Immune Response Targeting Sjögren’s Syndrome Antigen Ro52 Suppresses Tear Production in Female Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
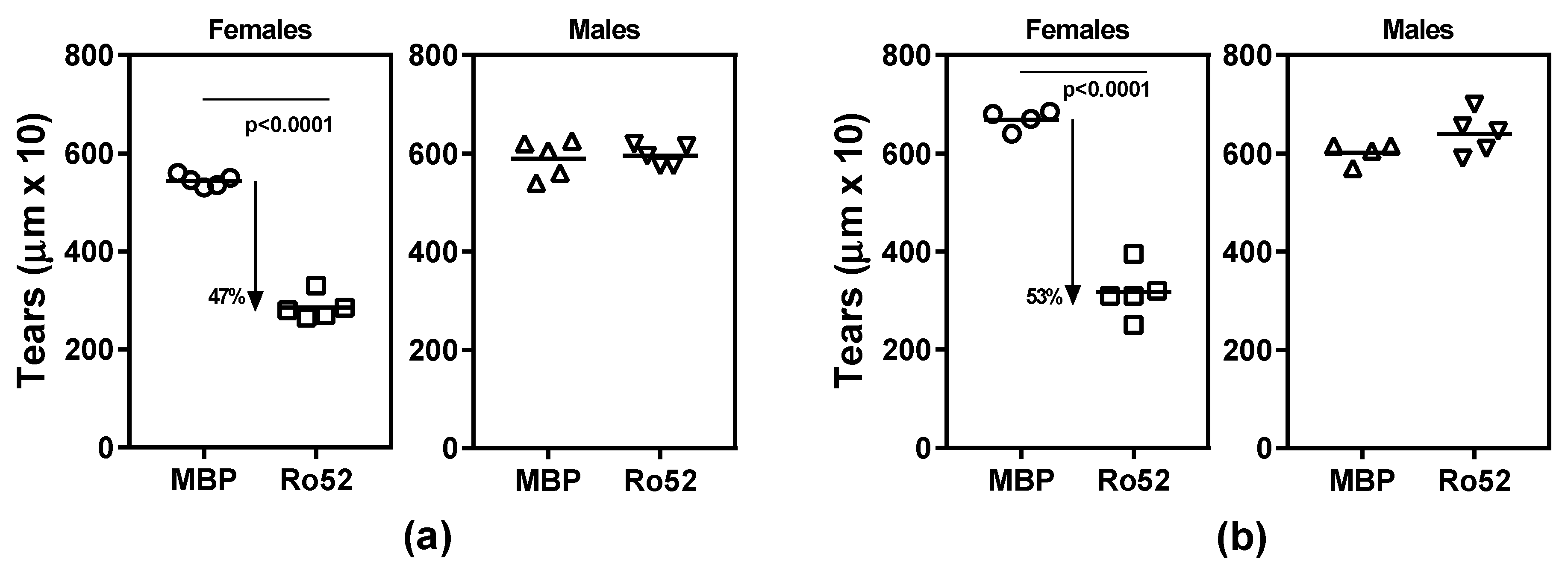
2.1. Ro52-Immunized NZM2758 Female Mice Develop Lacrimal Gland Dysfunction
2.2. Antibody Response to Ro52 Is Not Significantly Different between Female and Male Mice
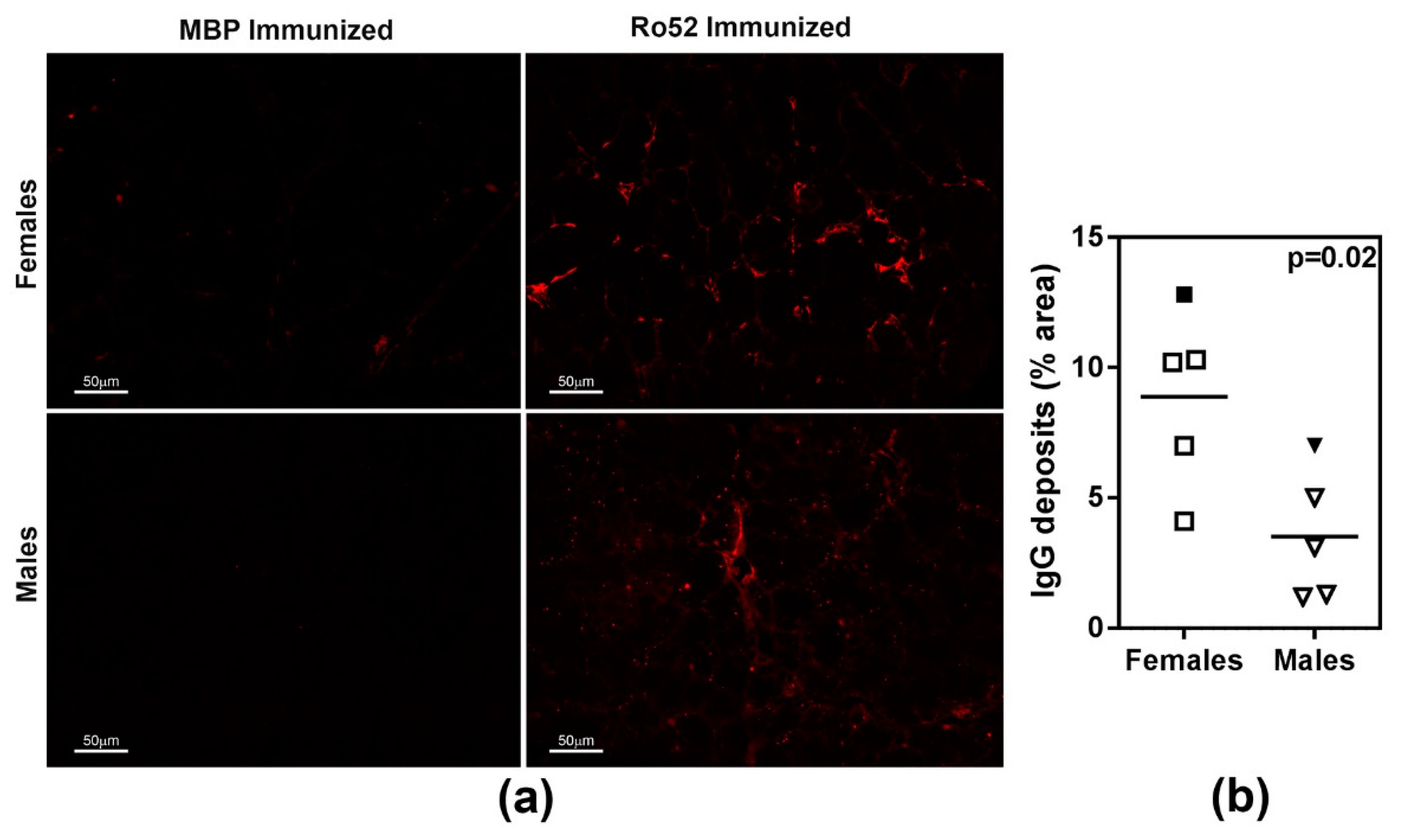
2.3. Female Mice Show Higher Severity of Antibody Deposits within Lacrimal Glands
2.4. Passive Transfer of Anti-Ro52 Immune Sera Recapitulates Lacrimal Gland Dysfunction in Female Mice
3. Discussion
4. Materials and Methods
4.1. Proteins and High Titer Immune Sera
4.2. Mice
4.3. Quantitative Immunoprecipitation Assay (IP)
4.4. Histopathology
4.5. Immunofluorescence Staining and Quantification of IgG Deposits in Lacrimal Glands
4.6. Passive Transfer of Rabbit Serum into Adjuvant Treated Mice
4.7. Statistical Methods
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| NZM | New Zealand Mixed |
| SSA | Sjögren’s Syndrome Antigen A |
| MBP | Maltose binding protein |
| IP | Immunoprecipitation |
| H&E | Hematoxylin and Eosin |
References
- Mariette, X.; Criswell, L.A. Primary Sjogren’s syndrome. N. Engl. J. Med. 2018, 378, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Foulks, G.N.; Forstot, S.L.; Donshik, P.C.; Forstot, J.Z.; Goldstein, M.H.; Lemp, M.A.; Nelson, J.D.; Nichols, K.K.; Pflugfelder, S.C.; Tanzer, J.M.; et al. Clinical guidelines for management of dry eye associated with Sjogren disease. Ocul. Surf. 2015, 13, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Szodoray, P.; Koczok, K.; Szanto, A.; Horvath, I.F.; Nakken, B.; Molnar, I.; Zeher, M. Autoantibodies to novel membrane and cytosolic antigens of the lachrymal gland in primary Sjogren’s syndrome. Clin. Rheumatol. 2008, 27, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Fernandez Castro, M.; Sanchez-Piedra, C.; Andreu, J.L.; Martinez Taboada, V.; Olive, A.; Rosas, J. Factors associated with severe dry eye in primary Sjogren’s syndrome diagnosed patients. Rheumatol. Int. 2018, 38, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef] [PubMed]
- Zoukhri, D.; Macari, E.; Kublin, C.L. A single injection of interleukin-1 induces reversible aqueous-tear deficiency, lacrimal gland inflammation, and acinar and ductal cell proliferation. Exp. Eye Res. 2007, 84, 894–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.H.; Prendergast, R.A.; Silverstein, A.M. Experimental autoimmune dacryoadenitis. I. Lacrimal gland disease in the rat. Investig. Ophthalmol. Vis. Sci. 1987, 28, 270–275. [Google Scholar]
- Liu, S.H.; Zhou, D.H. Experimental autoimmune dacryoadenitis: Purification and characterization of a lacrimal gland antigen. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2029–2036. [Google Scholar]
- Hou, A.; Bose, T.; Chandy, K.G.; Tong, L. A Chronic Autoimmune Dry Eye Rat Model with Increase in Effector Memory T Cells in Eyeball Tissue. J. Vis. Exp. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, G.; Ke, Y.; Sun, D.; Li, H.; Ihnen, M.; Jumblatt, M.M.; Foulks, G.; Wang, Y.; Bian, Y.; Kaplan, H.J.; et al. A new model of experimental autoimmune keratoconjunctivitis sicca (KCS) induced in Lewis rat by the autoantigen Klk1b22. Investig. Ophthalmol. Vis. Sci. 2009, 50, 2245–2254. [Google Scholar] [CrossRef] [PubMed]
- Mircheff, A.K.; Wang, Y.; Schechter, J.E.; Li, M.; Tong, W.; Attar, M.; Chengalvala, M.; Harmuth, J.; Prusakiewicz, J.J. Multiple Natural and Experimental Inflammatory Rabbit Lacrimal Gland Phenotypes. Ocul. Surf. 2016, 14, 460–483. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Koh, V.; Thong, B.Y. Review of autoantigens in Sjögren’s syndrome: An update. J. Inflamm. Res. 2017, 10, 97–105. [Google Scholar] [CrossRef] [PubMed]
- El Annan, J.; Jiang, G.; Wang, D.; Zhou, J.; Foulks, G.N.; Shao, H. Elevated immunoglobulin to tissue KLK11 in patients with Sjögren syndrome. Cornea 2013, 32, e90–e93. [Google Scholar] [CrossRef] [PubMed]
- Baldini, C.; Ferro, F.; Elefante, E.; Bombardieri, S. Biomarkers for Sjogren’s syndrome. Biomark. Med. 2018, 12, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Shiboski, S.C.; Shiboski, C.H.; Criswell, L.; Baer, A.; Challacombe, S.; Lanfranchi, H.; Schiodt, M.; Umehara, H.; Vivino, F.; Zhao, Y.; et al. American college of rheumatology classification criteria for Sjogren’s syndrome: A data-driven, expert consensus approach in the Sjogren’s international collaborative clinical alliance cohort. Arthritis Care Res. 2012, 64, 475–487. [Google Scholar] [CrossRef]
- Rasmussen, A.; Ice, J.A.; Li, H.; Grundahl, K.; Kelly, J.A.; Radfar, L.; Stone, D.U.; Hefner, K.S.; Anaya, J.M.; Rohrer, M.; et al. Comparison of the American-European consensus group Sjogren’s syndrome classification criteria to newly proposed American College of Rheumatology criteria in a large, carefully characterized sicca cohort. Ann. Rheum. Dis. 2014, 73, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, G.; Stein, A.J.; Fu, C.; Reinisch, K.M.; Wolin, S.L. Structural and biochemical basis for misfolded RNA recognition by the Ro autoantigen. Nat. Struct. Mol. Biol. 2006, 13, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Ozato, K.; Shin, D.M.; Chang, T.H.; Morse, H.C., 3rd. TRIM family proteins and their emerging roles in innate immunity. Nat. Rev. Immunol. 2008, 8, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, B.; Li, H.; Guo, M.; Wang, J.; Xu, Y.; Zou, X.; Deng, R.; Li, G.; Zhu, H. Trim21 promotes innate immune response to RNA viral infection through Lys27-linked polyubiquitination of MAVS. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Higgs, R.; Ni Gabhann, J.; Ben Larbi, N.; Breen, E.P.; Fitzgerald, K.A.; Jefferies, C.A. The E3 ubiquitin ligase Ro52 negatively regulates IFN-beta production post-pathogen recognition by polyubiquitin-mediated degradation of IRF3. J. Immunol. 2008, 181, 1780–1786. [Google Scholar] [CrossRef] [PubMed]
- Vinter, H.; Langkilde, A.; Ottosson, V.; Espinosa, A.; Wahren-Herlenius, M.; Raaby, L.; Johansen, C.; Iversen, L. TRIM21 is important in the early phase of inflammation in the imiquimod-induced psoriasis-like skin inflammation mouse model. Exp. Dermatol. 2017, 26, 713–720. [Google Scholar] [CrossRef] [PubMed]
- McEwan, W.A.; Tam, J.C.; Watkinson, R.E.; Bidgood, S.R.; Mallery, D.L.; James, L.C. Intracellular antibody-bound pathogens stimulate immune signaling via the Fc receptor TRIM21. Nat. Immunol. 2013, 14, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, A.Y.S. A review of the role and clinical utility of anti-Ro52/TRIM21 in systemic autoimmunity. Rheumatol. Int. 2017, 37, 1323–1333. [Google Scholar] [CrossRef] [PubMed]
- Oke, V.; Wahren-Herlenius, M. The immunobiology of Ro52 (TRIM21) in autoimmunity: A critical review. J. Autoimmun. 2012, 39, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Szczerba, B.M.; Kaplonek, P.; Wolska, N.; Podsiadlowska, A.; Rybakowska, P.D.; Dey, P.; Rasmussen, A.; Grundahl, K.; Hefner, K.S.; Stone, D.U.; et al. Interaction between innate immunity and ro52-induced antibody causes Sjogren’s syndrome-like disorder in mice. Ann. Rheum. Dis. 2016, 75, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Sroka, M.; Bagavant, H.; Biswas, I.; Ballard, A.; Deshmukh, U.S. Immune response against the coiled coil domain of Sjogren’s syndrome associated autoantigen Ro52 induces salivary gland dysfunction. Clin. Exp. Rheumatol. 2018, 36 (Suppl. 112), 41–46. [Google Scholar] [PubMed]
- Yu, X.; Riemekasten, G.; Petersen, F. Autoantibodies against muscarinic acetylcholine receptor m3 in Sjogren’s syndrome and corresponding mouse models. Front. Biosci. (Landmark Ed.) 2018, 23, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.H.; Brayer, J.; Cha, S.; Diggs, S.; Yasunari, U.; Hilal, G.; Peck, A.B.; Humphreys-Beher, M.G. Evidence for antimuscarinic acetylcholine receptor antibody-mediated secretory dysfunction in nod mice. Arthritis Rheum. 2000, 43, 2297–2306. [Google Scholar] [CrossRef]
- Bacman, S.; Sterin-Borda, L.; Camusso, J.J.; Arana, R.; Hubscher, O.; Borda, E. Circulating antibodies against rat parotid gland M3 muscarinic receptors in primary Sjogren’s syndrome. Clin. Exp. Immunol. 1996, 104, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Pelegrino, F.S.; Volpe, E.A.; Gandhi, N.B.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Deletion of interferon-gamma delays onset and severity of dacryoadenitis in CD25KO mice. Arthritis Res. Ther. 2012, 14, R234. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, J.; Huang, Q.; Deng, F.; Huang, R.; Zhao, W.; Yin, J.; Song, L.; Chen, J.; Gao, X.; et al. Autoantibodies against the second extracellular loop of M3R do neither induce nor indicate primary Sjogren’s syndrome. PLoS ONE 2016, 11, e0149485. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.E.; Schaumburg, C.S.; Siemasko, K.F.; Gao, J.; Wheeler, L.A.; Grupe, D.A.; De Paiva, C.S.; Calder, V.L.; Calonge, M.; Niederkorn, J.Y.; et al. Autoantibodies contribute to the immunopathogenesis of experimental dry eye disease. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2062–2075. [Google Scholar] [CrossRef] [PubMed]
- Bagavant, H.; Nandula, S.R.; Kaplonek, P.; Rybakowska, P.D.; Deshmukh, U.S. Alum, an aluminum-based adjuvant, induces Sjögren’s syndrome-like disorder in mice. Clin. Exp. Rheumatol. 2014, 32, 251–255. [Google Scholar] [PubMed]
- Sullivan, D.A.; Rocha, E.M.; Aragona, P.; Clayton, J.A.; Ding, J.; Golebiowski, B.; Hampel, U.; McDermott, A.M.; Schaumberg, D.A.; Srinivasan, S.; et al. TFOS DEWS II sex, gender, and hormones report. Ocul. Surf. 2017, 15, 284–333. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.M.; Jensen, R.V.; Liu, M.; Sullivan, B.D.; Lombardi, M.J.; Rowley, P.; Schirra, F.; Treister, N.S.; Suzuki, T.; Steagall, R.J.; et al. Influence of sex on gene expression in the mouse lacrimal gland. Exp. Eye Res. 2006, 82, 13–23. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trzeciak, M.; Bagavant, H.; Papinska, J.; Deshmukh, U.S. Immune Response Targeting Sjögren’s Syndrome Antigen Ro52 Suppresses Tear Production in Female Mice. Int. J. Mol. Sci. 2018, 19, 2935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102935
Trzeciak M, Bagavant H, Papinska J, Deshmukh US. Immune Response Targeting Sjögren’s Syndrome Antigen Ro52 Suppresses Tear Production in Female Mice. International Journal of Molecular Sciences. 2018; 19(10):2935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102935
Chicago/Turabian StyleTrzeciak, Marta, Harini Bagavant, Joanna Papinska, and Umesh S. Deshmukh. 2018. "Immune Response Targeting Sjögren’s Syndrome Antigen Ro52 Suppresses Tear Production in Female Mice" International Journal of Molecular Sciences 19, no. 10: 2935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102935