“Vessels in the Storm”: Searching for Prognostic and Predictive Angiogenic Factors in Colorectal Cancer

 , , , and
, , , and
Abstract
:
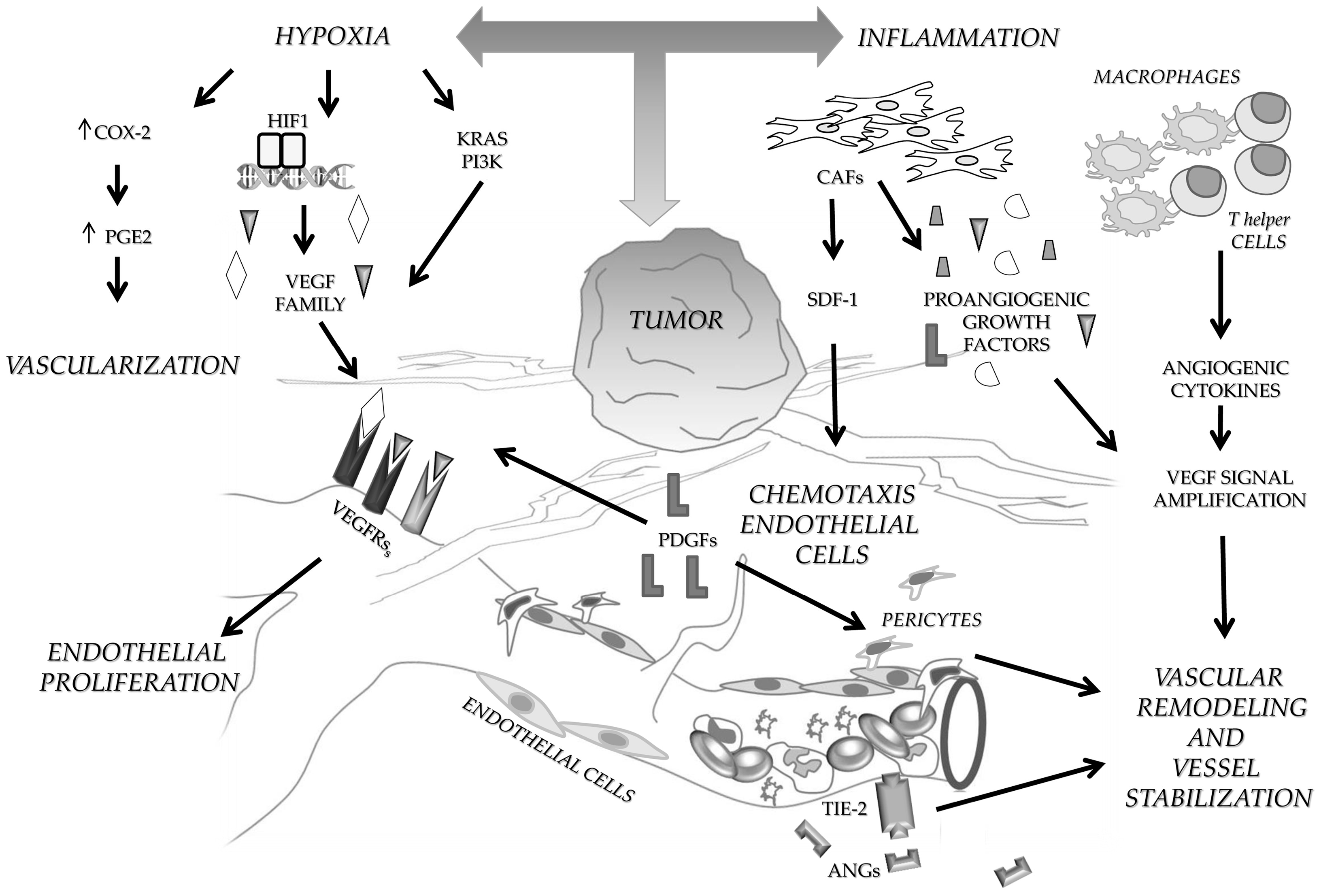
1. Angiogenesis in Colon Cancer
1.1. Angiogenic Growth Factors
1.1.1. VEGF-A
1.1.2. PlGF
1.1.3. ANGs
1.1.4. FGFs
1.1.5. PDGFs
1.2. Tumor Microenvironment and Angiogenesis
1.2.1. TAMs
1.2.2. CAFs
1.2.3. MDSCs and Lymphocytes
1.3. Angiogenesis Induced by Oncogenic Signaling
1.3.1. TGF-β1
1.3.2. WNT
1.3.3. KRAS
2. Targeting Angiogenesis in CRC: Current Clinical Outlook
2.1. Bevacizumab
2.2. Aflibercept
2.3. Vanucizumab
2.4. Regorafenib
2.5. TAS 102
2.6. Vandetanib
2.7. Nintedanib
2.8. Trebanabib
2.9. Vorinostat
2.10. Fruquintinib
2.11. Famitinib
3. Angiogenesis-Related Prognostic Biomarkers in CRC
3.1. VEGF Signaling
3.2. ANG Signaling
3.3. PDGF Signaling
3.4. FGF Signaling
3.5. TGF-β Signaling
3.6. Non-Coding RNA Signaling
3.7. Other Signaling Pathways
4. Response Biomarkers in Anti-Angiogenesis Therapy of CRC
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Kondo, Y.; Arii, S.; Furutani, M.; Isigami, S.; Mori, A.; Onodera, H.; Chiba, T.; Imamura, M. Implication of vascular endothelial growth factor and p53 status for angiogenesis in noninvasive colorectal carcinoma. Cancer 2000, 88, 1820–1827. [Google Scholar] [CrossRef]
- Kondo, Y.; Arii, S.; Mori, A.; Furutani, M.; Chiba, T.; Imamura, M. Enhancement of angiogenesis, tumor growth, and metastasis by transfection of vascular endothelial growth factor into LoVo human colon cancer cell line. Clin. Cancer Res. 2000, 6, 622–630. [Google Scholar] [PubMed]
- Takahashi, Y.; Kitadai, Y.; Bucana, C.D.; Cleary, K.R.; Ellis, L.M. Expression of vascular endothelial growth factor and its receptor, KDR, correlates with vascularity, metastasis, and proliferation of human colon cancer. Cancer Res. 1995, 55, 3964–3968. [Google Scholar] [PubMed]
- Hanrahan, V.; Currie, M.J.; Gunningham, S.P.; Morrin, H.R.; Scott, P.A.; Robinson, B.A.; Fox, S.B. The angiogenic switch for vascular endothelial growth factor (VEGF)-A, VEGF-B, VEGF-C, and VEGF-D in the adenoma-carcinoma sequence during colorectal cancer progression. J. Pathol. 2003, 200, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; De Marzo, A.M.; Laughner, E.; Lim, M.; Hilton, D.A.; Zagzag, D.; Buechler, P.; Isaacs, W.B.; Semenza, G.L.; Simons, J.W. Overexpression of hypoxia-inducible factor 1α in common human cancers and their metastases. Cancer Res. 1999, 59, 5830–5835. [Google Scholar] [PubMed]
- Ioannou, M.; Paraskeva, E.; Baxevanidou, K.; Simos, G.; Papamichali, R.; Papacharalambous, C.; Samara, M.; Koukoulis, G. HIF-1α in colorectal carcinoma: Review of the literature. J. Buon 2015, 20, 680–689. [Google Scholar] [PubMed]
- Cao, D.; Hou, M.; Guan, Y.-S.; Jiang, M.; Yang, Y.; Gou, H.-F. Expression of HIF-1α and VEGF in colorectal cancer: Association with clinical outcomes and prognostic implications. BMC Cancer 2009, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Tsujii, M.; Kawano, S.; Tsuji, S.; Sawaoka, H.; Hori, M.; DuBois, R.N. Cyclooxygenase Regulates Angiogenesis Induced by Colon Cancer Cells. Cell 1998, 93, 705–716. [Google Scholar] [CrossRef]
- Kaidi, A.; Qualtrough, D.; Williams, A.C.; Paraskeva, C. Direct transcriptional up-regulation of cyclooxygenase-2 by hypoxia-inducible factor (HIF)-1 promotes colorectal tumor cell survival and enhances HIF-1 transcriptional activity during hypoxia. Cancer Res. 2006, 66, 6683–6691. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Willard, M.; Simons, J. NS398 reduces hypoxia-inducible factor (HIF)-1α and HIF-1 activity: Multiple-level effects involving cyclooxygenase-2 dependent and independent mechanisms. Int. J. Cancer 2004, 112, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Chekhonin, V.P.; Shein, S.A.; Korchagina, A.A.; Gurina, O.I. VEGF in tumor progression and targeted therapy. Curr. Cancer Drug Targets 2013, 13, 423–443. [Google Scholar] [CrossRef] [PubMed]
- Eswarappa, S.M.; Potdar, A.A.; Koch, W.J.; Fan, Y.; Vasu, K.; Lindner, D.; Willard, B.; Graham, L.M.; DiCorleto, P.E.; Fox, P.L. Programmed Translational Readthrough Generates Anti-Angiogenic VEGF-Ax. Cell 2014, 157, 1605–1618. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, X.; Ge, J.; Hu, F.; Zhao, Y. Can Vascular Endothelial Growth Factor and Microvessel Density Be Used as Prognostic Biomarkers for Colorectal Cancer? A Systematic Review and Meta-Analysis. Sci. World J. 2014, 2014, 102736. [Google Scholar] [CrossRef] [PubMed]
- Bestas, R.; Kaplan, M.A.; Isikdogan, A. The correlation between serum VEGF levels and known prognostic risk factors in colorectal carcinoma. Hepatogastroenterology 2014, 61, 267–271. [Google Scholar] [PubMed]
- Uthoff, S.M.; Duchrow, M.; Schmidt, M.H.; Broll, R.; Bruch, H.P.; Strik, M.W.; Galandiuk, S. VEGF isoforms and mutations in human colorectal cancer. Int. J. Cancer 2002, 101, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.T.; Luque, A.; Lee, S.; Anderson, S.M.; Segura, T.; Iruela-Arispe, M.L. Anchorage of VEGF to the extracellular matrix conveys differential signaling responses to endothelial cells. J. Cell Biol. 2010, 188, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Brogan, I.J.; Khan, N.; Isaac, K.; Hutchinson, J.A.; Pravica, V.; Hutchinson, I.V. Novel polymorphisms in the promoter and 5’ UTR regions of the human vascular endothelial growth factor gene. Hum. Immunol. 1999, 60, 1245–1249. [Google Scholar] [CrossRef]
- Maltese, P.; Canestrari, E.; Ruzzo, A.; Graziano, F.; Falcone, A.; Loupakis, F.; Tonini, G.; Santini, D.; Magnani, M. VEGF gene polymorphisms and susceptibility to colorectal cancer disease in Italian population. Int. J. Colorectal Dis. 2009, 24, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Carmeliet, P.; Moons, L.; Luttun, A.; Vincenti, V.; Compernolle, V.; De Mol, M.; Wu, Y.; Bono, F.; Devy, L.; Beck, H.; et al. Synergism between vascular endothelial growth factor and placental growth factor contributes to angiogenesis and plasma extravasation in pathological conditions. Nat. Med. 2001, 7, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Autiero, M.; Waltenberger, J.; Communi, D.; Kranz, A.; Moons, L.; Lambrechts, D.; Kroll, J.; Plaisance, S.; de Mol, M.; Bono, F.; et al. Role of PlGF in the intra- and intermolecular cross talk between the VEGF receptors Flt1 and Flk1. Nat. Med. 2003, 9, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Sung, C.Y.; Son, M.W.; Ahn, T.S.; Jung, D.J.; Lee, M.S.; Baek, M.J. Expression of placenta growth factor in colorectal carcinomas. J. Korean Soc. Coloproctol. 2012, 28, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Tudisco, L.; Orlandi, A.; Tarallo, V.; De Falco, S. Hypoxia activates placental growth factor expression in lymphatic endothelial cells. Oncotarget 2017, 8, 32873–32883. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Mazzone, M.; Jonckx, B.; Carmeliet, P. FLT1 and its ligands VEGFB and PlGF: Drug targets for anti-angiogenic therapy? Nat. Rev. Cancer 2008, 8, 942–956. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Jonckx, B.; Mazzone, M.; Zacchigna, S.; Loges, S.; Pattarini, L.; Chorianopoulos, E.; Liesenborghs, L.; Koch, M.; De Mol, M.; et al. Anti-PlGF inhibits growth of VEGF(R)-inhibitor-resistant tumors without affecting healthy vessels. Cell 2007, 131, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Gale, N.W.; Thurston, G.; Hackett, S.F.; Renard, R.; Wang, Q.; McClain, J.; Martin, C.; Witte, C.; Witte, M.H.; Jackson, D.; et al. Angiopoietin-2 is required for postnatal angiogenesis and lymphatic patterning, and only the latter role is rescued by Angiopoietin-1. Dev. Cell 2002, 3, 411–423. [Google Scholar] [CrossRef]
- Maisonpierre, P.C.; Suri, C.; Jones, P.F.; Bartunkova, S.; Wiegand, S.J.; Radziejewski, C.; Compton, D.; McClain, J.; Aldrich, T.H.; Papadopoulos, N.; et al. Angiopoietin-2, a natural antagonist for Tie2 that disrupts in vivo angiogenesis. Science 1997, 277, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.A.; Liu, W.; Jung, Y.D.; Fan, F.; Reinmuth, N.; Bucana, C.D.; Ellis, L.M. Differential expression of angiopoietin-1 and angiopoietin-2 in colon carcinoma. A possible mechanism for the initiation of angiogenesis. Cancer 2001, 92, 1138–1143. [Google Scholar] [CrossRef]
- Seghezzi, G.; Patel, S.; Ren, C.J.; Gualandris, A.; Pintucci, G.; Robbins, E.S.; Shapiro, R.L.; Galloway, A.C.; Rifkin, D.B.; Mignatti, P. Fibroblast growth factor-2 (FGF-2) induces vascular endothelial growth factor (VEGF) expression in the endothelial cells of forming capillaries: An autocrine mechanism contributing to angiogenesis. J. Cell Biol. 1998, 141, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Simons, M. Fibroblast growth factor regulation of neovascularization. Curr. Opin. Hematol. 2008, 15, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Ueda, J.; Ishiwata, T. Fibroblast growth factor receptor 2: Expression, roles, and potential as a novel molecular target for colorectal cancer. Pathol. Res. Int. 2012, 2012, 574768. [Google Scholar] [CrossRef] [PubMed]
- Sonvilla, G.; Allerstorfer, S.; Heinzle, C.; Stattner, S.; Karner, J.; Klimpfinger, M.; Wrba, F.; Fischer, H.; Gauglhofer, C.; Spiegl-Kreinecker, S.; et al. Fibroblast growth factor receptor 3-IIIc mediates colorectal cancer growth and migration. Br. J. Cancer 2010, 102, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Narita, K.; Fujii, T.; Ishiwata, T.; Yamamoto, T.; Kawamoto, Y.; Kawahara, K.; Nakazawa, N.; Naito, Z. Keratinocyte growth factor induces vascular endothelial growth factor-A expression in colorectal cancer cells. Int. J. Oncol. 2009, 34, 355–360. [Google Scholar] [PubMed]
- Manzat Saplacan, R.M.; Balacescu, L.; Gherman, C.; Chira, R.I.; Craiu, A.; Mircea, P.A.; Lisencu, C.; Balacescu, O. The Role of PDGFs and PDGFRs in Colorectal Cancer. Mediat. Inflamm. 2017, 2017, 4708076. [Google Scholar] [CrossRef] [PubMed]
- Belizon, A.; Balik, E.; Horst, P.K.; Shantha Kumara, H.M.; Nasar, A.; Whelan, R.L. Platelet-derived growth factor (subtype BB) is elevated in patients with colorectal carcinoma. Dis. Colon Rectum 2009, 52, 1166–1171. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Multifarious functions of PDGFs and PDGFRs in tumor growth and metastasis. Trends Mol. Med. 2013, 19, 460–473. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; Somcio, R.J.; Stoeltzing, O.; Wey, J.; Fan, F.; Liu, W.; Bucana, C.; Ellis, L.M. Overexpression of PDGF-BB decreases colorectal and pancreatic cancer growth by increasing tumor pericyte content. J. Clin. Investig. 2007, 117, 2114–2122. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, C.; Berindan-Neagoe, I.; Burz, C.; Balacescu, O.; Balacescu, L.; Tudoran, O.; Asztalos, R.; Irimie, A. The clinical implications of platelet derived growth factor B, vascular endothelial growth factor and basic fibroblast growth factor in colorectal cancer. J. Buon 2011, 16, 274–276. [Google Scholar] [PubMed]
- Murdoch, C.; Muthana, M.; Coffelt, S.B.; Lewis, C.E. The role of myeloid cells in the promotion of tumour angiogenesis. Nat. Rev. Cancer 2008, 8, 618–631. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Montemurro, S.; Ruggieri, E.; Patruno, R.; Marech, I.; Cariello, M.; Vacca, A.; et al. Correlation between serum tryptase, mast cells positive to tryptase and microvascular density in colo-rectal cancer patients: Possible biological-clinical significance. PLoS ONE 2014, 9, e99512. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Patruno, R.; Sacco, R.; Marech, I.; Sammarco, G.; Zuccala, V.; Luposella, M.; Zizzo, N.; Gadaleta, C.; Porcelli, M.; et al. Mast cells positive to tryptase and tumour-associated macrophages correlate with angiogenesis in locally advanced colorectal cancer patients undergone to surgery. Expert Opin. Ther. Targets 2016, 20, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ma, T.; Shen, X.N.; Xia, X.F.; Xu, G.D.; Bai, X.L.; Liang, T.B. Macrophage-induced tumor angiogenesis is regulated by the TSC2-mTOR pathway. Cancer Res. 2012, 72, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Edin, S.; Wikberg, M.L.; Dahlin, A.M.; Rutegard, J.; Oberg, A.; Oldenborg, P.A.; Palmqvist, R. The distribution of macrophages with a M1 or M2 phenotype in relation to prognosis and the molecular characteristics of colorectal cancer. PLoS ONE 2012, 7, e47045. [Google Scholar] [CrossRef] [PubMed]
- Badawi, M.A.; Abouelfadl, D.M.; El-Sharkawy, S.L.; El-Aal, W.E.; Abbas, N.F. Tumor-Associated Macrophage (TAM) and Angiogenesis in Human Colon Carcinoma. Open Access Maced. J. Med. Sci. 2015, 3, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, J.; von Bernstorff, W.; Evert, K.; Albers, N.; Hadlich, S.; Hagemann, S.; Gunther, C.; van Rooijen, N.; Heidecke, C.D.; Partecke, L.I. Macrophages promote tumour growth and liver metastasis in an orthotopic syngeneic mouse model of colon cancer. Int. J. Colorectal Dis. 2013, 28, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Tsujino, T.; Seshimo, I.; Yamamoto, H.; Ngan, C.Y.; Ezumi, K.; Takemasa, I.; Ikeda, M.; Sekimoto, M.; Matsuura, N.; Monden, M. Stromal myofibroblasts predict disease recurrence for colorectal cancer. Clin. Cancer Res. 2007, 13, 2082–2090. [Google Scholar] [CrossRef] [PubMed]
- Torres, S.; Bartolome, R.A.; Mendes, M.; Barderas, R.; Fernandez-Acenero, M.J.; Pelaez-Garcia, A.; Pena, C.; Lopez-Lucendo, M.; Villar-Vazquez, R.; de Herreros, A.G.; et al. Proteome profiling of cancer-associated fibroblasts identifies novel proinflammatory signatures and prognostic markers for colorectal cancer. Clin. Cancer Res. 2013, 19, 6006–6019. [Google Scholar] [CrossRef] [PubMed]
- Chun, E.; Lavoie, S.; Michaud, M.; Gallini, C.A.; Kim, J.; Soucy, G.; Odze, R.; Glickman, J.N.; Garrett, W.S. CCL2 Promotes Colorectal Carcinogenesis by Enhancing Polymorphonuclear Myeloid-Derived Suppressor Cell Population and Function. Cell Rep. 2015, 12, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Montero, C.M.; Salem, M.L.; Nishimura, M.I.; Garrett-Mayer, E.; Cole, D.J.; Montero, A.J. Increased circulating myeloid-derived suppressor cells correlate with clinical cancer stage, metastatic tumor burden, and doxorubicin-cyclophosphamide chemotherapy. Cancer Immunol. Immunother. 2009, 58, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, A.G.; O’Byrne, K.J. Chronic immune activation and inflammation in the pathogenesis of AIDS and cancer. Adv. Cancer Res. 2002, 84, 231–276. [Google Scholar] [PubMed]
- Takahashi, H.; Numasaki, M.; Lotze, M.T.; Sasaki, H. Interleukin-17 enhances bFGF-, HGF- and VEGF-induced growth of vascular endothelial cells. Immunol. Lett. 2005, 98, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Vlajnic, T.; Andreozzi, M.C.; Schneider, S.; Tornillo, L.; Karamitopoulou, E.; Lugli, A.; Ruiz, C.; Zlobec, I.; Terracciano, L. VEGFA gene locus (6p12) amplification identifies a small but highly aggressive subgroup of colorectal cancer [corrected] patients. Mod. Pathol. 2011, 24, 1404–1412. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y.; Fujiki, K.; Duerr, E.-M.; Gala, M.; Jo, W.-S.; Zhang, X.; Chung, D.C. Hypoxic Regulation of Vascular Endothelial Growth Factor through the Induction of Phosphatidylinositol 3-Kinase/Rho/ROCK and c-Myc. J. Biol. Chem. 2006, 281, 13957–13963. [Google Scholar] [CrossRef] [PubMed]
- Pore, N.; Liu, S.; Shu, H.K.; Li, B.; Haas-Kogan, D.; Stokoe, D.; Milanini-Mongiat, J.; Pages, G.; O’Rourke, D.M.; Bernhard, E.; et al. Sp1 is involved in Akt-mediated induction of VEGF expression through an HIF-1-independent mechanism. Mol. Biol. Cell 2004, 15, 4841–4853. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.M.; Williams, A.C.; Game, S.M.; Paraskeva, C. Differential sensitivity of human colonic adenoma and carcinoma cells to transforming growth factor β (TGF-β): Conversion of an adenoma cell line to a tumorigenic phenotype is accompanied by a reduced response to the inhibitory effects of TGF-β. Oncogene 1991, 6, 1471–1476. [Google Scholar] [PubMed]
- Van Meeteren, L.A.; ten Dijke, P. Regulation of endothelial cell plasticity by TGF-β. Cell Tissue Res. 2012, 347, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Gille, H.; Kowalski, J.; Li, B.; LeCouter, J.; Moffat, B.; Zioncheck, T.F.; Pelletier, N.; Ferrara, N. Analysis of biological effects and signaling properties of FLT-1 (VEGFR-1) and KDR (VEGFR-2). A reassessment using novel receptor-specific vascular endothelial growth factor mutants. J. Biol. Chem. 2001, 276, 3222–3230. [Google Scholar] [CrossRef] [PubMed]
- Grady, W.M.; Myeroff, L.L.; Swinler, S.E.; Rajput, A.; Thiagalingam, S.; Lutterbaugh, J.D.; Neumann, A.; Brattain, M.G.; Chang, J.; Kim, S.J.; et al. Mutational inactivation of transforming growth factor β receptor type II in microsatellite stable colon cancers. Cancer Res. 1999, 59, 320–324. [Google Scholar] [PubMed]
- Levy, L.; Hill, C.S. Alterations in components of the TGF-β superfamily signaling pathways in human cancer. Cytokine Growth Factor Rev. 2006, 17, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Tsushima, H.; Kawata, S.; Tamura, S.; Ito, N.; Shirai, Y.; Kiso, S.; Imai, Y.; Shimomukai, H.; Nomura, Y.; Matsuda, Y.; et al. High levels of transforming growth factor β 1 in patients with colorectal cancer: Association with disease progression. Gastroenterology 1996, 110, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Matsuzaki, K.; Date, M.; Watanabe, T.; Shibano, K.; Nakagawa, T.; Yanagitani, S.; Amoh, Y.; Takemoto, H.; Ogata, N.; et al. Down-regulation of TGF-β receptors in human colorectal cancer: Implications for cancer development. Br. J. Cancer 1999, 80, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Bertolino, P.; Deckers, M.; Lebrin, F.; ten Dijke, P. Transforming Growth Factor-β Signal Transduction in Angiogenesis and Vascular Disorders. CHEST J. 2005, 128, 585S–590S. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, E.A.; Viloria-Petit, A.M.; Coomber, B.L. Colorectal carcinoma cell production of transforming growth factor β decreases expression of endothelial cell vascular endothelial growth factor receptor 2. Cancer 2011, 117, 5601–5611. [Google Scholar] [CrossRef] [PubMed]
- Grade, M.; Ghadimi, B.M.; Varma, S.; Simon, R.; Wangsa, D.; Barenboim-Stapleton, L.; Liersch, T.; Becker, H.; Ried, T.; Difilippantonio, M.J. Aneuploidy-Dependent Massive Deregulation of the Cellular Transcriptome and Apparent Divergence of the Wnt/β-catenin Signaling Pathway in Human Rectal Carcinomas. Cancer Res. 2006, 66, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Brabletz, T.; Hlubek, F.; Spaderna, S.; Schmalhofer, O.; Hiendlmeyer, E.; Jung, A.; Kirchner, T. Invasion and metastasis in colorectal cancer: Epithelial-mesenchymal transition, mesenchymal-epithelial transition, stem cells and β-catenin. Cells Tissues Organs 2005, 179, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Easwaran, V.; Lee, S.H.; Inge, L.; Guo, L.; Goldbeck, C.; Garrett, E.; Wiesmann, M.; Garcia, P.D.; Fuller, J.H.; Chan, V.; et al. β-Catenin regulates vascular endothelial growth factor expression in colon cancer. Cancer Res. 2003, 63, 3145–3153. [Google Scholar] [PubMed]
- Pate, K.T.; Stringari, C.; Sprowl-Tanio, S.; Wang, K.; TeSlaa, T.; Hoverter, N.P.; McQuade, M.M.; Garner, C.; Digman, M.A.; Teitell, M.A.; et al. Wnt signaling directs a metabolic program of glycolysis and angiogenesis in colon cancer. EMBO J. 2014, 33, 1454–1473. [Google Scholar] [CrossRef] [PubMed]
- Giles, R.H.; Lolkema, M.P.; Snijckers, C.M.; Belderbos, M.; van der Groep, P.; Mans, D.A.; van Beest, M.; van Noort, M.; Goldschmeding, R.; van Diest, P.J.; et al. Interplay between VHL/HIF1α and Wnt/β-catenin pathways during colorectal tumorigenesis. Oncogene 2006, 25, 3065–3070. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.; Zeindl-Eberhart, E.; Kirchner, T.; Jung, A. Frequency and type of KRAS mutations in routine diagnostic analysis of metastatic colorectal cancer. Pathol. Res. Pract. 2009, 205, 858–862. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.; Clements, J.; Dawson, E.; Bamford, S.; Webb, T.; Dogan, A.; Flanagan, A.; Teague, J.; Wooster, R.; Futreal, P.A.; et al. Cosmic 2005. Br. J. Cancer 2006, 94, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Ogino, S.; Meyerhardt, J.A.; Irahara, N.; Niedzwiecki, D.; Hollis, D.; Saltz, L.B.; Mayer, R.J.; Schaefer, P.; Whittom, R.; Hantel, A.; et al. KRAS mutation in stage III colon cancer and clinical outcome following intergroup trial CALGB 89803. Clin. Cancer Res. 2009, 15, 7322–7329. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Campbell, P.M.; Brekken, R.A.; Sung, B.; Ouellette, M.M.; Fleming, J.B.; Aggarwal, B.B.; Der, C.J.; Guha, S. K-Ras promotes angiogenesis mediated by immortalized human pancreatic epithelial cells through mitogen-activated protein kinase signaling pathways. Mol. Cancer Res. 2009, 7, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gaspard, J.P.; Chung, D.C. Regulation of vascular endothelial growth factor by the Wnt and K-Ras pathways in colonic neoplasia. Cancer Res. 2001, 61, 6050–6054. [Google Scholar] [PubMed]
- Pietrantonio, F.; Petrelli, F.; Coinu, A.; Di Bartolomeo, M.; Borgonovo, K.; Maggi, C.; Cabiddu, M.; Iacovelli, R.; Bossi, I.; Lonati, V.; et al. Predictive role of BRAF mutations in patients with advanced colorectal cancer receiving cetuximab and panitumumab: A meta-analysis. Eur. J. Cancer 2015, 51, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B., 3rd.; Venook, A.P.; Cederquist, L.; Chan, E.; Chen, Y.J.; Cooper, H.S.; Deming, D.; Engstrom, P.F.; Enzinger, P.C.; Fichera, A.; et al. Colon Cancer, Version 1.2017, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2017, 15, 370–398. [Google Scholar] [CrossRef]
- Rui-hua Xu, R.H.; Shen, L.; Wang, K.; Wu, G.; Shi, C.; Ding, K.; Lin, L.; Wang, J.Y.; Xiong, J.; Wu, G.; et al. A randomized, double-blind, parallel-group, placebo-controlled, multicenter, phase II clinical study of famitinib in the treatment of advanced metastatic colorectal cancer. J. Clin. Oncol. 2015, 33, 513. [Google Scholar]
- Li, J.; Qin, S.; Bai, Y.; Deng, Y.; Yang, L.; Xu, R.H.; Chen, Z.; Zhong, H.; Pan, H.; Shu, Y.; et al. A randomized, double-blind, placebo-controlled, multi-centered phase 3 trial comparing fruquintinib versus placebo plus best supportive care in Chinese patients with metastatic colorectal cancer (FRESCO). J. Clin. Oncol. 2017, 35, 3508. [Google Scholar]
- Van Cutsem, E.; Prenen, H.; D’Haens, G.; Bennouna, J.; Carrato, A.; Ducreux, M.; Bouche, O.; Sobrero, A.; Latini, L.; Staines, H.; et al. A phase I/II, open-label, randomised study of nintedanib plus mFOLFOX6 versus bevacizumab plus mFOLFOX6 in first-line metastatic colorectal cancer patients. Ann. Oncol. 2015, 26, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Yoshino, T.; Hocke, J.; Oum’Hamed, Z.; Studeny, M.; Tabernero, J. Rationale and Design for the LUME-Colon 1 Study: A Randomized, Double-Blind, Placebo-Controlled Phase III Trial of Nintedanib Plus Best Supportive Care Versus Placebo Plus Best Supportive Care in Patients With Advanced Colorectal Cancer Refractory to Standard Treatment. Clin. Colorectal Cancer 2016, 15, 91.e91–94.e91. [Google Scholar] [PubMed]
- Peeters, M.; Strickland, A.H.; Lichinitser, M.; Suresh, A.V.; Manikhas, G.; Shapiro, J.; Rogowski, W.; Huang, X.; Wu, B.; Warner, D.; et al. A randomised, double-blind, placebo-controlled phase 2 study of trebananib (AMG 386) in combination with FOLFIRI in patients with previously treated metastatic colorectal carcinoma. Br. J. Cancer 2013, 108, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Meyerhardt, J.A.; Ancukiewicz, M.; Abrams, T.A.; Schrag, D.; Enzinger, P.C.; Chan, J.A.; Kulke, M.H.; Wolpin, B.M.; Goldstein, M.; Blaszkowsky, L.; et al. Phase I study of cetuximab, irinotecan, and vandetanib (ZD6474) as therapy for patients with previously treated metastastic colorectal cancer. PLoS ONE 2012, 7, e38231. [Google Scholar] [CrossRef] [PubMed]
- Cabebe, E.C.; Fisher, G.A.; Sikic, B.I. A phase I trial of vandetanib combined with capecitabine, oxaliplatin and bevacizumab for the first-line treatment of metastatic colorectal cancer. Investig. New Drugs 2012, 30, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Bendell, J.C.; Sauri, T.; Cubillo, A.; Lopez-Lopez, C.; Alfonso, P.G.; Hussein, M.A.; Limon, M.L.; Cervantes, A.; Montagut, C.; Santos, C.; et al. Final results of the McCAVE trial: A double-blind, randomized phase 2 study of vanucizumab (VAN) plus FOLFOX vs. bevacizumab (BEV) plus FOLFOX in patients (pts) with previously untreated metastatic colorectal carcinoma (mCRC). J. Clin. Oncol. 2017, 25, 3539. [Google Scholar]
- Hutt, D.M.; Roth, D.M.; Vignaud, H.; Cullin, C.; Bouchecareilh, M. The histone deacetylase inhibitor, Vorinostat, represses hypoxia inducible factor 1 α expression through translational inhibition. PLoS ONE 2014, 9, e106224. [Google Scholar] [CrossRef] [PubMed]
- Wilson, P.M.; El-Khoueiry, A.; Iqbal, S.; Fazzone, W.; LaBonte, M.J.; Groshen, S.; Yang, D.; Danenberg, K.D.; Cole, S.; Kornacki, M.; et al. A phase I/II trial of vorinostat in combination with 5-fluorouracil in patients with metastatic colorectal cancer who previously failed 5-FU-based chemotherapy. Cancer Chemother. Pharmacol. 2010, 65, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, H.; Fehrenbacher, L.; Novotny, W.; Cartwright, T.; Hainsworth, J.; Heim, W.; Berlin, J.; Baron, A.; Griffing, S.; Holmgren, E.; et al. Bevacizumab plus Irinotecan, Fluorouracil, and Leucovorin for Metastatic Colorectal Cancer. N. Engl. J. Med. 2004, 350, 2335–2342. [Google Scholar] [CrossRef] [PubMed]
- Kabbinavar, F.F.; Hambleton, J.; Mass, R.D.; Hurwitz, H.I.; Bergsland, E.; Sarkar, S. Combined analysis of efficacy: The addition of bevacizumab to fluorouracil/leucovorin improves survival for patients with metastatic colorectal cancer. J. Clin. Oncol. 2005, 23, 3706–3712. [Google Scholar] [CrossRef] [PubMed]
- Sobrero, A.; Ackland, S.; Clarke, S.; Perez-Carrión, R.; Chiara, S.; Gapski, J.; Mainwaring, P.; Langer, B.; Young, S. Phase IV Study of Bevacizumab in Combination with Infusional Fluorouracil, Leucovorin and Irinotecan (FOLFIRI) in First-Line Metastatic Colorectal Cancer. Oncology 2009, 77, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Saltz, L.B.; Clarke, S.; Diaz-Rubio, E.; Scheithauer, W.; Figer, A.; Wong, R.; Koski, S.; Lichinitser, M.; Yang, T.S.; Rivera, F.; et al. Bevacizumab in combination with oxaliplatin-based chemotherapy as first-line therapy in metastatic colorectal cancer: A randomized phase III study. J. Clin. Oncol. 2008, 26, 2013–2019. [Google Scholar] [CrossRef] [PubMed]
- Hochster, H.S.; Hart, L.L.; Ramanathan, R.K.; Childs, B.H.; Hainsworth, J.D.; Cohn, A.L.; Wong, L.; Fehrenbacher, L.; Abubakr, Y.; Saif, M.W.; et al. Safety and efficacy of oxaliplatin and fluoropyrimidine regimens with or without bevacizumab as first-line treatment of metastatic colorectal cancer: Results of the TREE Study. J. Clin. Oncol. 2008, 26, 3523–3529. [Google Scholar] [CrossRef] [PubMed]
- Van Cutsem, E.; Rivera, F.; Berry, S.; Kretzschmar, A.; Michael, M.; DiBartolomeo, M.; Mazier, M.A.; Canon, J.L.; Georgoulias, V.; Peeters, M.; et al. Safety and efficacy of first-line bevacizumab with FOLFOX, XELOX, FOLFIRI and fluoropyrimidines in metastatic colorectal cancer: The BEAT study. Ann. Oncol. 2009, 20, 1842–1847. [Google Scholar] [CrossRef] [PubMed]
- Loupakis, F.; Cremolini, C.; Masi, G.; Lonardi, S.; Zagonel, V.; Salvatore, L.; Cortesi, E.; Tomasello, G.; Ronzoni, M.; Spadi, R.; et al. Initial therapy with FOLFOXIRI and bevacizumab for metastatic colorectal cancer. N. Engl. J. Med. 2014, 371, 1609–1618. [Google Scholar] [CrossRef] [PubMed]
- Bennouna, J.; Sastre, J.; Arnold, D.; Osterlund, P.; Greil, R.; Van Cutsem, E.; von Moos, R.; Vieitez, J.M.; Bouche, O.; Borg, C.; et al. Continuation of bevacizumab after first progression in metastatic colorectal cancer (ML18147): A randomised phase 3 trial. Lancet Oncol. 2013, 14, 29–37. [Google Scholar] [CrossRef]
- Tsai, H.L.; Lin, C.H.; Huang, C.W.; Yang, I.P.; Yeh, Y.S.; Hsu, W.H.; Wu, J.Y.; Kuo, C.H.; Tseng, F.Y.; Wang, J.Y. Decreased peritherapeutic VEGF expression could be a predictor of responsiveness to first-line FOLFIRI plus bevacizumab in mCRC patients. Int. J. Clin. Exp. Pathol. 2015, 8, 1900–1910. [Google Scholar] [PubMed]
- Van Cutsem, E.; Tabernero, J.; Lakomy, R.; Prenen, H.; Prausova, J.; Macarulla, T.; Ruff, P.; van Hazel, G.A.; Moiseyenko, V.; Ferry, D.; et al. Addition of aflibercept to fluorouracil, leucovorin, and irinotecan improves survival in a phase III randomized trial in patients with metastatic colorectal cancer previously treated with an oxaliplatin-based regimen. J. Clin. Oncol. 2012, 30, 3499–3506. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A.; van Cutsem, E.; Sobrero, A.; Siena, S.; Falcone, A.; Ychou, M.; Humblet, Y.; Bouche, O.; Mineur, L.; Barone, C.; et al. Regorafenib monotherapy for previously treated metastatic colorectal cancer (CORRECT): An international, multicentre, randomised, placebo-controlled, phase 3 trial. Lancet 2013, 381, 303–312. [Google Scholar] [CrossRef]
- Mayer, R.J.; van Cutsem, E.; Falcone, A.; Yoshino, T.; Garcia-Carbonero, R.; Mizunuma, N.; Yamazaki, K.; Shimada, Y.; Tabernero, J.; Komatsu, Y.; et al. Randomized trial of TAS-102 for refractory metastatic colorectal cancer. N. Engl. J. Med. 2015, 372, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Emura, T.; Suzuki, N.; Fujioka, A.; Ohshimo, H.; Fukushima, M. Potentiation of the antitumor activity of α, α, α-trifluorothymidine by the co-administration of an inhibitor of thymidine phosphorylase at a suitable molar ratio in vivo. Int. J. Oncol. 2005, 27, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.H.; Li, J.; Bai, Y.; Xu, J.; Liu, T.; Shen, L.; Wang, L.; Pan, H.; Cao, J.; Zhang, D.; et al. Safety and efficacy of fruquintinib in patients with previously treated metastatic colorectal cancer: A phase Ib study and a randomized double-blind phase II study. J. Hematol. Oncol. 2017, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, G.; Coviello, M.; Patruno, R.; Valerio, P.; Martino, D.; Milella, P.; Catalano, V.; Scotto, F.; de Ceglie, A.; Quaranta, M.; et al. Vascular endothelial growth factor concentrations in the plasma-activated platelets rich (P-APR) of healthy controls and colorectal cancer patients. Oncol. Rep. 2004, 12, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Coskun, O.; Oztopuz, O.; Ozkan, O.F. Determination of IL-6, TNF-α and VEGF levels in the serums of patients with colorectal cancer. Cell. Mol. Biol. 2017, 63, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Martins, S.F.; Garcia, E.A.; Luz, M.A.; Pardal, F.; Rodrigues, M.; Filho, A.L. Clinicopathological correlation and prognostic significance of VEGF-A, VEGF-C, VEGFR-2 and VEGFR-3 expression in colorectal cancer. Cancer Genom. Proteom. 2013, 10, 55–67. [Google Scholar]
- De Vita, F.; Orditura, M.; Lieto, E.; Infusino, S.; Morgillo, F.; Martinelli, E.; Castellano, P.; Romano, C.; Ciardiello, F.; Catalano, G.; et al. Elevated perioperative serum vascular endothelial growth factor levels in patients with colon carcinoma. Cancer 2004, 100, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Divella, R.; Daniele, A.; DE Luca, R.; Simone, M.; Naglieri, E.; Savino, E.; Abbate, I.; Gadaleta, C.D.; Ranieri, G. Circulating Levels of VEGF and CXCL1 Are Predictive of Metastatic Organotropismin in Patients with Colorectal Cancer. Anticancer Res. 2017, 37, 4867–4871. [Google Scholar] [PubMed]
- Kwon, K.A.; Kim, S.H.; Oh, S.Y.; Lee, S.; Han, J.Y.; Kim, K.H.; Goh, R.Y.; Choi, H.J.; Park, K.J.; Roh, M.S.; et al. Clinical significance of preoperative serum vascular endothelial growth factor, interleukin-6, and C-reactive protein level in colorectal cancer. BMC Cancer 2010, 10, 203. [Google Scholar] [CrossRef] [PubMed]
- Kjaer-Frifeldt, S.; Fredslund, R.; Lindebjerg, J.; Hansen, T.F.; Spindler, K.L.; Jakobsen, A. Danish Colorectal Cancer Group. Prognostic importance of VEGF-A haplotype combinations in a stage II colon cancer population. Pharmacogenomics 2012, 13, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Zygon, J.; Szajewski, M.; Kruszewski, W.J.; Rzepko, R. VEGF, Flt-1, and microvessel density in primary tumors as predictive factors of colorectal cancer prognosis. Mol. Clin. Oncol. 2017, 6, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Giatromanolaki, A.; Koukourakis, M.I.; Sivridis, E.; Chlouverakis, G.; Vourvouhaki, E.; Turley, H.; Harris, A.L.; Gatter, K.C. Activated VEGFR2/KDR pathway in tumour cells and tumour associated vessels of colorectal cancer. Eur. J. Clin. Investig. 2007, 37, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Casanovas, O.; Hicklin, D.J.; Bergers, G.; Hanahan, D. Drug resistance by evasion of antiangiogenic targeting of VEGF signaling in late-stage pancreatic islet tumors. Cancer Cell 2005, 8, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Arao, T.; Matsumoto, K.; Kimura, H.; Togashi, Y.; Hirashima, Y.; Horita, Y.; Iwasa, S.; Okita, N.T.; Honma, Y.; et al. Biomarkers of reactive resistance and early disease progression during chemotherapy plus bevacizumab treatment for colorectal carcinoma. Oncotarget 2014, 5, 2588–2595. [Google Scholar] [CrossRef] [PubMed]
- Alidzanovic, L.; Starlinger, P.; Schauer, D.; Maier, T.; Feldman, A.; Buchberger, E.; Stift, J.; Koeck, U.; Pop, L.; Gruenberger, B.; et al. The VEGF rise in blood of bevacizumab patients is not based on tumor escape but a host-blockade of VEGF clearance. Oncotarget 2016, 7, 57197–57212. [Google Scholar] [CrossRef] [PubMed]
- Martinetti, A.; Miceli, R.; Sottotetti, E.; di Bartolomeo, M.; de Braud, F.; Gevorgyan, A.; Dotti, K.F.; Bajetta, E.; Campiglio, M.; Bianchi, F.; et al. Circulating biomarkers in advanced colorectal cancer patients randomly assigned to three bevacizumab-based regimens. Cancers 2014, 6, 1753–1768. [Google Scholar] [CrossRef] [PubMed]
- Hegde, P.S.; Jubb, A.M.; Chen, D.; Li, N.F.; Meng, Y.G.; Bernaards, C.; Elliott, R.; Scherer, S.J.; Chen, D.S. Predictive impact of circulating vascular endothelial growth factor in four phase III trials evaluating bevacizumab. Clin. Cancer Res. 2013, 19, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Jubb, A.M.; Hurwitz, H.I.; Bai, W.; Holmgren, E.B.; Tobin, P.; Guerrero, A.S.; Kabbinavar, F.; Holden, S.N.; Novotny, W.F.; Frantz, G.D.; et al. Impact of vascular endothelial growth factor-A expression, thrombospondin-2 expression, and microvessel density on the treatment effect of bevacizumab in metastatic colorectal cancer. J. Clin. Oncol. 2006, 24, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Loupakis, F.; Ruzzo, A.; Salvatore, L.; Cremolini, C.; Masi, G.; Frumento, P.; Schirripa, M.; Catalano, V.; Galluccio, N.; Canestrari, E.; et al. Retrospective exploratory analysis of VEGF polymorphisms in the prediction of benefit from first-line FOLFIRI plus bevacizumab in metastatic colorectal cancer. BMC Cancer 2011, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Formica, V.; Palmirotta, R.; del Monte, G.; Savonarola, A.; Ludovici, G.; de Marchis, M.L.; Grenga, I.; Schirru, M.; Guadagni, F.; Roselli, M. Predictive value of VEGF gene polymorphisms for metastatic colorectal cancer patients receiving first-line treatment including fluorouracil, irinotecan, and bevacizumab. Int. J. Colorectal Dis. 2011, 26, 143–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loupakis, F.; Cremolini, C.; Yang, D.; Salvatore, L.; Zhang, W.; Wakatsuki, T.; Bohanes, P.; Schirripa, M.; Benhaim, L.; Lonardi, S.; et al. Prospective validation of candidate SNPs of VEGF/VEGFR pathway in metastatic colorectal cancer patients treated with first-line FOLFIRI plus bevacizumab. PLoS ONE 2013, 8, e66774. [Google Scholar] [CrossRef] [PubMed]
- Koutras, A.K.; Antonacopoulou, A.G.; Eleftheraki, A.G.; Dimitrakopoulos, F.I.; Koumarianou, A.; Varthalitis, I.; Fostira, F.; Sgouros, J.; Briasoulis, E.; Bournakis, E.; et al. Vascular endothelial growth factor polymorphisms and clinical outcome in colorectal cancer patients treated with irinotecan-based chemotherapy and bevacizumab. Pharmacogenom. J. 2012, 12, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Azuma, M.; Lurje, G.; Gordon, M.A.; Yang, D.; Pohl, A.; Ning, Y.; Bohanes, P.; Gerger, A.; Winder, T.; et al. Molecular predictors of combination targeted therapies (cetuximab, bevacizumab) in irinotecan-refractory colorectal cancer (BOND-2 study). Anticancer Res. 2010, 30, 4209–4217. [Google Scholar] [PubMed]
- Duda, D.G.; Willett, C.G.; Ancukiewicz, M.; di Tomaso, E.; Shah, M.; Czito, B.G.; Bentley, R.; Poleski, M.; Lauwers, G.Y.; Carroll, M.; et al. Plasma soluble VEGFR-1 is a potential dual biomarker of response and toxicity for bevacizumab with chemoradiation in locally advanced rectal cancer. Oncologist 2010, 15, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Cacev, T.; Radosevic, S.; Krizanac, S.; Kapitanovic, S. Influence of interleukin-8 and interleukin-10 on sporadic colon cancer development and progression. Carcinogenesis 2008, 29, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Di Salvatore, M.; Pietrantonio, F.; Orlandi, A.; del Re, M.; Berenato, R.; Rossi, E.; Caporale, M.; Guarino, D.; Martinetti, A.; Basso, M.; et al. IL-8 and eNOS polymorphisms predict bevacizumab-based first line treatment outcomes in RAS mutant metastatic colorectal cancer patients. Oncotarget 2017, 8, 16887–16898. [Google Scholar] [CrossRef] [PubMed]
- Tabernero, J.; Paccard, C.; Chiron, M.; Dochy, E.; van Cutsem, E. Placental growth factor and the angiogenic environment based on analysis of baseline plasma biomarkers from the VELOUR trial. J. Clin. Oncol. 2017, 35, 592. [Google Scholar] [CrossRef]
- Chung, Y.C.; Hou, Y.C.; Chang, C.N.; Hseu, T.H. Expression and prognostic significance of angiopoietin in colorectal carcinoma. J. Surg. Oncol. 2006, 94, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Jung, H.I.; Ahn, T.S.; Kim, H.J.; Lee, K.T.; Baek, M.J.; Bae, S.B. Expressions and Clinical Significances of Angiopoietin-1, Angiopoietin-2, and Tie-2 Receptor in Patients With Colorectal Cancer. Ann. Coloproctol. 2017, 33, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Toiyama, Y.; Tanaka, K.; Kitajima, T.; Shimura, T.; Kawamura, M.; Kawamoto, A.; Okugawa, Y.; Saigusa, S.; Hiro, J.; Inoue, Y.; et al. Elevated serum angiopoietin-like protein 2 correlates with the metastatic properties of colorectal cancer: A serum biomarker for early diagnosis and recurrence. Clin. Cancer Res. 2014, 20, 6175–6186. [Google Scholar] [CrossRef] [PubMed]
- Engin, H.; Ustundag, Y.; Tekin, I.O.; Gokmen, A.; Ertop, S.; Ilikhan, S.U. Plasma concentrations of angiopoietin-1, angiopoietin-2 and Tie-2 in colon cancer. Eur. Cytokine Netw. 2012, 23, 68–71. [Google Scholar] [PubMed]
- Manzat-Saplacan, R.M.; Balacescu, L.; Gherman, C.; Visan, S.; Chira, R.I.; Bintintan, A.; Nagy, G.; Popovici, C.; Valean, S.D.; Anca, C.; et al. Is there a correlation between peripheral blood expression of angiogenic transcriptional factors/receptors and colorectal cancer? J. Buon 2015, 20, 1193–1200. [Google Scholar] [PubMed]
- Kurokawa, Y.; Matsuura, N.; Kawabata, R.; Nishikawa, K.; Ebisui, C.; Yokoyama, Y.; Shaker, M.N.; Hamakawa, T.; Takahashi, T.; Takiguchi, S.; et al. Prognostic impact of major receptor tyrosine kinase expression in gastric cancer. Ann. Surg. Oncol. 2014, 21, S584–S590. [Google Scholar] [CrossRef] [PubMed]
- Katano, M.; Nakamura, M.; Fujimoto, K.; Miyazaki, K.; Morisaki, T. Prognostic value of platelet-derived growth factor-A (PDGF-A) in gastric carcinoma. Ann. Surg. 1998, 227, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, N.; Inokuchi, M.; Takagi, Y.; Sugita, H.; Kato, K.; Kojima, K.; Sugihara, K. Clinical significance of platelet derived growth factor-C and -D in gastric cancer. Oncol. Lett. 2015, 10, 3495–3501. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tanaka, F.; Yoshikawa, Y.; Mimori, K.; Inoue, H.; Yanaga, K.; Mori, M. PDGF-BB is a novel prognostic factor in colorectal cancer. Ann. Surg. Oncol 2008, 15, 2129–2136. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Yamamoto, M.; Minami, S.; Komuta, K.; Yamaguchi, J.; Furui, J.; Kanematsu, T. Human colon cancer produces a factor which induces the proliferation of venous endothelial cells. Oncol. Rep. 2001, 8, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, H.; Altuntas, F.; Akbulut, K.G.; Ozturk, G.; Cindoruk, M.; Unal, E.; Icli, F. Prognostic role of serum vascular endothelial growth factor, basic fibroblast growth factor and nitric oxide in patients with colorectal carcinoma. Cytokine 2002, 20, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.K.; Jung, K.U.; Choi, Y.L.; Hong, H.K.; Kim, S.H.; Yun, S.H.; Kim, H.C.; Lee, W.Y.; Cho, Y.B. Low expression of transforming growth factor β-1 in cancer tissue predicts a poor prognosis for patients with stage III rectal cancers. Oncology 2014, 86, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Yamakuchi, M.; Yagi, S.; Ito, T.; Lowenstein, C.J. MicroRNA-22 regulates hypoxia signaling in colon cancer cells. PLoS ONE 2011, 6, e20291. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Asai, T.; Koide, H.; Okamoto, A.; Maeda, N.; Tomita, K.; Dewa, T.; Minamino, T.; Oku, N. Advanced cancer therapy by integrative antitumor actions via systemic administration of miR-499. J. Control. Release 2014, 181, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Nagel, R.; le Sage, C.; Diosdado, B.; van der Waal, M.; Oude Vrielink, J.A.; Bolijn, A.; Meijer, G.A.; Agami, R. Regulation of the adenomatous polyposis coli gene by the miR-135 family in colorectal cancer. Cancer Res. 2008, 68, 5795–5802. [Google Scholar] [CrossRef] [PubMed]
- Monzo, M.; Navarro, A.; Bandres, E.; Artells, R.; Moreno, I.; Gel, B.; Ibeas, R.; Moreno, J.; Martinez, F.; Diaz, T.; et al. Overlapping expression of microRNAs in human embryonic colon and colorectal cancer. Cell Res. 2008, 18, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Guo, X.; Zhang, H.; Xiang, Y.; Chen, J.; Yin, Y.; Cai, X.; Wang, K.; Wang, G.; Ba, Y.; et al. Role of miR-143 targeting KRAS in colorectal tumorigenesis. Oncogene 2009, 28, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Okugawa, Y.; Toiyama, Y.; Goel, A. An update on microRNAs as colorectal cancer biomarkers: Where are we and what’s next? Expert Rev. Mol. Diagn. 2014, 14, 999–1021. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Liu, L.Z.; Qian, X.; Chen, Q.; Jiang, Y.; Li, D.; Lai, L.; Jiang, B.H. MiR-145 directly targets p70S6K1 in cancer cells to inhibit tumor growth and angiogenesis. Nucleic Acids Res. 2012, 40, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Shi, K.; Kang, X.; Li, W. Prognostic value of long non-coding RNA TUG1 in various tumors. Oncotarget 2017, 8, 65659–65667. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jin, M.; Xu, H.; Shimin, Z.; He, S.; Wang, L.; Zhang, Y. Clinicopathologic significance of HIF-1α, CXCR4, and VEGF expression in colon cancer. Clin. Dev. Immunol. 2010, 2010, 537531. [Google Scholar] [CrossRef] [PubMed]
- Mansour, R.N.; Enderami, S.E.; Ardeshirylajimi, A.; Fooladsaz, K.; Fathi, M.; Ganji, S.M. Evaluation of hypoxia inducible factor-1 α gene expression in colorectal cancer stages of Iranian patients. J. Cancer Res. Ther. 2016, 12, 1313–1317. [Google Scholar] [PubMed]
- Setrerrahmane, S.; Xu, H. Tumor-related interleukins: Old validated targets for new anti-cancer drug development. Mol. Cancer 2017, 16, 153. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, G.; Pallone, F.; Stolfi, C. The dual role of inflammation in colon carcinogenesis. Int. J. Mol. Sci. 2012, 13, 11071–11084. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Choi, I.; Ning, Y.; Kim, N.Y.; Khatchadourian, V.; Yang, D.; Chung, H.K.; Choi, D.; LaBonte, M.J.; Ladner, R.D.; et al. Interleukin-8 and its receptor CXCR2 in the tumour microenvironment promote colon cancer growth, progression and metastasis. Br. J. Cancer 2012, 106, 1833–1841. [Google Scholar] [CrossRef] [PubMed]
- Nastase, A.; Paslaru, L.; Herlea, V.; Ionescu, M.; Tomescu, D.; Bacalbasa, N.; Dima, S.; Popescu, I. Expression of interleukine-8 as an independent prognostic factor for sporadic colon cancer dissemination. J. Med. Life 2014, 7, 215–219. [Google Scholar] [PubMed]
- Nagasaki, T.; Hara, M.; Nakanishi, H.; Takahashi, H.; Sato, M.; Takeyama, H. Interleukin-6 released by colon cancer-associated fibroblasts is critical for tumour angiogenesis: Anti-interleukin-6 receptor antibody suppressed angiogenesis and inhibited tumour-stroma interaction. Br. J. Cancer 2014, 110, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, Y.; Jo, W.S.; Duerr, E.M.; Gala, M.; Li, J.; Zhang, X.; Zimmer, M.A.; Iliopoulos, O.; Zukerberg, L.R.; Kohgo, Y.; et al. Induction of interleukin-8 preserves the angiogenic response in HIF-1α-deficient colon cancer cells. Nat. Med. 2005, 11, 992–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Caro, G.; Carvello, M.; Pesce, S.; Erreni, M.; Marchesi, F.; Todoric, J.; Sacchi, M.; Montorsi, M.; Allavena, P.; Spinelli, A. Circulating Inflammatory Mediators as Potential Prognostic Markers of Human Colorectal Cancer. PLoS ONE 2016, 11, e0148186. [Google Scholar]
- Hara, M.; Nagasaki, T.; Shiga, K.; Takahashi, H.; Takeyama, H. High serum levels of interleukin-6 in patients with advanced or metastatic colorectal cancer: The effect on the outcome and the response to chemotherapy plus bevacizumab. Surg. Today 2017, 47, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Marisi, G.; Scarpi, E.; Passardi, A.; Nanni, O.; Ragazzini, A.; Valgiusti, M.; Casadei Gardini, A.; Neri, L.M.; Frassineti, G.L.; Amadori, D.; et al. Circulating VEGF and eNOS variations as predictors of outcome in metastatic colorectal cancer patients receiving bevacizumab. Sci. Rep. 2017, 7, 1293. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.O.; Catalano, P.J.; Symonds, K.E.; Varey, A.H.; Ramani, P.; O’Dwyer, P.J.; Giantonio, B.J.; Meropol, N.J.; Benson, A.B.; Harper, S.J. Association between VEGF splice isoforms and progression-free survival in metastatic colorectal cancer patients treated with bevacizumab. Clin. Cancer Res. 2012, 18, 6384–6391. [Google Scholar] [CrossRef] [PubMed]
- Bunni, J.; Shelley-Fraser, G.; Stevenson, K.; Oltean, S.; Salmon, A.; Harper, S.J.; Carter, J.G.; Bates, D.O. Circulating levels of anti-angiogenic VEGF-A isoform (VEGF-Axxxb) in colorectal cancer patients predicts tumour VEGF-A ratios. Am. J. Cancer Res. 2015, 5, 2083–2089. [Google Scholar] [PubMed]
- Zhang, S.D.; McCrudden, C.M.; Meng, C.; Lin, Y.; Kwok, H.F. The significance of combining VEGFA, FLT1, and KDR expressions in colon cancer patient prognosis and predicting response to bevacizumab. Onco Targets Ther. 2015, 8, 835–843. [Google Scholar] [PubMed]
- Jurgensmeier, J.M.; Schmoll, H.J.; Robertson, J.D.; Brooks, L.; Taboada, M.; Morgan, S.R.; Wilson, D.; Hoff, P.M. Prognostic and predictive value of VEGF, sVEGFR-2 and CEA in mCRC studies comparing cediranib, bevacizumab and chemotherapy. Br. J. Cancer 2013, 108, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Bergers, G.; Hanahan, D. Modes of resistance to anti-angiogenic therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef] [PubMed]
- Lieu, C.H.; Tran, H.; Jiang, Z.Q.; Mao, M.; Overman, M.J.; Lin, E.; Eng, C.; Morris, J.; Ellis, L.; Heymach, J.V.; et al. The association of alternate VEGF ligands with resistance to anti-VEGF therapy in metastatic colorectal cancer. PLoS ONE 2013, 8, e77117. [Google Scholar] [CrossRef] [PubMed]
- Boisen, M.K.; Dehlendorff, C.; Linnemann, D.; Nielsen, B.S.; Larsen, J.S.; Osterlind, K.; Nielsen, S.E.; Tarpgaard, L.S.; Qvortrup, C.; Pfeiffer, P.; et al. Tissue microRNAs as predictors of outcome in patients with metastatic colorectal cancer treated with first line Capecitabine and Oxaliplatin with or without Bevacizumab. PLoS ONE 2014, 9, e109430. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.F.; Carlsen, A.L.; Heegaard, N.H.; Sorensen, F.B.; Jakobsen, A. Changes in circulating microRNA-126 during treatment with chemotherapy and bevacizumab predicts treatment response in patients with metastatic colorectal cancer. Br. J. Cancer 2015, 112, 624–629. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Compound | Category | Pro-Angiogenic Targets | Combination Drugs | Clinical Use | Route of Administration | Ref. |
|---|---|---|---|---|---|---|
| Aflibercept | ”Trap”-inhibitor, recombinant fusion protein. | VEGF-A/B, PlGF | Fluoropyrimidines, folinic acid and irinotecan | Second line, after oxaliplatin; regardless K/NRAS and BRAF genotype | Intravenous | [75] |
| Bevacizumab | Recombinant humanized—mAb | VEGF-A | Fluoropyrimidines, folinic acid, irinotecan and/or oxaliplatin | First and second line; regardless K/NRAS and BRAF genotype | Intravenous | [75] |
| Famitinib | Small molecule, multiple kinase inhibitor | VEGRs, PDGFRs, c-KIT, FLT3, RET | Alone | No, experimental use | Oral | [76] |
| Fruquintinib | Small molecule, multiple kinase inhibitor | VEGRs | Alone | No, experimental use | Oral | [77] |
| Nintedanib | Small molecule, multiple kinase inhibitor | VEGFR 1-3, FGFRs, PDGFRs | 5FU, folinic acid and oxaliplatin | No, experimental use | Oral | [78,79] |
| Regorafenib | Small molecule, multiple kinase inhibitor | KIT, RET, PDGFR, FGFR BRAF | Alone | After progression to other conventional treatments | Oral | [75] |
| Trebanabib | Fusion protein | ANG-1/2, TIE2 | 5FU, folinic acid and irinotecan | No, experimental use | Oral | [80] |
| Trifluridine/tipiracil | Antimetabolite | PD-ECGF | Alone | After progression to other conventional treatments | Oral | [75] |
| Vandetanib | Small molecule, multiple kinase inhibitor | VEGFR-2, EGFR | Irinotecan and cetuximab; capecitabine, oxaliplatin and bevacizumab | Not in use, due to unsafe toxicity profile | Oral | [81,82] |
| Vanucizumab | Bi-specific monoclonal antibody | VEGF-A, ANG-2 | FOLFOX | No, experimental use | Intravenous | [83] |
| Vorinostat | Histone deacetylase inhibitors | HIF1-α | 5FU | No, experimental use | Oral | [84,85] |
| Biomarker | Drug/Treatment | Prognostic Value | Predictive Value | Ref. |
|---|---|---|---|---|
| Circulating VEGF-A | FOLFIRI plus bevacizumab | Yes | Yes (VEGF-A ↑ during treatment and poor prognosis) | [110] |
| XELOX, FOLFOX6 or FOLFIRI/FOLFOXIRI plus bevacizumab | Yes | No | [111] | |
| Tissue VEGF expression | Randomized, 3-arms, phase II trial plus bevacizumab; multiple treatments | Yes | N/V | [112,113] |
| FOLFIRI plus bevacizumab | N/V | Yes (↓ peri-therapeutic VEGF-A expression predicts responsiveness to bevacizumab and PFS) | [94] | |
| IFL | NO | NO (bevacizumab improves survival regardless VEGF levels) | [114] | |
| SNPs VEGF | FOLFIRI plus bevacizumab, retrospective analysis | N/V | Yes (VEGF GP −1498 T/T genotype was associated with shorter PFS) | [115,116] |
| FOLFIRI plus bevacizumab | N/V | No | [117] | |
| FOLFIRI AND XELIRI plus Bevacizumab | N/V | Yes (the VEGF GP 1154 G/G is associated with OS) | [118] | |
| Retrospective analysis | Yes (VEGF-2578 is associated with time of recurrence) | N/V | [106] | |
| FOLFIRI plus bevacizumab | N/V | Yes (VEGF GPs −1154 G/G −152 G/G is predictive for PFS) | [116] | |
| Tissue VEGFR1,VEGFR2, VEGFR3 expression | CBI or CB * | Yes (VEGFR2 could predict clinical outcome in mCRC | N/V | [119] |
| Four cycles of therapy plus Bevacizumab | N/V | YES (Pretreatment ↑ soluble VEGFR-1 is associated with higher response to therapy) | [120] | |
| Tissue IL-8 and SNP expression | Retrospective analysis ** | Yes | N/V | [121] |
| Bevacizumab-based first line treatment | N/V | YES (IL-8 GPs-51 T/A and A/A are associated with shorter PFS and OS) | [122] | |
| Serum PLGF | FOLFIRI plus bevacizumab/aflibercept | N/V | YES (↑PLGF and VEGF correlate with response in patients previous treated with bevacizumab) | [123] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelucci, A.; Delle Monache, S.; Cortellini, A.; Di Padova, M.; Ficorella, C. “Vessels in the Storm”: Searching for Prognostic and Predictive Angiogenic Factors in Colorectal Cancer. Int. J. Mol. Sci. 2018, 19, 299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19010299
Angelucci A, Delle Monache S, Cortellini A, Di Padova M, Ficorella C. “Vessels in the Storm”: Searching for Prognostic and Predictive Angiogenic Factors in Colorectal Cancer. International Journal of Molecular Sciences. 2018; 19(1):299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19010299
Chicago/Turabian StyleAngelucci, Adriano, Simona Delle Monache, Alessio Cortellini, Monica Di Padova, and Corrado Ficorella. 2018. "“Vessels in the Storm”: Searching for Prognostic and Predictive Angiogenic Factors in Colorectal Cancer" International Journal of Molecular Sciences 19, no. 1: 299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19010299