenopus laevis. C. R. Biol. 2015, 338, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Trinchella, F.; Riggio, M.; Filosa, S.; Parisi, E.; Scudiero, R. Molecular cloning and sequencing of metallothionein in squamates: New insights into the evolution of the metallothionein genes in vertebrates. Gene 2008, 423, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Trinchella, F.; Esposito, M.G.; Scudiero, R. Metallothionein primary structure in amphibians: Insights from comparative evolutionary analysis in vertebrates. C. R. Biol. 2012, 335, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Riggio, M.; Trinchella, F.; Parisi, E.; Filosa, S.; Scudiero, R. Accumulation of zinc, copper and metallothionein mRNA in lizard ovary proceeds without a concomitant increase in metallothionein content. Mol. Reprod. Dev. 2003, 66, 347–382. [Google Scholar] [CrossRef] [PubMed]
- Trinchella, F.; Riggio, M.; Filosa, S.; Volpe, M.G.; Parisi, E.; Scudiero, R. Cadmium distribution and metallothionein expression in lizard tissues following acute and chronic cadmium intoxication. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Scudiero, R.; Filosa, S.; Motta, C.M.; Simoniello, P.; Trinchella, F. Cadmium in the wall lizard Podarcis Sicula: Morphological and molecular effects on embryonic and adult tissues. In Reptiles: Biology, Behavior and Conservation; Baker, K.J., Ed.; Nova Science Publishers: Happauge, NY, USA, 2011; pp. 147–162. ISBN 978-1-61324-740-2. [Google Scholar]
- Verderame, M.; Limatola, E.; Scudiero, R. The terrestrial lizard Podarcis sicula as experimental model in emerging pollutants evaluation. In Ecotoxicology and Genotoxicology: Non-Traditional Terrestrial Models; Larramendy, M., Ed.; Royal Society of Chemistry (RSC) Publishing: London, UK, 2017; Chapter 12; ISBN 978-1-78262-811-8. [Google Scholar]
- Simoniello, P.; Filosa, S.; Riggio, M.; Scudiero, R.; Tammaro, S.; Trinchella, F.; Motta, C.M. Responses to cadmium intoxication in the liver of the wall lizard Podarcis Sicula. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 194–203. [Google Scholar] [CrossRef] [PubMed]
- Simoniello, P.; Filosa, S.; Scudiero, R.; Trinchella, F.; Motta, C.M. Cadmium impairment of reproduction in the female wall lizard Podarcis sicula. Environ. Toxicol. 2013, 28, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Carnevali, O.; Mosconi, G.; Angelini, F.; Limatola, E.; Ciarcia, G.; Polzonetti-Magni, A. Plasma vitellogenin and 17β-estradiol levels during the annual reproductive cycle of Podarcis s. sicula Raf. Gen. Comp. Endocrinol. 1991, 84, 337–343. [Google Scholar] [CrossRef]
- Borrelli, L.; de Stasio, R.; Motta, C.M.; Parisi, E.; Filosa, S. Seasonal-dependent effect of temperature on the response of adenylate cyclase to FSH stimulation in the oviparous lizard, Podarcis sicula. J. Endocrinol. 2000, 167, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Motta, C.M.; Castriota Scanderbeg, M.; Filosa, S.; Andreuccetti, P. Role of pyriform cells during the growth of oocytes in the lizard Podarcis sicula. J. Exp. Zool. A Ecol. Genet. Physiol. 1995, 273, 247–256. [Google Scholar] [CrossRef]
- Simoniello, P.; Motta, C.M.; Scudiero, R.; Trinchella, F.; Filosa, S. Spatiotemporal changes in metallothionein gene expression during embryogenesis in the wall lizard Podarcis sicula. J. Exp. Zool. 2010, 313A, 410–420. [Google Scholar] [CrossRef] [PubMed]
- De Moor, C.H.; Richter, J.D. Translational control in vertebrate development. Int. Rev. Cytol. 2001, 203, 567–608. [Google Scholar] [CrossRef] [PubMed]
- Pedrini-Martha, V.; Schnegg, R.; Baurand, P.E.; de Vaufleury, A.; Dallinger, R. The physiological role and toxicological significance of the non-metal-selective cadmium/copper-metallothionein isoform differ between embryonic and adult helicid snails. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, M.P.; Chernoff, S.B.; Klaassen, C.D. Cadmium-binding proteins of rat testes. Characterization of a low-molecular-mass protein that lacks identity with metallothionein. Biochem. J 1984, 220, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Waalkes, M.P.; Perantoni, A.; Palmer, A.E. Isolation and partial characterization of the low-molecular-mass zinc/cadmium-binding protein from the testes of the patas monkey (Erythrocebus patas). Distinction from metallothionein. Biochem. J. 1988, 256, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Nishimura, N.; Tohyama, C. Localization of metallothionein in the genital organs of the male rat. J. Histochem. Cytochem. 1990, 38, 927–933. [Google Scholar] [CrossRef] [PubMed]
- De, S.K.; Enders, G.C.; Andrews, G.K. High levels of metallothionein messenger RNAs in male germ cells of the adult mouse. Mol. Endocrinol. 1991, 5, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Tohyama, C.; Nishimura, N.; Suzuki, J.S.; Karasawa, M.; Nishimura, H. Metallothionein mRNA in the testis and prostate of the rat detected by digoxigenin-labeled riboprobe. Histochemistry 1994, 101, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, J.S.; Kodama, N.; Molotkov, A.; Aoki, E.; Tohyama, C. Isolation and identification of metallothionein isoforms (MT-1 and MT-2) in the rat testis. Biochem. J. 1998, 334, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.L.; Sheng, Z.; Liu, G.D.; Long, L.L.; Wang, Y.F.; Yang, W.X.; Zhu, J.Q. Cloning, characterization and cadmium inducibility of metallothionein in the testes of the mudskipper Boleophthalmus pectinirostris. Ecotoxicol. Environ. Saf. 2015, 119, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Durnam, D.M.; Palmiter, R.D. Transcriptional regulation of the mouse metallothionein-I gene by heavy metals. J. Biol. Chem. 1981, 256, 5712–5716. [Google Scholar] [PubMed]
- Rosati, L.; Prisco, M.; Coraggio, F.; Valiante, S.; Scudiero, R.; Laforgia, V.; Andreuccetti, P.; Agnese, M. PACAP and PAC1 receptor in the reproductive cycle of male lizard Podarcis sicula. Gen. Comp. Endocrinol. 2014, 205, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Verderame, M.; Angelini, F.; Limatola, E. Spermatogenic waves and expression of AR and ERs in germ cells of Podarcis sicula. Int. J. Zool. 2014, 2014. [Google Scholar] [CrossRef]
- Verderame, M.; Limatola, E.; Scudiero, R. Metallothionein expression and synthesis in the testis of the lizard Podarcis sicula under natural conditions and following estrogenic exposure. Eur. J. Histochem. 2017, 61, 90–95. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [PubMed]
- Sørensen, M.B.; Stoltenberg, M.; Henriksén, K.; Ernst, E.; Danscher, G.; Parvinen, M. Histochemical tracing of zinc ions in the rat testis. Mol. Hum. Reprod. 1998, 4, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Cano-Gauci, D.F.; Sarkar, B. Reversible zinc exchange between metallothionein and the estrogen receptor zinc finger. FEBS Lett. 1996, 386, 1–4. [Google Scholar] [CrossRef]
- Kelly, E.J.; Quaife, C.J.; Froelick, G.J.; Palmiter, R.D. Metallothionein I and II protect against zinc deficiency and zinc toxicity in mice. J. Nutr. 1996, 126, 1782–1790. [Google Scholar] [PubMed]
- Andrews, G.K.; Huet-Hudson, Y.M.; Paria, B.C.; McMaster, M.T.; De, S.K.; Dey, S.K. Metallothionein gene expression and metal regulation during preimplantation mouse embryo development (MT mRNA during early development). Dev. Biol. 1991, 145, 13–27. [Google Scholar] [CrossRef]
- Andersen, R.D.; Piletz, J.E.; Birren, B.W.; Herschman, H.R. Levels of metallothionein messenger RNA in foetal, neonatal and maternal rat liver. Eur. J. Biochem. 1983, 131, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Andrews, G.K.; Lee, D.K.; Ravindra, R.; Lichtlen, P.; Sirito, M.; Sawadogo, M.; Schaffner, W. The transcription factors MTF-1 and USF1 cooperate to regulate mouse metallothionein-I expression in response to the essential metal zinc in visceral endoderm cells during early development. EMBO J. 2001, 20, 1114–1122. [Google Scholar] [CrossRef] [PubMed]
- McAleer, M.F.; Tuan, R.S. Cytotoxicant-induced trophoblast dysfunction and abnormal pregnancy outcomes: Role of zinc and metallothionein. Birth Defects Res. C Embryo Today 2004, 72, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Bush, P.G.; Mayhew, T.M.; Abramovich, D.R.; Aggett, P.J.; Burke, M.D.; Page, K.R. A quantitative study on the effects of maternal smoking on placental morphology and cadmium concentration. Placenta 2000, 21, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Guirlet, E.; Das, K.; Thomé, J.P.; Girondot, M. Maternal transfer of chlorinated contaminants in the leatherback turtles, Dermochelys coriacea, nesting in French Guiana. Chemosphere 2010, 79, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Marco, A.; López-Vicente, M.; Pérez-Mellado, V. Arsenic uptake by reptile flexible-shelled eggs from contaminated nest substrates and toxic effect on embryos. Bull. Environ. Contam. Toxicol. 2004, 72, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Marco, A.; López-Vicente, M.; Pérez-Mellado, V. Soil acidification negatively affects embryonic development of flexible-shelled lizard eggs. Herpetol. J. 2005, 15, 107–111. [Google Scholar]
- Trinchella, F.; Cannetiello, M.; Simoniello, P.; Filosa, S.; Scudiero, R. Differential gene expression profiles in embryos of the lizard Podarcis sicula under in ovo exposure to cadmium. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Simoniello, P.; Motta, C.M.; Scudiero, R.; Trinchella, F.; Filosa, S. Cadmium-induced teratogenicity in lizard embryos: Correlation with metallothionein gene expression. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 119–127. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Control | Acute Cd-Exposure | Chronic Cd-Exposure |
|---|---|---|---|
| mRNA/Protein | mRNA/Protein | mRNA/Protein | |
| Brain | +++/n.d. | No change/n.d. | No change/n.d. |
| Liver | ++/+++ | +3-fold/+2-fold | No change/+2-fold |
| Kidney | ++/n.d. | +4-fold/n.d. | +4-fold/n.d. |
| Ovary | +/not present | +3-fold/+++ | +4-fold/++ |
| Gut | +/n.d. | n.d./n.d. | +30-fold/n.d. |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scudiero, R.; Verderame, M.; Motta, C.M.; Simoniello, P. Unravelling the Role of Metallothionein on Development, Reproduction and Detoxification in the Wall Lizard Podarcis sicula. Int. J. Mol. Sci. 2017, 18, 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071569
Scudiero R, Verderame M, Motta CM, Simoniello P. Unravelling the Role of Metallothionein on Development, Reproduction and Detoxification in the Wall Lizard Podarcis sicula. International Journal of Molecular Sciences. 2017; 18(7):1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071569
Chicago/Turabian StyleScudiero, Rosaria, Mariailaria Verderame, Chiara Maria Motta, and Palma Simoniello. 2017. "Unravelling the Role of Metallothionein on Development, Reproduction and Detoxification in the Wall Lizard Podarcis sicula" International Journal of Molecular Sciences 18, no. 7: 1569. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18071569