Novel Insights into the Adipokinome of Obese and Obese/Diabetic Mouse Models

 , , and
, , and
Abstract
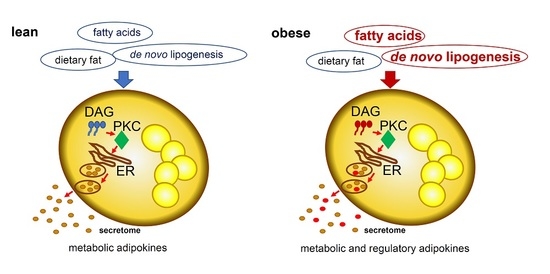
:
1. Introduction
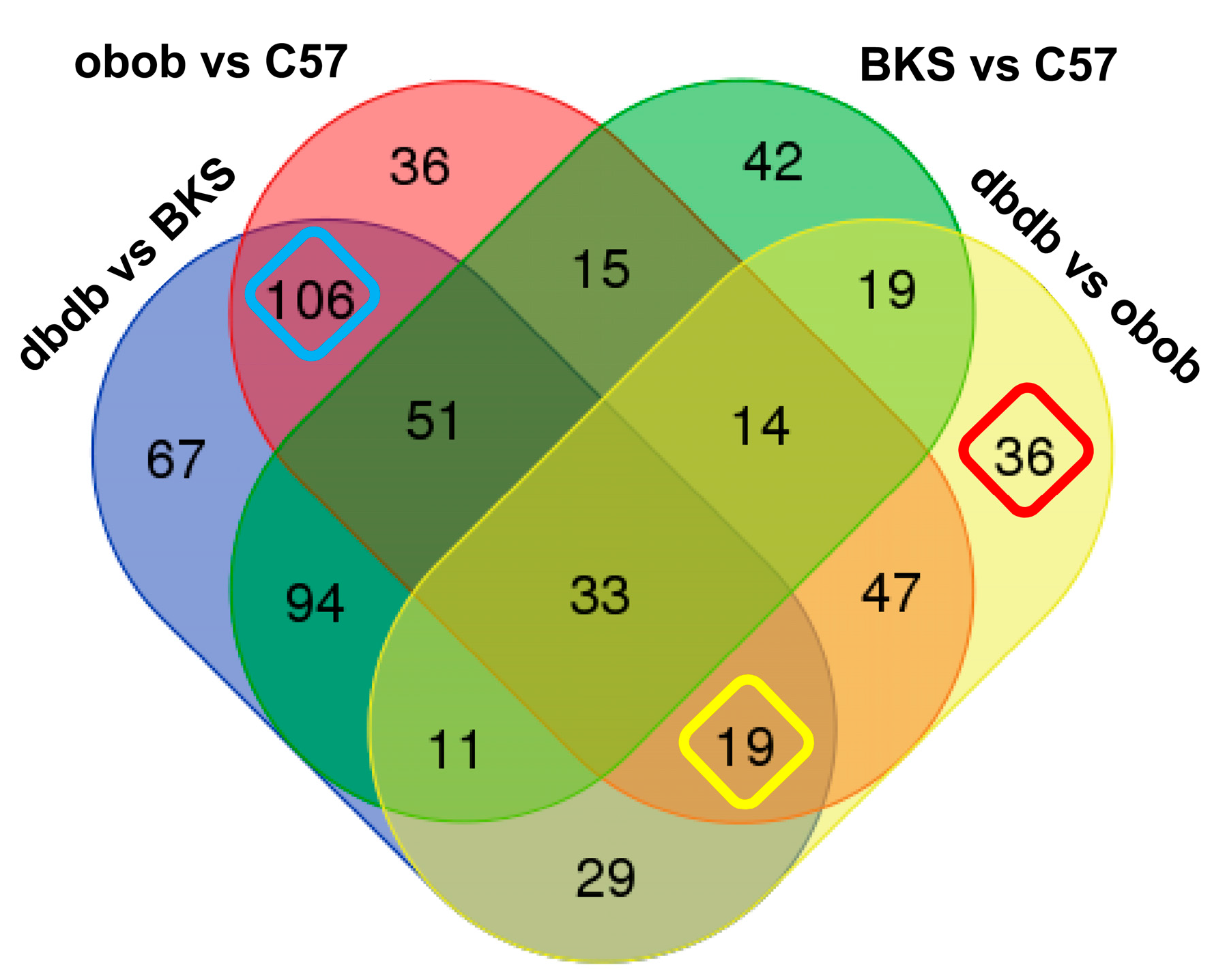
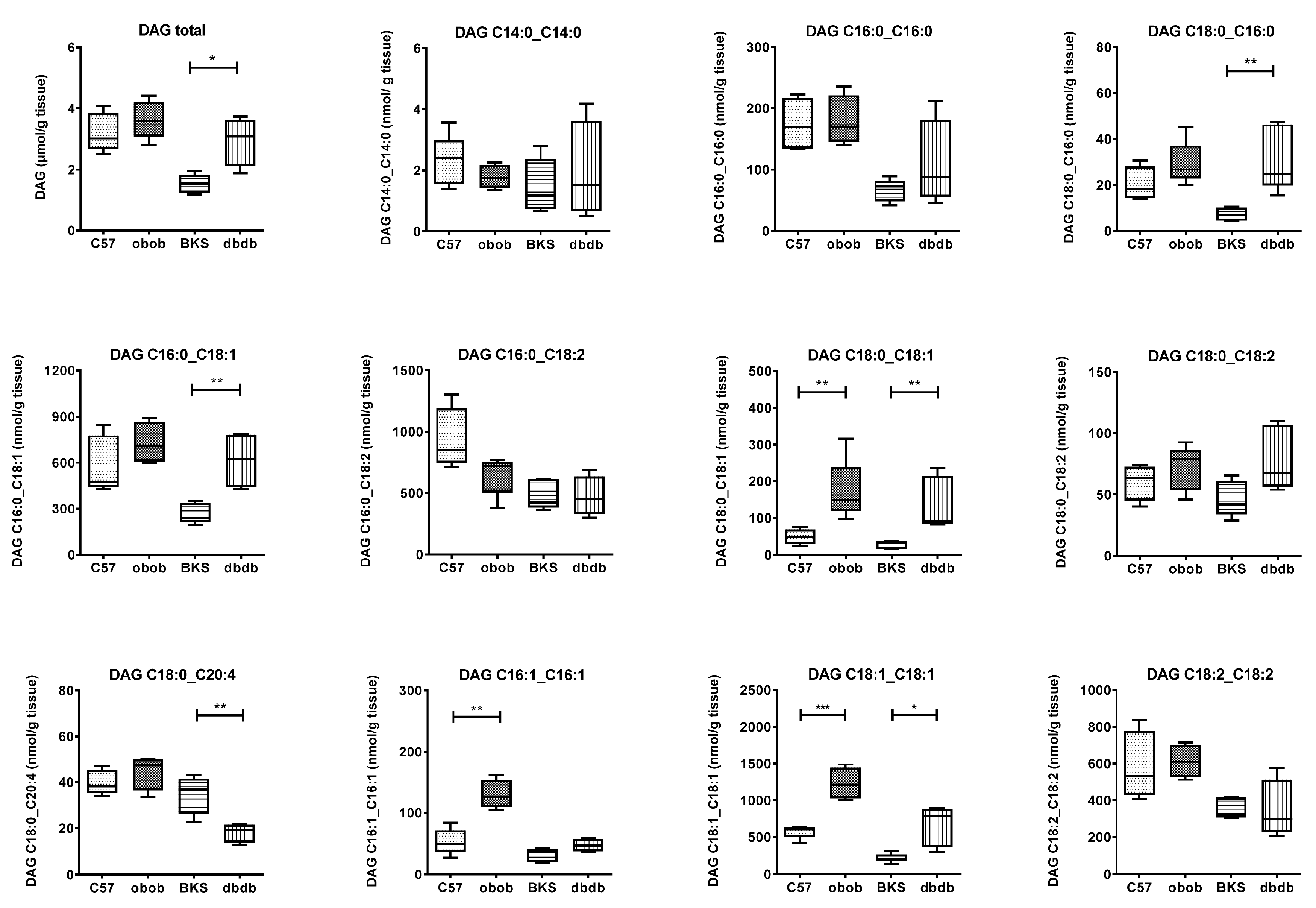
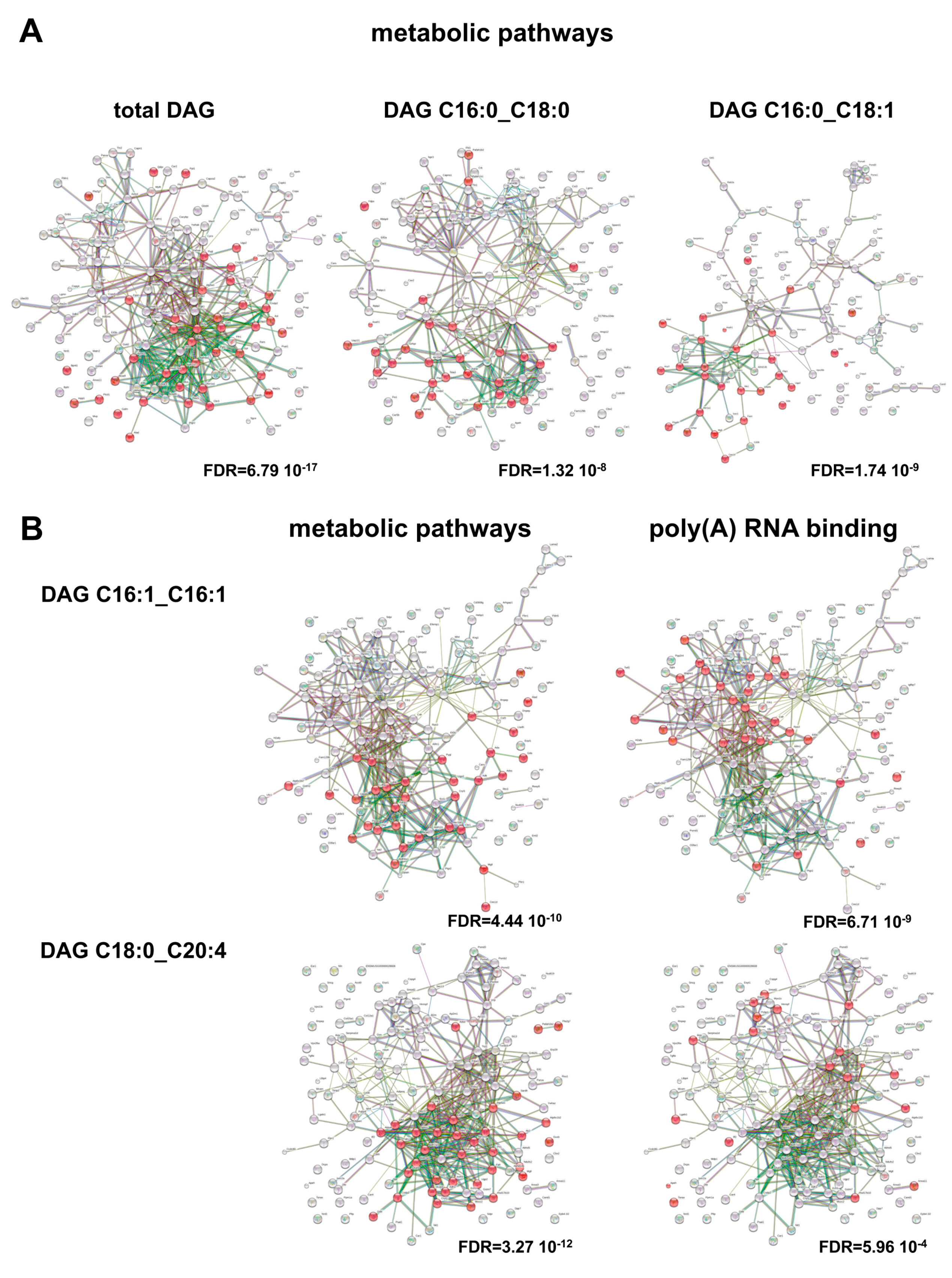
2. Results and Discussion
3. Materials and Methods
3.1. Mouse Models
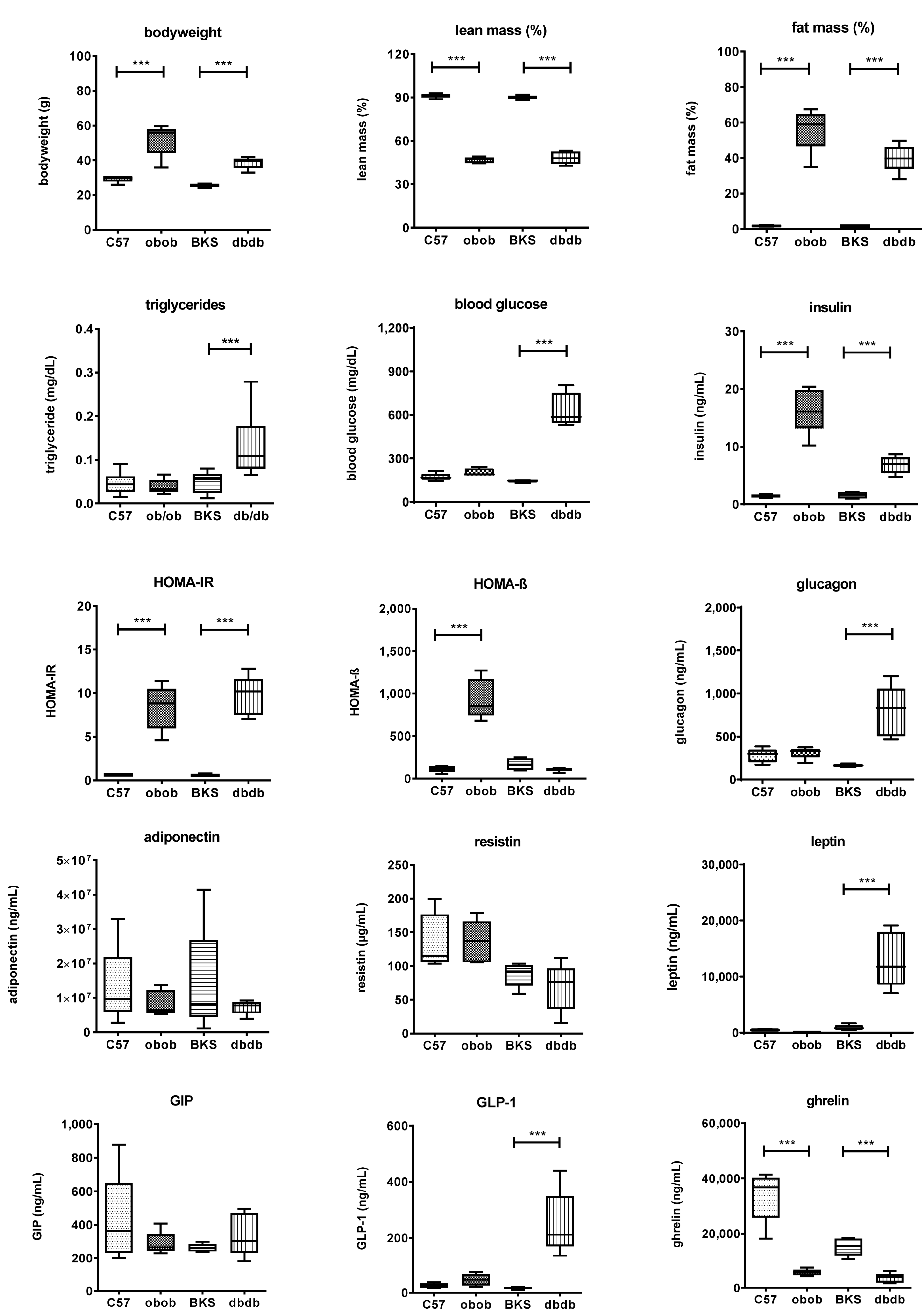
3.2. Metabolic Characterization of the Mouse Models
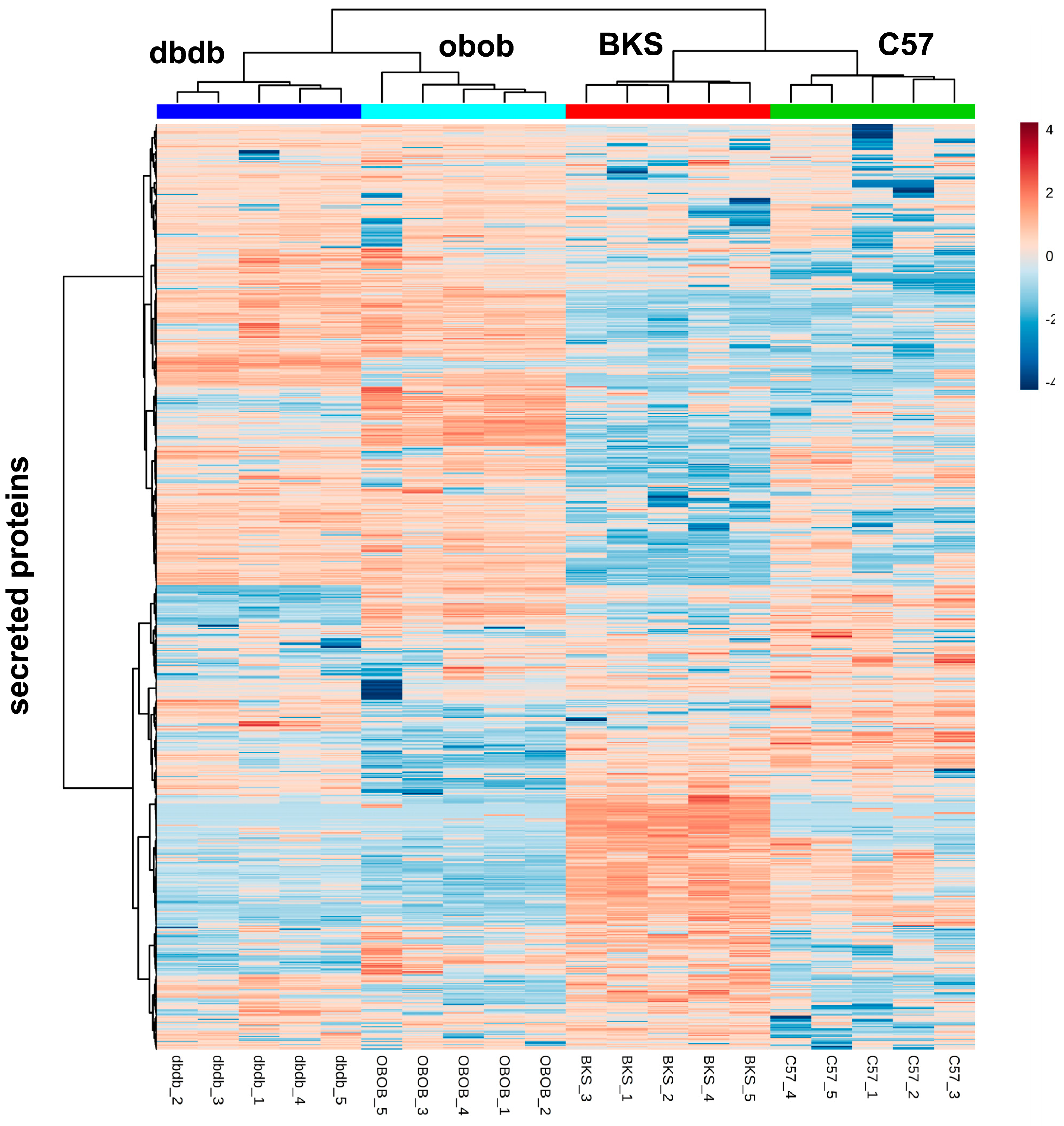
3.3. Secretome Profiling by Liquid Chromatography (LC)-Electrospray Ionization (ESI)-MS/MS and Data Analyses
3.4. Lipid Analysis of Adipose Tissue
3.5. Prediction and Annotation of Secretory Proteins
3.6. Web-Based Functional Annotation
3.7. Statistical Methods
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Meldrum, D.R.; Morris, M.A.; Gambone, J.C. Obesity pandemic: Causes, consequences, and solutions-but do we have the will? Fertil. Steril. 2017, 107, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.E.; do Carmo, J.M.; da Silva, A.A.; Wang, Z.; Hall, M.E. Obesity-induced hypertension: Interaction of neurohumoral and renal mechanisms. Circ. Res. 2015, 116, 991–1006. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, A.E.; Goodwin, P.J.; Chlebowski, R.T.; Pan, K.; Stambolic, V.; Dowling, R.J. Association of obesity-related metabolic disruptions with cancer risk and Outcome. J. Clin. Oncol. 2016, 34, 4249–4255. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, S.; Goeddeke, S.; Poschmann, G.; Dicken, H.D.; Jacob, S.; Nitzgen, U.; Passlack, W.; Stühler, K.; Ouwens, D.M.; Al-Hasani, H.; et al. Identification of novel adipokines differential regulated in C57BL/Ks and C57BL/6. Arch. Physiol. Biochem. 2014, 120, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Coleman, D.L.; Hummel, K.P. Symposium IV: Diabetic syndrome in animals. Influence of genetic background on the expression of mutations at the diabetes locus in the mouse. II. Studies on background modifiers. Isr. J. Med. Sci. 1975, 11, 708–713. [Google Scholar] [PubMed]
- Hummel, K.P.; Coleman, D.L.; Lane, P.W. The influence of genetic background on expression of mutations at the diabetes locus in the mouse. I. C57BL-KsJ and C57BL-6J strains. Biochem. Genet. 1972, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Unger, R.H.; Clark, G.O.; Scherer, P.E.; Orci, L. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Biochim. Biophys. Acta 2010, 1801, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.C.; Shulmann, G.I. Roles of Diacylglycerols and Ceramides in Hepatic Insulin Resistance. Trends Pharmacol. Sci. 2017, 38, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Sampson, S.R.; Cooper, D.R. Specific protein kinase C isoforms as transducers and modulators of insulin signaling. Mol. Genet. Metab. 2006, 89, 32–47. [Google Scholar] [CrossRef] [PubMed]
- Leiter, E.H.; Chapman, H.D.; Coleman, D.L. The influence of genetic background on the expression of mutations at the diabetes locus in the mouse. V. Interaction between the db gene and hepatic sex steroid sulfotransferases correlates with gender-dependent susceptibility to hyperglycemia. Endocrinology 1989, 124, 912–922. [Google Scholar] [CrossRef] [PubMed]
- Leiter, E.H.; Reifsnyder, P.C.; Flurkey, K.; Partke, H.-J.; Junger, E.; Herberg, L. NIDDM genes in mice. Deleterious synergism by both parental genomes contributes to diabetic thresholds. Diabetes 1998, 47, 1287–1295. [Google Scholar] [CrossRef] [PubMed]
- Schattenberg, P.; Gallem, R. Steatohepatitis: Of Mice and Man Animal Models of Non-Alcoholic Liver and Metabolic Syndrome. Dig. Dis. 2010, 28, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M.; Leibel, R.L.; Siegel, D.S.; Walsh, J.; Bahary, N. Molecular mapping of the mouse ob mutation. Genomics 1991, 11, 1054–1062. [Google Scholar] [CrossRef]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin- and leptin receptor-deficient rodent models: Relevance for human type 2 diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Rabouille, C. Pathways of Unconventional Protein Secretion. Trends Cell Biol. 2017, 27, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.; Wang, Y.; Chandrasekaran, S.; Witten, D.M.; Price, N.D. Molecular signatures from omics data: From chaos to consensus. Biotechnol. J. 2012, 8, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Diacylglycerol activation of protein kinase Cε and hepatic insulin resistance. Cell Metab. 2012, 15, 574–584. [Google Scholar] [CrossRef] [PubMed]
- Itani, S.I.; Ruderman, N.B.; Schmieder, F.; Boden, G. Lipid-induced insulin resistance in human muscle is associated with changes in diacylglycerol, protein kinase C, and IκB-α. Diabetes 2002, 51, 2005–2011. [Google Scholar] [CrossRef] [PubMed]
- Kumashiro, N.; Erion, D.M.; Zhang, D.; Kahn, M.; Beddow, S.A.; Chu, X.; Still, C.D.; Gerhard, G.S.; Han, X.; Dziura, J.; et al. Cellular mechanism of insulin resistance in nonalcoholic fatty liver disease. Proc. Natl. Acad. Sci. USA 2011, 108, 16381–16385. [Google Scholar] [CrossRef] [PubMed]
- Magkos, F.; Su, X.; Bradley, D.; Fabbrini, E.; Conte, C.; Eagon, J.C.; Varela, J.E.; Brunt, E.M.; Patterson, B.W.; Klein, S. Intrahepatic diacylglycerol content is associated with hepatic insulin resistance in obese subjects. Gastroenterology 2012, 142, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Sriram, G.; Martinez, J.A.; McCabe, E.R.; Liao, J.C.; Dipple, K.M. Single-gene disorders: What role could moonlighting enzymes play? Am. J. Hum. Genet. 2005, 76, 911–924. [Google Scholar] [CrossRef] [PubMed]
- Castello, A.; Hentze, M.W.; Preiss, T. Metabolic Enzymes Enjoying New Partnerships as RNA-Binding Proteins. Trends Endocrinol. Metab. 2015, 12, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Kao, A.W.; Noda, Y.; Johnson, J.H.; Pessin, J.E.; Saltiel, A.R. Aldolase mediates the association of F-actin with the insulin-responsive glucose transporter GLUT4. J. Biol. Chem. 1999, 274, 17742–17747. [Google Scholar] [CrossRef] [PubMed]
- Miles, L.A.; Dahlberg, C.M.; Plescia, J.; Felez, J.; Kato, K.; Plow, E.F. Role of cell-surface lysines in plasmin(ogen)-binding to cells: Identification of α-enolase as a candidate plasminogen receptor. Biochemistry 1991, 30, 1682–1691. [Google Scholar] [CrossRef] [PubMed]
- Redlitz, A.; Fowler, B.J.; Plow, E.F.; Miles, L.A. The role of an enolase-related molecule in plasminogen binding to cells. Eur. J. Biochem. 1995, 227, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Knebel, B.; Hartwig, S.; Haas, J.; Lehr, S.; Goeddeke, S.; Susanto, F.; Bohne, L.; Jacob, S.; Koellmer, C.; Nitzgen, U.; et al. Peroxisomes compensate hepatic lipid overflow in mice with fatty liver. Biochim. Biophys. Acta 2015, 1851, 965–976. [Google Scholar] [CrossRef] [PubMed]
- Göddeke, S.; Kotzka, J.; Lehr, S. Investigating the adipose tissue secretome: A protocol to generate high-quality samples appropriate for comprehensive proteomic profiling. Methods Mol. Biol. 2015, 1295, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [PubMed]
- Bendtsen, J.D.; Jensen, L.J.; Blom, N.; von Heijne, G.; Brunak, S. Feature-based prediction of non-classical and leaderless protein secretion. Protein Eng. Des. Sel. 2004, 17, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A web-based compendium of exosomal cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein–protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Names | Protein ID | Gene Names | BKS_C57 | BKS_C57 |
|---|---|---|---|---|
| Log2 Fold Change | p-Value (Welch Test) | |||
| Complement factor D | P03953 | Cfd | −5.42 | 1.49 × 106 |
| cAMP-dependent protein kinase type II-β | P31324 | Prkar2b | −2.47 | 2.73 × 106 |
| Apolipoprotein E | P08226 | Apoe | −2.17 | 7.51 × 104 |
| Receptor expression-enhancing protein 6 | Q9JM62 | Reep6 | −2.14 | 9.00 × 104 |
| Lipoprotein lipase | P11152 | Lpl | −2.03 | 8.64 × 103 |
| Prolargin | Q9JK53 | Prelp | −1.90 | 8.59 × 104 |
| 26S protease regulatory subunit 10B | P62334 | Psmc6 | −1.89 | 6.65 × 105 |
| Coiled-coil domain-containing protein 80 | Q8R2G6 | Ccdc80 | −1.89 | 2.33 × 104 |
| Pentraxin-related protein PTX3 | P48759 | Ptx3 | −1.87 | 3.35 × 102 |
| Tenascin | Q80YX1 | Tnc | −1.86 | 6.09 × 105 |
| Inositol polyphosphate 1-phosphatase | P49442 | Inpp1 | 1.52 | 1.24 × 105 |
| Carbonic anhydrase 2 | P00920 | Ca2 | 1.57 | 1.29 × 104 |
| Dolichyl-diphosphooligosaccharide-protein glycosyltransferase 48 kDa subunit | O54734 | Ddost | 1.71 | 9.47 × 105 |
| Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 2 | Q9DBG6 | Rpn2 | 1.72 | 4.48 × 104 |
| Nodal modulator 1 | Q6GQT9 | Nomo1 | 1.78 | 1.74 × 105 |
| 60S ribosomal protein L12 | P35979 | Rpl12 | 1.86 | 9.57 × 105 |
| Ketohexokinase | P97328 | Khk | 1.94 | 9.40 × 105 |
| Carbonyl reductase 3 | Q8K354 | Cbr3 | 1.98 | 1.06 × 104 |
| Carbonic anhydrase 1 | P13634 | Ca1 | 2.38 | 3.45 × 107 |
| Glutathione S-transferase θ-2 | Q61133 | Gstt2 | 3.25 | 4.80 × 109 |
| Protein Names | Protein IDs | Gene Names | obob_C57 | obob_C57 |
| Complement factor D | P03953 | Cfd | −9.46 | 5.52 × 1010 |
| Collagen α-1(XII) chain | Q60847 | Col12a1 | −6.39 | 1.12 × 109 |
| Collagen α-5(VI) chain | A6H584 | Col6a5 | −6.20 | 1.22 × 109 |
| Angiotensinogen | P11859 | Agt | −4.43 | 2.77 × 107 |
| Fructose-1,6-bisphosphatase 1 | Q9QXD6 | Fbp1 | −3.94 | 1.45 × 107 |
| Carboxypeptidase Q | Q9WVJ3 | Cpq | −3.64 | 1.15 × 106 |
| α-Amylase 1 | P00687 | Amy1 | −3.64 | 1.84 × 105 |
| Coiled-coil domain-containing protein 80 | Q8R2G6 | Ccdc80 | −3.47 | 2.67 × 107 |
| Tissue α-l-fucosidase | Q99LJ1 | Fuca1 | −3.28 | 9.84 × 107 |
| Ganglioside GM2 activator | Q60648 | Gm2a | −3.26 | 2.29 × 106 |
| Actin-related protein 2/3 complex subunit 3 | Q9JM76 | Arpc3 | 1.94 | 4.89 × 105 |
| NADH-cytochrome b5 reductase 3 | Q9DCN2 | Cyb5r3 | 2.07 | 5.69 × 105 |
| Epoxide hydrolase 1 | Q9D379 | Ephx1 | 2.12 | 3.21 × 108 |
| Acyl-CoA dehydrogenase 9, mitochondrial | Q8JZN5 | Acad9 | 2.18 | 5.98 × 108 |
| Serum deprivation-response protein | Q63918 | Sdpr | 2.22 | 2.03 × 106 |
| Serpin H1 | P19324 | Serpinh1 | 2.25 | 3.14 × 105 |
| Galectin-3 | P16110 | Lgals3 | 2.25 | 6.50 × 107 |
| GTP:AMP phosphotransferase AK3, mitochondrial | Q9WTP7 | Ak3 | 2.28 | 2.06 × 107 |
| Apolipoprotein A-IV | P06728 | Apoa4 | 2.42 | 6.29 × 107 |
| Interleukin-6 | P08505 | Il6 | 3.19 | 6.96 × 107 |
| Protein Names | Protein IDs | Gene Names | dbdb_BKS | dbdb_BKS |
| Collagen α-5(VI) chain | A6H584 | Col6a5 | −7.87 | 3.53 × 1011 |
| Fructose-1,6-bisphosphatase 1 | Q9QXD6 | Fbp1 | −4.10 | 1.64 × 107 |
| Tissue α-l-fucosidase | Q99LJ1 | Fuca1 | −3.92 | 4.32 × 108 |
| Carboxypeptidase Q | Q9WVJ3 | Cpq | −3.34 | 1.63 × 106 |
| Complement factor D | P03953 | Cfd | −3.33 | 2.71 × 104 |
| Ganglioside GM2 activator | Q60648 | Gm2a | −3.28 | 1.18 × 106 |
| Carboxylesterase 1D | Q8VCT4 | Ces1d | −2.97 | 1.49 × 1011 |
| Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit 2 | Q9DBG6 | Rpn2 | −2.87 | 2.22 × 104 |
| Fructose-1,6-bisphosphatase isozyme 2 | P70695 | Fbp2 | −2.80 | 5.61 × 105 |
| Angiotensinogen | P11859 | Agt | −2.44 | 2.67 × 104 |
| Phospholipid transfer protein | P55065 | Pltp | 2.77 | 1.62 × 102 |
| Serum deprivation-response protein | Q63918 | Sdpr | 2.86 | 5.17 × 107 |
| Polymerase I and transcript release factor | O54724 | Ptrf | 2.93 | 1.87 × 108 |
| Platelet-activating factor acetylhydrolase | Q60963 | Pla2g7 | 2.97 | 4.03 × 103 |
| Vimentin | P20152 | Vim | 3.12 | 8.70 × 1011 |
| C-C motif chemokine 2 | P10148 | Ccl2 | 3.37 | 1.23 × 102 |
| Prolargin | Q9JK53 | Prelp | 3.39 | 1.65 × 106 |
| cAMP-dependent protein kinase type II-β | P31324 | Prkar2b | 3.70 | 4.52 × 109 |
| Growth-regulated α protein | P12850 | Cxcl1 | 4.36 | 1.13 × 106 |
| Interleukin-6 | P08505 | Il6 | 5.35 | 2.09 × 1010 |
| Protein Names | Protein IDs | Gene Names | dbdb_obob | dbdb_obob |
| Transthyretin | P07309 | Ttr | −3.06 | 9.14 × 108 |
| ATP-citrate synthase | Q91V92 | Acly | −2.98 | 4.13 × 1011 |
| Sarcosine dehydrogenase, mitochondrial | Q99LB7 | Sardh | −2.34 | 3.51 × 106 |
| GTP:AMP phosphotransferase AK3, mitochondrial | Q9WTP7 | Ak3 | −2.15 | 1.91 × 107 |
| Serpin H1 | P19324 | Serpinh1 | −2.01 | 2.23 × 104 |
| Acyl-CoA dehydrogenase 9, mitochondrial | Q8JZN5 | Acad9 | −1.99 | 2.95 × 107 |
| Vitamin D-binding protein | P21614 | Gc | −1.90 | 9.29 × 107 |
| 3-Hydroxyisobutyrate dehydrogenase, mitochondrial | Q99L13 | Hibadh | −1.86 | 3.24 × 104 |
| Pyruvate dehydrogenase E1 mitochondrial | P35486 | Pdha1 | −1.81 | 1.05 × 103 |
| Citrate synthase, mitochondrial | Q9CZU6 | Cs | −1.70 | 1.07 × 105 |
| Calmodulin | P62204 | Calm1 | 1.51 | 3.03 × 105 |
| Laminin subunit β-2 | Q61292 | Lamb2 | 1.55 | 7.46 × 107 |
| C-X-C motif chemokine 5;GCP-2(1-78);GCP-2(9-78) | P50228 | Cxcl5 | 1.77 | 4.79 × 104 |
| Prolargin | Q9JK53 | Prelp | 1.82 | 3.83 × 103 |
| Pentraxin-related protein PTX3 | P48759 | Ptx3 | 1.84 | 9.82 × 103 |
| Desmin | P31001 | Des | 2.09 | 5.46 × 107 |
| Lactotransferrin | P08071 | Ltf | 2.15 | 9.64 × 104 |
| Collagen α-1(XII) chain | Q60847 | Col12a1 | 4.45 | 1.31 × 107 |
| Protein Names | Protein ID | Gene Names | SP(+) | SP(−) | NP |
|---|---|---|---|---|---|
| BKS_C57 | |||||
| Tyrosine-protein phosphatase non-receptor type 6 | P29351 | Ptpn6 | + | ||
| NADPH-cytochrome P450 reductase | P37040 | Por | + | ||
| Vacuolar protein sorting-associated protein 13C | Q8BX70 | Vps13c | + | ||
| Annexin A11 | P97384 | Anxa11 | + | ||
| Epidermal growth factor receptor substrate 15 | P42567 | Eps15 | + | ||
| Adenosine deaminase | P03958 | Ada | + | ||
| Pyruvate dehydrogenase E1 subunit α, mitochondrial | P35486 | Pdha1 | + | ||
| Carnitine O-acetyltransferase | P47934 | Crat | + | ||
| Histidine triad nucleotide-binding protein 1 | P70349 | Hint1 | + | ||
| Heat shock protein 75 kDa, mitochondrial | Q9CQN1 | Trap1 | + | ||
| COP9 signalosome complex subunit 8 | Q8VBV7 | Cops8 | + | ||
| Endothelial cell-selective adhesion molecule | Q925F2 | Esam | + | ||
| Carbonic anhydrase 4 | Q64444 | Ca4 | + | ||
| Arsenite methyltransferase | Q91WU5 | As3mt | + | ||
| Deoxyguanosine kinase, mitochondrial | Q9QX60 | Dguok | + | ||
| H-2 class I histocompatibility antigen, D-B α chain | P01899 | H2-D1 | + | ||
| Phosphoserine phosphatase | Q99LS3 | Psph | + | ||
| Mannosyl-oligosaccharide 1,2-α-mannosidase IA | P45700 | Man1a1 | + | ||
| Band 4.1-like protein 2 | O70318 | Epb41l2 | + | ||
| Glucosamine-6-phosphate isomerase 1 | O88958 | Gnpda1 | + | ||
| β-Galactosidase | P23780 | Glb1 | + | ||
| Semaphorin-7A | Q9QUR8 | Sema7a | + | ||
| Very long-chain acyl-CoA dehydrogenase, mitochondrial | P50544 | Acadvl | + | ||
| Inter-α-trypsin inhibitor heavy chain H1 | Q61702 | Itih1 | + | ||
| Glutathione S-transferase θ-1 | Q64471 | Gstt1 | + | ||
| Catenin β-1 | Q02248 | Ctnnb1 | + | ||
| BTB/POZ domain-containing protein KCTD12 | Q6WVG3 | Kctd12 | + | ||
| Retinol-binding protein 2 | Q08652 | Rbp2 | + | ||
| Small nuclear ribonucleoprotein Sm D3 | P62320 | Snrpd3 | + | ||
| Coronin-7 | Q9D2V7 | Coro7 | + | ||
| DNA topoisomerase 2-β | Q64511 | Top2b | + | ||
| Mast cell protease 2 | P15119 | Mcpt2 | + | ||
| Retinol-binding protein 1 | Q00915 | Rbp1 | + | ||
| Phosphoglycolate phosphatase | Q8CHP8 | Pgp | + | ||
| β-Hexosaminidase subunit β | P20060 | Hexb | + | ||
| Putative hydroxypyruvate isomerase | Q8R1F5 | Hyi | + | ||
| S-adenosylmethionine synthase isoform type-2 | Q3THS6 | Mat2a | + | ||
| Bisphosphoglycerate mutase | P15327 | Bpgm | + | ||
| obob_C57 | |||||
| Semaphorin-7A | Q9QUR8 | Sema7a | + | ||
| α-Methylacyl-CoA racemase | O09174 | Amacr | + | ||
| Leukemia inhibitory factor | P09056 | Lif | + | ||
| Fructose-1,6-bisphosphatase isozyme 2 | P70695 | Fbp2 | + | ||
| Thiosulfate sulfurtransferase | P52196 | Tst | + | ||
| Eukaryotic translation initiation factor 3 subunit C | Q8R1B4 | Eif3c | + | ||
| Tyrosine-protein phosphatase non-receptor type 6 | P29351 | Ptpn6 | + | ||
| Signal transducer and activator of transcription 1 | P42225 | Stat1 | + | ||
| Fibromodulin | P50608 | Fmod | + | ||
| Growth-regulated α protein | P12850 | Cxcl1 | + | ||
| Plasminogen activator inhibitor 2, macrophage | P12388 | Serpinb2 | + | ||
| 1-Acyl-sn-glycerol-3-phosphate acyltransferase β | Q8K3K7 | Agpat2 | + | ||
| BTB/POZ domain-containing protein KCTD12 | Q6WVG3 | Kctd12 | + | ||
| C-X-C motif chemokine 3 | Q6W5C0 | Cxcl3 | + | ||
| Epidermal growth factor receptor substrate 15 | P42567 | Eps15 | + | ||
| 26S proteasome non-ATPase regulatory subunit 13 | Q9WVJ2 | Psmd13 | + | ||
| Glutathione S-transferase Mu 7 | Q80W21 | Gstm7 | + | ||
| Hereditary hemochromatosis protein homolog | P70387 | Hfe | + | ||
| Very long-chain specific acyl-CoA dehydrogenase, mitochondrial | P50544 | Acadvl | + | ||
| Cadherin-16 | O88338 | Cdh16 | + | ||
| C-X-C motif chemokine 5 | P50228 | Cxcl5 | + | ||
| α-1-Antitrypsin 1-5 | Q00898 | Serpina1e | + | ||
| dbdb_BKS | |||||
| Tyrosine-protein phosphatase non-receptor type 6 | P29351 | Ptpn6 | + | ||
| Hereditary hemochromatosis protein homolog | P70387 | Hfe | + | ||
| Proteasome activator complex subunit 3 | P61290 | Psme3 | + | ||
| C-C motif chemokine 9 | P51670 | Ccl9 | + | ||
| Tripeptidyl-peptidase 2 | Q64514 | Tpp2 | + | ||
| Vacuolar protein sorting-associated protein 13C | Q8BX70 | Vps13c | + | ||
| Fibromodulin | P50608 | Fmod | + | ||
| Eukaryotic translation initiation factor 2 subunit 1 | Q6ZWX6 | Eif2s1 | + | ||
| Metalloproteinase inhibitor 2 | P25785 | Timp2 | + | ||
| Mannosyl-oligosaccharide 1,2-α-mannosidase IA | P45700 | Man1a1 | + | ||
| Importin-9 | Q91YE6 | Ipo9 | + | ||
| Carnitine O-acetyltransferase | P47934 | Crat | + | ||
| Small glutamine-rich tetratricopeptide repeat-containing protein α | Q8BJU0 | Sgta | + | ||
| T-complex protein 1 subunit ζ | P80317 | Cct6a | + | ||
| Epidermal growth factor receptor substrate 15 | P42567 | Eps15 | + | ||
| Arginase-1 | Q61176 | Arg1 | + | ||
| Granulocyte colony-stimulating factor | P09920 | Csf3 | + | ||
| AP-2 complex subunit mu | P84091 | Ap2m1 | + | ||
| Phosphoserine phosphatase | Q99LS3 | Psph | + | ||
| Histidine triad nucleotide-binding protein 1 | P70349 | Hint1 | + | ||
| Plasminogen activator inhibitor 2, macrophage | P12388 | Serpinb2 | + | ||
| Basigin | P18572 | Bsg | + | ||
| Dynactin subunit 2 | Q99KJ8 | Dctn2 | + | ||
| COP9 signalosome complex subunit 8 | Q8VBV7 | Cops8 | + | ||
| Coatomer subunit ζ-1 | P61924 | Copz1 | + | ||
| 6-Pyruvoyl tetrahydrobiopterin synthase | Q9R1Z7 | Pts | + | ||
| Inter-α-trypsin inhibitor heavy chain H1 | Q61702 | Itih1 | + | ||
| Annexin A11 | P97384 | Anxa11 | + | ||
| Plastin-1 | Q3V0K9 | Pls1 | + | ||
| Eosinophil cationic protein 1 | P97426 | Ear1 | + | ||
| Isopentenyl-diphosphate δ-isomerase 1 | P58044 | Idi1 | + | ||
| Cadherin-1 | P09803 | Cdh1 | + | ||
| 4-Hydroxy-2-oxoglutarate aldolase, mitochondrial | Q9DCU9 | Hoga1 | + | ||
| Nucleoside diphosphate-linked moiety X motif 19, mitochondrial | P11930 | Nudt19 | + | ||
| Dolichyl-diphosphooligosaccharide-protein glycosyltransferase 48 kDa subunit | O54734 | Ddost | + | ||
| Thiosulfate sulfurtransferase | P52196 | Tst | + | ||
| Cysteine and glycine-rich protein 1 | P97315 | Csrp1 | + | ||
| N(G),N(G)-Dimethylarginine dimethylaminohydrolase 1 | Q9CWS0 | Ddah1 | + | ||
| Small nuclear ribonucleoprotein Sm D3 | P62320 | Snrpd3 | + | ||
| SUMO-conjugating enzyme UBC9 | P63280 | Ube2i | + | ||
| Band 4.1-like protein 2 | O70318 | Epb41l2 | + | ||
| Semaphorin-7A | Q9QUR8 | Sema7a | + | ||
| Adenosine deaminase | P03958 | Ada | + | ||
| Coronin-7 | Q9D2V7 | Coro7 | + | ||
| Nodal modulator 1 | Q6GQT9 | Nomo1 | + | ||
| Phosphoglycolate phosphatase | Q8CHP8 | Pgp | + | ||
| C-X-C motif chemokine 3 | Q6W5C0 | Cxcl3 | + | ||
| β-Hexosaminidase subunit β | P20060 | Hexb | + | ||
| Putative hydrolase RBBP9 | O88851 | Rbbp9 | + | ||
| Ketohexokinase | P97328 | Khk | + | ||
| Interleukin-1 receptor antagonist protein | P25085 | Il1rn | + | ||
| α-Methylacyl-CoA racemase | O09174 | Amacr | + | ||
| Protein kinase C δ-binding protein | Q91VJ2 | Prkcdbp | + | ||
| Retinol-binding protein 1 | Q00915 | Rbp1 | + | ||
| S-adenosylmethionine synthase isoform type-2 | Q3THS6 | Mat2a | + | ||
| Glutathione S-transferase θ-2 | Q61133 | Gstt2 | + | ||
| Cadherin-16 | O88338 | Cdh16 | + | ||
| Bisphosphoglycerate mutase | P15327 | Bpgm | + | ||
| C-X-C motif chemokine 5 | P50228 | Cxcl5 | + | ||
| α-1-Antitrypsin 1-5 | Q00898 | Serpina1e | + | ||
| dbdb_obob | |||||
| Deoxyguanosine kinase, mitochondrial | Q9QX60 | Dguok | + | ||
| Semaphorin-7A | Q9QUR8 | Sema7a | + | ||
| Glutathione S-transferase Mu 7 | Q80W21 | Gstm7 | + | ||
| Phosphoglucomutase-like protein 5 | Q8BZF8 | Pgm5 | + | ||
| 1-Acyl-sn-glycerol-3-phosphate acyltransferase β | Q8K3K7 | Agpat2 | + | ||
| Regulator of microtubule dynamics protein 3 | Q3UJU9 | Rmdn3 | + | ||
| Eosinophil cationic protein 1 | P97426 | Ear1 | + | ||
| H-2 class I histocompatibility antigen, D-B α chain | P01899 | H2-D1 | + | ||
| Interleukin-1 receptor antagonist protein | P25085 | Il1rn | + | ||
| Putative hydroxypyruvate isomerase | Q8R1F5 | Hyi | + |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knebel, B.; Goeddeke, S.; Poschmann, G.; Markgraf, D.F.; Jacob, S.; Nitzgen, U.; Passlack, W.; Preuss, C.; Dicken, H.-D.; Stühler, K.; et al. Novel Insights into the Adipokinome of Obese and Obese/Diabetic Mouse Models. Int. J. Mol. Sci. 2017, 18, 1928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091928
Knebel B, Goeddeke S, Poschmann G, Markgraf DF, Jacob S, Nitzgen U, Passlack W, Preuss C, Dicken H-D, Stühler K, et al. Novel Insights into the Adipokinome of Obese and Obese/Diabetic Mouse Models. International Journal of Molecular Sciences. 2017; 18(9):1928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091928
Chicago/Turabian StyleKnebel, Birgit, Simon Goeddeke, Gereon Poschmann, Daniel F. Markgraf, Sylvia Jacob, Ulrike Nitzgen, Waltraud Passlack, Christina Preuss, Hans-Dieter Dicken, Kai Stühler, and et al. 2017. "Novel Insights into the Adipokinome of Obese and Obese/Diabetic Mouse Models" International Journal of Molecular Sciences 18, no. 9: 1928. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091928