Non-Clinical Studies for Evaluation of 8-C-Rhamnosyl Apigenin Purified from Peperomia obtusifolia against Acute Edema

 and
and 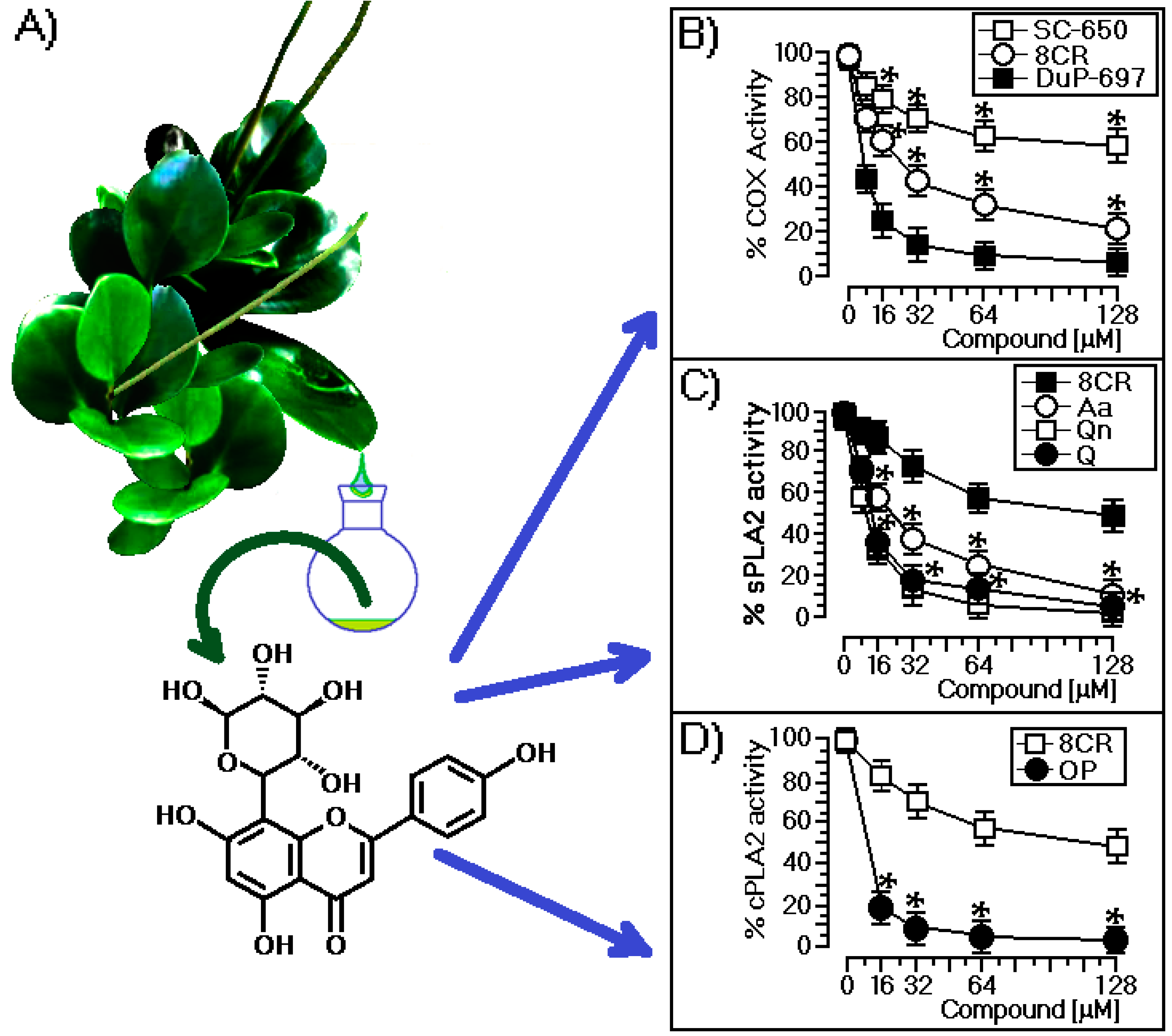{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
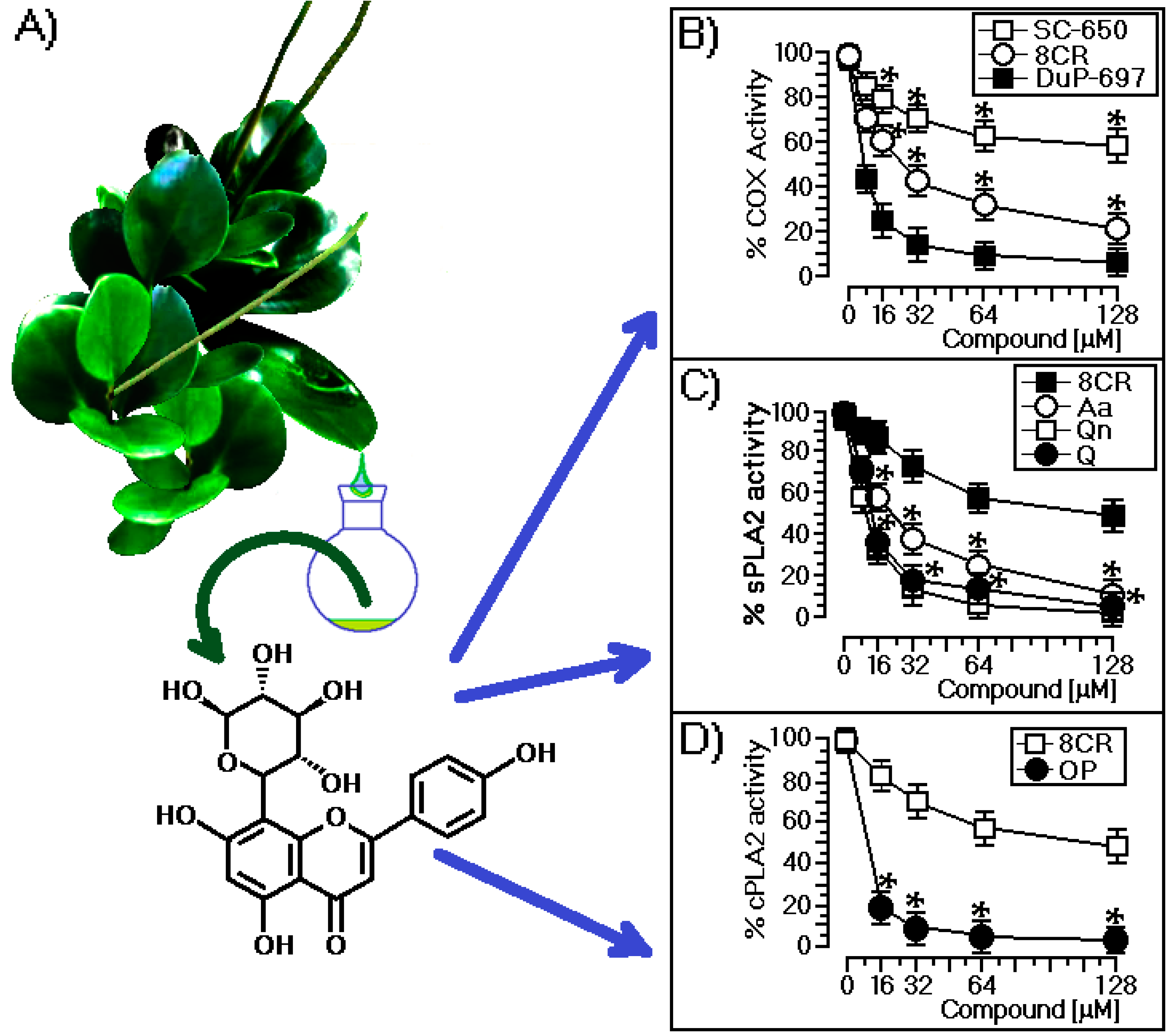
2.1. Structural and Biological Characterization of 8-C-Rhamnosyl Apigenin (8CR)
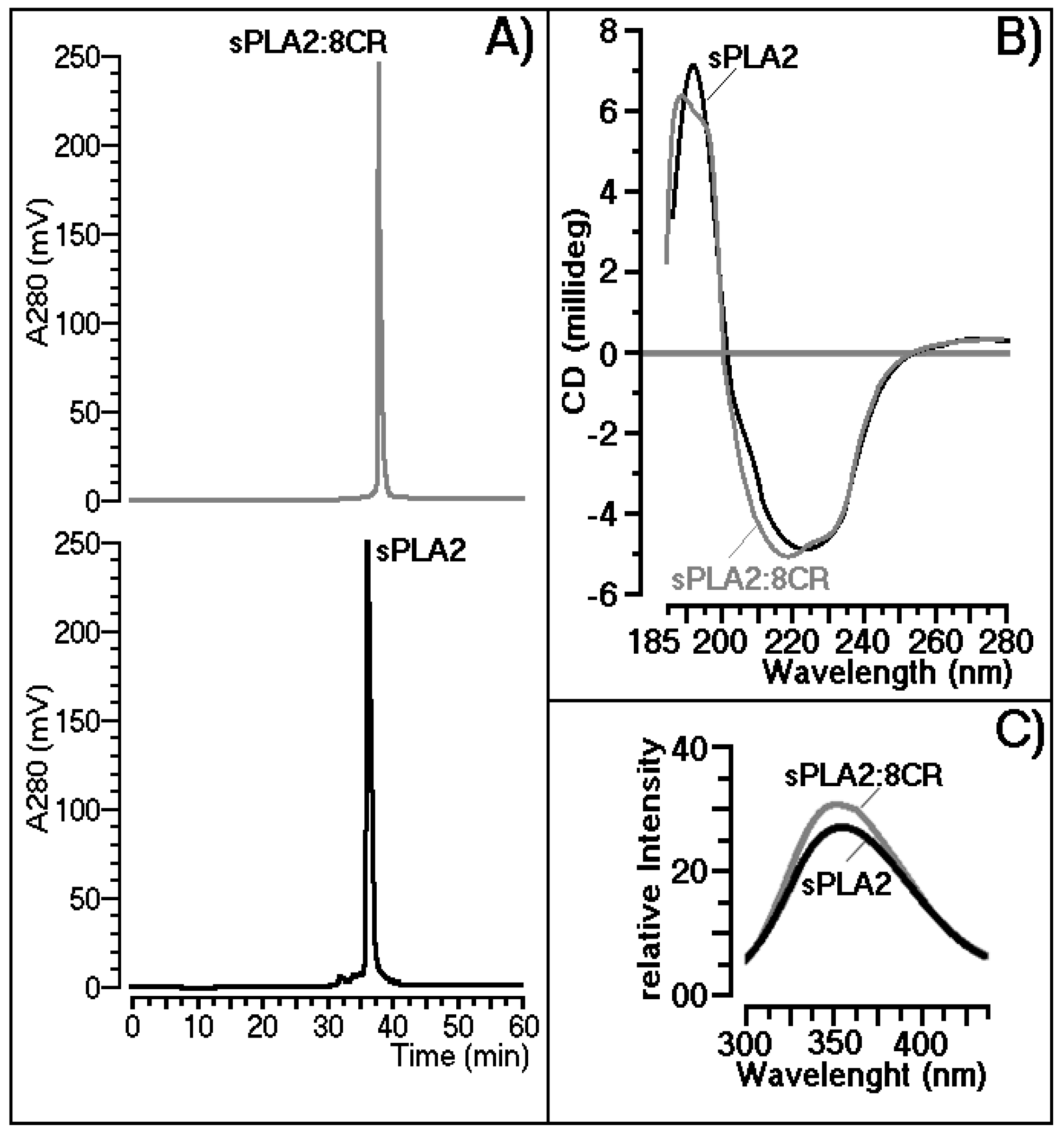
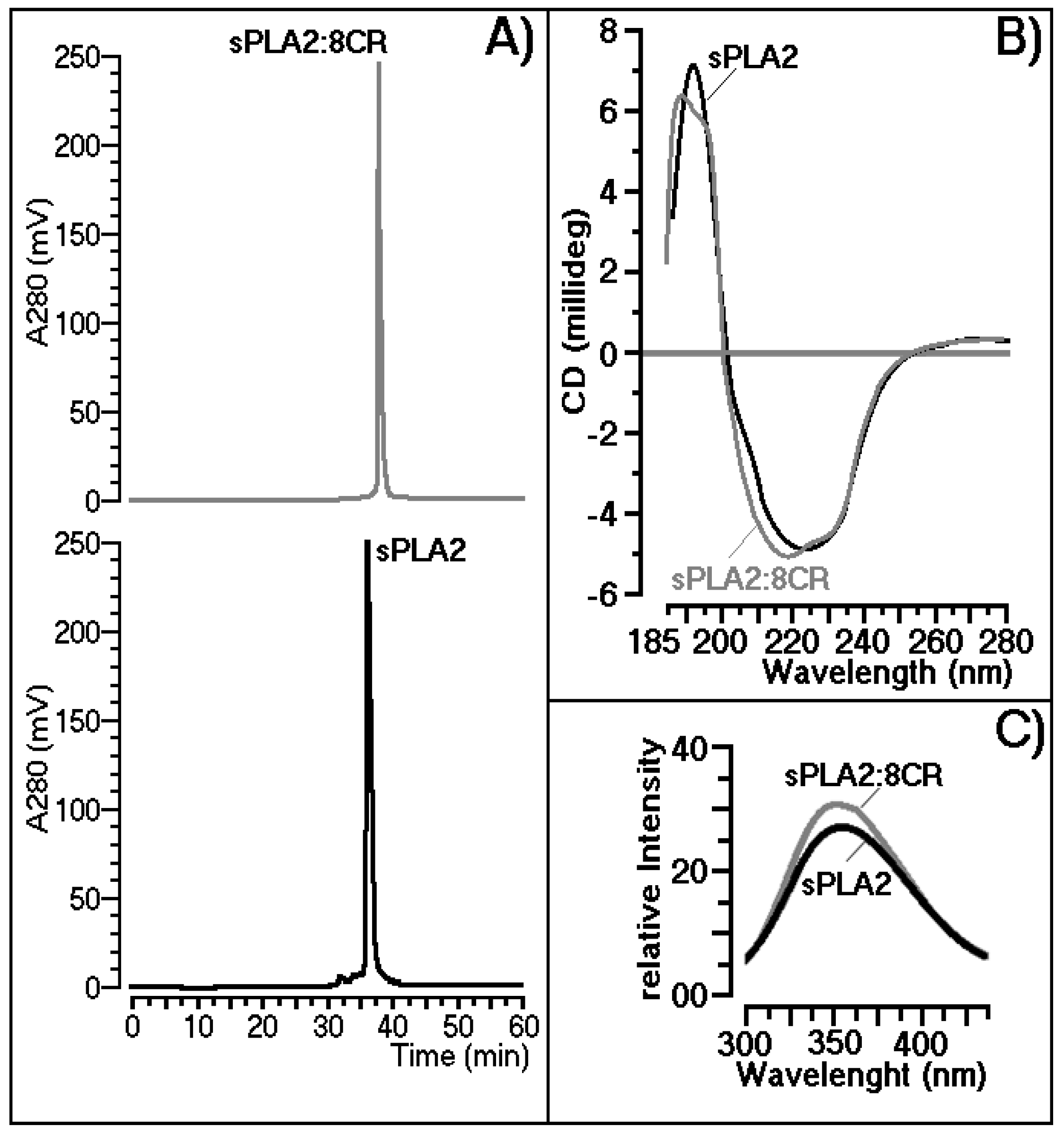
2.2. Structural Shifts Induced by 8CR on the sPLA2 Molecule
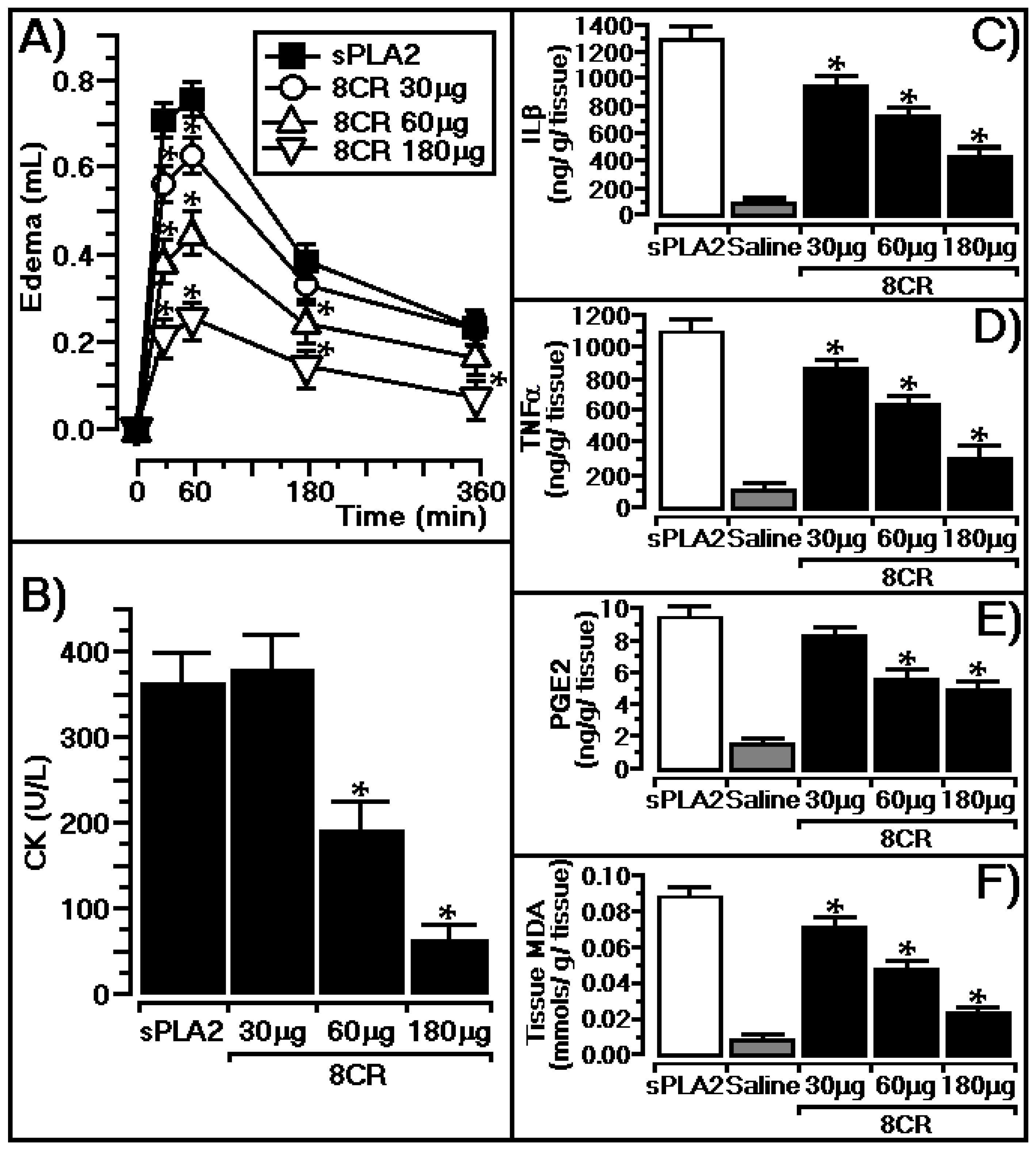
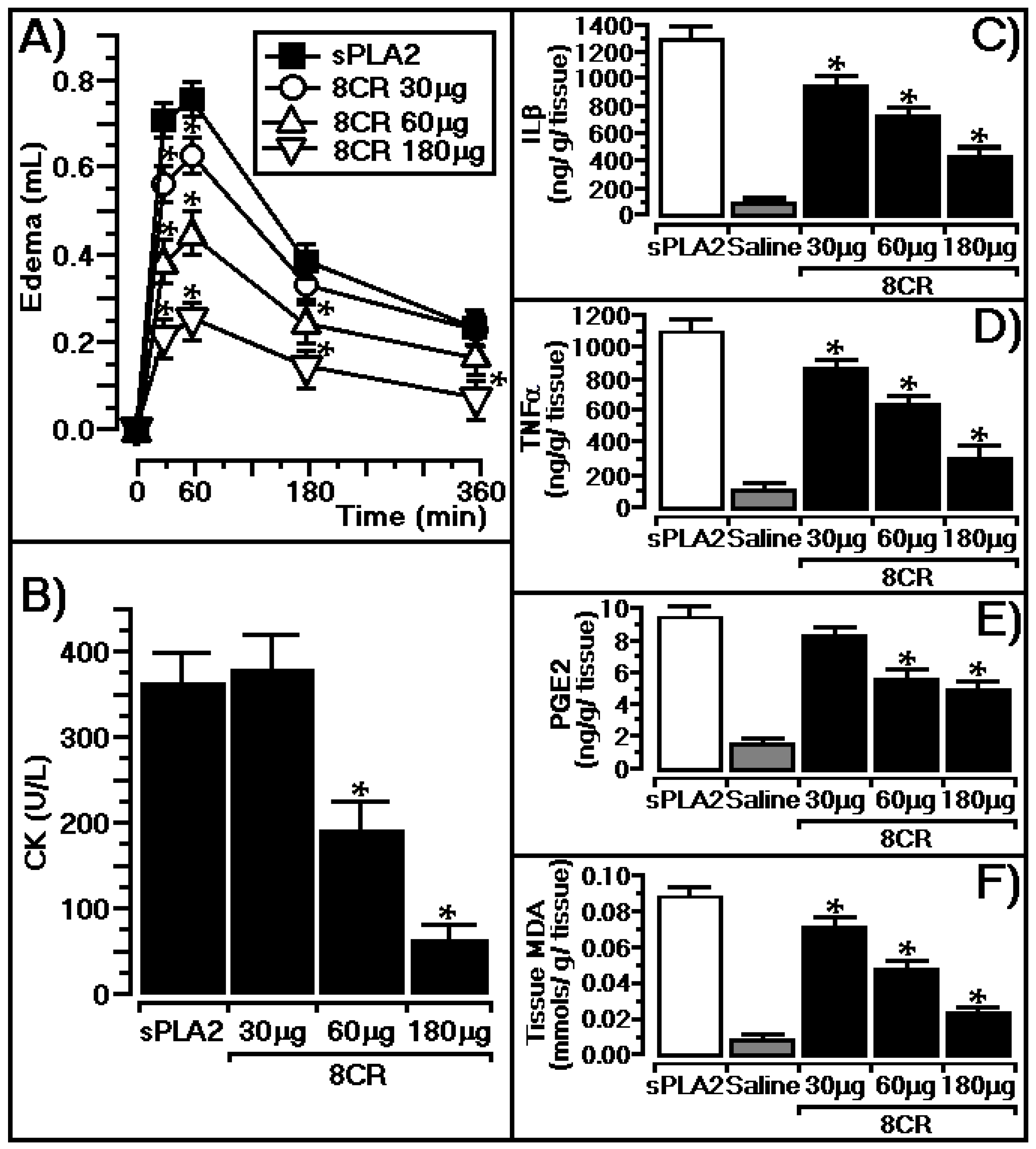
2.3. Protective Effect of 8CR against the Pharmacological Effects of sPLA2
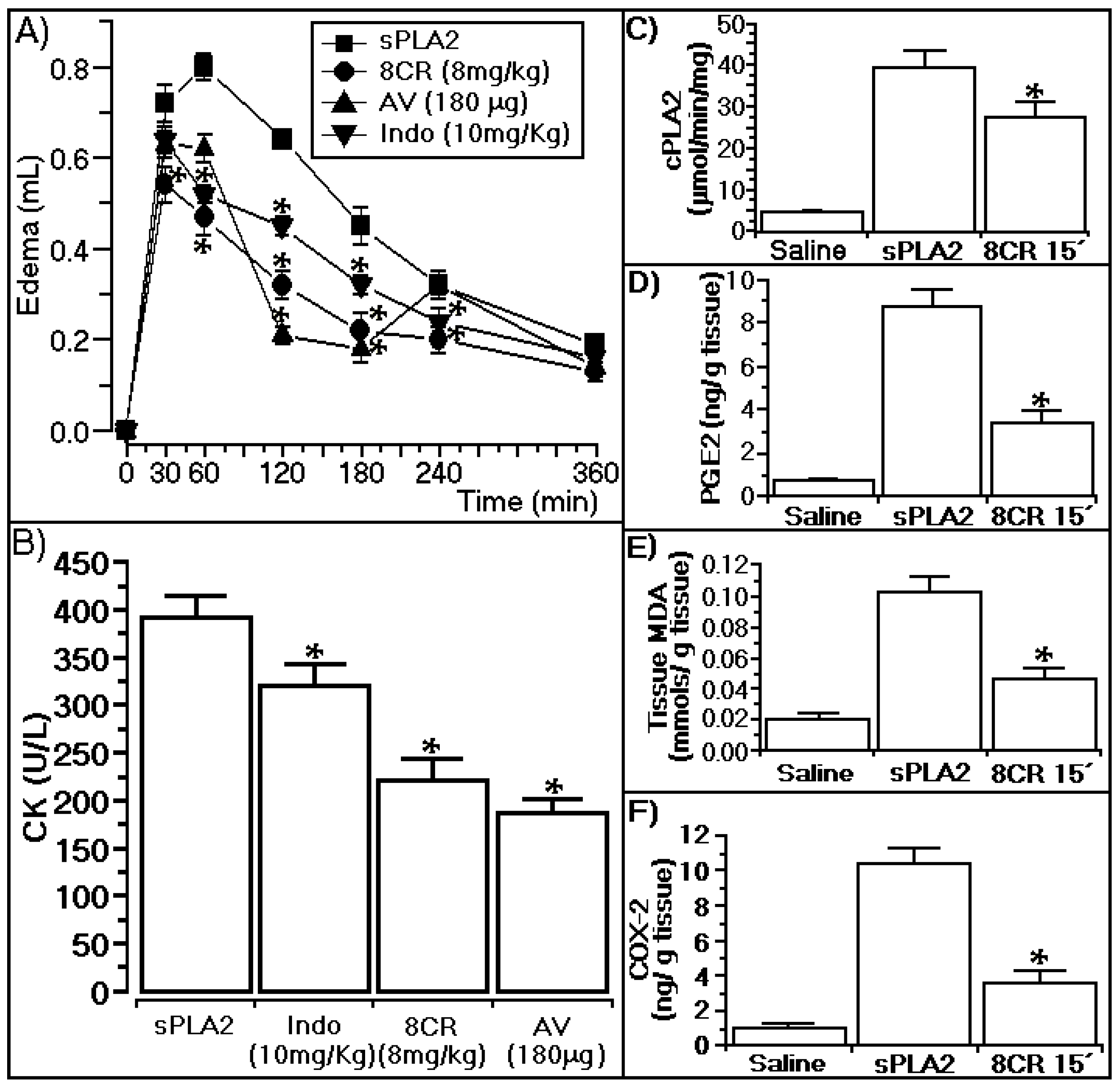
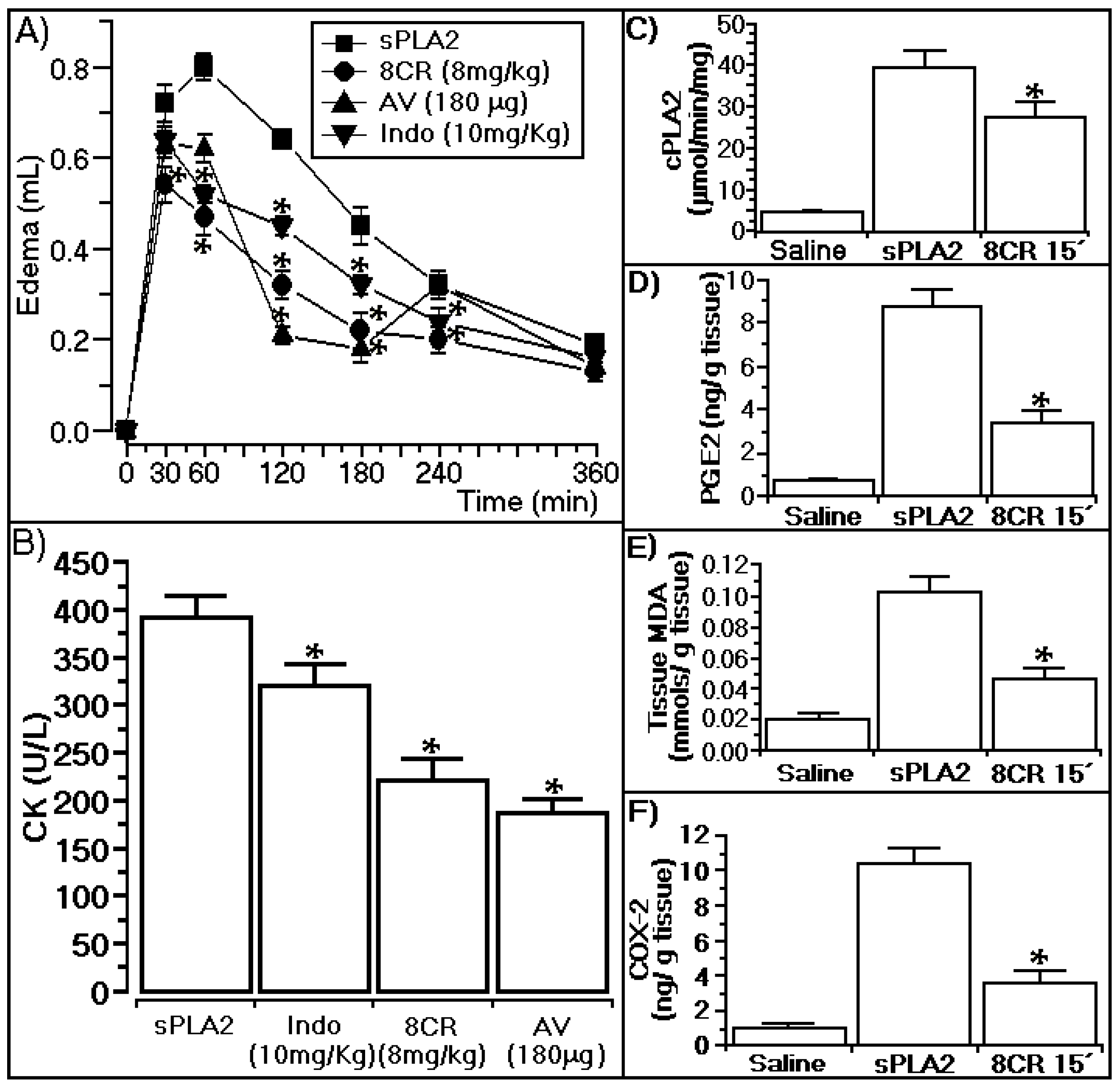
2.4. Antiophidian Activity Effect of 8CR against the Pharmacological Effects of sPLA2
3. Discussion
3.1. Structural and Biological Characterization of 8CR
3.2. Structural Shifts Induced by 8CR on the sPLA2 Molecule
3.3. In Vivo Protective Effect of 8CR against the Pharmacological Effects of sPLA2
3.4. Anti-Ophidian Effect of 8CR against the Pharmacological Effects of sPLA2
4. Materials and Methods
4.1. Materials and General Experimental Procedures
4.2. Extraction and Purification of 8CR
4.3. Purification of Secretory Phospholipase A2
4.4. Cytosolic PLA2, sPLA2, and COX-1 and -2
4.5. Pharmacological Assay and Biochemical Assays
4.5.1. Paw Edema
4.5.2. Evaluation of Myonecrosis
4.5.3. Determination of IL-1β, TNF-α, Prostaglandin E2 (PGE-2), and Malondialdehyde (MDA) Levels in Mice Paws
4.6. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SC-560 | 5-(4-Chlorophenyl)-1-(4-methoxyphenyl)-3-(trifluoromethyl)-1H-pyrazole) |
| 8CR | 8-C-Rhamnosyl apigenin |
| cPLA2 | Citossolic phospholipase A2 |
| sPLA2 | Secretory phospholipase A2 |
| DuP-697 | 5-Bromo-2-(4-fluorophenyl)-3-(4-(methylsulfonyl)phenyl)-thiophene) |
| Aa | Aristolochic acid |
| CK | Ceratine kinase |
| Cdt | Crotalus durissus terrificus |
| CD | Circular dichroism |
| IL-1β | Interleukin 1β |
| PG | Prostaglandins |
| COX | Cyclooxygenase |
| AA | Arachidonic acid |
| OP | Oleyloxyethyl phosphorylcholine |
| SEM | Standard Error of Mean |
References
- Marcussi, S.; Sant’Ana, C.D.; Oliveira, C.Z.; Rueda, A.Q.; Menaldo, D.L.; Beleboni, R.O.; Stabeli, R.G.; Giglio, J.R.; Fontes, M.R.; Soares, A.M. Snake venom phospholipase A2 inhibitors: Medicinal chemistry and therapeutic potential. Curr. Top. Med. Chem. 2007, 7, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Perumal Samy, R.; Gopalakrishnakone, P.; Chow, V.T. Therapeutic application of natural inhibitors against snake venom phospholipase A2. Bioinformation 2012, 8, 48–57. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, L.; Cintra, A.C.; Veronese, E.L.; Nomizo, A.; Franco, J.J.; Arantes, E.C.; Giglio, J.R.; Sampaio, S.V. Anticrotalic and antitumoral activities of gel filtration fractions of aqueous extract from Tabernaemontana catharinensis (Apocynaceae). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 137, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, S.C.; Hyslop, S.; Fontes, M.R.; Prado-Franceschi, J.; Zambelli, V.O.; Magro, A.J.; Brigatte, P.; Gutierrez, V.P.; Cury, Y. Crotoxin: Novel activities for a classic β-neurotoxin. Toxicon 2010, 55, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Exeter, H.J.; Folkersen, L.; Palmen, J.; Franco-Cereceda, A.; Cooper, J.A.; Kalea, A.Z.; Hooft, F.V.; Eriksson, P.; Humphries, S.E.; Talmud, P.J. Functional analysis of two PLA2G2A variants associated with secretory phospholipase A2-IIA levels. PLoS ONE 2012, 7, e41139. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, B.S.; Fawzy, A.A.; Franson, R.C. Edema-inducing activity of phospholipase A2 purified from human synovial fluid and inhibition by aristolochic acid. Inflammation 1988, 12, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Baranowska-Bosiacka, I.; Gutowska, I.; Chlubek, D. The effect of reactive oxygen species on the synthesis of prostanoids from arachidonic acid. J. Physiol. Pharmacol. 2013, 64, 409–421. [Google Scholar] [PubMed]
- Sun, G.Y.; Chuang, D.Y.; Zong, Y.; Jiang, J.; Lee, J.C.M.; Gu, Z.; Simonyl, A. Role of cytosolic phospholipase A2 in oxidative and inflammatory signaling pathways in different cell types in the central nervous system. Mol. Neurobiol. 2014, 50, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Velozo, L.S.; Ferreira, M.J.; Santos, M.I.; Moreira, D.L.; Emerenciano, V.P.; Kaplan, M.A. Unusual chromenes from Peperomia blanda. Phytochemistry 2006, 67, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Velozo, L.S.; Ferreira, M.J.; Santos, M.I.; Moreira, D.L.; Guimarães, E.F.; Emerenciano, V.P.; Kaplan, M.A. C-glycosyl flavones from Peperomia blanda. Fitoterapia 2009, 80, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Z.; Tong, A.P.; Huang, J. Two New Norlignans and a New Lignanamide from Peperomia tetraphylla. Chem. Biodivers. 2012, 9, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Da Silva Mota, J.; Leite, A.C.; Junior, J.M.B.; López, S.N.; Ambrósio, D.L.; Passerini, G.D.; Kato, M.J.; da Silva Bolzani, V.; Barretto Cicarelli, R.M.; Furlan, M. In Vitro trypanocidal activity of phenolic derivatives from Peperomia obtusifolia. Planta Med. 2009, 75, 620–623. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Asai, F.; Iinuma, M. Phenolic compounds from Peperomia obtusifolia. Phytochemistry 1998, 49, 229–232. [Google Scholar] [CrossRef]
- Wang, Q.W.; Yu, D.H.; Lin, M.G.; Zhao, M.; Zhu, W.J.; Lu, Q.; Li, G.X.; Wang, C.; Yang, Y.F.; Qin, X.M.; et al. Antiangiogenic polyketides from Peperomia dindygulensis Miq. Molecules 2012, 17, 4474–4483. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, E.A.; Reigada, J.B.; Correia, M.V.; Young, M.C.; Guimarães, E.F.; Franchi, G.C., Jr.; Nowill, A.E.; Lago, J.H.; Yamaguchi, L.F.; Kato, M.J. Antifungal and cytotoxic 2-acylcyclohexane-1,3-diones from Peperomia alata and P. trineura. J. Nat. Prod. 2014, 77, 1377–1382. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.M., Jr.; Batista, A.N.; Mota, J.S.; Cass, Q.B.; Kato, M.J.; Bolzani, V.S.; Freedman, T.B.; López, S.N.; Furlan, M.; Nafie, L.A. Structure elucidation and absolute stereochemistry of isomeric monoterpene chromane esters. J. Org. Chem. 2011, 76, 2603–2612. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.M., Jr.; Batista, A.N.L.; Kato, M.J.; Bolzani, V.S.; López, S.N.; Laurence, A.; Nafie, L.A.; Furlan, M. Further monoterpene chromane esters from Peperomia obtusifolia: VCDdetermination of the absolute configuration of a new diastereomeric mixture. Tetrahedron Lett. 2012, 53, 6051–6054. [Google Scholar] [CrossRef]
- Batista, A.N.L.; Santos-Pinto, J.R.A.D.; Batista, J.M., Jr.; Souza-Moreira, T.M.; Santoni, M.M.; Zanelli, C.F.; Kato, M.J.; López, S.N.; Palma, M.S.; Furlan, M. The Combined Use of Proteomics and Transcriptomics Reveals a Complex Secondary Metabolite Network in Peperomia obtusifolia. J. Nat. Prod. 2017, 80, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Cotrim, C.A.; de Oliveira, S.C.; Diz Filho, E.B.; Fonseca, F.V.; Baldissera, L., Jr.; Antunes, E.; Ximenes, R.M.; Monteiro, H.S.; Rabello, M.M.; Hernandes, M.Z.; et al. Quercetin as an inhibitor of snake venom secretory phospholipase A2. Chem. Biol. Interact. 2011, 189, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, P.K. Carbone-13 NMR of Flavonoids; Elsevier Science Publishing Co. Inc.: New York, NY, USA, 1989. [Google Scholar]
- Pires, C.L.; Rodrigues, S.D.; Bristot, D.; Gaeta, H.; Toyama, D.; Lobo-Farias, W.; Toyama, M. Sulfated polysaccharide extracted of the green algae Caulerpa racemosa increase the enzymatic activity and paw edema induced by sPLA2 from Crotalus durissus terrificus venom. Brazilian. J. Pharmacogn. 2013, 23, 635–645. [Google Scholar] [CrossRef]
- Moreira, V.; Gutiérrez, J.M.; Soares, A.M.; Zamunér, S.R.; Purgatto, E.; de Teixeira, C.F. Secretory phospholipases A2 isolated from Bothrops asper and from Crotalus durissus terrificus snake venoms induce distinct mechanisms for biosynthesis of prostaglandins E2 and D2 and expression of cyclooxygenases. Toxicon 2008, 52, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Andrade, E.L.; Bento, A.F.; Cavalli, J.; Oliveira, S.K.; Freitas, C.S.; Marcon, R.; Schwanke, R.C.; Siqueira, J.M.; Calixto, J.B. Non-clinical studies required for new drug development—Part I: Early in silico and in vitro studies, new target discovery and validation, proof of principles and robustness of animal studies. Br. J. Med. Biol. Res. 2016, 49, e5644. [Google Scholar] [CrossRef] [PubMed]
- Johansson, J.U.; Woodling, N.S.; Shi, J.; Andreasson, K. Inflammatory cyclooxygenase activity and PGE-2 signaling in models of Alzheimer’s disease. Curr. Immunol. Rev. 2015, 11, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.W.; Misso, N.; Aggarwal, S.; Thompson, P.; Walters, E.H. Oxidative stress and lipid-derived inflammatory mediators during acute exacerbations of cystic fibrosis. Respirology 2007, 12, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Elmarakby, A.A.; Sullivan, J.C. Relationship between oxidative stress and inflammatory cytokines in diabetic nephropathy. Cardiovasc. Ther. 2012, 30, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Tovar-Gijón, C.E.; Hernández-Carlos, B.; Burgueño-Tapia, E.; Cedillo-Portugal, E.; Joseph-Nathan, P. A new C-Glycosylflavone from Encyclia michuacana. J. Mol. Struct. 2006, 783, 96–100. [Google Scholar] [CrossRef]
- Roque, A.T.; Gambeloni, R.Z.; Felitti, S.; Ribeiro, M.L.; Santos, J.C. Inflammation-induced oxidative stress in breast cancer patients. Med. Oncol. 2015, 32, 263. [Google Scholar] [CrossRef] [PubMed]
- Toyama, M.H.; de Oliveira, D.G.; Beriam, L.O.; Novello, J.C.; Rodrigues-Simioni, L.; Marangoni, S. Structural, enzymatic and biological properties of new PLA2 isoform from Crotalus durissus terrificus venom. Toxicon 2003, 41, 1033–1038. [Google Scholar] [CrossRef]
- Toyama, D.O.; Ferreira, M.J.; Romoff, P.; Fávero, O.A.; Gaeta, H.H.; Toyama, M.H. Effect of chlorogenic acid (5-Caffeoylquinic acid) isolated from Baccharis oxyodonta on the structure and pharmacological activities of secretory phospholipase A2 from Crotalus durissus terrificus. Biomed. Res. Int. 2014, 2014, 726585. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamayose, C.I.; Romoff, P.; Toyama, D.O.; Gaeta, H.H.; Costa, C.R.C.; Belchor, M.N.; Ortolan, B.D.; Velozo, L.S.M.; Kaplan, M.A.C.; Ferreira, M.J.P.; et al. Non-Clinical Studies for Evaluation of 8-C-Rhamnosyl Apigenin Purified from Peperomia obtusifolia against Acute Edema. Int. J. Mol. Sci. 2017, 18, 1972. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091972
Tamayose CI, Romoff P, Toyama DO, Gaeta HH, Costa CRC, Belchor MN, Ortolan BD, Velozo LSM, Kaplan MAC, Ferreira MJP, et al. Non-Clinical Studies for Evaluation of 8-C-Rhamnosyl Apigenin Purified from Peperomia obtusifolia against Acute Edema. International Journal of Molecular Sciences. 2017; 18(9):1972. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091972
Chicago/Turabian StyleTamayose, Cinthia I., Paulete Romoff, Daniela O. Toyama, Henrique H. Gaeta, Caroline R. C. Costa, Mariana N. Belchor, Bruna D. Ortolan, Leosvaldo S. M. Velozo, Maria A. C. Kaplan, Marcelo J. P. Ferreira, and et al. 2017. "Non-Clinical Studies for Evaluation of 8-C-Rhamnosyl Apigenin Purified from Peperomia obtusifolia against Acute Edema" International Journal of Molecular Sciences 18, no. 9: 1972. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091972