To Wnt or Lose: The Missing Non-Coding Linc in Colorectal Cancer


1
Department of Experimental Therapeutics, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
2
Nanfang Hospital, Southern Medical University/The First School of Clinical Medicine, Southern Medical University, Guangzhou 510515, Guangdong, China
3
Research Unit of Non-Coding RNA and Genome Editing in Cancer, Division of Oncology, Medical University of Graz, Graz 8010, Austria
4
The Center for RNA Interference and Non-Coding RNAs, The University of Texas MD Anderson Cancer Center, Houston, TX 77030, USA
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2017, 18(9), 2003; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18092003
Submission received: 9 August 2017
/
Revised: 9 September 2017
/
Accepted: 13 September 2017
/
Published: 20 September 2017
(This article belongs to the Collection Regulation by Non-coding RNAs)
Abstract
:Colorectal cancer (CRC) is the third most frequent cancer and one of the leading causes for cancer-related mortality. Aberrant activation of the Wnt signaling is an essential initiating factor in colon carcinogenesis, and a driving force of CRC progression. Recently, long non-coding RNAs (lncRNAs) have emerged as significant players in CRC pathogenesis through diversified mechanisms. Although both Wnt signaling and lncRNAs represent interesting research areas for CRC, an effort of directly connecting these two areas is lacking. To fill in the knowledge gap, we focus on the reported findings of lncRNAs that regulate Wnt signaling or essential Wnt signaling targets. These include several newly discovered lncRNAs originated from the amplified cancer-associated chromosome 8q24 region that surrounds the essential Wnt target MYC gene, lncRNAs reported to be involved in CRC stem cells, and several individual lncRNAs connected to Wnt signaling through other mechanisms. This review will provide essential information that assists in understanding the missing link of lncRNAs to the classical Wnt signaling in CRC.
1. Introduction
Colorectal cancer (CRC), accounting for 8% of new cancer cases, is the third most frequent cancer, and one of the leading cause of cancer-related mortalities in the United States [1]. Despite the fact that the CRC incidence rates declined in people aged 50 years or older, CRC incidence rates increased by 22% from 2000 to 2013 in adults younger than 50 years in the United States [2]. Similarly, CRC mortality rates increased by 13% in those less than 50 years old during the same period [2]. The rise of CRC with early age at diagnosis underlines the need for exploration of new avenues for disease understanding, and the development of innovative detection and intervention strategies.
Aberrant Wnt signaling is the initiating and driving event underlying the vast majority of CRC carcinogenesis [3,4,5]. Although the essential molecules of Wnt signaling have been well characterized, how this pathway is fine-tuned at other layers remains largely unexplored. Non-coding RNA transcripts such as microRNAs have been revealed to restrain or activate Wnt signaling, by controlling the expression of Wnt signaling proteins [6,7,8]. More recently, long non-coding RNAs (lncRNAs), defined as being at least 200 nucleotides in length, show significant association with CRC incidence, the extent of malignancy, and patient prognosis [9,10]. However, the mechanisms underlying lncRNA involvement in CRC lag far behind its discovery pace and remain largely uncharacterized, partially because of the lack of a unified molecular mechanism.
We reason that because Wnt signaling is an initiating force in CRC carcinogenesis, lncRNAs that participate in this pathway might represent the novel research avenue for mechanisms of understanding the other regulatory layers of CRC. We acknowledge that many excellent reviews have covered each topic of Wnt signaling and lncRNA independently, and refer the readers to this literature for more in-depth information [9,11,12]. In this review, we focus on the knowledge connecting these two topics, and aim to review recent findings linking lncRNAs with essential Wnt signaling. Herein, we briefly introduce the individual topic, summarize the typical examples that connect these two topics, and discuss the clinical application potential of lncRNAs in CRC. We further separated these lncRNAs into several categories: those transcribed from the 8q24 cancer risk region, associated with CRC stem cells, and others. To keep this review concise while informative, we list only the essential findings limited by our own knowledge, and apologize to those whose work was not referenced due to space restrictions.
2. Wnt: The Initiating and Driving Force of Colorectal Cancer (CRC)
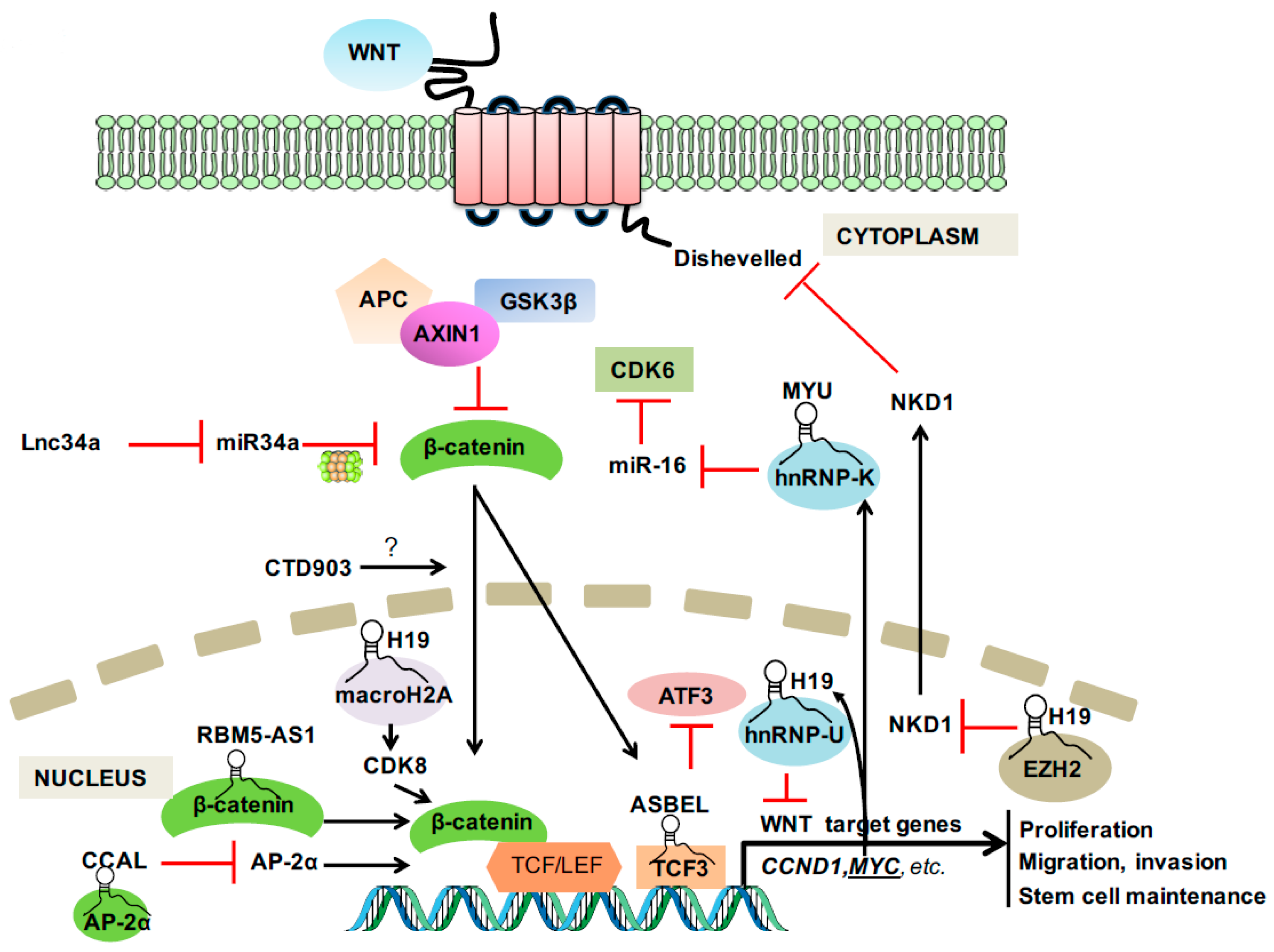
In 1982, the first Wnt family member Int1 (now known as Wnt1) was identified as a mouse proto-oncogene that is activated by the integration of mouse mammary tumor virus [13]. Subsequent research demonstrated that Int1 is a vertebrate homologue of the previously identified fly Wingless (Wg) gene, from which the term Wnt originates [14]. As the most well-known and best characterized Wnt signaling pathway, the canonical Wnt signaling that involves β-catenin and members of the lymphocyte-enhancer-binding factor (LEF)/T-cell factor (TCF) family is the fundamental driving force of CRC [15]. In the presence of a Wnt ligand binding to its receptor complex, β-catenin is rescued from ubiquitination and proteasomal breakdown by the destruction complex comprising adenomatous polyposis coli (APC), AXIN1, and GSK3β [16,17]. The stabilized β-catenin translocates to nucleus and forms a complex with LEF/TCF transcription factors to activate the transcription of a wide range of Wnt target genes [18] (Figure 1).
The Cancer Genome Atlas (TCGA) consortium revealed that the Wnt signaling pathway was altered in more than 90% of CRC tumors, with mutational inactivation of the APC tumor suppressor gene, located at chromosome 5q21-q22, in ~80% of cases [19]. As an initiating event in both familial adenomatous polyposis and sporadic CRCs [20,21], the mutational inactivation of APC leads to the accumulation of β-catenin in the nucleus, a hallmark of the canonical Wnt signaling, and the transcriptional activation of Wnt target genes by β-catenin/TCF complex [22,23]. Wnt signaling is essential in maintaining the stem cell niche, and high Wnt activity was reported to accurately define the CRC stem cell population [24]. Experimentally, restoration of APC reverted CRC tumorigenic lesions by re-establishing the normal crypt homeostasis, even in mice harboring oncogenic Kras and mutated p53 [25]. This experimental finding not only reinforces the essential suppressor function of APC in CRC initiation, but also revealed the critical importance of APC-regulated Wnt signaling in CRC progression. As classical downstream targets that respond to Wnt signaling, CCND1 and MYC are established drivers in CRC formation by regulating cell growth, apoptosis, migration, invasion and stem cell maintenance [26,27].
3. LncRNAs: The Emerging Dark Matters That Matter
The vast majority of the human genome is transcribed into RNA transcripts, but only a small proportion of these RNA molecules are translated into proteins [28]. The genes that do not code for proteins produce non-coding RNAs (ncRNAs) as the final output. As one type of ncRNA, microRNA (miRNA) received much attention in the last 15 years since the discovery of their cancer involvement [29]. More recently, long ncRNAs (lncRNAs), containing no less than 200 nucleotides, have emerged as important new players in cancer [12,30]. According to their genomic features, these lncRNAs can be further classified into long intergenic ncRNAs (lincRNAs), transcribed ultraconserved regions (T-UCRs), circular RNAs, promoter-associated RNAs, enhancer-associated RNAs, and others [12,31]. In this review, we use the term lncRNA for consistency, although most of the examples in the text are lincRNAs.
A previous study summarized the diverse molecular mechanisms of lncRNAs into four archetypes [32]: (i) lncRNA can serve as a molecular sensor to deliver the signal from the hint of cellular context in a temporal and spatial manner; (ii) lncRNA can serve as decoy to interfere the function of proteins, or that of miRNAs by sponging the miRNAs via sequence complementarity; (iii) lncRNA can guide chromatin-modifying proteins onto target genes, either locally in cis or distantly in trans, respectively; (iv) lncRNAs can bridge multiple proteins together to modify chromatin or stabilize subcellular structures. It should be noted that these archetypes are not exclusive, and a single lncRNA may have multiple mechanisms.
4. LncRNAs in Wnt Signaling and CRC
Since Wnt signaling is an essential pathway in CRC carcinogenesis and progression, it is not surprising that many CRC-associated lncRNAs exert their function via this pathway. The dynamic molecular mechanisms of lncRNAs also render them large diversity in regulating Wnt activity or essential Wnt downstream targets. The 8q24 region represents one of the most frequently amplified cancer-associated regions in CRC, and contains the MYC oncogene [33]. Reports in the last several years revealed that the 8q24 region is an oasis for long non-coding RNAs [34,35,36,37,38]. As such, we discuss this unique group of lncRNAs separately. We summarize three lncRNAs in the subsection “CRC stem cells—related lncRNAs” and several lncRNAs that do not share common features in the subsection “Others” (Table 1).
4.1. LncRNAs from 8q24 Region
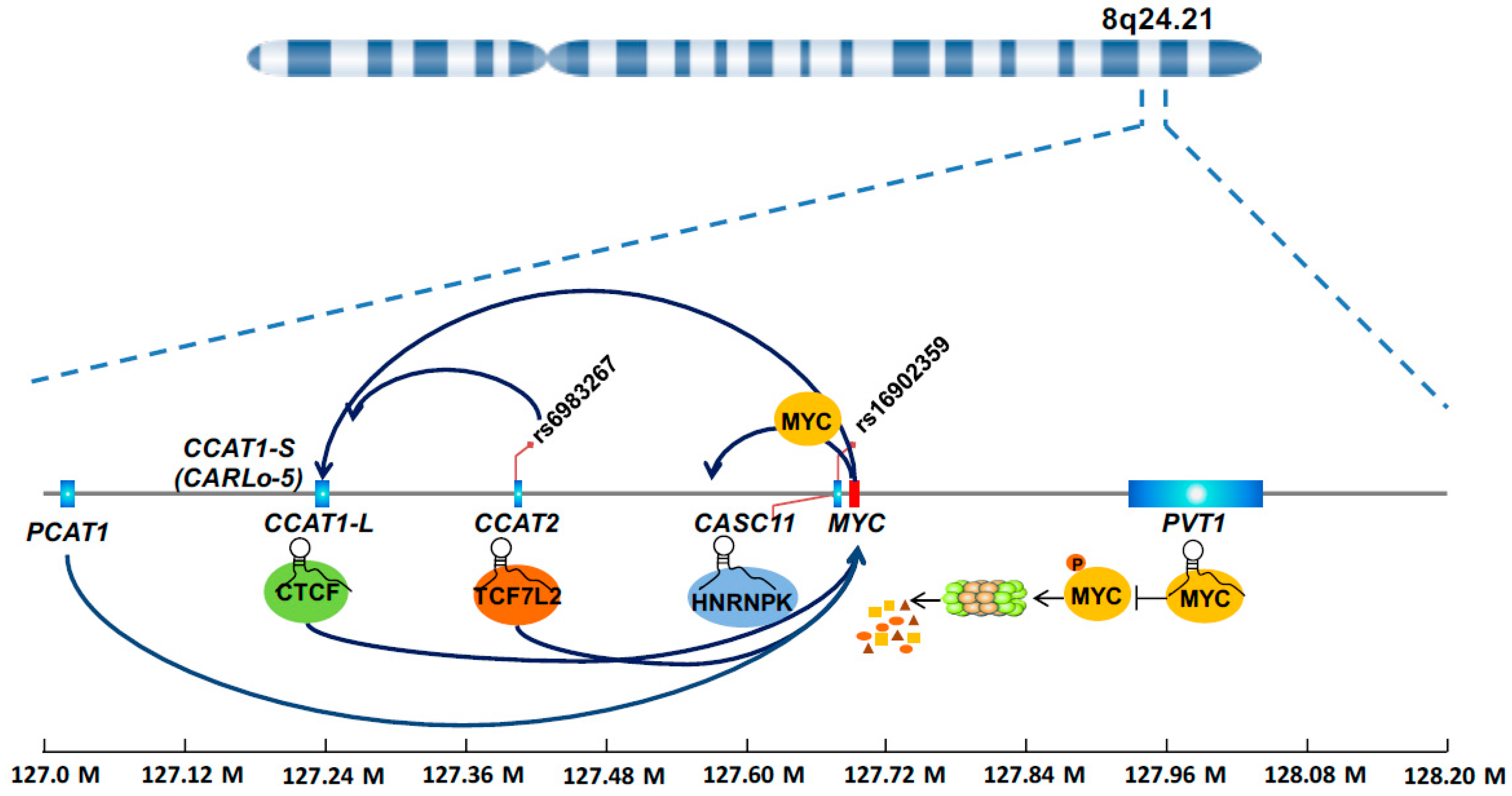
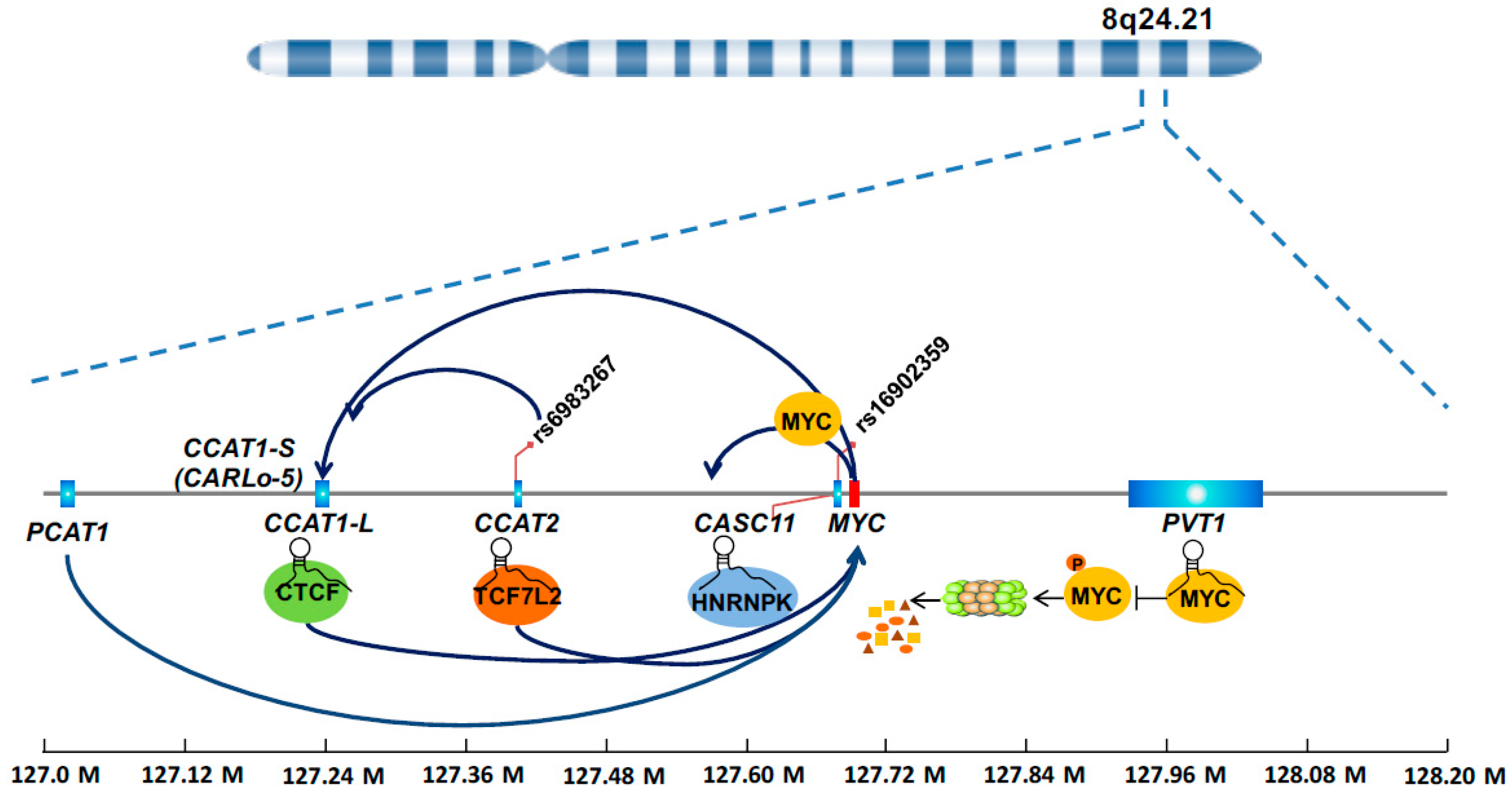
The chromosome 8q24 is frequently amplified in human cancer. Particularly, the 8q24.21 genomic region that spans almost 2 Mb but represents a desert for protein coding genes and attracts much attention for several reasons. (i) This region contains the MYC gene, which is a classical Wnt signaling target and an essential oncogene [27,39]. (ii) Genome-wide association studies consistently suggested that multiple single nucleotide polymorphisms (SNPs) in such region are associated with CRC risk [40,41,42]. (iii) DNA elements in this region have various enhancer activities that are affected by SNPs [43,44], and more recently this region was proposed as a typical example of a super-enhancer [45]. These observations, originating from a different angle but pointing to the same genomic locus, indicate that important unidentified molecular culprits reside in such region. Indeed, the last decade witnessed an explosion of discoveries of lncRNAs in the 8q24.21 region, most of which show relevance with multiple types of cancers including CRC. Not surprisingly, a large proportion of these lncRNAs were found to regulate MYC, a protein-coding gene in the region, in one way or another (Figure 2). We can envision that strong enhancer activity in this region promotes transcription of lncRNAs because of the presence of abundant transcriptional factors and mediator proteins. These lncRNAs in turn increase the enhancer activity by forming the chromatin loop or bridging the protein partners, thus creating a positive feedback mechanism in controlling MYC expression. Because of their sensitivity in responding to oncogenic signals, the lncRNAs themselves are often found to be prognosis factors in predicting the outcome of CRC patients.
4.1.1. CCAT1
The CCAT1 gene is located 515 kb upstream of the MYC oncogene, encodes a short isoform CCAT1-S and a long isoform CCAT1-L. CCAT1-S, also known as CARLo-5, is upregulated in all disease stages, including pre-malignant conditions, during CRC transformation [37]. A meta-analysis suggests significant association of increased CCAT1 expression in tumor samples with poor survival of cancer patients [46]. The expression of CCAT1-S is significantly correlated with the allele status of the SNP rs6983267, located telomeric of CCAT1-S [37]. In addition, the genomic region containing rs6983267 forms a chromatin loop with the promoter of CCAT1-S gene, suggesting a long-range interaction of rs6983267-containing region with CCAT1-S promoter in regulating its expression [37]. Experimentally, knockdown of CCAT1-S decreased CRC cell growth in vitro and in vivo [37]. Similar to its shorter isoform, CCAT1-L is highly expressed in CRC tumors, and promotes CRC growth in xenograft mouse models [47]. CCAT1-L enhances MYC transcription, as elegantly demonstrated by genome engineered cell lines that overexpressed CCAT1-L at its own gene loci [47]. Chromosome conformation capture assay suggests that CCAT1-L facilitates the formation of a long-range physical interaction loop between the MYC enhancer and its promoter [47]. Molecular mechanism study revealed an interaction of CCAT1-L with CTCF, an essential protein regulating 3D structure of the chromatin [47]. The CCAT1 gene is also transcriptionally regulated by MYC [34]. These results reveal a complex molecular interaction connecting SNP, enhancer, lncRNA, and protein in controlling MYC expression and CRC growth.
4.1.2. CCAT2
The CCAT2 gene was identified by the Calin laboratory based on several previous observations [38]. First, this high degree of conservation of this genomic region among mammals suggests the functional importance of this locus and associated transcripts [48]. Second, the rs6983267 SNP in this region is one of the most consistently reported, predisposing SNPs in prostate cancer and CRC [41,49,50]. Third, this region, 335 kb centromeric from the MYC oncogene possesses strong enhancer activity that is influenced by the rs6983267 SNP variants [43,44]. After cloning and characterization of the gene, CCAT2 was found to express at higher levels in microsatellite-stable CRC tumors that exhibit chromosomal instability (CIN), than in microsatellite-instable tumors or normal mucosae that lack the CIN feature [38]. This led to the discovery of CCAT2 initiation of CIN via cell model systems. Experimental data suggest that CCAT2 not only exerts cis regulatory effects on the nearby gene MYC, but also interacts with TCF7L2 protein to exert trans regulatory effects on Wnt signaling [38]. Together with the DNA element with enhancer activity influenced by the SNP, the CCAT2 RNA contributes to a unique Wnt signaling regulatory network. This DNA-RNA regulatory network may be essential in CRC, as deletion of this genomic region, which results in loss of both DNA elements and RNA transcripts, reduces the number of the intestine polyps in ApcMin/+ mice [51]. Recently, this non-coding RNA was reported to exert allele-specific effects on cancer metabolism by interaction with the splicing protein CFIm and ensuring alternative splicing of glutaminase [52]. In addition, multiple meta-analysis studies proved the prognostic value of CCAT2 in predicting cancer patient survival [53,54,55,56].
4.1.3. CASC11
CASC11, also known as CARLo-7, is located ~2.1 kb upstream of the MYC gene. The CASC11 gene encompasses the lymphoma predisposition SNP rs16902359. Similar to the above CCAT transcripts, CASC11 is overexpressed in CRC tumors, and high CASC11 correlates with large primary tumors and metastasis to lymph nodes [35]. Ectopic expression of CASC11 promotes CRC growth and metastasis in vitro and in vivo [35]. Mechanism study showed that CASC11 interacts with and increase the stability of heterogeneous ribonucleoprotein K (hnRNP-K), which protects β-catenin from degradation by the destruction complex, and consequently activates WNT/β-catenin signaling [35]. Forming a feedback mechanism, MYC protein binds to the promoter of the CASC11 gene to activate its transcription [35].
4.1.4. PVT1
Different from most of the non-coding genes upstream of MYC, PVT1 is located 100–500 kb downstream of the MYC gene. In addition, different from CCAT1 and CCAT2 that regulate MYC transcription, PVT1 controls MYC protein levels by protecting the MYC protein from degradation [36]. Specifically, the physical interaction between PVT1 RNA and MYC protein interferes with its phosphorylation at threonine 58, which is essential in leading to MYC protein degradation [36]. As revealed by in vivo chromosome engineering, PVT1 is indispensable for MYC-induced cancer promoting effect [36]. The copy number of PVT1 and MYC gene was co-increased in nearly all CRC cases with MYC gene amplification [36]. Depletion of PVT1 reduced the tumorigenic capacity of HCT116, a MYC-driven CRC cell line [36]. In addition, multiple studies reported significant association between PVT1 expression and CRC malignancies, and pointed to PVT1 as a potential diagnostic and prognostic marker in CRC [36,57]. Interestingly, the PVT1 gene locus also harbors a cluster of six annotated microRNA genes (namely, miR-1204, miR-1205, miR-1206, miR-1207-5p, miR-1207-3p, and miR-1208), of which the function remains to be clarified [58].
4.1.5. PCAT1
The PCAT1 gene is located ~725 kb upstream of the MYC oncogene. Besides the involvement of PCAT1 in prostate cancer [59,60], PCAT1 was also found to be overexpressed in CRC tumors [61]. Experimental data showed that downregulation of PCAT1 inhibits CRC growth in vitro and in vivo, partially via its regulatory effect on MYC [62]. High levels of PCAT1 expression in primary CRC tumors were significantly associated with distal metastasis of CRC. Moreover, multivariable analysis revealed that increased PCAT1 expression was an independent factor for poor prognosis in CRC patients [61].
4.2. CRC Stem Cell—Associated LncRNAs
Wnt signaling is a determining factor in CRC stem cell maintenance [24,63]. It controls not only essential stem cell genes such as LGR5, but also regulates asymmetric division of CRC stem cells [64,65]. The control of CRC stem cells by Wnt signaling offers growth and selection advantages, which may underlie the resistance of CRC tumors to chemotherapeutic drugs [66,67]. Several lncRNAs have been reported to control the fate of the CRC stem cells.
4.2.1. Lnc34a
Loss of miR-34a is commonly seen in many types of cancer [68,69]. Several reports showed that miR-34a directly targets genes involved in Wnt signaling, including Wnt ligands and the essential β-catenin/TCF7L2 components, resulting in suppression of Wnt activity [7,70,71]. Recently, miR-34a was revealed to be directly involved in controlling CRC stem cell asymmetric division by forming a feedforwarded loop targeting Numb and Notch [72]. In an effort to trace the mechanism of reduced miR-34a expression in CRC cells, a new lncRNA termed lnc34a, transcribed in the opposite orientation from the miR-34a, was identified as a key regulator of miR-34a [73]. Lnc34a interacts with several epigenetic regulators, namely, Dnmt3a, HDAC1, and PHB2, to silence the transcription of miR-34a independent of the p53 protein [73]. The functional importance of lnc34a was demonstrated by its enrichment in CRC stem cells, and its ability to initiate asymmetric division by suppressing miR-34a [73]. Interestingly, lnc34a is distributed unevenly during cell division, and represses the transcription of miR-34a in only one daughter cell [73]. Mouse studies proved the function of Lnc34a in regulating self-renewal of cancer stem cell and CRC growth [73]. Concordantly, lnc34a expression was found to be upregulated in clinical samples of late-stage CRCs [73]. This provides an example of lncRNA-miRNA interaction in maintenance of cancer stem cell feature by regulating Wnt and Notch signaling.
4.2.2. RBM5-AS1
The lncRNA RBM5-AS1 was found to be enriched during sphere formation of colon cancer initialing cells [74]. Silencing of RBM5-AS1 impaired Wnt signaling, while overexpression enhances Wnt signaling in CRC cells [74]. The RBM5-AS1 activity on Wnt signaling is critical for enabling the function of CRC stem cells, as loss of RBM5-AS1 impaired the spheroid formation in multiple CRC cell lines [74]. Mechanism study revealed that RBM5-AS1 physically interacts with β-catenin, and promotes the interaction of β-catenin with the TCF7L2 complex [74]. As a result, Wnt target genes such as SGK1, YAP1 and MYC are transcriptionally activated by RBM5-AS1 [74].
4.2.3. WiNTRLINC1
ASCL2 is an essential transcription factor in controlling the stemness of intestinal cells in response to Wnt signaling [65]. Using ChIP-seq with antibodies against RNA polymerase II, WiNTRLINC1 (WNT-regulated lincRNA 1), located ~60 kb away from the ASCL2 gene, was identified as one of the direct β-catenin/TCF7L2 targets in CRC [75]. WiNTRLINC1 physically interacts with β-catenin/TCF7L2 to facilitate the looping of regulatory elements, and consequently activate the transcription of the ASCL2 gene [75]. The regulatory network of Wnt-WiNTRLINC1-ASCL2-stemness is further enhanced with the transcriptional activation of WiNTRLINC1 by ASCL2 [75]. The expression levels of WiNTRLINC1 and ASCL2 were both increased in clinical CRC tumors, and high levels of WiNTRLINC1 were correlated with increased metastatic potential and worse prognosis of CRC patients [75].
4.3. Others
4.3.1. H19
H19 is one of the first imprinted non-coding genes discovered in 1990s [76]. H19 is exclusively transcribed from the maternally inherited allele and participates as a key factor in embryonic development [77,78]. Various reports have suggested the involvement of H19 in human cancer [77,79,80,81]. H19 exerts its function by interaction with EZH2 [82], sponging microRNAs such as let7 and miR-106a [83,84], or production of miR-675 as a primary transcript [85]. Studies from our own work using TCGA CRC data identified H19 as a top candidate in association with worse CRC survival [86]. Knockdown of H19 caused a dramatic reduction of CRC cell proliferation and migration [86]. An unbiased approach with microarray analysis revealed not only the known mechanisms of H19 regulation on let7 and MYC, but also a novel mechanism where H19 regulates β-catenin activity via modulating CDK8 expression [87], which is probably a consequence of H19 interaction with the repressive histone variant macroH2A [86,88]. This study, together with other findings that H19 regulates Wnt signaling by interaction with hnRNP resulting in suppressed expression of Wnt genes in liver cells [89], and by interaction with EZH2, leading to Wnt activation through NKD1 repression in bladder cancer [82], provides a vivid example of diverse mechanisms by a single lncRNA. H19 itself was transcriptionally controlled by the MYC protein, thus forming a connecting loop of Wnt-MYC-H19-Wnt [90]. Multiple studies, including our own work, suggest that H19 is an independent prognostic marker for CRC survival [86]. Combined analysis of H19 with its molecular targets significantly improved the prediction power to a level comparable to stage, validating the clinical significance of CDK8-β-catenin regulation by H19 in CRC [86].
4.3.2. CCAL
LncRNA expression profiling of normal, adenoma, and carcinoma tissues identified CCAL as a crucial regulator of CRC carcinogenesis [66]. In addition, high CCAL levels in the CRC tumor correspond to short overall survival and poor response to adjuvant chemotherapy [66]. The interaction of CCAL with AP-2α protein promotes the degradation of AP-2α, a negative regulator of β-catenin/TCF7L2 interaction in CRC, and thus indirectly activates Wnt signaling [66]. As a consequence, the multidrug resistance (MDR1) gene, which is a Wnt target that encodes the P-glycoprotein 1, is activated [66]. The effect on MDR1 offers a possible explanation on the observed association between CCAL expression and therapeutic outcome in patients with CRC.
4.3.3. CTD903
CTD903 is transcribed from the region of chromosome 14q11.2. Ectopic expression of CTD903 inhibits cell proliferation and cell motility of CRC cells [91]. Cell line model showed that downregulation of CTD903 results in the activation of Wnt/β-catenin signaling, and consequently leads to epithelial mesenchymal transition (EMT), as evidenced by the increase of Twist, Snail, and Vimentin, and reduction of the epithelial marker ZO-1 [91]. This effect of CTD903 on Wnt and EMT provides a possible explanation on the observed suppression of CRC cell invasion by CTD903 [91]. Concordantly, CTD903 predicts the favorable prognosis of CRC patients [91]. The exact molecular mechanism by which CTD903 inhibits Wnt signaling remains to be characterized.
4.3.4. ASBEL
In an effort to identify Wnt-regulated lncRNAs in CRC, ASBEL (a lncRNA also known as BTG3-AS1) was revealed to be a direct target of β-catenin by RNA-seq and ChIP-seq analysis [92]. Knockdown of ASBEL retarded tumor growth in a xenograft mouse model of CRC [92]. Mechanism studies showed that ASBEL forms a complex with TCF3, a transcription factor that is transcriptionally activated by β-catenin, to cooperatively suppress ATF3 gene transcription [92]. This β-catenin-ASBEL-TCF3-ATF3 signaling was demonstrated to be required for CRC proliferation [92]. Together, these data suggest an important role of lncRNA in promoting CRC formation by mediating and executing Wnt activity.
4.3.5. MYU
MYU (c-Myc-upregulated lncRNA), originating from opposite strand of the VPS9D1 gene, is transcriptionally activated by the Wnt target MYC [93]. Downregulation of MYU inhibits the proliferation of MYC-overexpressing cells in vitro and retards xenograft CRC tumor growth in vivo [93]. MYU interacts with the protein hnRNP-K, which binds to the 3′ untranslated region (UTR) of the CDK6 mRNA to prevent the degradation of CDK6 by miR-16 [93]. Consequently, MYU stabilizes CDK6 and controls the cell transition from G1 to S phase [93]. The mechanism of MYC-MYU-hnRNP-K-CDK6 provides another layer of complexity connecting Wnt signaling and CRC growth.
5. Potential Clinical Application
Once dark genome matter, lncRNA emerged as an important layer of regulation of the essential signaling in CRC [94]. Many of the Wnt-associated lncRNAs exhibit significant association with clinical parameters of CRC. Being potential candidates, lncRNAs have several advantages as cancer biomarkers. First, tissue-specific patterns of lncRNA expression suggests that lncRNAs may be over-represented in specific types of tumor, as exemplified by the successful development of PCA3 as cancer biomarker to help determine the necessity for repeated prostate biopsies [95]. Second, lncRNAs are sensitive signaling molecules in response to tissue-specific and context-specific stimuli [32]. Genentech researchers identified CCAT1 as a reliable marker for predicting the response to JQ1, a chemical inhibitor of bromodomain containing 4 (BRD4), suggesting the biomarker potential of CCAT1 in stratifying patients for clinical trials [96]. Third, the fact that some lncRNA genes contain predisposition SNPs offers a unique opportunity for using both DNA and RNA information as biomarkers [31]. As an example, the combined detection of rs6983267 allele status and CCAT2 levels might strengthen the disease connection. Indeed, a large-scale study showed that the preventative effect of aspirin in CRC was affected by the rs6983267 status [97]. We hypothesize that the CCAT2 transcript might work together with its DNA counterpart to regulate nearby genes, and influence the effect of aspirin in preventing CRC. Last, a combination of lncRNA profiling with other RNA or protein expression might improve the biomarker sensitivity or specificity. The finding that levels of H19 and its targets together have a prediction power for CRC prognosis similar to tumor stage supports this possibility, and highlights the importance of mechanism understanding in translational applications [86].
Because of the diverse mechanisms of lncRNAs in fine-tuning Wnt signaling, lncRNAs can be potential therapeutic targets. For instance, the multifaceted function of H19 (sponging let7 [84], regulating methylation at a genome wide level [98], targeting CDK8-β-catenin signaling [86], and regulating the cell cycle [86]) in CRC makes it possible to tackle multiple oncogenic mechanisms with a single hit on this lncRNA. The unique mechanism of PVT1 in controlling MYC protein stability can also be explored for therapeutic modulation of MYC protein, which is hard to inhibit by small molecule chemicals [99]. Similarly, finding out how the transcription of lnc34a is controlled might offer novel insights and strategies to target CRC cancer stem cells.
There are tremendous challenges ahead for the application of the lncRNA in clinical settings: their expression levels are usually low; some intron-derived or nuclear-localized lncRNAs are unstable with a half-life less than 2 h (http://stability.matticklab.com) [100]; their tissue-specific expression pattern may render complexity in the interpretation of profiling by mixed cell population; targeting lncRNAs that reside in the nucleus is still facing many technically difficulties and uncertainties [101]. However, with the technical advances in sequencing and in situ hybridization, it is possible to obtain precise tissue-specificity information and spatial expression patterns. Similarly, with the technical advance on siRNA, antisense oligonucleotides (ASOs), and delivery systems, it is possible to overcome technical difficulties and safety concerns. For instance, most ASOs are taken up by the clearance organs such as liver, and thus, targeting CRC represents unmet challenges [102]. The conjugation of colon specific ligands, similar as the improved distribution seen in the hepatocytes by GalNAc ligand [103], might be one strategy for enhancing the targeting potency of siRNA or ASOs for CRC. Understanding the mechanism such as the effect of protein interactions on cellular uptake of modified ASOs is critical in optimizing the delivery of oligonucleotides [104].
6. Conclusions
As an essential signaling in CRC initiation and progression, Wnt signaling is involved in all key aspects of cancer biology related to tumor growth, metastasis, and therapeutic response. As such, any layer of regulation of this signaling has significance in determining the cancer cell fate, and the outcome of CRC patients. The emerging concept of lncRNAs as important players in CRC by regulating Wnt singling deserves the attention of both academia and the biotechnological industry. Currently, the mechanism of understanding lncRNAs in CRC is still in its early infancy, and this represents one of the bottlenecks in bringing them to clinical applications. We propose that the mechanism of elucidation should be facilitated by unbiased and high-throughput experimental analysis including gene expression microarray analysis, RNA sequencing, methylation profiling, protein interaction identification, genomic occupation by lncRNAs, and others. In addition, large-scale database analysis will reveal the clinical significance of lncRNA, and offer hints on the molecular mechanisms. Finally, more effort should be put into the study of the secondary and higher-order structure of lncRNAs, which is critical for detailed understanding of lncRNA function at a molecular level. We expect that the advances on mechanism of understanding lncRNAs in Wnt signaling might bring novel candidates as biomarkers and therapeutics for CRC. We predict that microdissection and single-cell sequencing might represent a trend in fully extracting gene expression information, and foresee that breakthroughs in using oligonucleotides as drugs in the near future will also help achieve the full potential of lncRNA targeting as cancer therapy.
Acknowledgments
Work in George A. Calin’s laboratory is supported by National Institutes of Health (NIH/NCATS) grant UH3TR00943-01 through the NIH Common Fund, Office of Strategic Coordination (OSC), the NIH/NCI grant 1 R01 CA182905-01, a U54 grant—UPR/MDACC Partnership for Excellence in Cancer Research 2016 Pilot Project, a Team DOD (CA160445P1) grant, a Ladies Leukemia League grant, a CLL Moonshot Flagship project, a SINF 2017 grant, and the Estate of C. G. Johnson, Jr. Martin Pichler was supported by the funding of an Erwin Schroedinger Scholarship of the Austrian Science Funds (No. J3389-B23).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fedewa, S.A.; Ahnen, D.J.; Meester, R.G. S.; Barzi, A.; Jemal, A. Colorectal cancer statistics, 2017. CA Cancer J. Clin. 2017, 67, 177–193. [Google Scholar] [CrossRef] [PubMed]
- Goss, K.H.; Groden, J. Biology of the adenomatous polyposis coli tumor suppressor. J. Clin. Oncol. 2000, 18, 1967–1979. [Google Scholar] [CrossRef] [PubMed]
- Giles, R.H.; van Es, J.H.; Clevers, H. Caught up in a Wnt storm: Wnt signaling in cancer. Biochim. Biophys. Acta 2003, 1653, 1–24. [Google Scholar] [CrossRef]
- Groden, J.; Thliveris, A.; Samowitz, W.; Carlson, M.; Gelbert, L.; Albertsen, H.; Joslyn, G.; Stevens, J.; Spirio, L.; Robertson, M.; et al. Identification and characterization of the familial adenomatous polyposis coli gene. Cell 1991, 66, 589–600. [Google Scholar] [CrossRef]
- Nagel, R.; le Sage, C.; Diosdado, B.; van der Waal, M.; Oude Vrielink, J.A.; Bolijn, A.; Meijer, G.A.; Agami, R. Regulation of the adenomatous polyposis coli gene by the miR-135 family in colorectal cancer. Cancer Res. 2008, 68, 5795–5802. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, H.S.; Kim, N.G.; Lee, I.; Choi, H.S.; Li, X.Y.; Kang, S.E.; Cha, S.Y.; Ryu, J.K.; Na, J.M.; et al. p53 and microRNA-34 are suppressors of canonical Wnt signaling. Sci. Signal. 2011, 4, ra71. [Google Scholar] [CrossRef] [PubMed]
- Hwang, W.L.; Jiang, J.K.; Yang, S.H.; Huang, T.S.; Lan, H.Y.; Teng, H.W.; Yang, C.Y.; Tsai, Y.P.; Lin, C.H.; Wang, H.W.; et al. MicroRNA-146a directs the symmetric division of Snail-dominant colorectal cancer stem cells. Nat. Cell Biol. 2014, 16, 268–280. [Google Scholar] [CrossRef] [PubMed]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Smolle, M.; Uranitsch, S.; Gerger, A.; Pichler, M.; Haybaeck, J. Current status of long non-coding RNAs in human cancer with specific focus on colorectal cancer. Int. J. Mol. Sci. 2014, 15, 13993–14013. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Varmus, H.E. Many tumors induced by the mouse mammary tumor virus contain a provirus integrated in the same region of the host genome. Cell 1982, 31, 99–109. [Google Scholar] [CrossRef]
- Rijsewijk, F.; Schuermann, M.; Wagenaar, E.; Parren, P.; Weigel, D.; Nusse, R. The Drosophila homolog of the mouse mammary oncogene int-1 is identical to the segment polarity gene wingless. Cell 1987, 50, 649–657. [Google Scholar] [CrossRef]
- Kikuchi, A.; Yamamoto, H.; Sato, A.; Matsumoto, S. New insights into the mechanism of Wnt signaling pathway activation. Int. Rev. Cell Mol. Biol. 2011, 291, 21–71. [Google Scholar] [PubMed]
- Hart, M.J.; de los Santos, R.; Albert, I.N.; Rubinfeld, B.; Polakis, P. Downregulation of β-catenin by human Axin and its association with the APC tumor suppressor, β-catenin and GSK3β. Curr. Biol. 1998, 8, 573–581. [Google Scholar] [CrossRef]
- Kishida, S.; Yamamoto, H.; Ikeda, S.; Kishida, M.; Sakamoto, I.; Koyama, S.; Kikuchi, A. Axin, a negative regulator of the Wnt signaling pathway, directly interacts with adenomatous polyposis coli and regulates the stabilization of β-catenin. J. Biol. Chem. 1998, 273, 10823–10826. [Google Scholar] [CrossRef] [PubMed]
- Brannon, M.; Gomperts, M.; Sumoy, L.; Moon, R.T.; Kimelman, D. A β-catenin/XTcf-3 complex binds to the siamois promoter to regulate dorsal axis specification in Xenopus. Genes Dev. 1997, 11, 2359–2370. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas, N. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337. [Google Scholar]
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. APC mutations occur early during colorectal tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Burt, R.W.; Leppert, M.F.; Slattery, M.L.; Samowitz, W.S.; Spirio, L.N.; Kerber, R.A.; Kuwada, S.K.; Neklason, D.W.; Disario, J.A.; Lyon, E.; et al. Genetic testing and phenotype in a large kindred with attenuated familial adenomatous polyposis. Gastroenterology 2004, 127, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Morin, P.J.; Sparks, A.B.; Korinek, V.; Barker, N.; Clevers, H.; Vogelstein, B.; Kinzler, K.W. Activation of β-catenin-Tcf signaling in colon cancer by mutations in β-catenin or APC. Science 1997, 275, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Korinek, V.; Barker, N.; Morin, P.J.; van Wichen, D.; de Weger, R.; Kinzler, K.W.; Vogelstein, B.; Clevers, H. Constitutive transcriptional activation by a β-catenin-Tcf complex in APC−/− colon carcinoma. Science 1997, 275, 1784–1787. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, L.; de Sousa, E.M.F.; van der Heijden, M.; Cameron, K.; de Jong, J.H.; Borovski, T.; Tuynman, J.B.; Todaro, M.; Merz, C.; Rodermond, H.; et al. Wnt activity defines colon cancer stem cells and is regulated by the microenvironment. Nat. Cell Biol. 2010, 12, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Dow, L.E.; O’Rourke, K.P.; Simon, J.; Tschaharganeh, D.F.; van Es, J.H.; Clevers, H.; Lowe, S.W. Apc Restoration Promotes Cellular Differentiation and Reestablishes Crypt Homeostasis in Colorectal Cancer. Cell 2015, 161, 1539–1552. [Google Scholar] [CrossRef] [PubMed]
- Tetsu, O.; McCormick, F. β-catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [PubMed]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Vincent, K.; Pichler, M.; Fodde, R.; Berindan-Neagoe, I.; Slack, F.J.; Calin, G.A. Junk DNA and the long non-coding RNA twist in cancer genetics. Oncogene 2015, 34, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Beroukhim, R.; Mermel, C.H.; Porter, D.; Wei, G.; Raychaudhuri, S.; Donovan, J.; Barretina, J.; Boehm, J.S.; Dobson, J.; Urashima, M.; et al. The landscape of somatic copy-number alteration across human cancers. Nature 2010, 463, 899–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Qi, H.; Luo, F.; Xu, H.; Ling, M.; Qin, Y.; Yang, P.; Liu, X.; Yang, Q.; Xue, J.; et al. Feedback circuitry via let-7c between lncRNA CCAT1 and c-Myc is involved in cigarette smoke extract-induced malignant transformation of HBE cells. Oncotarget 2017, 8, 19285–19297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhou, C.; Chang, Y.; Zhang, Z.; Hu, Y.; Zhang, F.; Lu, Y.; Zheng, L.; Zhang, W.; Li, X.; Li, X. Long non-coding RNA CASC11 interacts with hnRNP-K and activates the WNT/β-catenin pathway to promote growth and metastasis in colorectal cancer. Cancer Lett. 2016, 376, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Cui, R.; Jeon, Y.J.; Lee, J.H.; Lee, J.H.; Sim, H.; Park, J.K.; Fadda, P.; Tili, E.; Nakanishi, H.; et al. Long-range interaction and correlation between MYC enhancer and oncogenic long noncoding RNA CARLo-5. Proc. Natl. Acad. Sci. USA 2014, 111, 4173–4178. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafa, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, N.; Penn, L.Z. Reflecting on 25 years with MYC. Nat. Rev. Cancer 2008, 8, 976–990. [Google Scholar] [CrossRef] [PubMed]
- Tenesa, A.; Farrington, S.M.; Prendergast, J.G.; Porteous, M.E.; Walker, M.; Haq, N.; Barnetson, R.A.; Theodoratou, E.; Cetnarskyj, R.; Cartwright, N.; et al. Genome-wide association scan identifies a colorectal cancer susceptibility locus on 11q23 and replicates risk loci at 8q24 and 18q21. Nat. Genet. 2008, 40, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, I.; Webb, E.; Carvajal-Carmona, L.; Broderick, P.; Kemp, Z.; Spain, S.; Penegar, S.; Chandler, I.; Gorman, M.; Wood, W.; et al. A genome-wide association scan of tag SNPs identifies a susceptibility variant for colorectal cancer at 8q24.21. Nat. Genet. 2007, 39, 984–988. [Google Scholar] [CrossRef] [PubMed]
- Zanke, B.W.; Greenwood, C.M.; Rangrej, J.; Kustra, R.; Tenesa, A.; Farrington, S.M.; Prendergast, J.; Olschwang, S.; Chiang, T.; Crowdy, E.; et al. Genome-wide association scan identifies a colorectal cancer susceptibility locus on chromosome 8q24. Nat. Genet. 2007, 39, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, M.M.; Ahmadiyeh, N.; Jia, L.; Herman, P.; Verzi, M.P.; Doddapaneni, H.; Beckwith, C.A.; Chan, J.A.; Hills, A.; Davis, M.; et al. The 8q24 cancer risk variant rs6983267 shows long-range interaction with MYC in colorectal cancer. Nat. Genet. 2009, 41, 882–884. [Google Scholar] [CrossRef] [PubMed]
- Tuupanen, S.; Turunen, M.; Lehtonen, R.; Hallikas, O.; Vanharanta, S.; Kivioja, T.; Bjorklund, M.; Wei, G.; Yan, J.; Niittymaki, I.; et al. The common colorectal cancer predisposition SNP rs6983267 at chromosome 8q24 confers potential to enhanced Wnt signaling. Nat. Genet. 2009, 41, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Hnisz, D.; Abraham, B.J.; Lee, T.I.; Lau, A.; Saint-Andre, V.; Sigova, A.A.; Hoke, H.A.; Young, R.A. Super-enhancers in the control of cell identity and disease. Cell 2013, 155, 934–947. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Wu, F.; Gao, F.; Qing, X.; Shao, Z. Prognostic value of long non-coding RNA CCAT1 expression in patients with cancer: A meta-analysis. PLoS ONE 2017, 12, e0179346. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.F.; Yin, Q.F.; Chen, T.; Zhang, Y.; Zhang, X.O.; Wu, Z.; Zhang, S.; Wang, H.B.; Ge, J.; Lu, X.; et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014, 24, 513–531. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Liu, C.G.; Ferracin, M.; Hyslop, T.; Spizzo, R.; Sevignani, C.; Fabbri, M.; Cimmino, A.; Lee, E.J.; Wojcik, S.E.; et al. Ultraconserved regions encoding ncRNAs are altered in human leukemias and carcinomas. Cancer Cell 2007, 12, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Haiman, C.A.; Le Marchand, L.; Yamamato, J.; Stram, D.O.; Sheng, X.; Kolonel, L.N.; Wu, A.H.; Reich, D.; Henderson, B.E. A common genetic risk factor for colorectal and prostate cancer. Nat. Genet. 2007, 39, 954–956. [Google Scholar] [CrossRef] [PubMed]
- Yeager, M.; Orr, N.; Hayes, R.B.; Jacobs, K.B.; Kraft, P.; Wacholder, S.; Minichiello, M.J.; Fearnhead, P.; Yu, K.; Chatterjee, N.; et al. Genome-wide association study of prostate cancer identifies a second risk locus at 8q24. Nat. Genet. 2007, 39, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Sur, I.K.; Hallikas, O.; Vaharautio, A.; Yan, J.; Turunen, M.; Enge, M.; Taipale, M.; Karhu, A.; Aaltonen, L.A.; Taipale, J. Mice lacking a Myc enhancer that includes human SNP rs6983267 are resistant to intestinal tumors. Science 2012, 338, 1360–1363. [Google Scholar] [CrossRef] [PubMed]
- Redis, R.S.; Vela, L.E.; Lu, W.; Ferreira de Oliveira, J.; Ivan, C.; Rodriguez-Aguayo, C.; Adamoski, D.; Pasculli, B.; Taguchi, A.; Chen, Y.; et al. Allele-Specific Reprogramming of Cancer Metabolism by the Long Non-coding RNA CCAT2. Mol. Cell 2016, 61, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Liang, H.; Cui, X.; Han, C.; Hao, C.; Huo, K. Long noncoding RNA CCAT2 can predict metastasis and a poor prognosis: A meta-analysis. Clin. Chim. Acta 2017, 468, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Matsuyama, T.; Toiyama, Y.; Takahashi, N.; Ishikawa, T.; Uetake, H.; Yamada, Y.; Kusunoki, M.; Calin, G.; Goel, A. CCAT1 and CCAT2 long noncoding RNAs, located within the 8q.24.21 ’gene desert’, serve as important prognostic biomarkers in colorectal cancer. Ann. Oncol. 2017, 28, 1882–1888. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.H.; Fang, H.; Ji, C.X.; Xie, H.; Xiao, B.; Zhu, X.G. Long noncoding RNA CCAT2 can predict metastasis and poor prognosis: A meta-analysis. Clin. Chim. Acta 2017, 466, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Hou, Y.C.; Fu, L.N.; Wang, Y.Q.; Liu, Q.Q.; Xiong, H.; Chen, Y.X.; Fang, J.Y. Long Noncoding RNA CCAT2 as a Potential Novel Biomarker to Predict the Clinical Outcome of Cancer Patients: A Meta-Analysis. J. Cancer 2017, 8, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Sawada, G.; Kurashige, J.; Uchi, R.; Matsumura, T.; Ueo, H.; Takano, Y.; Eguchi, H.; Sudo, T.; Sugimachi, K.; et al. Amplification of PVT-1 is involved in poor prognosis via apoptosis inhibition in colorectal cancers. Br. J. Cancer 2014, 110, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Huppi, K.; Volfovsky, N.; Runfola, T.; Jones, T.L.; Mackiewicz, M.; Martin, S.E.; Mushinski, J.F.; Stephens, R.; Caplen, N.J. The identification of microRNAs in a genomically unstable region of human chromosome 8q24. Mol. Cancer Res. 2008, 6, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Prensner, J.R.; Iyer, M.K.; Balbin, O.A.; Dhanasekaran, S.M.; Cao, Q.; Brenner, J.C.; Laxman, B.; Asangani, I.A.; Grasso, C.S.; Kominsky, H.D.; et al. Transcriptome sequencing across a prostate cancer cohort identifies PCAT-1, an unannotated lincRNA implicated in disease progression. Nat. Biotechnol. 2011, 29, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Ahmed, M.; Zhang, F.; Yao, C.Q.; Li, S.; Liang, Y.; Hua, J.; Soares, F.; Sun, Y.; Langstein, J.; et al. Modulation of long noncoding RNAs by risk SNPs underlying genetic predispositions to prostate cancer. Nat. Genet. 2016, 48, 1142–1150. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Chen, Y.; Liao, X.; Liu, D.; Li, F.; Ruan, H.; Jia, W. Overexpression of long noncoding RNA PCAT-1 is a novel biomarker of poor prognosis in patients with colorectal cancer. Med. Oncol. 2013, 30, 588. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Liu, X.; Tang, Y.; Zhao, Z.; Zhang, J.; Feng, Y. Down regulation of the long non-coding RNA PCAT-1 induced growth arrest and apoptosis of colorectal cancer cells. Life Sci. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ordonez-Moran, P.; Dafflon, C.; Imajo, M.; Nishida, E.; Huelsken, J. HOXA5 Counteracts Stem Cell Traits by Inhibiting Wnt Signaling in Colorectal Cancer. Cancer Cell 2015, 28, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Es, J.H.; Kuipers, J.; Kujala, P.; van den Born, M.; Cozijnsen, M.; Haegebarth, A.; Korving, J.; Begthel, H.; Peters, P.J.; Clevers, H. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature 2007, 449, 1003–1007. [Google Scholar] [CrossRef] [PubMed]
- Van der Flier, L.G.; van Gijn, M.E.; Hatzis, P.; Kujala, P.; Haegebarth, A.; Stange, D.E.; Begthel, H.; van den Born, M.; Guryev, V.; Oving, I.; et al. Transcription factor achaete scute-like 2 controls intestinal stem cell fate. Cell 2009, 136, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Yang, Y.; Wang, F.; Moyer, M.P.; Wei, Q.; Zhang, P.; Yang, Z.; Liu, W.; Zhang, H.; Chen, N.; et al. Long non-coding RNA CCAL regulates colorectal cancer progression by activating Wnt/β-catenin signaling pathway via suppression of activator protein 2α. Gut 2016, 65, 1494–1504. [Google Scholar] [CrossRef] [PubMed]
- Kendziorra, E.; Ahlborn, K.; Spitzner, M.; Rave-Frank, M.; Emons, G.; Gaedcke, J.; Kramer, F.; Wolff, H.A.; Becker, H.; Beissbarth, T.; et al. Silencing of the Wnt transcription factor TCF4 sensitizes colorectal cancer cells to (chemo-) radiotherapy. Carcinogenesis 2011, 32, 1824–1831. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Ren, Z.J.; Tang, J.H. MicroRNA-34a: A potential therapeutic target in human cancer. Cell Death Dis. 2014, 5, e1327. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Hashimi, S.T.; Fulcher, J.A.; Chang, M.H.; Gov, L.; Wang, S.; Lee, B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood 2009, 114, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Fu, H.; Liu, Q.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Downregulation of CCND1 and CDK6 by miR-34a induces cell cycle arrest. FEBS Lett. 2008, 582, 1564–1568. [Google Scholar] [CrossRef] [PubMed]
- Bu, P.; Wang, L.; Chen, K.Y.; Srinivasan, T.; Murthy, P.K.; Tung, K.L.; Varanko, A.K.; Chen, H.J.; Ai, Y.; King, S.; et al. A miR-34a-Numb Feedforward Loop Triggered by Inflammation Regulates Asymmetric Stem Cell Division in Intestine and Colon Cancer. Cell Stem Cell 2016, 18, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bu, P.; Ai, Y.; Srinivasan, T.; Chen, H.J.; Xiang, K.; Lipkin, S.M.; Shen, X. A long non-coding RNA targets microRNA miR-34a to regulate colon cancer stem cell asymmetric division. Elife 2016, 5, e14620. [Google Scholar] [CrossRef] [PubMed]
- Di Cecilia, S.; Zhang, F.; Sancho, A.; Li, S.; Aguilo, F.; Sun, Y.; Rengasamy, M.; Zhang, W.; del Vecchio, L.; Salvatore, F.; et al. RBM5-AS1 Is Critical for Self-Renewal of Colon Cancer Stem-like Cells. Cancer Res. 2016, 76, 5615–5627. [Google Scholar] [CrossRef] [PubMed]
- Giakountis, A.; Moulos, P.; Zarkou, V.; Oikonomou, C.; Harokopos, V.; Hatzigeorgiou, A.G.; Reczko, M.; Hatzis, P. A Positive Regulatory Loop between a Wnt-Regulated Non-coding RNA and ASCL2 Controls Intestinal Stem Cell Fate. Cell Rep. 2016, 15, 2588–2596. [Google Scholar] [CrossRef] [PubMed]
- Brannan, C.I.; Dees, E.C.; Ingram, R.S.; Tilghman, S.M. The product of the H19 gene may function as an RNA. Mol. Cell. Biol. 1990, 10, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shields, T.; Crenshaw, T.; Hao, Y.; Moulton, T.; Tycko, B. Imprinting of human H19: Allele-specific CpG methylation, loss of the active allele in Wilms tumor, and potential for somatic allele switching. Am. J. Hum. Genet. 1993, 53, 113–124. [Google Scholar] [PubMed]
- Zhang, Y.; Tycko, B. Monoallelic expression of the human H19 gene. Nat. Genet. 1992, 1, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Raveh, E.; Matouk, I.J.; Gilon, M.; Hochberg, A. The H19 Long non-coding RNA in cancer initiation, progression and metastasis—A proposed unifying theory. Mol. Cancer 2015, 14, 184. [Google Scholar] [CrossRef] [PubMed]
- Medrzycki, M.; Zhang, Y.; Zhang, W.; Cao, K.; Pan, C.; Lailler, N.; McDonald, J.F.; Bouhassira, E.E.; Fan, Y. Histone H1.3 suppresses H19 noncoding RNA expression and cell growth of ovarian cancer cells. Cancer Res. 2014, 74, 6463–6473. [Google Scholar] [CrossRef] [PubMed]
- Moulton, T.; Crenshaw, T.; Hao, Y.; Moosikasuwan, J.; Lin, N.; Dembitzer, F.; Hensle, T.; Weiss, L.; McMorrow, L.; Loew, T.; et al. Epigenetic lesions at the H19 locus in Wilms’ tumour patients. Nat. Genet. 1994, 7, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lin, Y.; Yang, X.; Wu, X.; He, X. Long noncoding RNA H19 regulates EZH2 expression by interacting with miR-630 and promotes cell invasion in nasopharyngeal carcinoma. Biochem. Biophys. Res. Commun. 2016, 473, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.; Brunschweiger, A.; Brummer, A.; Guennewig, B.; Mittal, N.; Kishore, S.; Tsikrika, P.; Gerber, A.P.; Zavolan, M.; Hall, J. miR-CLIP capture of a miRNA targetome uncovers a lincRNA H19-miR-106a interaction. Nat. Chem. Biol. 2015, 11, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Kallen, A.N.; Zhou, X.B.; Xu, J.; Qiao, C.; Ma, J.; Yan, L.; Lu, L.; Liu, C.; Yi, J.S.; Zhang, H.; et al. The imprinted H19 lncRNA antagonizes Let-7 microRNAs. Mol. Cell 2013, 52, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuka, M.; Ling, H.; Ivan, C.; Pichler, M.; Matsushita, D.; Goblirsch, M.; Stiegelbauer, V.; Shigeyasu, K.; Zhang, X.; Chen, M.; et al. H19 Noncoding RNA, an Independent Prognostic Factor, Regulates Essential Rb-E2F and CDK8-β-Catenin Signaling in Colorectal Cancer. EBioMedicine 2016, 13, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Firestein, R.; Bass, A.J.; Kim, S.Y.; Dunn, I.F.; Silver, S.J.; Guney, I.; Freed, E.; Ligon, A.H.; Vena, N.; Ogino, S.; et al. CDK8 is a colorectal cancer oncogene that regulates β-catenin activity. Nature 2008, 455, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Goldberg, M.S.; Cumberland, L.K.; Ratnakumar, K.; Segura, M.F.; Emanuel, P.O.; Menendez, S.; Vardabasso, C.; Leroy, G.; Vidal, C.I.; et al. The histone variant macroH2A suppresses melanoma progression through regulation of CDK8. Nature 2010, 468, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, X.; Liu, Y.; Yuan, J.; Yang, F.; Huang, J.; Meng, Q.; Zhou, C.; Liu, F.; Ma, J.; et al. Long noncoding RNA H19 inhibits the proliferation of fetal liver cells and the Wnt signaling pathway. FEBS Lett. 2016, 590, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Barsyte-Lovejoy, D.; Lau, S.K.; Boutros, P.C.; Khosravi, F.; Jurisica, I.; Andrulis, I.L.; Tsao, M.S.; Penn, L.Z. The c-Myc oncogene directly induces the H19 noncoding RNA by allele-specific binding to potentiate tumorigenesis. Cancer Res. 2006, 66, 5330–5337. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Yu, X.; Ni, B.; Chen, D.; Yang, Z.; Huang, J.; Wang, J.; Chen, D.; Wang, L. Overexpression of long non-coding RNA-CTD903 inhibits colorectal cancer invasion and migration by repressing Wnt/β-catenin signaling and predicts favorable prognosis. Int. J. Oncol. 2016, 48, 2675–2685. [Google Scholar] [CrossRef] [PubMed]
- Taniue, K.; Kurimoto, A.; Takeda, Y.; Nagashima, T.; Okada-Hatakeyama, M.; Katou, Y.; Shirahige, K.; Akiyama, T. ASBEL-TCF3 complex is required for the tumorigenicity of colorectal cancer cells. Proc. Natl. Acad. Sci. USA 2016, 113, 12739–12744. [Google Scholar]
- Kawasaki, Y.; Komiya, M.; Matsumura, K.; Negishi, L.; Suda, S.; Okuno, M.; Yokota, N.; Osada, T.; Nagashima, T.; Hiyoshi, M.; et al. MYU, a Target lncRNA for Wnt/c-Myc Signaling, Mediates Induction of CDK6 to Promote Cell Cycle Progression. Cell Rep. 2016, 16, 2554–2564. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Girnita, L.; Buda, O.; Calin, G.A. Non-coding RNAs: The cancer genome dark matter that matters! Clin. Chem. Lab. Med. 2017, 55, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Rittenhouse, H.; Blase, A.; Shamel, B.; Schalken, J.; Groskopf, J. The long and winding road to FDA approval of a novel prostate cancer test: Our story. Clin. Chem. 2013, 59, 32–34. [Google Scholar] [CrossRef] [PubMed]
- McCleland, M.L.; Mesh, K.; Lorenzana, E.; Chopra, V.S.; Segal, E.; Watanabe, C.; Haley, B.; Mayba, O.; Yaylaoglu, M.; Gnad, F.; et al. CCAT1 is an enhancer-templated RNA that predicts BET sensitivity in colorectal cancer. J. Clin. Investig. 2016, 126, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Nan, H.; Morikawa, T.; Suuriniemi, M.; Imamura, Y.; Werner, L.; Kuchiba, A.; Yamauchi, M.; Hunter, D.J.; Kraft, P.; Giovannucci, E.L.; et al. Aspirin use, 8q24 single nucleotide polymorphism rs6983267, and colorectal cancer according to CTNNB1 alterations. J. Natl. Cancer Inst. 2013, 105, 1852–1861. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yang, L.; Zhong, T.; Mueller, M.; Men, Y.; Zhang, N.; Xie, J.; Giang, K.; Chung, H.; Sun, X.; et al. H19 lncRNA alters DNA methylation genome wide by regulating S-adenosylhomocysteine hydrolase. Nat. Commun. 2015, 6, 10221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dang, C.V.; Reddy, E.P.; Shokat, K.M.; Soucek, L. Drugging the ‘undruggable’ cancer targets. Nat. Rev. Cancer 2017, 17, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.B.; Johnston, R.L.; Inostroza-Ponta, M.; Fox, A.H.; Fortini, E.; Moscato, P.; Dinger, M.E.; Mattick, J.S. Genome-wide analysis of long noncoding RNA stability. Genome Res. 2012, 22, 885–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, H.; Fabbri, M.; Calin, G.A. MicroRNAs and other non-coding RNAs as targets for anticancer drug development. Nat. Rev. Drug Discov. 2013, 12, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Geary, R.S.; Norris, D.; Yu, R.; Bennett, C.F. Pharmacokinetics, biodistribution and cell uptake of antisense oligonucleotides. Adv. Drug Deliv. Rev. 2015, 87, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Prakash, T.P.; Graham, M.J.; Yu, J.; Carty, R.; Low, A.; Chappell, A.; Schmidt, K.; Zhao, C.; Aghajan, M.; Murray, H.F.; et al. Targeted delivery of antisense oligonucleotides to hepatocytes using triantennary N-acetyl galactosamine improves potency 10-fold in mice. Nucleic Acids Res. 2014, 42, 8796–8807. [Google Scholar] [CrossRef] [PubMed]
- Crooke, S.T.; Wang, S.; Vickers, T.A.; Shen, W.; Liang, X.H. Cellular uptake and trafficking of antisense oligonucleotides. Nat. Biotechnol. 2017, 35, 230–237. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Long non-coding RNA (LncRNA) involvement in canonical WNT signaling. Arrow in black: activate; T in Red: inhibit.
Figure 1.
Long non-coding RNA (LncRNA) involvement in canonical WNT signaling. Arrow in black: activate; T in Red: inhibit.

Figure 2.
8q24 lncRNAs in MYC regulation.

{kind=link}
{kind=link}
Table 1.
LncRNAs involved in Wnt signaling and CRC.
| LncRNA | Loci | Length | Indentification Method | Mechanism Related to Wnt | Clinical Relevance |
|---|---|---|---|---|---|
| 8q24 region | |||||
| CCAT1-S (CARLo-5) | 8q24.21 | 2628 nt | RACE qRT-PCR | Transcription of CCAT1-S is influenced by the allele status of the single nucleotide polymorphisms (SNP) rs6983267 via a long-range interaction of CCAT1-S promoter with rs6938267-containing region. | Promotes CRC growth and invasion; Increased expression correlates with poor prognosis. |
| CCAT1-L | 8q24.21 | 5200 nt | RNA-seq qRT-PCR Northern blot RACE | Interacts with CTCF to faciliate chromatin looping connecting MYC enhancer and promoter, resulting in MYC transcription. | |
| CCAT2 | 8q24.21 | 340 nt | qRT-PCR Northern blot RACE | Interacts with TCF7L2 to promote MYC and other Wnt target gene transcription. Spans the SNP rs6983267 alleles that responds differentially to Wnt signaling. | Promotes CRC growth and metastasis; Increased expression correlates with poor prognosis. |
| CASC11 (CARLo-7) | 8q24.21 | 872 nt | qRT-PCR | Interacts with heterogeneous ribonucleoprotein K (hnRNP-K) to protects β-catenin from degradation, and consequently activates Wnt signaling. MYC binds to the promoter of CASC11 to activate its transcription. | Promotes CRC growth and invasion; Increased expression correlates with CRC size, invasion, and lymph metastasis. |
| PVT1 | 8q24.21 | 1957 nt | RACE Northern blot qRT-PCR | Interacts with MYC protein to prevent MYC phosphorylation and degradation. | Promotes CRC growth. Increased expression correlates with poor prognosis. |
| PCAT1 | 8q24.21 | 1992 nt | qRT-PCR, RNA-seq | Increases MYC expression. | Promotes CRC growth. |
| CRC stem cell | |||||
| Lnc34a | 1p36.22? | 693 nt | qRT-PCR RACE | Interacts with Dnmt3a, HDAC1, and PHB2E to epigenetically silences miR-34a expression, resulting in CRC stem cell asymmetric division. | Enriched in CRC stem cells, and upregulated in late-stage CRCs. |
| RBM5-AS1 | 3p21.31 | 1386 nt | lncRNA array RNA-Seq qRT-PCR | Interacts with β-catenin, and promotes the transcriptional activity of β-catenin/TCF7L2 complex. | |
| WiNTRLINC1 | 11p15.5 | 4117 nt | qRT-PCR Northern blot | Interacts with TCF7L2/β-catenin to form chromatin loop and activate ASCL2 transcription. | Increased expression correlates with metastatic potential and poor prognosis. |
| Others | |||||
| H19 | 11q15.5 | 6295 nt | RACE cloning Northern blot qRT-PCR | Interacts with macroH2A to derepress transcription of CDK8, which positively regulates β-catenin activity. Interacts with hnRNP U to repress Wnt gene transcription. Interacts with EZH2 to repress NKD1 transcription, resulting in Wnt activation. Antagonizes the inhibition of let7 on MYC, which regulates H19 transcription. | Increased expression correlates with poor prognosis independent of other factors. |
| CCAL | Chr3 | 1933 nt | Microarray RACE qRT-PCR | Interacts with and degrades AP-2α, a negative regulator of Wnt activity, resulting in increased MDR1 transcription. | Increased expression correlates with poor prognosis and poor response to adjuvant chemotherapy |
| CTD903 | 14q11.2 | 903 nt | Microarray qRT-PCR | Inhibits Wnt signaling and EMT by unknown mechanisms. | Increased expression correlates with favorable prognosis. |
| ASBEL | 21q21.1 | 2000 nt | qRT-PCR Northern blot | Interacts with TCF3 to repress ATF3 transcription. | |
| MYU | 16q24.3 | 6310 nt | RNA-seq | Upregulated by MYC. Interacts with hnRNP-K to stabilize CDK6 mRNA. |
* RACE: Rapid amplification of cDNA ends; CTCF: CCCTC-binding factor.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, P.; Pichler, M.; Chen, M.; Calin, G.A.; Ling, H. To Wnt or Lose: The Missing Non-Coding Linc in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18092003
AMA Style
Shen P, Pichler M, Chen M, Calin GA, Ling H. To Wnt or Lose: The Missing Non-Coding Linc in Colorectal Cancer. International Journal of Molecular Sciences. 2017; 18(9):2003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18092003
Chicago/Turabian StyleShen, Peng, Martin Pichler, Meng Chen, George A. Calin, and Hui Ling. 2017. "To Wnt or Lose: The Missing Non-Coding Linc in Colorectal Cancer" International Journal of Molecular Sciences 18, no. 9: 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18092003
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.