Investigation of Antimicrobial Peptide Genes Associated with Fungus and Insect Resistance in Maize

Abstract
:
1. Introduction
2. Results
2.1. Identification of Maize AMPs
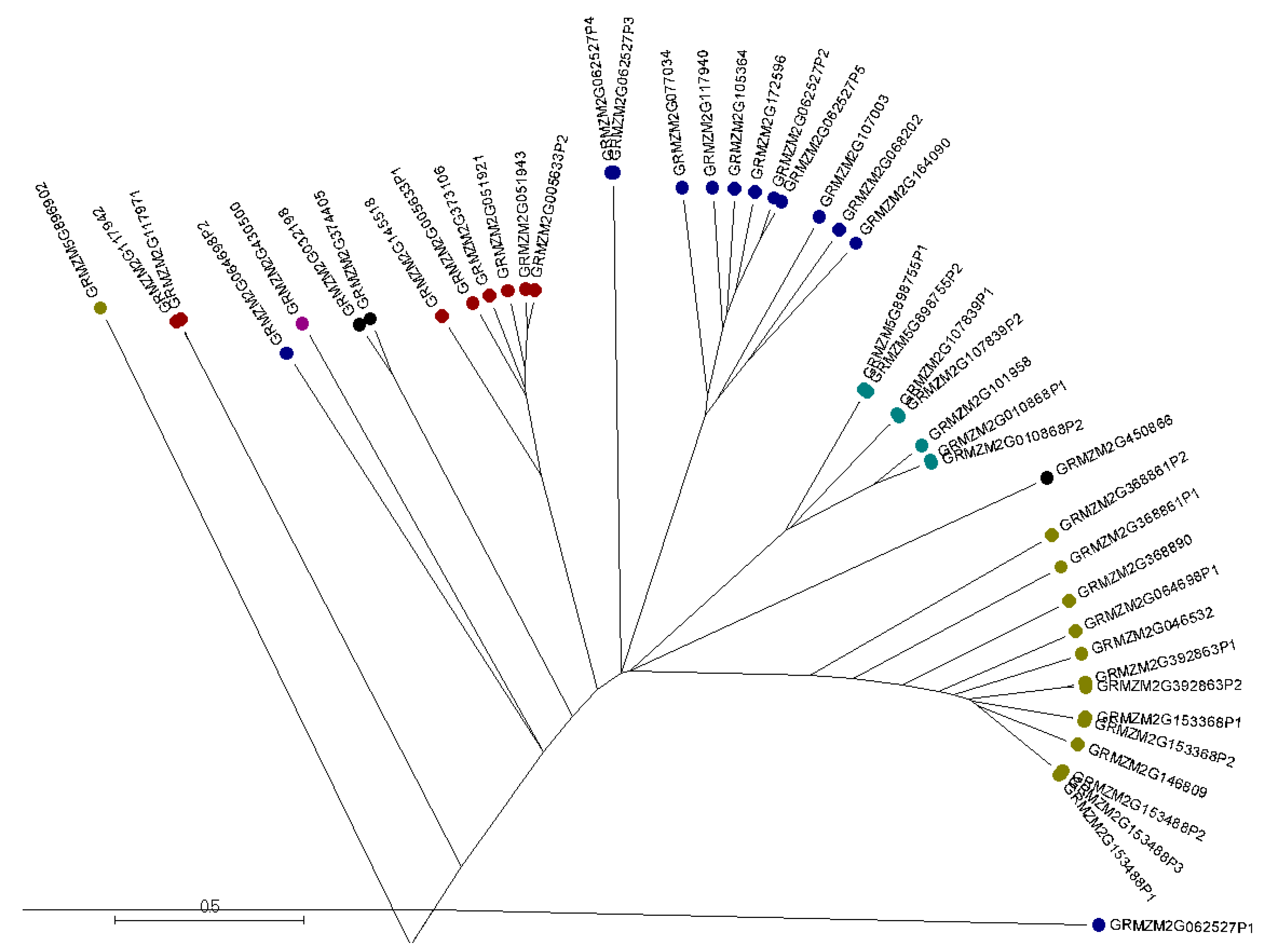
2.2. Phylogenetic Analysis and Protein Motif Detection of Maize AMPs
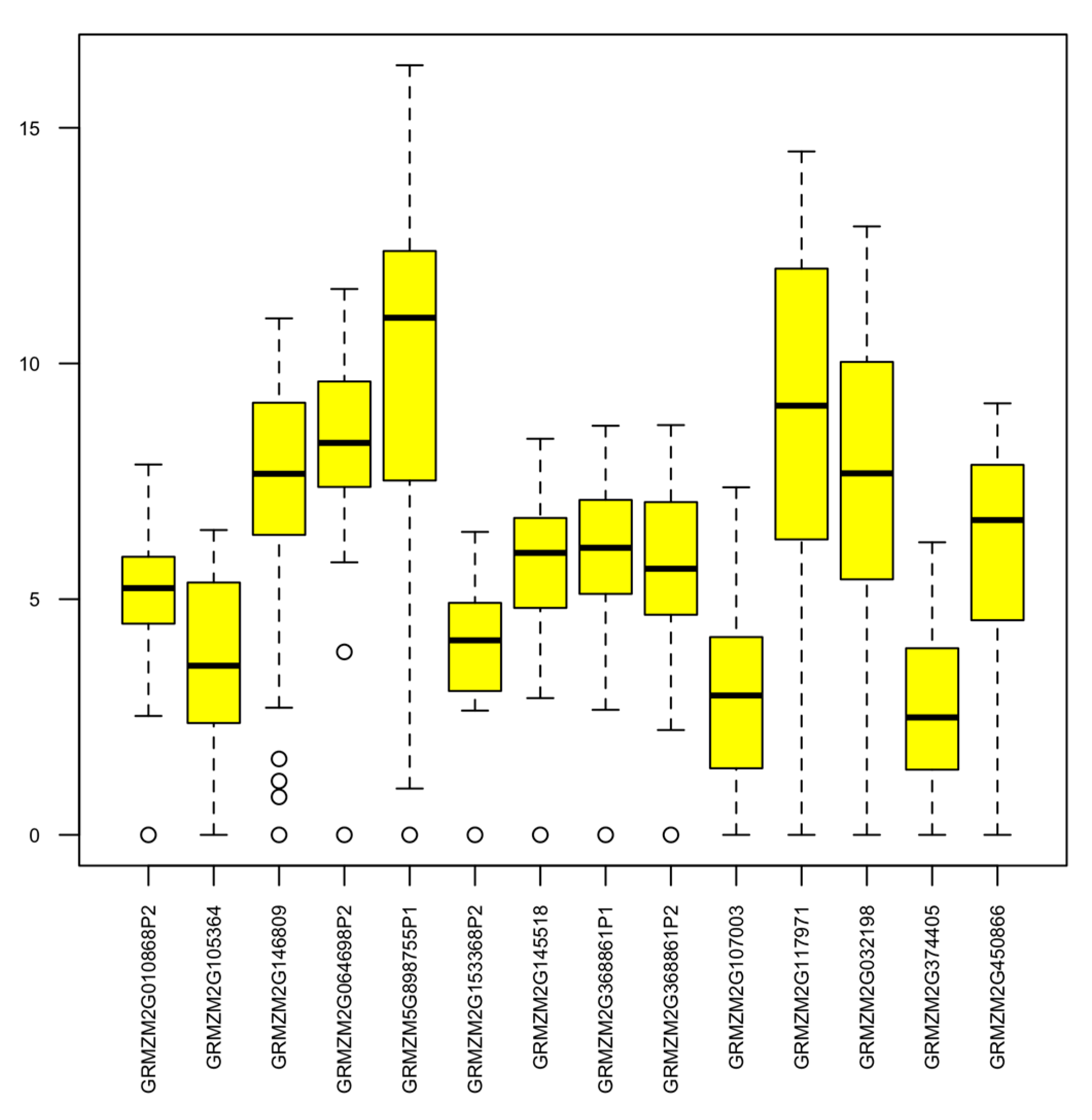
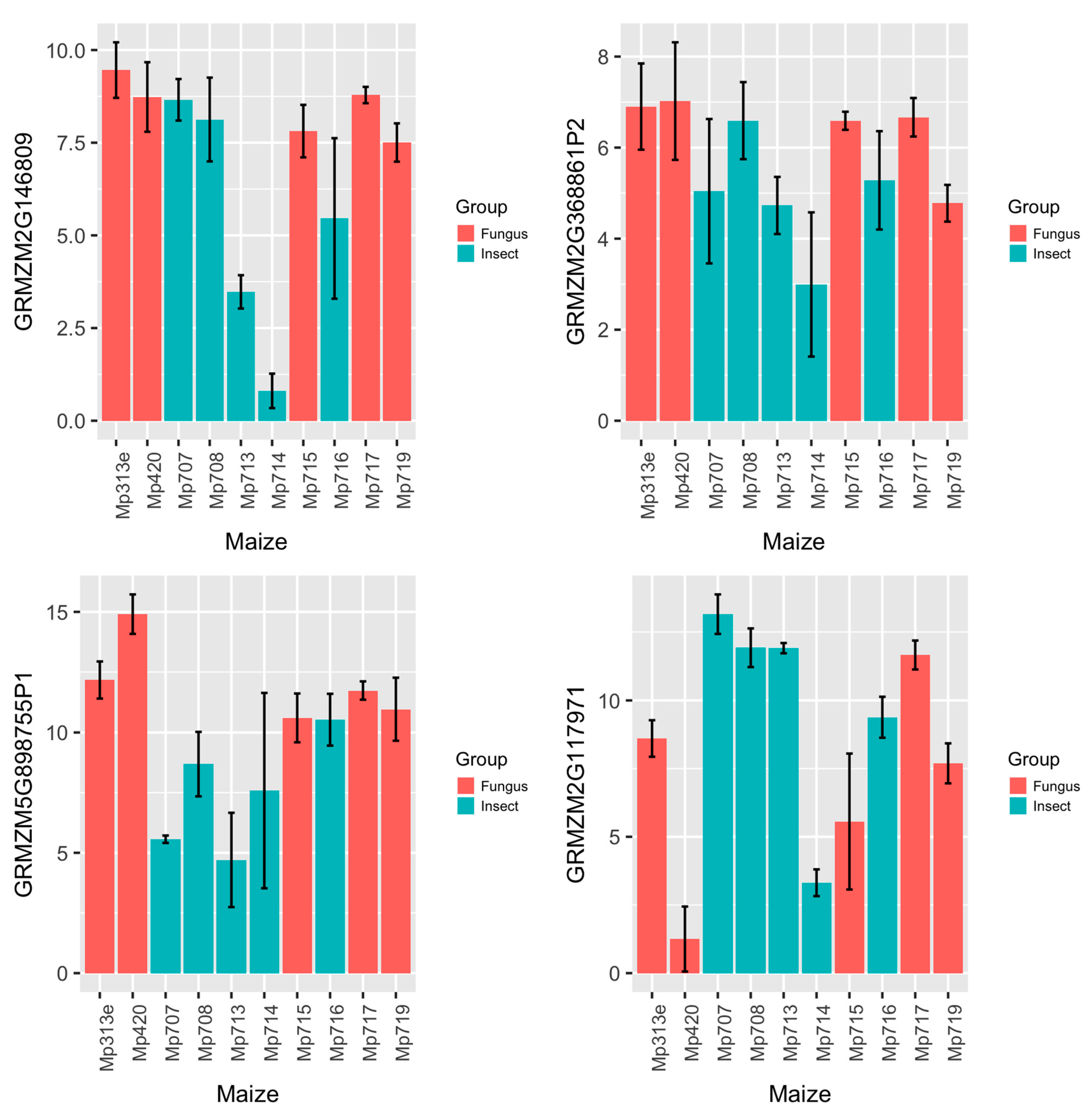
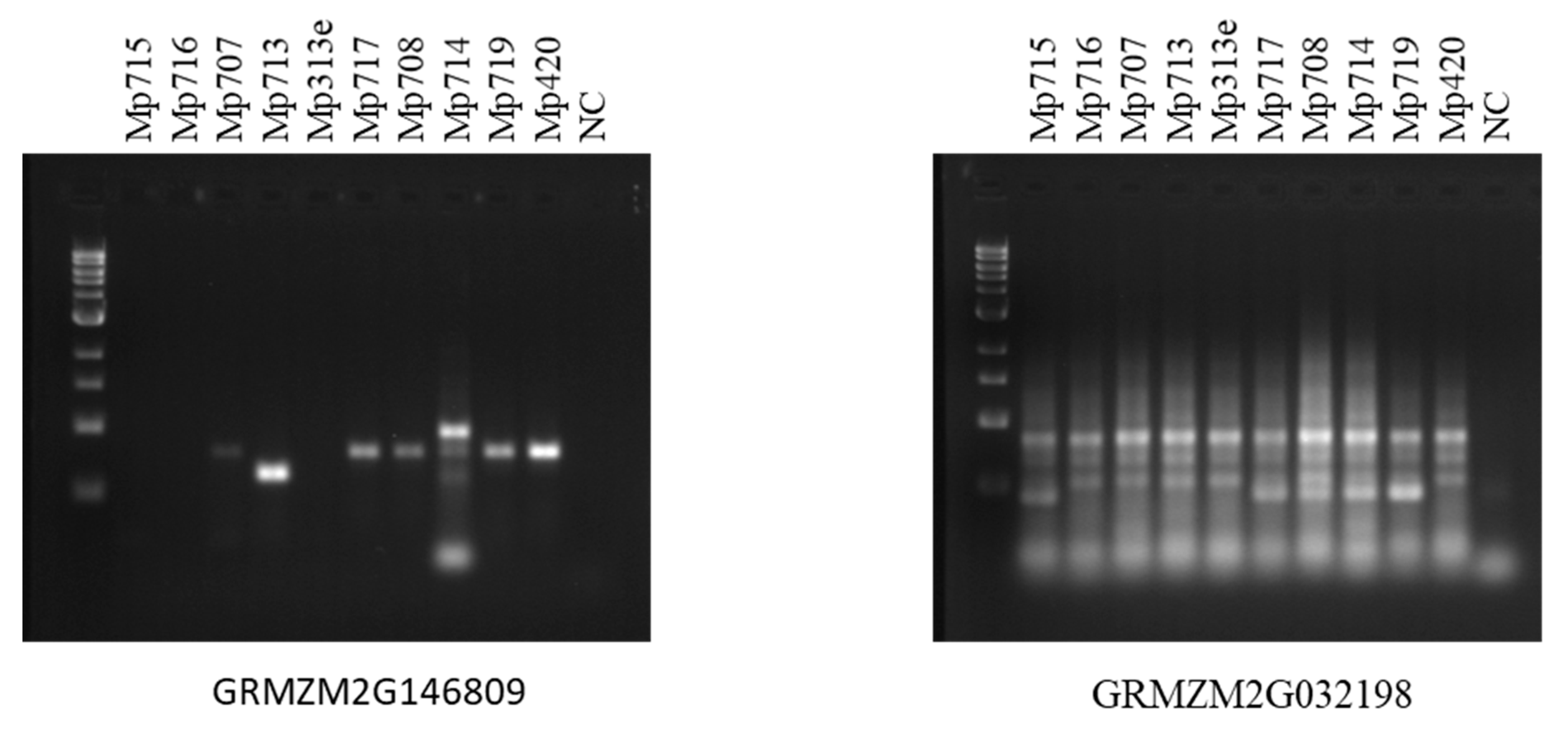
2.3. Expression Analysis of Maize AMP Genes and Polymorphism Study of Their Genomic Sequences
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. DNA Extraction
4.3. Database Search for Maize (Zea mays) Antimicrobial Peptide Gene and Protein Sequences
4.4. Design of Genomic DNA and cDNA Primers for Maize AMPs
4.5. Polymerase Chain Reaction for Genomic DNA
4.6. RNA Extraction
4.7. Synthesis of cDNA and qRT-PCR Analysis
4.8. Statistical and Bioinformatics Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tam, J.P.; Wang, S.; Wong, K.H.; Tan, W.L. Antimicrobial Peptides from Plants. Pharmaceuticals 2015, 8, 711–757. [Google Scholar] [CrossRef] [PubMed]
- Agriculture, R.G.; Canada, A. Host Defense Peptides and Their Potential as Therapeutic Agents; Epand, R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 111–136. [Google Scholar]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant antimicrobial peptides. Folia Microbiol. 2014, 59, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.F.S.; Silva, O.N.; Viana, J.C.; Porto, W.F.; Migliolo, L.; da Cunha, N.B.; Gomes, N.; Fensterseifer, I.C.M.; Colgrave, M.L.; Craik, D.J.; et al. Characterization of a Bioactive Acyclotide from Palicourea rigida. J. Nat. Prod. 2016, 79, 2767–2773. [Google Scholar] [CrossRef] [PubMed]
- Hammami, R.; Ben Hamida, J.; Vergoten, G.; Fliss, I. PhytAMP: A database dedicated to antimicrobial plant peptides. Nucleic Acids Res. 2009, 37, D963–D968. [Google Scholar] [CrossRef] [PubMed]
- Goyal, R.K.; Mattoo, A.K. Multitasking antimicrobial peptides in plant development and host defense against biotic/abiotic stress. Plant Sci. 2014, 228, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Herbel, V.; Sieber-Frank, J.; Wink, M. The antimicrobial peptide snakin-2 is upregulated in the defense response of tomatoes (Solanum lycopersicum) as part of the jasmonate-dependent signaling pathway. J. Plant Physiol. 2017, 208, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ravipati, A.S.; Poth, A.G.; Troeira Henriques, S.; Bhandari, M.; Huang, Y.-H.; Nino, J.; Colgrave, M.L.; Craik, D.J. Understanding the Diversity and Distribution of Cyclotides from Plants of Varied Genetic Origin. J. Nat. Prod. 2017, 80, 1522–1530. [Google Scholar] [CrossRef] [PubMed]
- Barbeta, B.L.; Marshall, A.T.; Gillon, A.D.; Craik, D.J.; Anderson, M.A. Plant cyclotides disrupt epithelial cells in the midgut of lepidopteran larvae. Proc. Natl. Acad. Sci. USA 2008, 105, 1221–1225. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J. Host-defense activities of cyclotides. Toxins 2012, 4, 139–156. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.W.; Cemazar, M.; Anderson, M.A.; Craik, D.J. Insecticidal plant cyclotides and related cystine knot toxins. Toxicon 2007, 49, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Troeira Henriques, S.; Craik, D.J. Cyclotide Structure and Function: The Role of Membrane Binding and Permeation. Biochemistry 2017, 56, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Weidmann, J.; Craik, D.J. Discovery, structure, function, and applications of cyclotides: Circular proteins from plants. J. Exp. Bot. 2016, 67, 4801–4812. [Google Scholar] [CrossRef] [PubMed]
- Daly, N.L.; Gunasekera, S.; Clark, R.J.; Lin, F.; Wade, J.D.; Anderson, M.A.; Craik, D.J. The N-terminal pro-domain of the kalata B1 cyclotide precursor is intrinsically unstructured. Biopolymers 2016, 106, 825–833. [Google Scholar] [CrossRef] [PubMed]
- Mulvenna, J.P.; Mylne, J.S.; Bharathi, R.; Burton, R.A.; Shirley, N.J.; Fincher, G.B.; Anderson, M.A.; Craik, D.J. Discovery of cyclotide-like protein sequences in graminaceous crop plants: Ancestral precursors of circular proteins? Plant Cell 2006, 18, 2134–2144. [Google Scholar] [CrossRef] [PubMed]
- Nahirñak, V.; Rivarola, M.; Gonzalez de Urreta, M.; Paniego, N.; Hopp, H.E.; Almasia, N.I.; Vazquez-Rovere, C. Genome-wide Analysis of the Snakin/GASA Gene Family in Solanum tuberosum cv. Kennebec. Am. J. Potato Res. 2016, 93, 172–188. [Google Scholar] [CrossRef]
- Kaur, J.; Fellers, J.; Adholeya, A.; Velivelli, S.L.S.; El-Mounadi, K.; Nersesian, N.; Clemente, T.; Shah, D. Expression of apoplast-targeted plant defensin MtDef4.2 confers resistance to leaf rust pathogen Puccinia triticina but does not affect mycorrhizal symbiosis in transgenic wheat. Transgenic Res. 2017, 26, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Menzel, L.P.; Chowdhury, H.M.; Masso-Silva, J.A.; Ruddick, W.; Falkovsky, K.; Vorona, R.; Malsbary, A.; Cherabuddi, K.; Ryan, L.K.; DiFranco, K.M.; et al. Potent in vitro and in vivo antifungal activity of a small molecule host defense peptide mimic through a membrane-active mechanism. Sci. Rep. 2017, 7, 4353. [Google Scholar] [CrossRef] [PubMed]
- Vriens, K.; Cammue, B.; Thevissen, K. Antifungal Plant Defensins: Mechanisms of Action and Production. Molecules 2014, 19, 12280–12303. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Hu, S.-H.; Martin, J.L.; Sjögren, T.; Hajdu, J.; Bohlin, L.; Claeson, P.; Göransson, U.; Rosengren, K.J.; Tang, J.; et al. Combined X-ray and NMR analysis of the stability of the cyclotide cystine knot fold that underpins its insecticidal activity and potential use as a drug scaffold. J. Biol. Chem. 2009, 284, 10672–10683. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.F.; Miranda, V.J.; Pinto, M.F.S.; Dohms, S.M.; Franco, O.L. High-performance computational analysis and peptide screening from databases of cyclotides from poaceae. Biopolymers 2016, 106, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Mylne, J.S.; Daly, N.L. Cyclotides: Macrocyclic peptides with applications in drug design and agriculture. Cell. Mol. Life Sci. 2010, 67, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Sousa, D.A.; Porto, W.F.; Silva, M.Z.; da Silva, T.R.; Franco, O.L. Influence of Cysteine and Tryptophan Substitution on DNA-Binding Activity on Maize α-Hairpinin Antimicrobial Peptide. Molecules 2016, 21, 1062. [Google Scholar] [CrossRef] [PubMed]
- Andorf, C.M.; Cannon, E.K.; Portwood, J.L.; Gardiner, J.M.; Harper, L.C.; Schaeffer, M.L.; Braun, B.L.; Campbell, D.A.; Vinnakota, A.G.; Sribalusu, V.V.; et al. MaizeGDB update: New tools, data and interface for the maize model organism database. Nucleic Acids Res. 2016, 44, D1195–D1201. [Google Scholar] [CrossRef] [PubMed]
- Walley, J.W.; Sartor, R.C.; Shen, Z.; Schmitz, R.J.; Wu, K.J.; Urich, M.A.; Nery, J.R.; Smith, L.G.; Schnable, J.C.; Ecker, J.R.; et al. Integration of omic networks in a developmental atlas of maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinforma. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.D.; Stephens, R.M. Sequence logos: A new way to display consensus sequences. Nucleic Acids Res. 1990, 18, 6097–6100. [Google Scholar] [CrossRef] [PubMed]
- Farhadpour, M.; Hashempour, H.; Talebpour, Z.; A-Bagheri, N.; Shushtarian, M.S.; Gruber, C.W.; Ghassempour, A. Microwave-assisted extraction of cyclotides from Viola ignobilis. Anal. Biochem. 2016, 497, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Hidden Markov models. Curr. Opin. Struct. Biol. 1996, 6, 361–365. [Google Scholar] [CrossRef]
- Berrocal-Lobo, M.; Segura, A.; Moreno, M.; López, G.; García-Olmedo, F.; Molina, A. Snakin-2, an antimicrobial peptide from potato whose gene is locally induced by wounding and responds to pathogen infection. Plant Physiol. 2002, 128, 951–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, P.W.R.; Yang, S.-H.; Molina, A.; López, G.; Middleditch, M.; Brimble, M.A. Plant antimicrobial peptides snakin-1 and snakin-2: Chemical synthesis and insights into the disulfide connectivity. Chemistry 2014, 20, 5102–5110. [Google Scholar] [CrossRef] [PubMed]
- Games, P.D.; DaSilva, E.Q.G.; de Oliveira Barbosa, M.; Almeida-Souza, H.O.; Fontes, P.P.; DeMagalhães, M.J.; Pereira, P.R.G.; Prates, M.V.; Franco, G.R.; Faria-Campos, A.; et al. Computer aided identification of a Hevein-like antimicrobial peptide of bell pepper leaves for biotechnological use. BMC Genomics 2016, 17, 999. [Google Scholar] [CrossRef] [PubMed]
- Epple, P.; Apel, K.; Bohlmann, H. An Arabidopsis thaliana thionin gene is inducible via a signal transduction pathway different from that for pathogenesis-related proteins. Plant Physiol. 1995, 109, 813–820. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize Gene ID | Maize Chromosome | BLAST From Seq | Origin |
|---|---|---|---|
| Hevein | |||
| GRMZM2G005633P1 | Ch10 | ctb1-chitinase B1 | Beta vulgaris |
| GRMZM2G005633P2 | Ch10 | ctb1-chitinase B1 | Beta vulgaris |
| GRMZM2G373106 | Ch8 | ctb1-chitinase B1 | Beta vulgaris |
| GRMZM2G117942 | Ch4 | Q8H950 | Eutrema wasabi |
| GRMZM2G145518 | Ch6 | Q8H950 | Eutrema wasabi |
| GRMZM2G051921 | Ch2 | Q8H950 | Eutrema wasabi |
| GRMZM2G051943 | Ch2 | cta1-chitinase | Beta vulgaris |
| GRMZM2G117971 | Ch4 | Q8H950 | Eutrema wasabi |
| Snakin | |||
| GRMZM2G105364 | Ch9 | Q0VYL5 | Fagus sylvatica |
| GRMZM2G068202 | Ch2 | Q19VG5 | Zea mays |
| GRMZM2G117940 | Ch1 | O49134 | Fragaria ananassa |
| GRMZM2G172596 | Ch10 | Q0VYL5 | Fagus sylvatica |
| GRMZM2G062527P1 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G062527P2 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G062527P3 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G062527P4 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G062527P5 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G077034 | Ch5 | O49134 | Fragaria ananassa |
| GRMZM2G164090 | Ch6 | Q19VG5 | Zea mays |
| GRMZM2G107003 | Ch2 | Q19VG5 | Zea mays |
| LTP | |||
| GRMZM2G010868P1 | Ch3 | Q2XX14 | Zea mays |
| GRMZM2G010868P2 | Ch3 | Q2XX14 | Zea mays |
| GRMZM2G101958 | Ch10 | Q2XX14 | Zea mays |
| GRMZM5G898755P1 | Ch10 | Q2XX14 | Zea mays |
| GRMZM5G898755P2 | Ch10 | Q2XX14 | Zea mays |
| GRMZM2G107839P1 | Ch3 | Q2XX25 | Zea mays |
| GRMZM2G107839P2 | Ch3 | Q2XX25 | Zea mays |
| Defensin | |||
| GRMZM2G368890 | Ch10 | P81008 | Zea mays |
| GRMZM2G392863P1 | Ch1 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM2G392863P2 | Ch1 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM2G146809 | Ch10 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM5G896902 | Ch2 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM2G153488P1 | Ch5 | P82781 | Arabidopsis thaliana |
| GRMZM2G153488P2 | Ch5 | P82781 | Arabidopsis thaliana |
| GRMZM2G153488P3 | Ch5 | P82781 | Arabidopsis thaliana |
| GRMZM2G064698P1 | Ch2 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM2G064698P2 | Ch2 | Q9ZUL7 | Arabidopsis thaliana |
| GRMZM2G153368P1 | Ch5 | P82781 | Arabidopsis thaliana |
| GRMZM2G153368P2 | Ch5 | P82781 | Arabidopsis thaliana |
| GRMZM2G368861P1 | Ch10 | P81008 | Zea mays |
| GRMZM2G368861P2 | Ch10 | P81008 | Zea mays |
| GRMZM2G046532 | Ch7 | P81008 | Zea mays |
| Beta-Barrelin | |||
| GRMZM2G430500 | Ch2 | P80915 | Macadamia integrifolia |
| Cyclotide | |||
| GRMZM2G032198 | Ch3 | P56879 | Chassalia parviflora |
| GRMZM2G374405 | Ch3 | P56879 | Chassalia parviflora |
| GRMZM2G450866 | Ch7 | P85233 | Viola hederacea |
| Maize Genes | Primers |
|---|---|
| cDNA Primers | |
| Hevein | |
| GRMZM2G117942F | TACATCGATCGGTTGCCAAA |
| GRMZM2G117942R | TTCTGCTGCGGGTTGTAGA |
| GRMZM2G145518F | TTCTCCAAGCACAGGAGACA |
| GRMZM2G145518R | ACGCCTCACTTCCCACTGTA |
| GRMZM2G117971F | TATGGATGTGATCCCACACG |
| GRMZM2G117971R | AGTGGACGACACATATTCGAGA |
| Snakin | |
| GRMZM2G105364F | TGGAATGCTACCAGCCAGAT |
| GRMZM2G105364R | CGGGATGTTCCTCATCAATC |
| GRMZM2G172596F | CTGCTCCTCTGCTTCCTGTT |
| GRMZM2G172596R | GTTCTTGTAGCCCTCGTGCTT |
| GRMZM2G107003F | CGCCACGTTTTGTATGATCC |
| GRMZM2G107003R | ACACAGACCCATCAACGTCA |
| LTP | |
| GRMZM2G101958F | CATATGTGACCGTGTGTTCCA |
| GRMZM2G101958R | CTCGCCCAGCTTTGTTTTAT |
| GRMZM2G010868P1F | TTGGCACCAAGCACTAAAGA |
| GRMZM2G010868P1R | TCCCAAATCATCCCCTAGAA |
| GRMZM2G010868P2F | CCTGCAACTGCCTCAAGAAC |
| GRMZM2G010868P2R | TGCATGCATACTACCCTACCTG |
| GRMZM5G898755P1F | TCGACTGCACCAAGATCAAC |
| GRMZM5G898755P1R | TCTGATGCATGACACACACG |
| GRMZM5G898755P2F | AGCAGCACCTCAATGTCCTT |
| GRMZM5G898755P2R | CATGCATATGTACGGCGAAT |
| GRMZM2G107839P1F | CTCCGGTTTGCAGAAACAAC |
| GRMZM2G107839P1R | CTAGGCATCAGCACAGTCCA |
| GRMZM2G107839P2F | GATCCACCTACTTGTTCAGACAG |
| GRMZM2G107839P2R | CATCTCCTCTGATCGTCCTTT |
| Defensin | |
| GRMZM2G368890F | GCCGGAATATGTGGACGAT |
| GRMZM2G368890R | ACATGCAGACCCCCTTGAA |
| GRMZM2G392863P1F | TGTTGTACGTACGTCTGCCTCT |
| GRMZM2G392863P1R | AACAATCAGCGTCGTCTCTT |
| GRMZM2G392863P2F | CCGCTGAGATCCTAGGAAGA |
| GRMZM2G392863P2R | CTGATGAGTCCACAGCACAGA |
| GRMZM2G146809F | GGTCCGTTTGCGTTTGTTTC |
| GRMZM2G146809R | GGTTCATCAATGCAACGAGAC |
| GRMZM5G896902F | AGAAGGACAGCGAGCGATT |
| GRMZM5G896902R | CCGGGAGTAGGTTAATTTAGCA |
| GRMZM2G153488P1F | GTTGTACTTTCTGCATCCGTTG |
| GRMZM2G153488P1R | TTGGTCATCAAGTTCCCTAGC |
| GRMZM2G153488P2F | AGCCTTACGTAGCGAAGCTC |
| GRMZM2G153488P2R | AGCAACGAGGAGTTGAGTCG |
| GRMZM2G153488P3F | ATAAACCGTGGCTCTGGTTC |
| GRMZM2G153488P3R | TTGCTCTGAGCTTCGCTACG |
| GRMZM2G064698P1F | AGTTCGTGAATCCCTGAAGC |
| GRMZM2G064698P1R | ATTCCCTTGCCTGTGCCATA |
| GRMZM2G368861P1F | GATAGTGACGTACGCGCAAC |
| GRMZM2G368861P1R | GCATACGATCTGACGCTCAT |
| GRMZM2G368861P2F | GCGATGGAGCTCATCAAGTC |
| GRMZM2G368861P2R | GTCCATGAGGCAGCAGAAAT |
| GRMZM2G046532F | GGTGCCCATACCATAGCTTC |
| GRMZM2G046532R | TAACAAACGAGCAGGAGGAG |
| GRMZM2G064698P2F | AGTTCGTGAATCCCTGAAGC |
| GRMZM2G064698P2R | ATTCCCTTGCCTGTGCCATA |
| GRMZM2G153368P1F | AAGAAGCCTTGCTAGTTCATCG |
| GRMZM2G153368P1R | CCCAGCAATTTAAGGACTGC |
| GRMZM2G153368P2F | GTACGTACTCGTACCAGGCAGA |
| GRMZM2G153368P2R | GCATGGCTACTCCCATTTTG |
| β-Barrelin | |
| GRMZM2G430500F | CTCGGGGGATACGTCGAT |
| GRMZM2G430500R | TGGGTGTCCTCGAAAACTTG |
| Cyclotide | |
| GRMZM2G032198F | GTGTTTGGCCTGGACTTCAT |
| GRMZM2G032198R | GGCGTCACGAGTTTATTTCA |
| GRMZM2G374405F | GTCCCCTGTTTTGAATCCTG |
| GRMZM2G374405R | TTCACACGTAACGGGATCAG |
| GRMZM2G450866F | GGGCTTGTTGCAGTGGTAGT |
| GRMZM2G450866R | CGATCTTGTGACGGTTCAGC |
| Genomic DNA Primers | |
| Defensin | |
| GRMZM2G146809F2 | GGCCAAGTATACTCGCCAGA |
| GRMZM2G146809R2 | TCGAAGGGTTATTGCATTCC |
| Cyclotide | |
| GRMZM2G032198F2 | GTTGGGAGCAAAGCAAAGAG |
| GRMZM2G032198R2 | GAGGAGCAGGCGATTGAGTA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noonan, J.; Williams, W.P.; Shan, X. Investigation of Antimicrobial Peptide Genes Associated with Fungus and Insect Resistance in Maize. Int. J. Mol. Sci. 2017, 18, 1938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091938
Noonan J, Williams WP, Shan X. Investigation of Antimicrobial Peptide Genes Associated with Fungus and Insect Resistance in Maize. International Journal of Molecular Sciences. 2017; 18(9):1938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091938
Chicago/Turabian StyleNoonan, Joseph, William Paul Williams, and Xueyan Shan. 2017. "Investigation of Antimicrobial Peptide Genes Associated with Fungus and Insect Resistance in Maize" International Journal of Molecular Sciences 18, no. 9: 1938. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091938