Identification of Circulating miRNAs Differentially Regulated by Opioid Treatment

Abstract
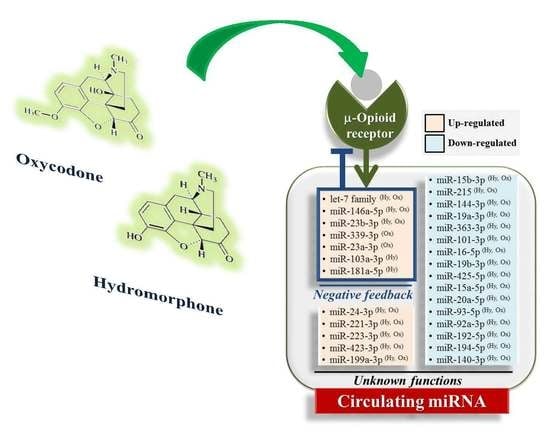
:
1. Introduction
2. Results
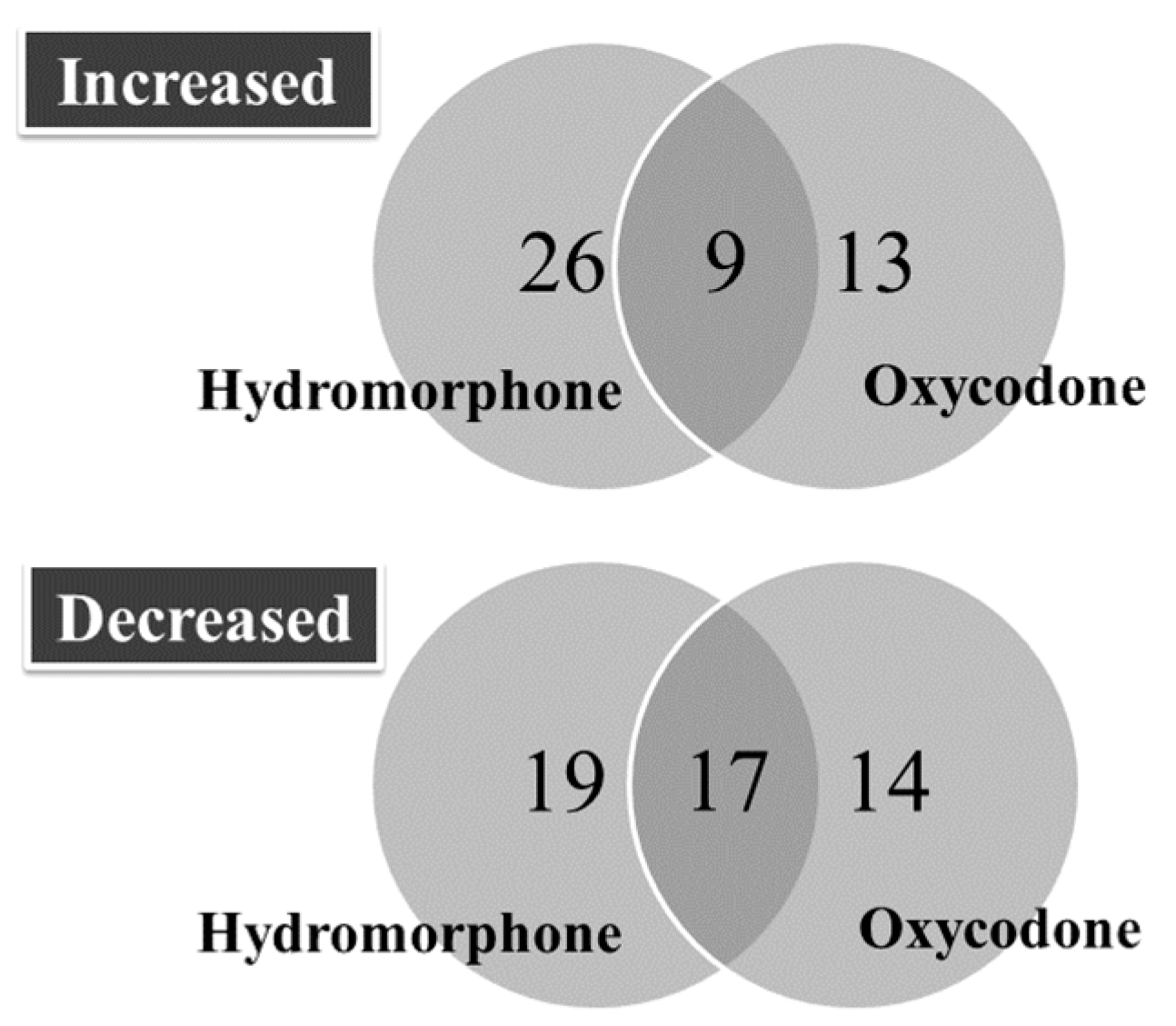
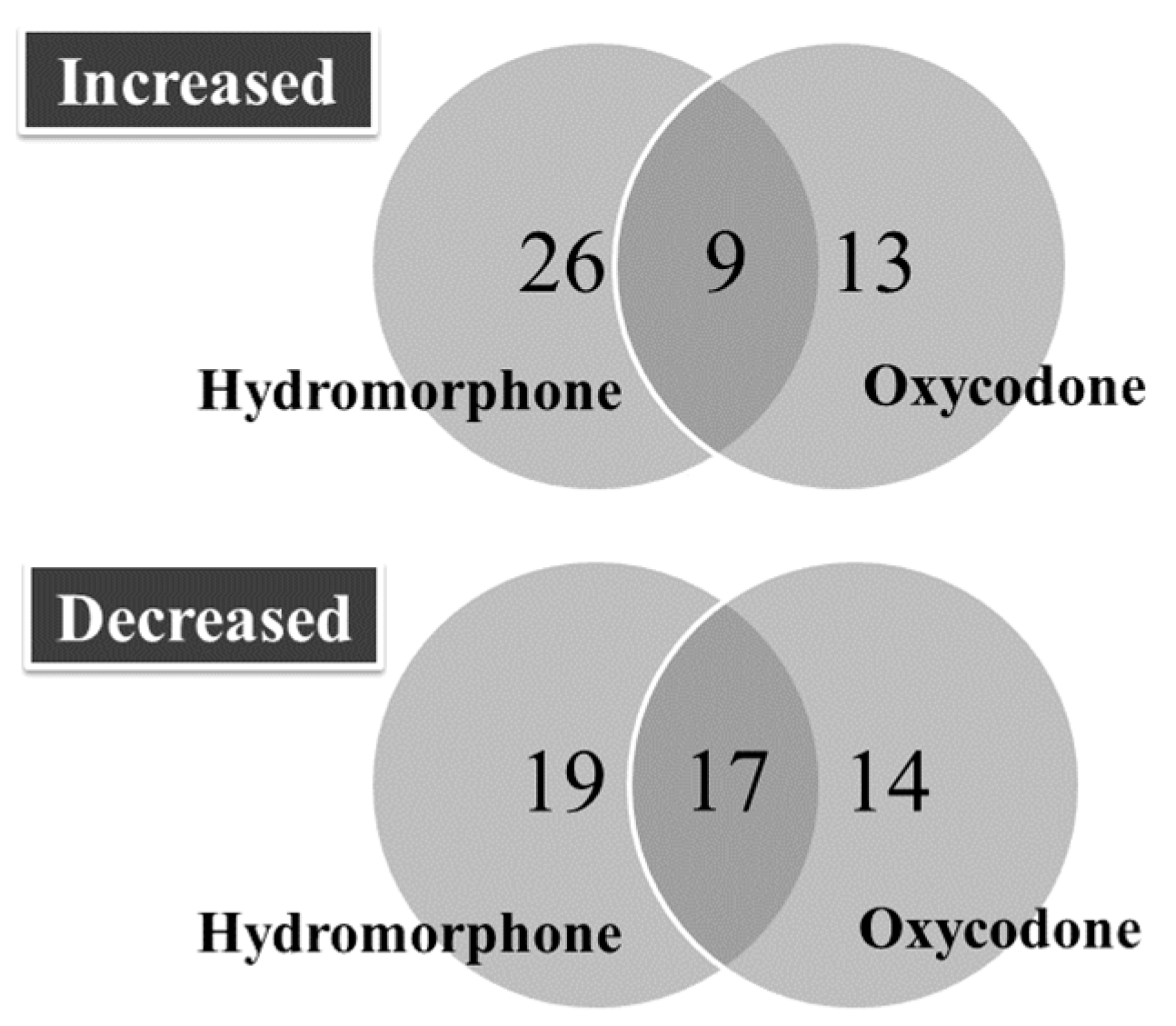
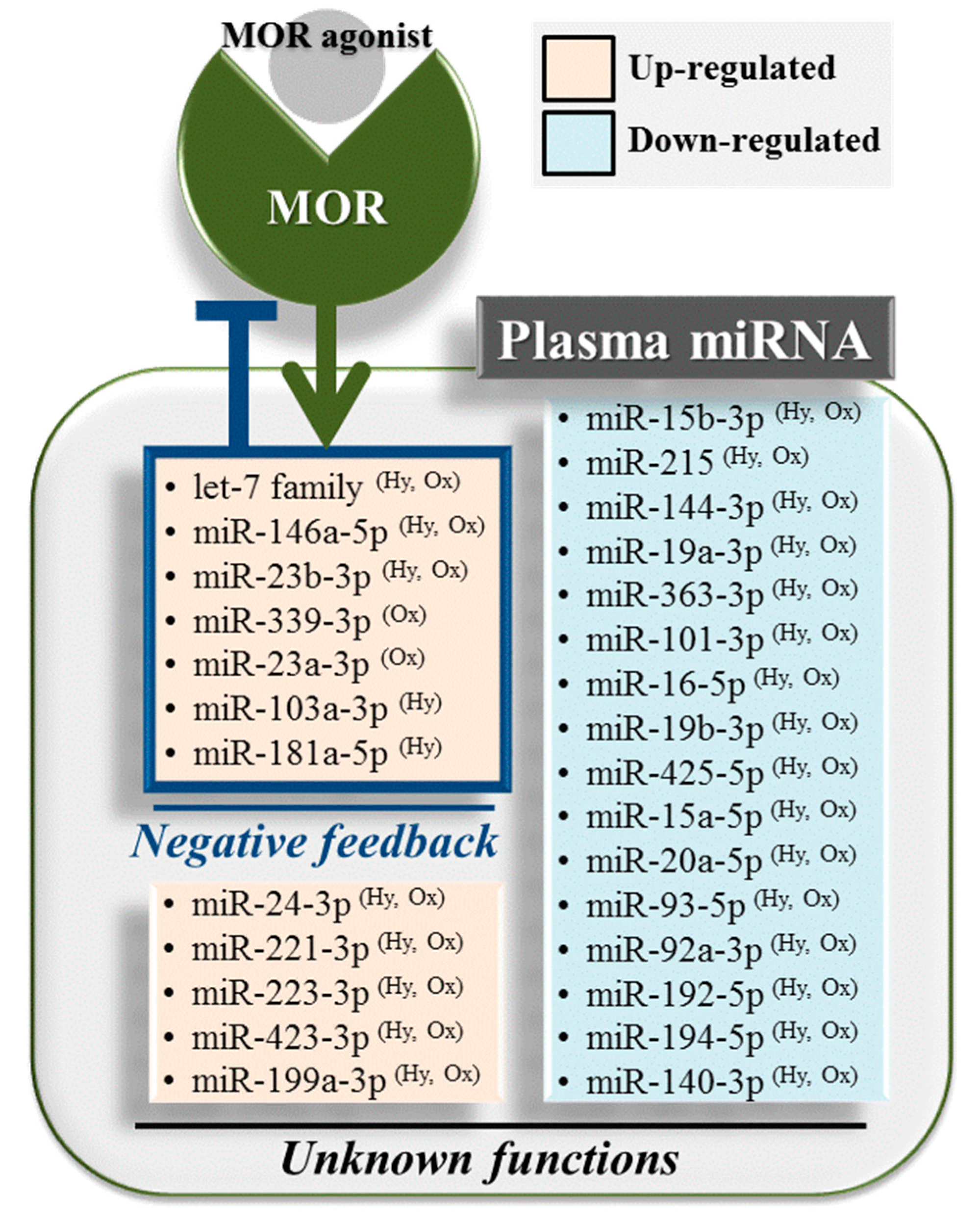
2.1. Overview of Differentially Regulated miRNAs
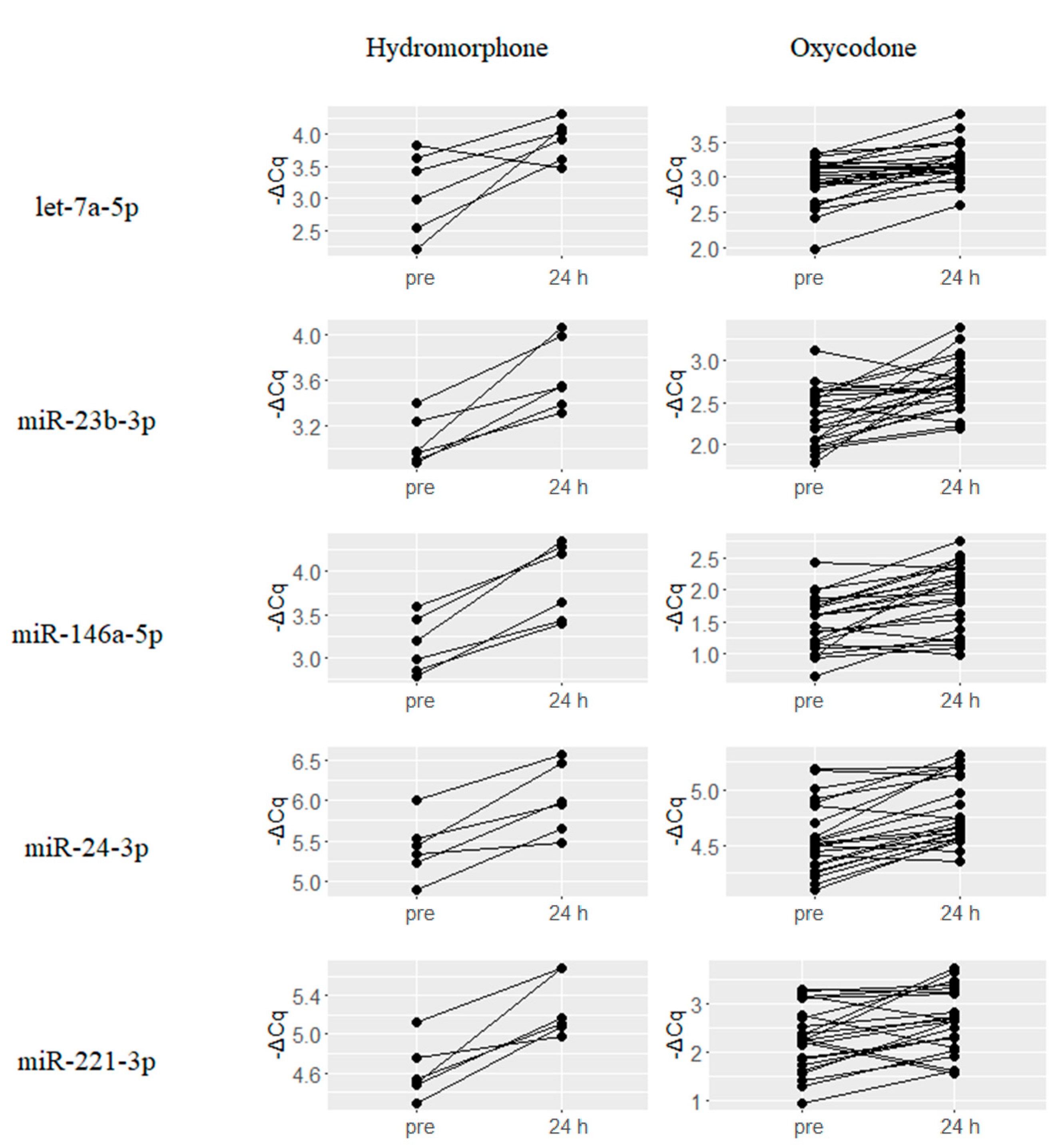
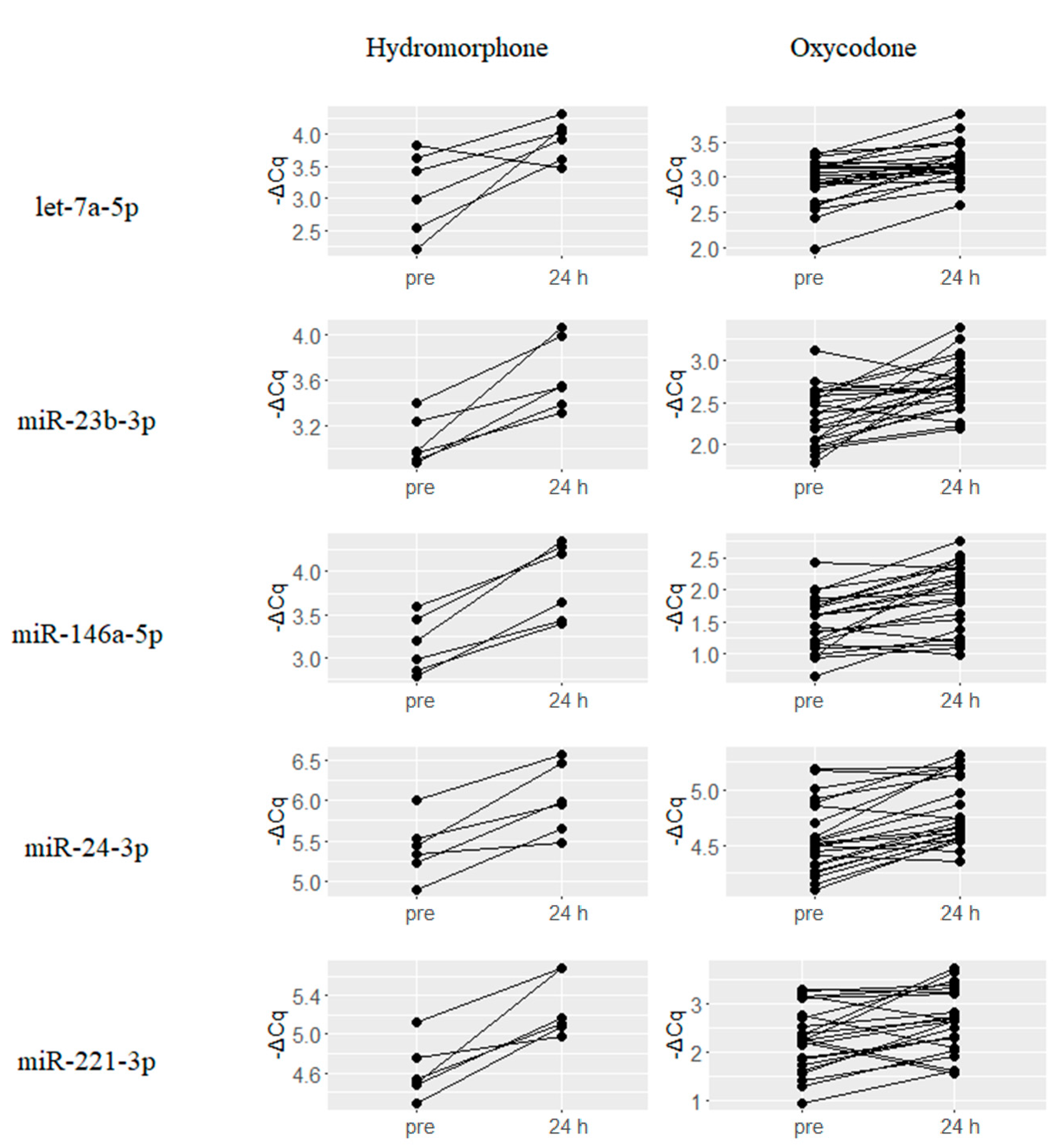
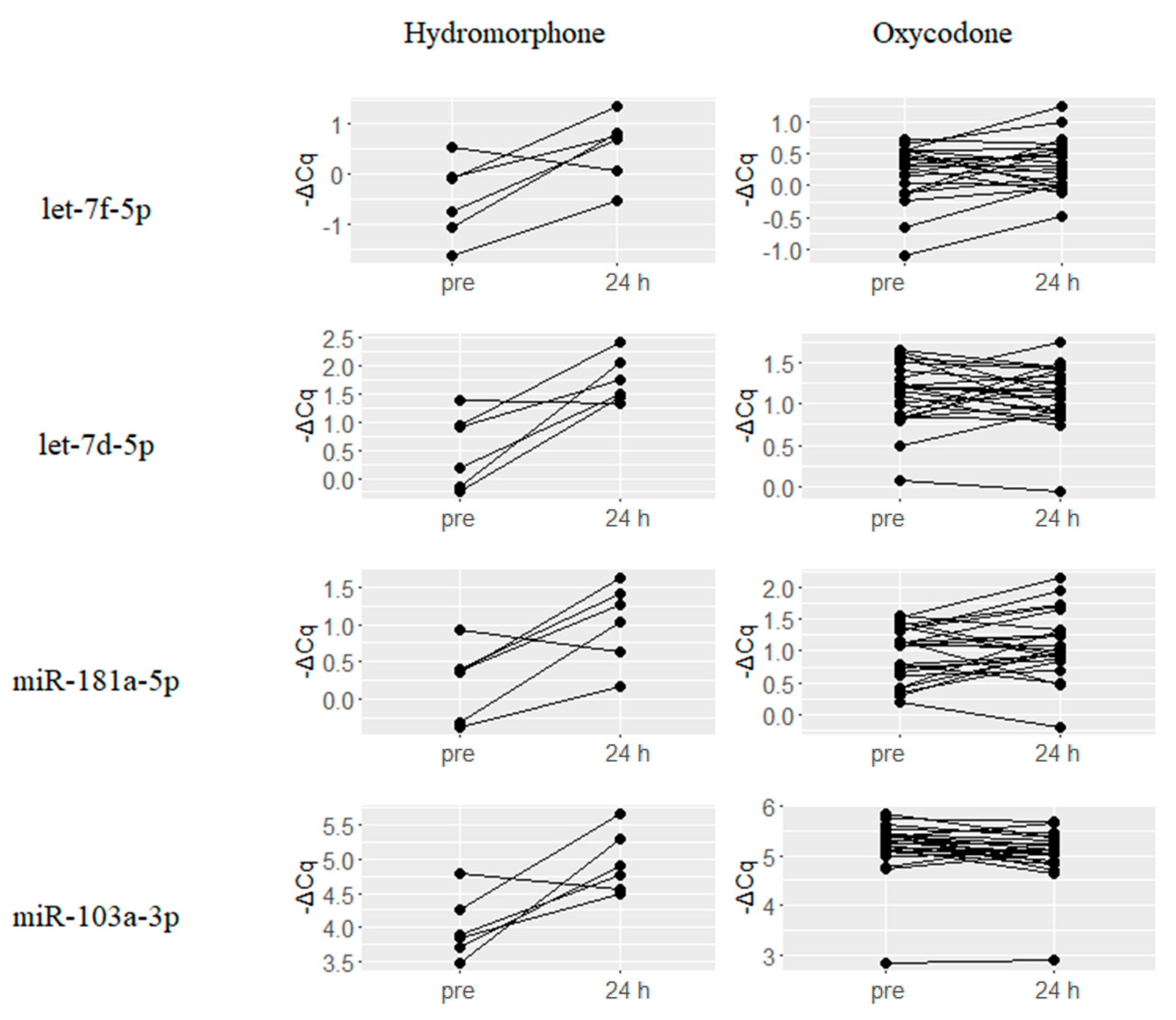
2.2. Upregulated miRNAs upon Hydromorphone and Oxycodone Treatment
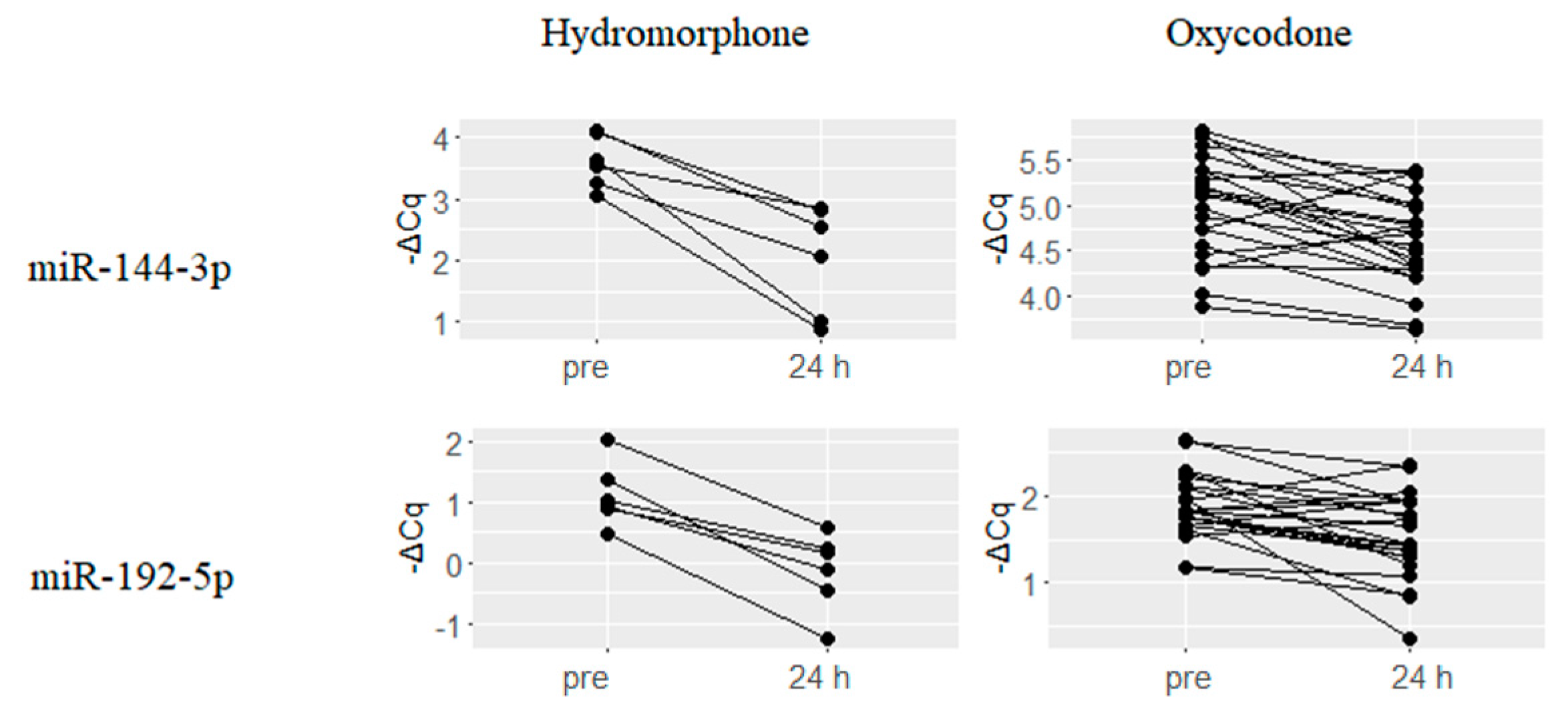
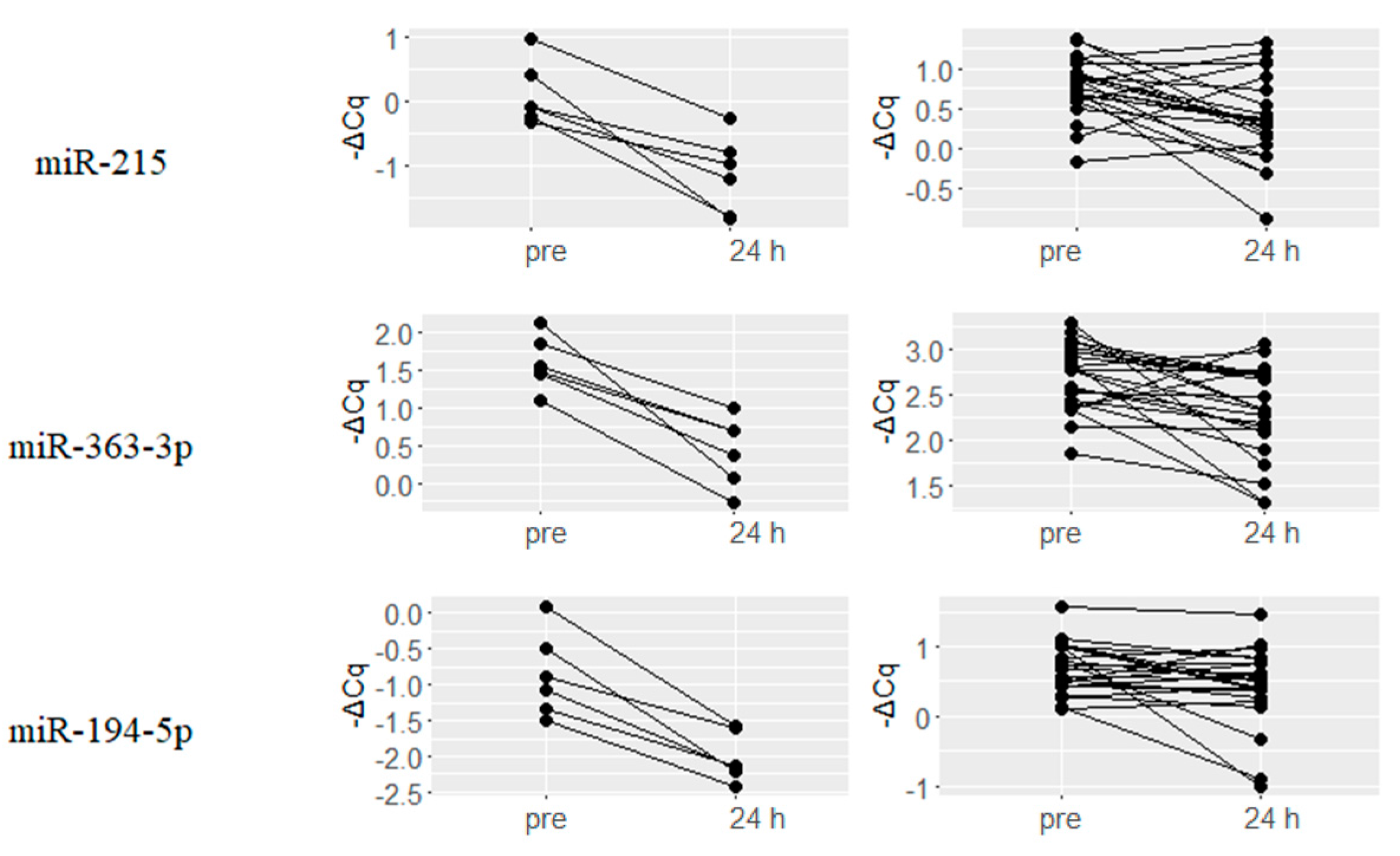
2.3. Downregulated miRNAs upon Hydromorphone and Oxycodone Treatment
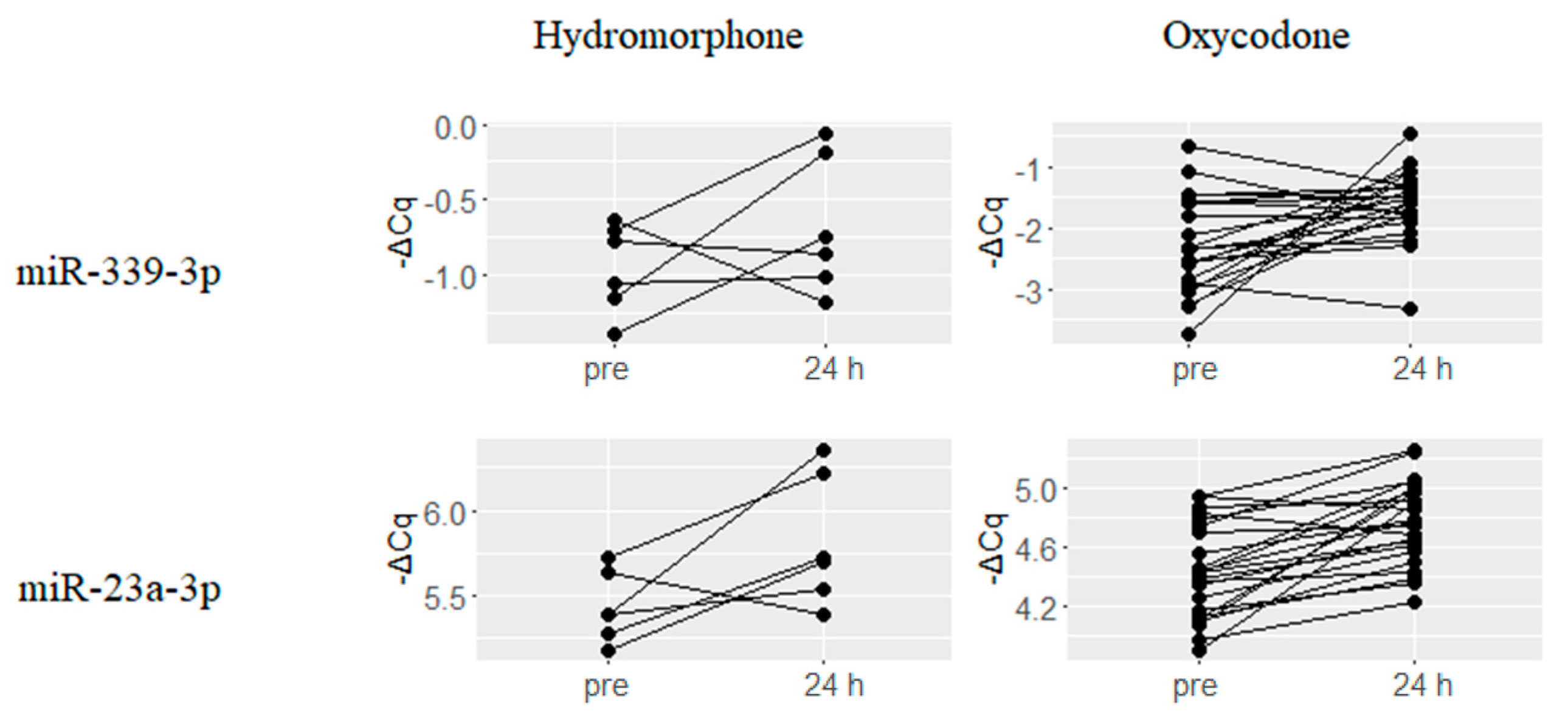
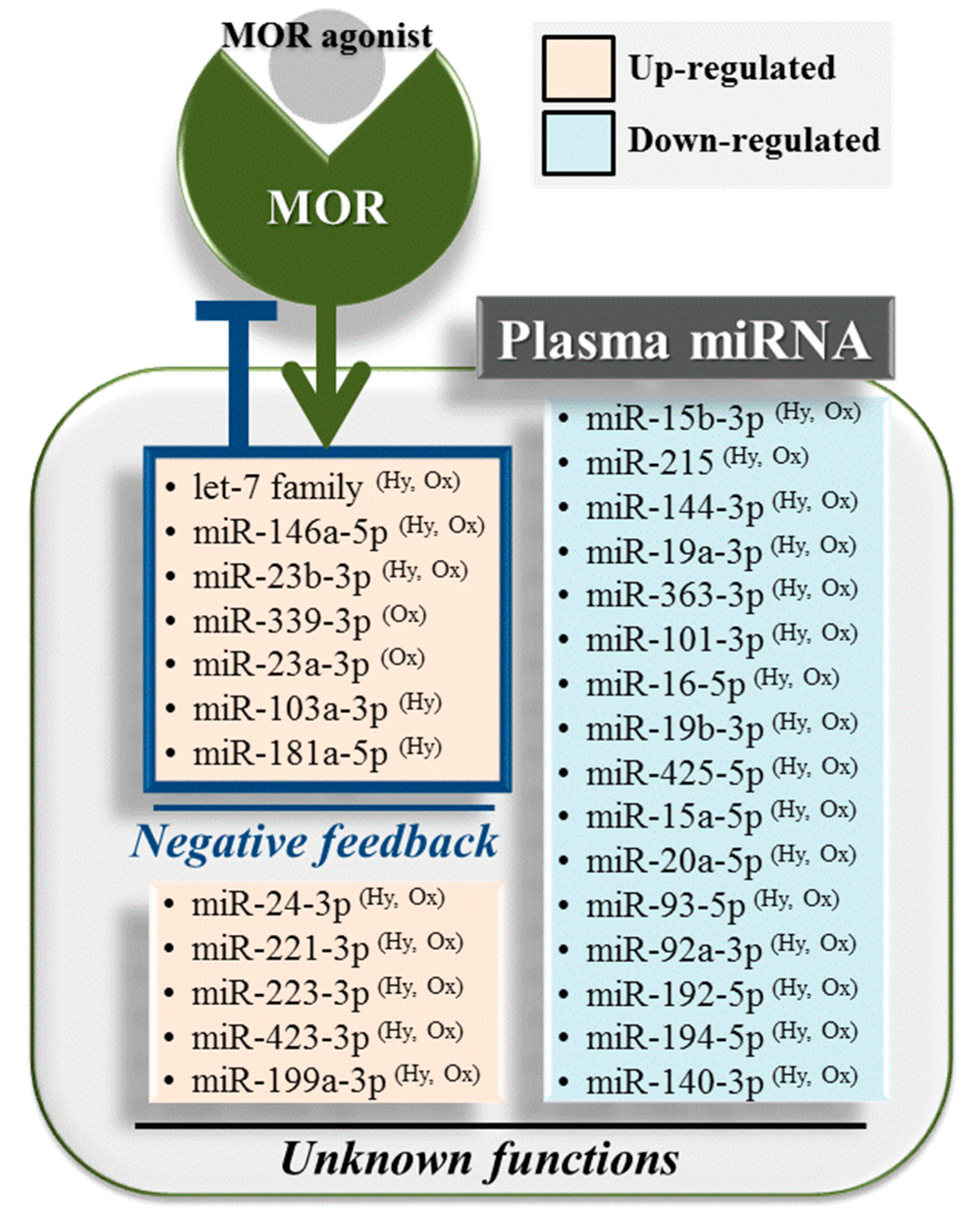
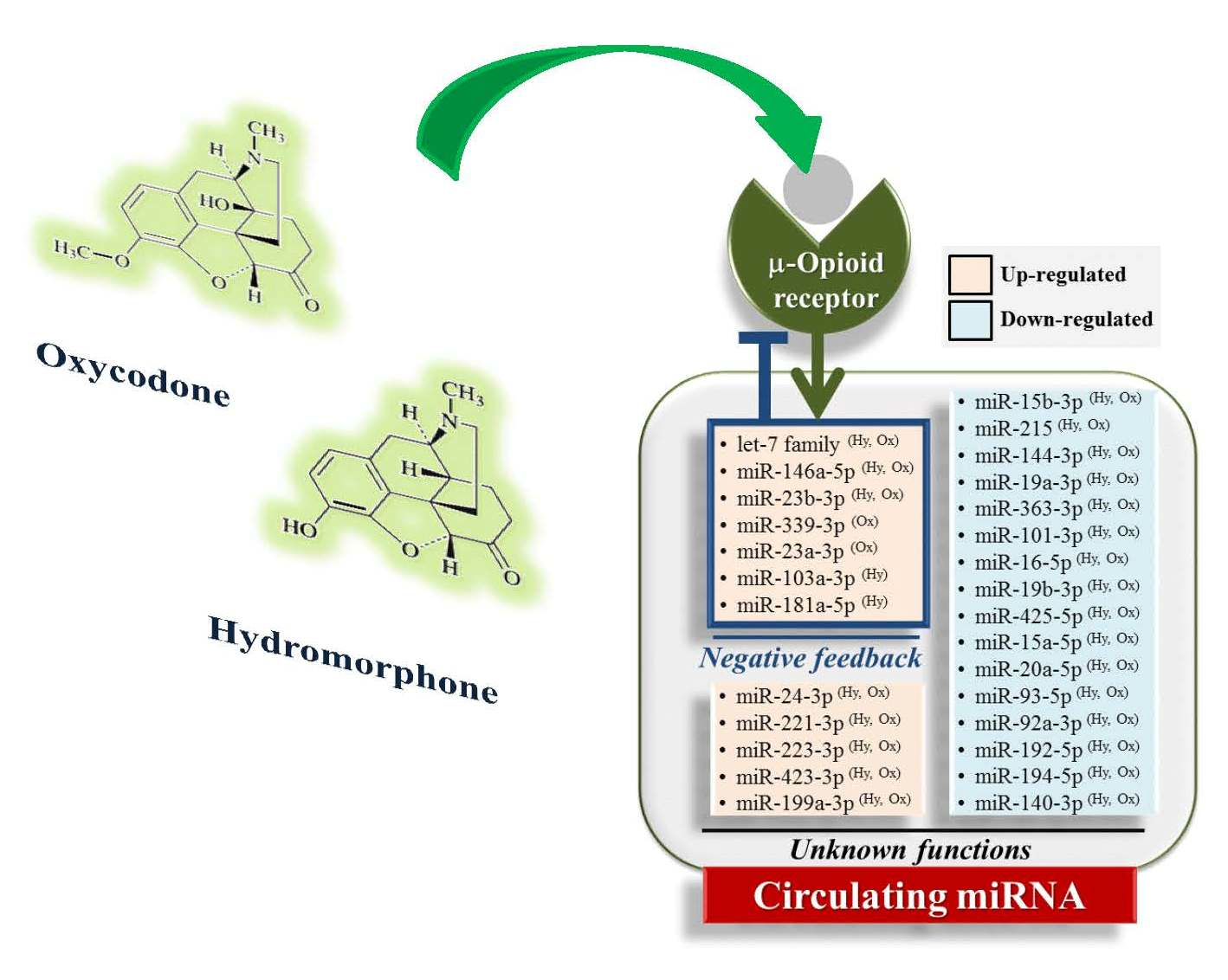
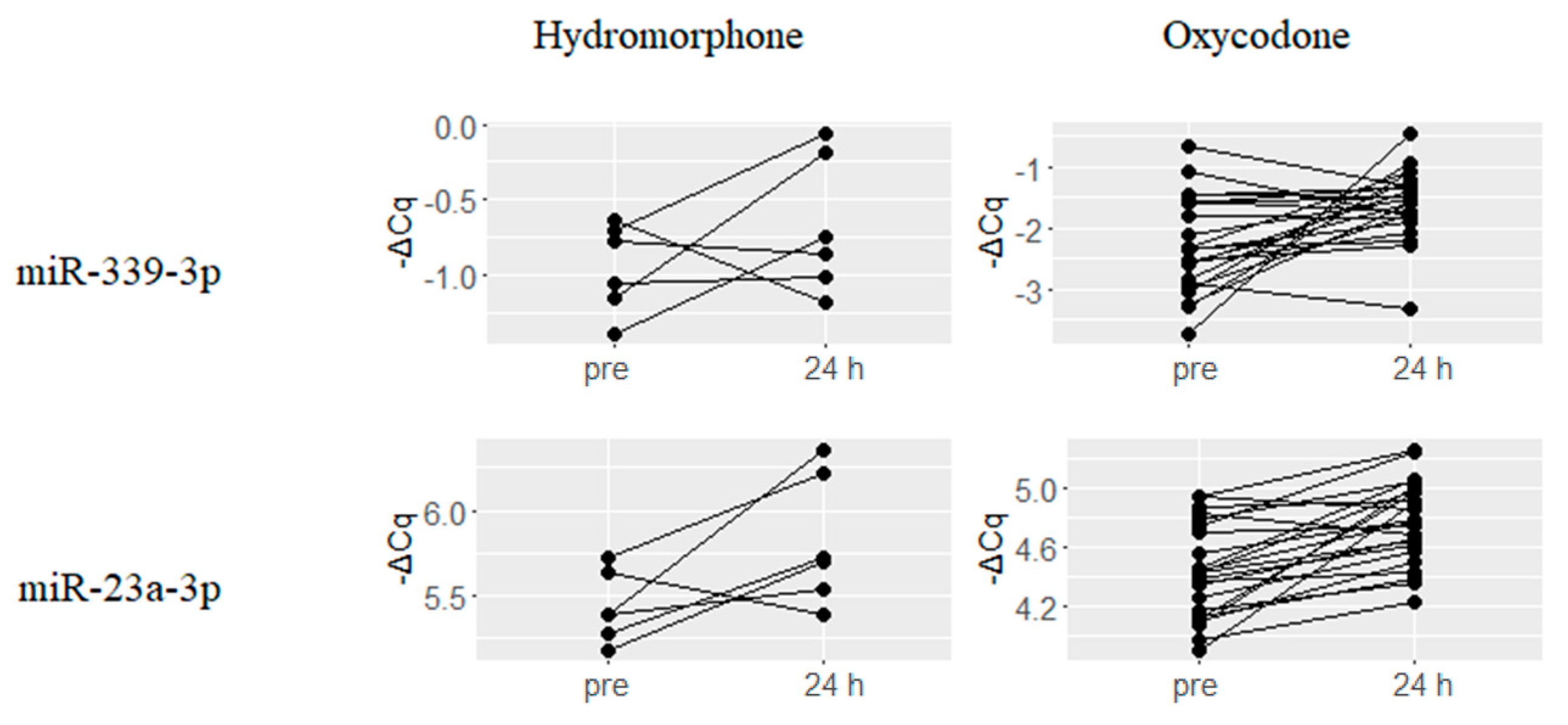
2.4. Hydromorphone- or Oxycodone-Induced miRNA Upregulation Is Associated with μ-Opioid Receptor (MOR) Signaling
3. Discussion
4. Materials and Methods
4.1. Summary of Clinical Studies
4.1.1. Hydromorphone Study (Japic-132167)
4.1.2. Oxycodone Study (Japic-163475)
4.2. miRNA Extraction from Plasma and qRT-PCR Analysis
4.3. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CNS | central nervous system |
| ER | extended release |
| miRNA | microRNA |
| MOR | μ-opioid receptor |
| PCR | polymerase chain reaction |
| qRT-PCR | quantitative real-time PCR |
| UTR | untranslated region |
References
- Al-Hasani, R.; Bruchas, M.R. Molecular mechanisms of opioid receptor-dependent signaling and behavior. Anesthesiology 2011, 115, 1363–1381. [Google Scholar] [CrossRef] [PubMed]
- Barbierato, M.; Zusso, M.; Skaper, S.D.; Giusti, P. MicroRNAs: Emerging role in the endogenous mu opioid system. CNS Neurol. Disord. Drug Targets 2015, 14, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Bruehl, S.; Apkarian, A.V.; Ballantyne, J.C.; Berger, A.; Borsook, D.; Chen, W.G.; Farrar, J.T.; Haythornthwaite, J.A.; Horn, S.D.; Iadarola, M.J.; et al. Personalized medicine and opioid analgesic prescribing for chronic pain: Opportunities and challenges. J. Pain 2013, 14, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, J.; Anderson, J.; Baumgartner, R.; Coimbra, A.; Schwarz, A.J.; Pendse, G.; Wallin, D.; Nutile, L.; Bishop, J.; George, E.; et al. Modulation of CNS pain circuitry by intravenous and sublingual doses of buprenorphine. NeuroImage 2012, 59, 3762–3773. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, J.; Anderson, J.; Schwarz, A.J.; Coimbra, A.; Baumgartner, R.; Pendse, G.; George, E.; Nutile, L.; Wallin, D.; Bishop, J.; et al. Imaging drugs with and without clinical analgesic efficacy. Neuropsychopharmacology 2011, 36, 2659–2673. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef] [PubMed]
- Cortez, M.A.; Bueso-Ramos, C.; Ferdin, J.; Lopez-Berestein, G.; Sood, A.K.; Calin, G.A. MicroRNAs in body fluids—The mix of hormones and biomarkers. Nat. Rev. Clin. Oncol. 2011, 8, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed]
- Im, H.I.; Kenny, P.J. MicroRNAs in neuronal function and dysfunction. Trends Neurosci. 2012, 35, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Sen, S. MicroRNA as biomarkers and diagnostics. J. Cell. Physiol. 2016, 231, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.P.; Diallo, A.; Cruceanu, C.; Fiori, L.M.; Laboissiere, S.; Guillet, I.; Fontaine, J.; Ragoussis, J.; Benes, V.; Turecki, G.; et al. Biomarker discovery: Quantification of microRNAs and other small non-coding RNAs using next generation sequencing. BMC Med. Genom. 2015, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Brase, J.C.; Wuttig, D.; Kuner, R.; Sultmann, H. Serum microRNAs as non-invasive biomarkers for cancer. Mol. Cancer 2010, 9, 306. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, J.D.; Chevillet, J.R.; Kroh, E.M.; Ruf, I.K.; Pritchard, C.C.; Gibson, D.F.; Mitchell, P.S.; Bennett, C.F.; Pogosova-Agadjanyan, E.L.; Stirewalt, D.L.; et al. Argonaute2 complexes carry a population of circulating microRNAs independent of vesicles in human plasma. Proc. Natl. Acad. Sci. USA 2011, 108, 5003–5008. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Li, J.; Huang, B.; Liu, J.; Chen, X.; Chen, X.M.; Xu, Y.M.; Huang, L.F.; Wang, X.Z. Exosomes: Novel biomarkers for clinical diagnosis. Sci. World J. 2015, 2015, 657086. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, A.; Filant, J.; Moxley, K.M.; Sood, A.; McMeekin, S.; Ramesh, R. Exosomes: A role for naturally occurring nanovesicles in cancer growth, diagnosis and treatment. Curr. Gene Ther. 2015, 15, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular miRNAs: The mystery of their origin and function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Higa, G.S.; de Sousa, E.; Walter, L.T.; Kinjo, E.R.; Resende, R.R.; Kihara, A.H. MicroRNAs in neuronal communication. Mol. Neurobiol. 2014, 49, 1309–1326. [Google Scholar] [CrossRef] [PubMed]
- Trescot, A.M.; Datta, S.; Lee, M.; Hansen, H. Opioid pharmacology. Pain Physician 2008, 11, S133–S153. [Google Scholar] [PubMed]
- Toyama, K.; Uchida, N.; Ishizuka, H.; Sambe, T.; Kobayashi, S. Single-dose evaluation of safety, tolerability and pharmacokinetics of newly formulated hydromorphone immediate-release and hydrophilic matrix extended-release tablets in healthy Japanese subjects without co-administration of an opioid antagonist. J. Clin. Pharmacol. 2015, 55, 975–984. [Google Scholar] [CrossRef] [PubMed]
- Toyama, K.; Furuie, H.; Ishizuka, H. Bioequivalence of an extended-release oxycodone hydrochloride tablet to a marketed reference product OXYCONTIN® in healthy Japanese subjects under fasting and fed conditions. Drugs R D 2017. [Google Scholar] [CrossRef]
- Fromm, B.; Billipp, T.; Peck, L.E.; Johansen, M.; Tarver, J.E.; King, B.L.; Newcomb, J.M.; Sempere, L.F.; Flatmark, K.; Hovig, E.; et al. A uniform system for the annotation of vertebrate microRNA genes and the evolution of the human microRNAome. Annu. Rev. Genet. 2015, 49, 213–242. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, C.; Kirkmire, C.M.; Wang, Z.J. Regulation of opioid tolerance by let-7 family microRNA targeting the mu opioid receptor. J. Neurosci. 2010, 30, 10251–10258. [Google Scholar] [CrossRef] [PubMed]
- Dave, R.S.; Khalili, K. Morphine treatment of human monocyte-derived macrophages induces differential miRNA and protein expression: Impact on inflammation and oxidative stress in the central nervous system. J. Cell. Biochem. 2010, 110, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cao, D.L.; Jiang, B.C.; Yang, T.; Gao, Y.J. MicroRNA-146a-5p attenuates neuropathic pain via suppressing TRAF6 signaling in the spinal cord. Brain Behav. Immun. 2015, 49, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, L.; Law, P.Y.; Wei, L.N.; Loh, H.H. Long-term morphine treatment decreases the association of mu-opioid receptor (MOR1) mRNA with polysomes through miRNA23b. Mol. Pharmacol. 2009, 75, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, Y.; Zheng, H.; Loh, H.H.; Law, P.Y. Morphine modulates mouse hippocampal progenitor cell lineages by upregulating miR-181a level. Stem Cells 2014, 32, 2961–2972. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekar, V.; Dreyer, J.L. MicroRNAs miR-124, let-7d and miR-181a regulate cocaine-induced plasticity. Mol. Cell. Neurosci. 2009, 42, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Xu, J.; Xu, M.; Pasternak, G.W.; Pan, Y.X. Morphine regulates expression of mu-opioid receptor MOR-1A, an intron-retention carboxyl terminal splice variant of the mu-opioid receptor (OPRM1) gene via miR-103/miR-107. Mol. Pharmacol. 2014, 85, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Hwang, C.K.; Zheng, H.; Wagley, Y.; Lin, H.Y.; Kim do, K.; Law, P.Y.; Loh, H.H.; Wei, L.N. MicroRNA 339 down-regulates mu-opioid receptor at the post-transcriptional level in response to opioid treatment. FASEB J. 2013, 27, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Zeng, Y.; Zhang, X.; Chu, J.; Loh, H.H.; Law, P.Y. mu-Opioid receptor agonists differentially regulate the expression of miR-190 and NeuroD. Mol. Pharmacol. 2010, 77, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Boyerinas, B.; Park, S.M.; Hau, A.; Murmann, A.E.; Peter, M.E. The role of let-7 in cell differentiation and cancer. Endocr. Relat. Cancer 2010, 17, F19–F36. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Inoue, K.; Fujiwara, A.; Hatakeyama, K.; Kanto, K.; Watanabe, Y.; Muramatsu, K.; Fukuda, Y.; Ogura, S.; Yamaguchi, K.; et al. Let-7 microRNA family is selectively secreted into the extracellular environment via exosomes in a metastatic gastric cancer cell line. PLoS ONE 2010, 5, e13247. [Google Scholar] [CrossRef] [PubMed]
- Kiyosawa, N.; Manabe, S.; Yamoto, T.; Sanbuissho, A. Practical application of toxicogenomics for profiling toxicant-induced biological perturbations. Int. J. Mol. Sci. 2010, 11, 3397–3412. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Thomson, T.M.; Sewer, A.; Drubin, D.A.; Mathis, C.; Weisensee, D.; Pratt, D.; Hoeng, J.; Peitsch, M.C. Assessment of network perturbation amplitudes by applying high-throughput data to causal biological networks. BMC Syst. Biol. 2012, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Data Analysis Guide For the miRCURY LNA™ Universal RT MicroRNA Ready-to-Use PCR Panels Using Exiqon GenEx Software Version 3 (February 2014). Available online: https://www.exiqon.com/ls/Documents/Scientific/Exiqon-data-analysis-guide.pdf (accessed on 4 July 2017).
- Mestdagh, P.; van Vlierberghe, P.; de Weer, A.; Muth, D.; Westermann, F.; Speleman, F.; Vandesompele, J. A novel and universal method for microRNA RT-qPCR data normalization. Genome Biol. 2009, 10, R64. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hydromorphone (N = 6) | Oxycodone (N = 23) | ||||
|---|---|---|---|---|---|
| miRNA | ΔCq(24 h–Pre) | q-Value | miRNA | ΔCq(24 h–Pre) | q-Value |
| hsa-miR-127-3p | −2.85 | 0.003 | hsa-miR-361-3p | −1.18 | 0.005 |
| hsa-miR-182-5p | −1.87 | 0.044 | hsa-miR-208a | −0.89 | 0.003 |
| hsa-miR-339-5p | −1.47 | 0.006 | hsa-miR-339-3p | −0.69 | 0.027 |
| hsa-miR-374b-5p | −1.39 | 0.003 | hsa-miR-10b-5p | −0.62 | 0.002 |
| hsa-miR-28-5p | −1.33 | 0.009 | hsa-miR-28-3p | −0.59 | 0.042 |
| hsa-miR-551b-3p | −1.33 | 0.008 | hsa-miR-125a-5p | −0.51 | 0.004 |
| hsa-let-7d-5p | −1.25 | 0.003 | hsa-miR-342-3p | −0.49 | 0 |
| hsa-miR-26a-5p | −1.15 | 0.009 | hsa-miR-150-5p | −0.48 | 0.001 |
| hsa-miR-382-5p | −1.09 | 0.037 | hsa-miR-146b-5p | −0.46 | 0.005 |
| hsa-let-7f-5p | −1.05 | 0.018 | hsa-miR-197-3p | −0.40 | 0.01 |
| hsa-miR-154-5p | −1.01 | 0.039 | hsa-miR-146a-5p | −0.40 | 0.001 |
| hsa-let-7e-5p | −1.00 | 0.042 | hsa-miR-23b-3p | −0.38 | 0.001 |
| hsa-miR-191-5p | −0.98 | 0.013 | hsa-miR-221-3p | −0.36 | 0.027 |
| hsa-miR-103a-3p | −0.95 | 0.004 | hsa-miR-23a-3p | −0.33 | 0.001 |
| hsa-miR-423-3p | −0.91 | 0.004 | hsa-miR-199a-3p | −0.32 | 0.031 |
| hsa-miR-1 | −0.87 | 0.043 | hsa-let-7a-5p | −0.29 | 0.027 |
| hsa-miR-766-3p | −0.84 | 0.04 | hsa-miR-29a-3p | −0.29 | 0.027 |
| hsa-let-7a-5p | −0.80 | 0.009 | hsa-miR-223-3p | −0.27 | 0.011 |
| hsa-miR-181a-5p | −0.79 | 0.035 | hsa-miR-24-3p | −0.25 | 0.001 |
| hsa-miR-199a-3p | −0.77 | 0.003 | hsa-miR-142-3p | −0.24 | 0.011 |
| hsa-miR-146a-5p | −0.75 | 0.004 | hsa-miR-423-3p | −0.23 | 0.038 |
| hsa-miR-328 | −0.73 | 0.002 | hsa-miR-126-3p | −0.17 | 0.033 |
| hsa-let-7c | −0.73 | 0.019 | |||
| hsa-miR-221-3p | −0.67 | 0.012 | |||
| hsa-miR-151a-5p | −0.67 | 0.009 | |||
| hsa-miR-142-5p | −0.63 | 0.026 | |||
| hsa-miR-24-3p | −0.61 | 0.005 | |||
| hsa-miR-324-5p | −0.60 | 0.035 | |||
| hsa-miR-107 | −0.60 | 0.022 | |||
| hsa-miR-151a-3p | −0.60 | 0.011 | |||
| hsa-miR-23b-3p | −0.58 | 0.005 | |||
| hsa-miR-146b-5p | −0.56 | 0.035 | |||
| hsa-miR-223-3p | −0.52 | 0.02 | |||
| hsa-miR-652-3p | −0.49 | 0.006 | |||
| hsa-miR-140-5p | −0.41 | 0.043 | |||
| Hydromorphone (N = 6) | Oxycodone (N = 23) | ||||
|---|---|---|---|---|---|
| miRNA | ΔCq(24 h–Pre) | q-Value | miRNA | ΔCq(24 h–Pre) | q-Value |
| hsa-miR-885-5p | 2.47 | 0.011 | hsa-miR-182-5p | 1.34 | 0.004 |
| hsa-miR-122-5p | 1.71 | 0.001 | hsa-miR-15b-3p | 0.6 | 0 |
| hsa-miR-375 | 1.61 | 0.002 | hsa-miR-451a | 0.41 | 0.002 |
| hsa-miR-144-3p | 1.58 | 0.003 | hsa-miR-215 | 0.4 | 0.003 |
| hsa-miR-451a | 1.39 | 0.003 | hsa-miR-144-3p | 0.4 | 0.003 |
| hsa-let-7b-3p | 1.3 | 0.011 | hsa-miR-19a-3p | 0.39 | 0.001 |
| hsa-miR-192-5p | 1.26 | 0.001 | hsa-miR-363-3p | 0.39 | 0.003 |
| hsa-miR-215 | 1.25 | 0 | hsa-miR-192-5p | 0.35 | 0.003 |
| hsa-miR-363-3p | 1.17 | 0.003 | hsa-miR-101-3p | 0.34 | 0.003 |
| hsa-miR-194-5p | 1.14 | 0.004 | hsa-miR-16-5p | 0.33 | 0.003 |
| hsa-miR-660-5p | 1.14 | 0.003 | hsa-miR-106b-5p | 0.33 | 0.003 |
| hsa-miR-140-3p | 1.11 | 0.004 | hsa-miR-19b-3p | 0.32 | 0.002 |
| hsa-miR-378a-3p | 1.1 | 0 | hsa-miR-185-5p | 0.32 | 0.005 |
| hsa-miR-486-5p | 1.08 | 0.006 | hsa-miR-17-5p | 0.29 | 0.038 |
| hsa-miR-424-5p | 1.02 | 0.003 | hsa-miR-324-3p | 0.29 | 0.006 |
| hsa-miR-144-5p | 1 | 0.03 | hsa-miR-425-5p | 0.29 | 0.004 |
| hsa-miR-16-5p | 1 | 0.003 | hsa-miR-18b-5p | 0.27 | 0.004 |
| hsa-miR-502-3p | 0.98 | 0.012 | hsa-miR-18a-5p | 0.27 | 0.003 |
| hsa-miR-16-2-3p | 0.97 | 0.011 | hsa-miR-15a-5p | 0.27 | 0.01 |
| hsa-miR-99a-5p | 0.95 | 0.004 | hsa-miR-106a-5p | 0.26 | 0.005 |
| hsa-miR-25-3p | 0.95 | 0.003 | hsa-miR-20a-5p | 0.26 | 0.01 |
| hsa-miR-365a-3p | 0.93 | 0.042 | hsa-miR-194-5p | 0.25 | 0.031 |
| hsa-miR-15a-5p | 0.91 | 0.003 | hsa-miR-30b-5p | 0.25 | 0.004 |
| hsa-miR-10b-5p | 0.88 | 0.042 | hsa-miR-140-3p | 0.24 | 0.031 |
| hsa-miR-92a-3p | 0.83 | 0.003 | hsa-miR-29b-3p | 0.24 | 0.028 |
| hsa-miR-101-3p | 0.78 | 0.011 | hsa-miR-93-5p | 0.23 | 0.032 |
| hsa-miR-20b-5p | 0.75 | 0.044 | hsa-miR-92a-3p | 0.23 | 0.028 |
| hsa-miR-210 | 0.74 | 0.012 | hsa-miR-484 | 0.21 | 0.014 |
| hsa-miR-497-5p | 0.73 | 0.042 | hsa-let-7g-5p | 0.21 | 0.014 |
| hsa-miR-19b-3p | 0.72 | 0.018 | hsa-miR-148b-3p | 0.2 | 0.017 |
| hsa-miR-30a-5p | 0.66 | 0.03 | hsa-miR-30e-5p | 0.2 | 0.01 |
| hsa-miR-19a-3p | 0.61 | 0.019 | |||
| hsa-miR-15b-3p | 0.59 | 0.036 | |||
| hsa-miR-425-5p | 0.42 | 0.031 | |||
| hsa-miR-93-5p | 0.37 | 0.042 | |||
| hsa-miR-20a-5p | 0.35 | 0.046 | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyama, K.; Kiyosawa, N.; Watanabe, K.; Ishizuka, H. Identification of Circulating miRNAs Differentially Regulated by Opioid Treatment. Int. J. Mol. Sci. 2017, 18, 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091991
Toyama K, Kiyosawa N, Watanabe K, Ishizuka H. Identification of Circulating miRNAs Differentially Regulated by Opioid Treatment. International Journal of Molecular Sciences. 2017; 18(9):1991. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091991
Chicago/Turabian StyleToyama, Kaoru, Naoki Kiyosawa, Kenji Watanabe, and Hitoshi Ishizuka. 2017. "Identification of Circulating miRNAs Differentially Regulated by Opioid Treatment" International Journal of Molecular Sciences 18, no. 9: 1991. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18091991