Differential Effect of Cobalt and Chromium Ions as Well as CoCr Particles on the Expression of Osteogenic Markers and Osteoblast Function

Abstract
:1. Introduction
2. Results
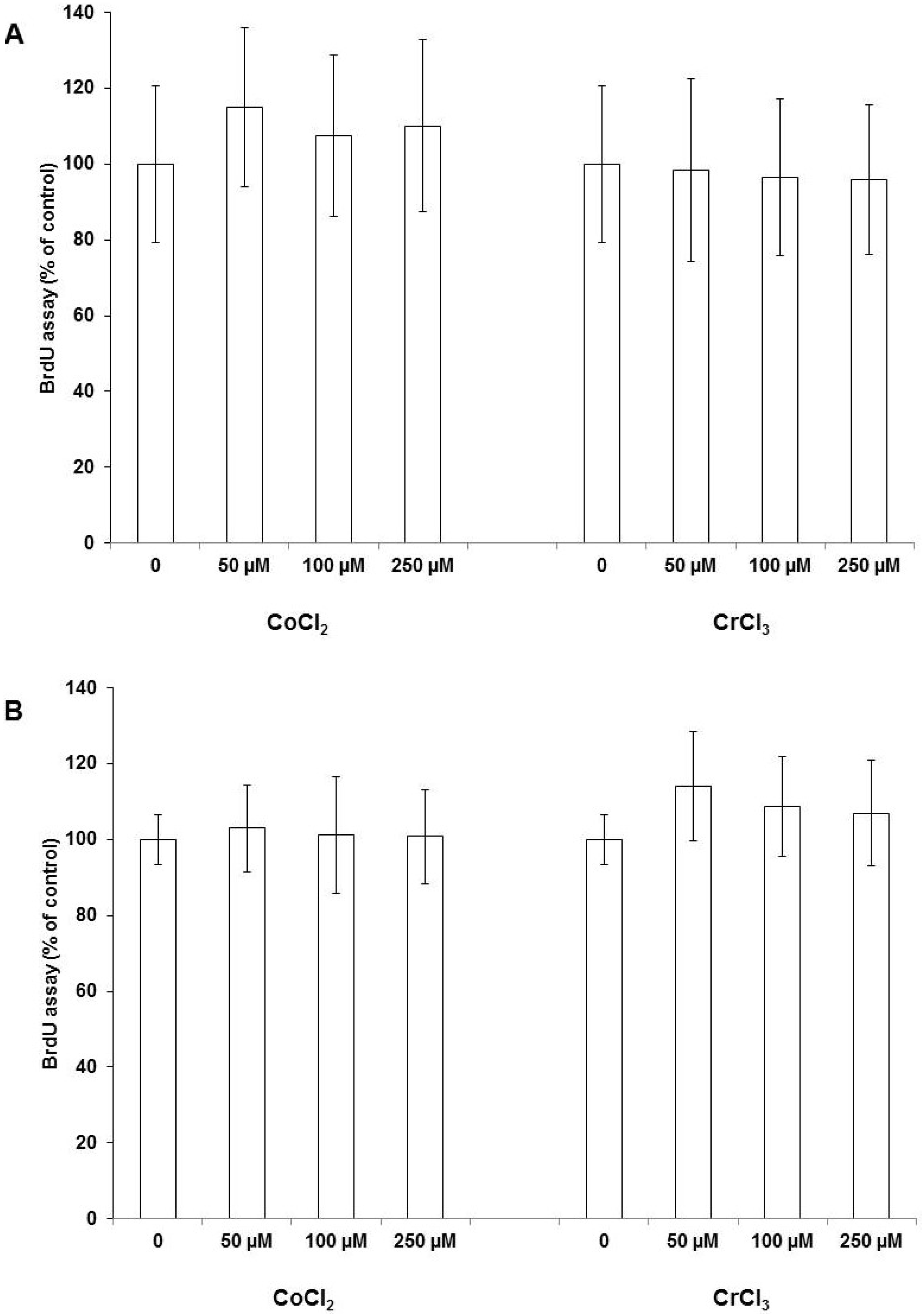
2.1. Cell Proliferation
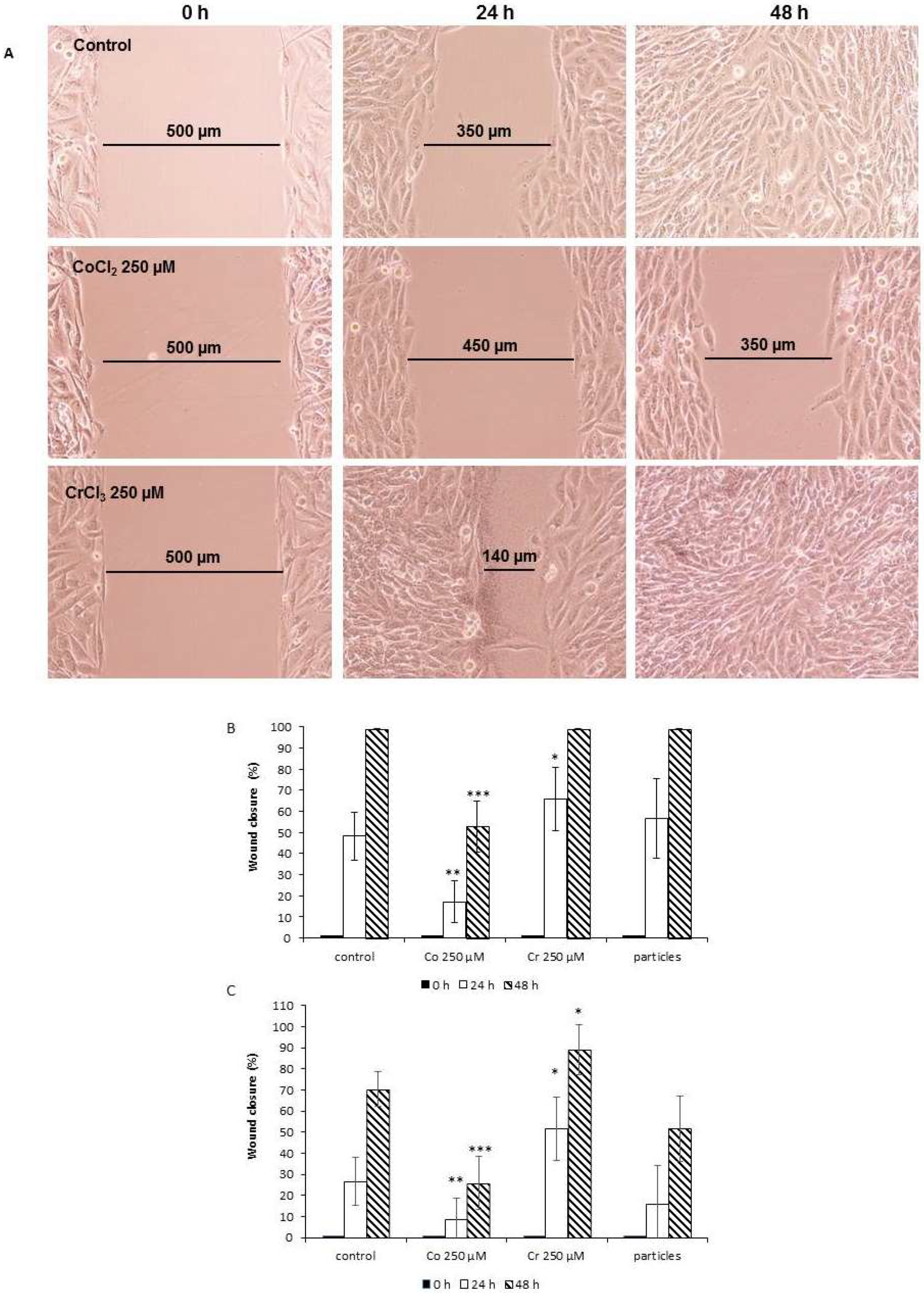
2.2. Cell Migration
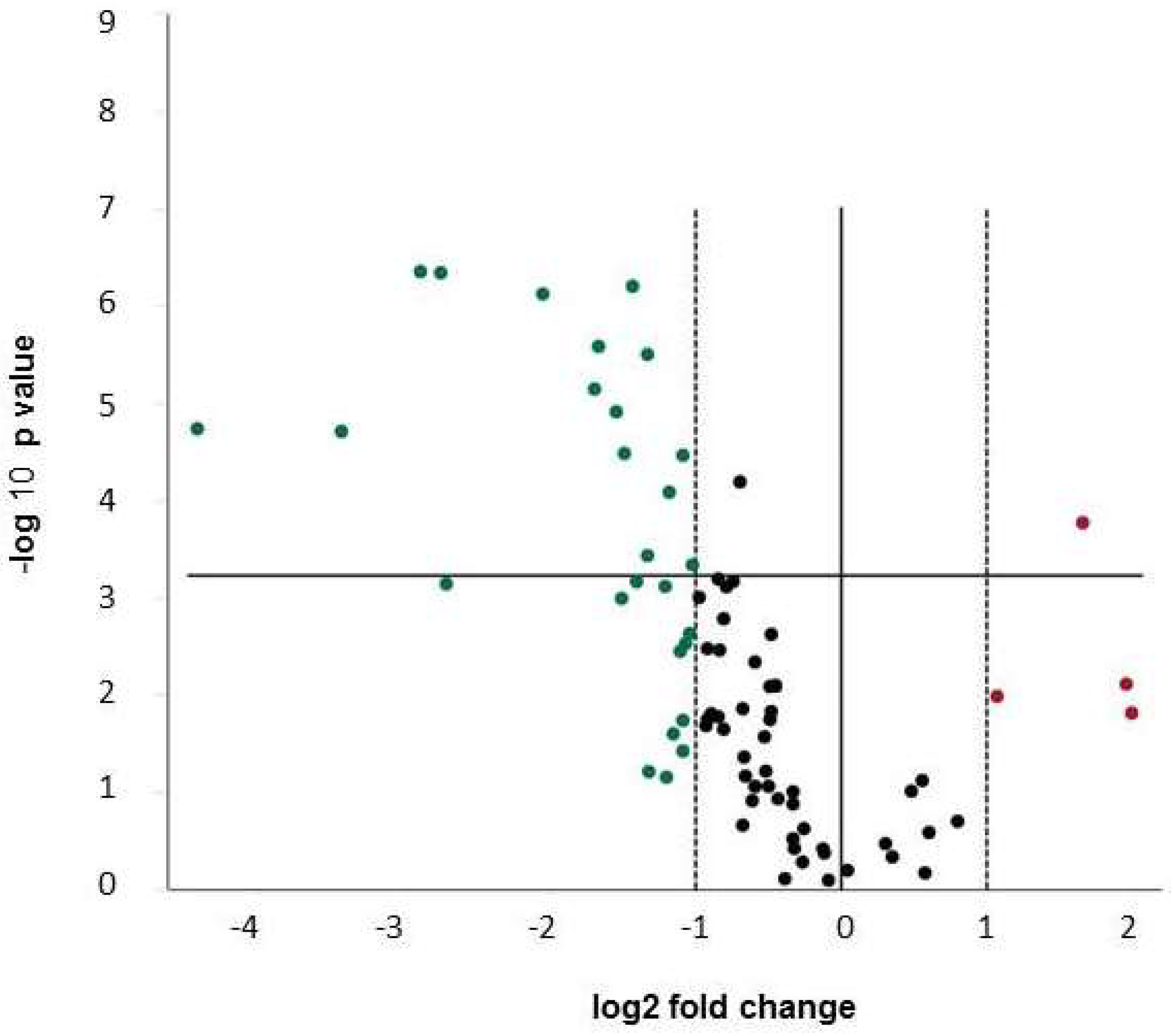
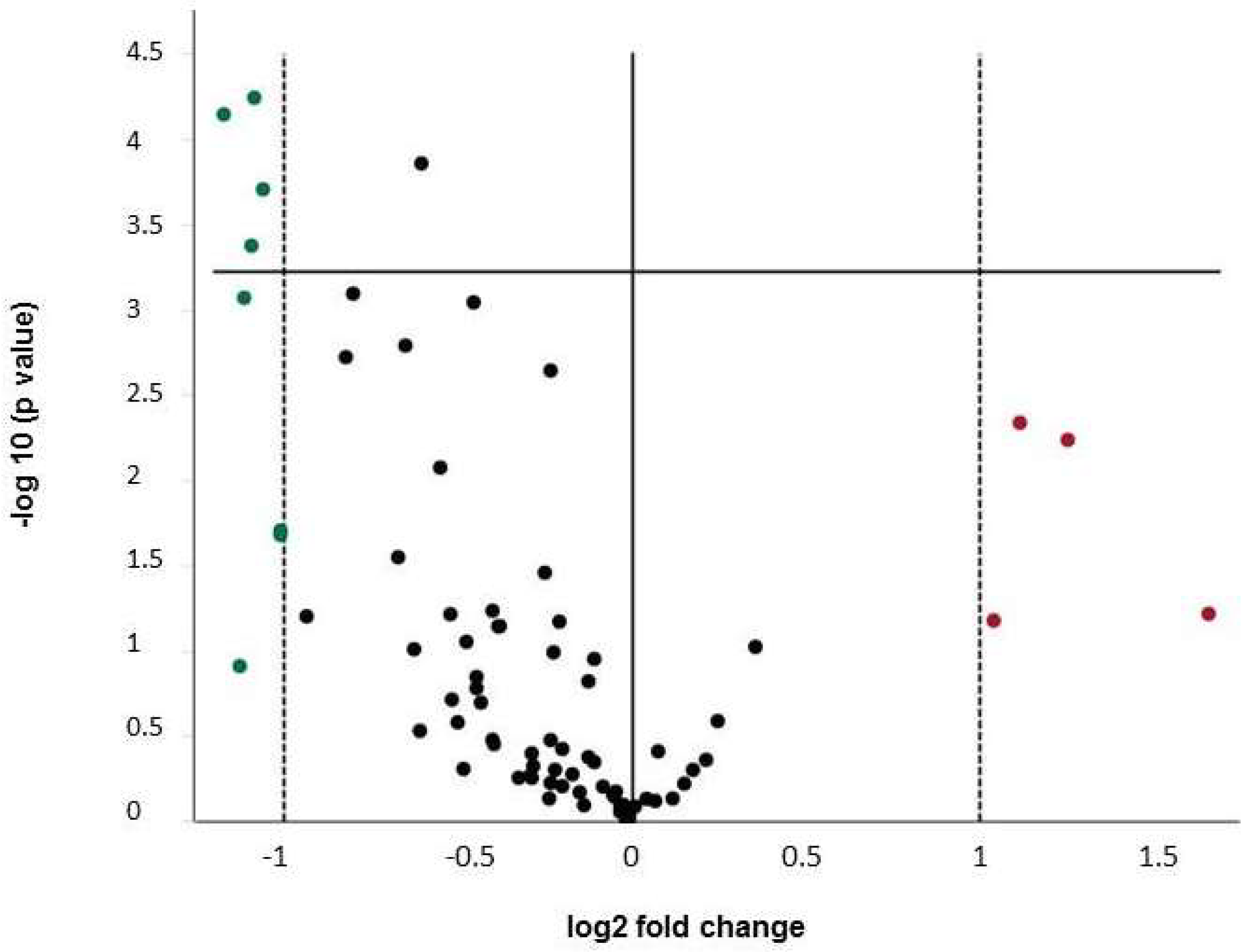
2.3. Expression of Osteogenic Markers
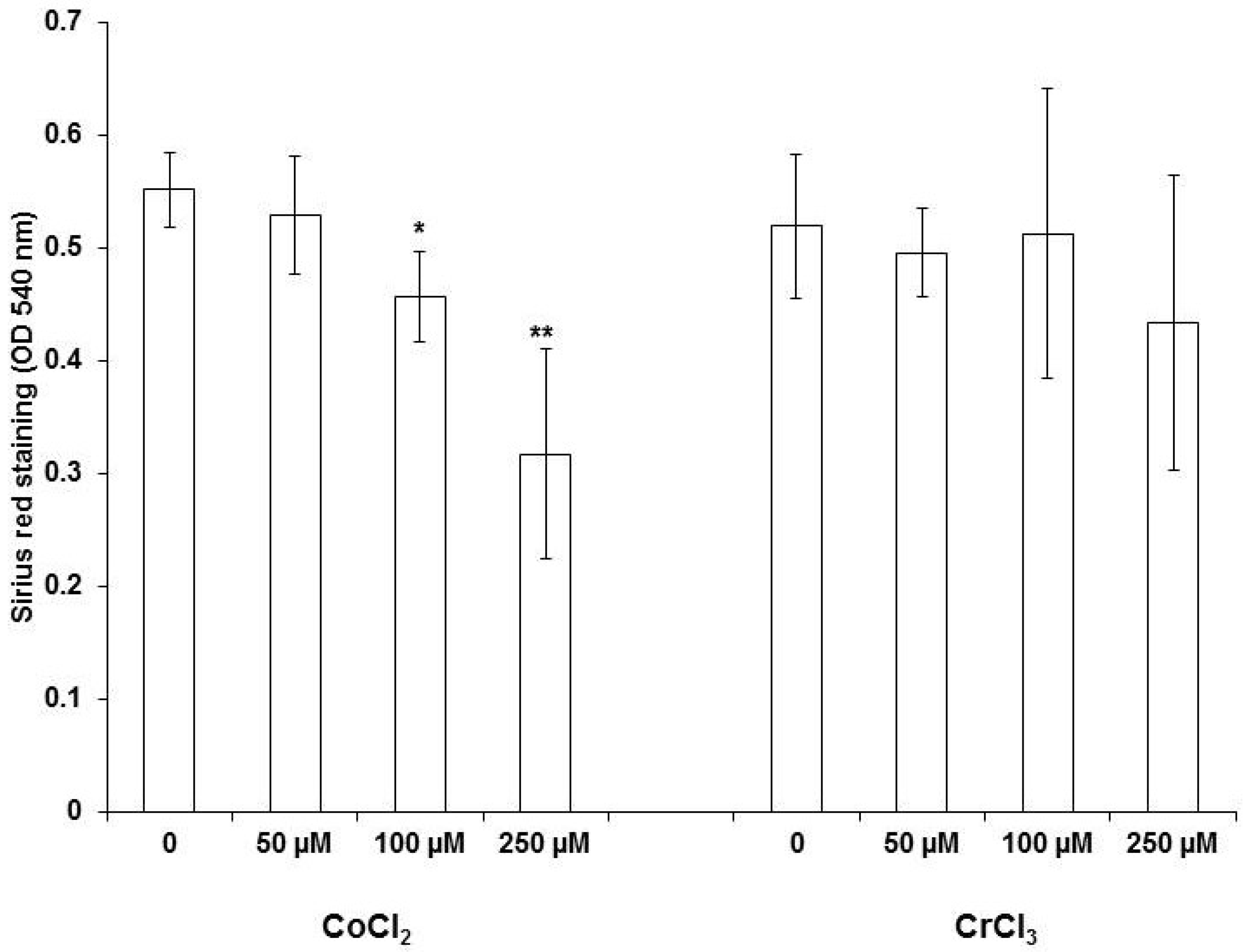
2.4. Collagen Staining with Sirius Red
3. Discussion
4. Material and Methods
4.1. Cell Culture
4.2. Preparation of CoCl2 and CrCl3 Solutions
4.3. Preparation of Particles
4.4. Cell Viability
4.5. RT2 Profiler PCR Array (PAHS-026ZF)—Human Osteogenesis
4.6. Cell Migration Assay
4.7. Detection of Proliferation
4.8. Detection of Collagen Secretion
4.9. Statistics
4.10. Analysis of Array Data
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of Variance |
| ARMD | Adverse Reactions on Metallic wear Debris |
| BMP | Bone Marrow Protein |
| BrdU | Bromodeoxyuridine |
| COL | Collagen |
| COMP | Cartilage Oligomeric Matrix Protein |
| CXCR4 | C-X-C chemokine Receptor type 4 |
| DMEM | Dulbecco’s Modified Eagle Medium |
| ECM | Extracellular Matrix |
| EDX | Energy Dispersive X-ray spectroscopy |
| FCS | Fetal Calf Serum |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HIF | Hypoxia-Inducible Factor(s) |
| HRP | Horseradish peroxidase |
| M-o-M | Metal-on-metal |
| PBS | Phosphate Buffered Saline |
| PCNA | Proliferating Cell Nuclear Antigen |
| PCR | Polymerase Chain Reaction |
| RANK | Receptor Activator of NF-κB |
| RANKL | Receptor Activator of NF-κB Ligand |
| RT-PCR | Real-Time PCR |
| SMAD | related to “MAD”(mothers against decapentaplegic) of D. Melanogaster and “Sma”(small body size) of C. elegans |
| TGF | Transforming Growth Factor |
| THA | Total Hip Arthroplasty |
| WST | Water Soluble Tetrazolium |
References
- Ebramzadeh, E.; Campbell, P.A.; Takamura, K.M.; Lu, Z.; Sangiorgio, S.N.; Kalma, J.J.; De Smet, K.A.; Amstutz, H.C. Failure modes of 433 metal-on-metal hip implants: How, why and wear. Orthop. Clin. N. Am. 2012, 42, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Morrey, B.F.; Berry, D.J.; An, K.N.; Kitaoka, H.B.; Pagnano, M.W. Joint Replacement Arthroplasty Basic Science, Hip, Knee and Ankle; Lippincott, Williams & Wilkins: Philadelphia, PA, USA, 2011; Volume 2, Chapter 51; pp. 510–514. [Google Scholar]
- Brodner, W.; Bitzan, P.; Meisinger, V.; Kaider, A.; Gottsauner-Wolf, F.; Kotz, R.M. Serum cobalt levels after metal-on metal total hip arthroplasty. J. Bone Joint Surg. Am. 2003, 85, 2168–2173. [Google Scholar] [CrossRef] [PubMed]
- Willert, H.G.; Semlitsch, M. Reactions of the articular capsule to wear products of artificial joint prostheses. J. Biomed. Mater. Res. 1977, 11, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Catelas, I.; Wimmer, M.A.; Utzschneider, S. Polyethylene and metal wear particles: Characteristics and biological effects. Semin. Immunopathol. 2011, 33, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Lhotka, C.; Szekeres, T.; Steffan, I.; Zhuber, K.; Zweymüller, K. Four-year study of cobalt and chromium blood levels in patients managed with two different metal-on-metal total hip replacements. J. Orthop. Res. 2003, 21, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Back, D.L.; Young, D.A.; Shimmin, A.J. How do serum cobalt and chromium levels change after metal-on-metal hip resurfacing? Clin. Orthop. Relat. Res. 2005, 438, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Hallab, N.J.; Anderson, S.; Caicedo, M.; Skipor, A.; Campbell, P.; Jacobs, J.J. Immune responses correlate with serum-metal in metal-on-metal hip arthroplasty. J. Arthroplasty 2004, 8, 88–93. [Google Scholar] [CrossRef]
- Andrews, R.E.; Shah, K.M.; Wilkinson, J.M.; Gartland, A. Effects of cobalt and chromium ions at clinically equivalent concentrations after metal-on-metal hip replacement on human osteoblasts and osteoclasts: Implications for skeletal health. Bone 2011, 49, 717–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapraz, F.; Besnardeau, L.; Lepage, T. Patterning of the dorsal-ventral axis in echinoderms: Insights into the evolution of the BMP-chordin signaling network. PLoS Biol. 2009, 7, e1000248. [Google Scholar] [CrossRef] [PubMed]
- Siegel, P.M.; Massagué, J. Cytostatic and apoptotic actions of TGF-beta in homeostasis and cancer. Nat. Rev. Cancer 2003, 3, 807–821. [Google Scholar] [CrossRef] [PubMed]
- Leask, A.; Abraham, D.J. TGF-beta signaling and the fibrotic response. FASEB J. 2004, 18, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Loeys, B.L.; Mortier, G.; Dietz, H.C. Bone lessons from Marfan syndrome and related disorders: Fibrillin, TGF-B and BMP at the balance of too long and too short. Pediatr. Endocrinol. Rev. 2013, 10, 417–423. [Google Scholar] [PubMed]
- Li, G.; Virdi, A.S.; Ashhurst, D.E.; Simpson, A.H.; Triffitt, J.T. Tissues formed during distraction osteogenesis in the rabbit are determined by the distraction rate: Localization of the cells that express the mRNAs and the distribution of types I and II. Cell Biol. Int. 2000, 24, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Birk, D.E.; Mayne, R. Localization of collagen types I, III and V during tendon development. Changes in collagen types I and III are correlated with changes in fibril diameter. Eur. J. Cell Biol. 1997, 72, 352–361. [Google Scholar] [PubMed]
- Henkel, W.; Glanville, R.W. Covalent crosslinking between molecules of type I and type III collagen. The involvement of the N-terminal, nonhelical regions of the alpha 1 (I) and alpha 1 (III) chains in the formation of intermolecular crosslinks. Eur. J. Biochem. 1982, 122, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Keene, D.R.; Sakai, L.Y.; Bächinger, H.P.; Burgeson, R.E. Type III collagen can be present on banded collagen fibrils regardless of fibril diameter. J. Cell Biol. 1987, 105, 2393–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wälchli, C.; Koch, M.; Chiquet, M.; Odermatt, B.F.; Trueb, B. Tissue-specific expression of the fibril-associated collagens XII and XIV. J. Cell Sci. 1994, 107, 669–681. [Google Scholar] [PubMed]
- Lisignoli, G.; Lambertini, E.; Manferdini, C.; Gabusi, E.; Penolazzi, L.; Paolella, F.; Angelozzi, M.; Casagranda, V.; Piva, R. Collagen type XV and the ‘osteogenic status’. J. Cell Mol. Med. 2017, 21, 2236–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodman, S.B.; Ma, T. Cellular chemotaxis induced by wear particles from joint replacements. Biomaterials 2010, 31, 5045–5050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyskova, T.; Gallo, J.; Kriegova, E. The Role of the Chemokine System in Tissue Response to Prosthetic By-products Leading to Periprosthetic Osteolysis and Aseptic Loosening. Front. Immunol. 2017, 8, 1026. [Google Scholar] [CrossRef] [PubMed]
- Alrabeah, G.O.; Brett, P.; Knowles, J.C.; Petridis, H. The effect of metal ions released from different dental implant-abutment couples on osteoblast function and secretion of bone resorbing mediators. J. Dent. 2017, 66, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Drynda, S.; Drynda, A.; Feuerstein, B.; Kekow, J.; Lohmann, C.H.; Bertrand, J. The effects of cobalt and chromium ions on transforming growth factor-beta patterns and mineralization in human osteoblast-like MG63 and SaOs-2 cells. J. Biomed. Mater. Res. A 2018. [Google Scholar] [CrossRef] [PubMed]
- Willert, H.G.; Buchhorn, G.H.; Fayyazi, A.; Flury, R.; Windler, M.; Köster, G.; Lohmann, C.H. Metal-on-metal bearings and hypersensitivity in patients with artificial hip joints. A clinical and histomorphological study. J. Bone Joint Surg. Am. 2005, 87, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Natu, S.; Sidaginamale, R.P.; Gandhi, J.; Langton, D.J.; Nargol, A.V. Adverse reactions to metal debris: Histopathological features of periprosthetic soft tissue reactions seen in association with failed metal on metal hip arthroplasties. J. Clin. Pathol. 2012, 65, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.M.; Ostlere, S.J.; McLardy-Smith, P.; Athanasou, N.A.; Gill, H.S.; Murray, D.W. “Asymptomatic” pseudotumors after metal-on-metal hip resurfacing arthroplasty: Prevalence and metal ion study. Arthroplasty 2011, 26, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Lehtovirta, L.; Reito, A.; Parkkinen, J.; Peräniemi, S.; Vepsäläinen, J.; Eskelinen, A. Association between periprosthetic tissue metal content, whole blood and synovial fluid metal ion levels and histopathological findings in patients with failed metal-on metal hip replacement. PLoS ONE 2018, 13, e0197614. [Google Scholar] [CrossRef] [PubMed]
- Drynda, A.; Singh, G.; Buchhorn, G.H.; Awiszus, F.; Ruetschi, M.; Feuerstein, B.; Kliche, S.; Lohmann, C.H. Metallic wear debris may regulate CXCR4 expression in vitro and in vivo. J. Biomed. Mater. Res. A 2015, 103, 1940–1948. [Google Scholar] [CrossRef] [PubMed]
- Drynda, A.; Ren, Q.; Buchhoern, G.H.; Lohmann, C.H. The induction of CXCR4 expression in human osteoblast-like cells (MG63) by CoCr particles is regulated by the PLC-DAG-PKC pathway. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 2326–2332. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.H.; Park, S.A.; Kim, S.M.; Lim, J.Y.; Jeong, C.H.; Jun, J.A.; Oh, J.H.; Park, S.H.; Oh, W.I.; Jeun, S.S. Migration of human umbilical cord blood mesenchymal stem cells mediated by stromal cell-derived factor-1/CXCR4 axis via Akt, ERK, and p38 signal transduction pathways. Biochem. Biophys. Res. Commun. 2010, 398, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Heino, T.J.; Hentunen, T.A.; Väänänen, H.K. Osteocytes inhibit osteoclastic bone resorption through transforming growth factor-beta: Enhancement by estrogen. J. Cell Biochem. 2002, 85, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Burch, M.L.; Zheng, W.; Little, P.J. Smad linker region phosphorylation in the regulation of extracellular matrix synthesis. Cell. Mol. Life Sci. 2011, 68, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.L.; Semenza, G.L. Desferrioxamine induces erythropoietin gene expression and hypoxia-inducible factor 1 DNA-binding activity: Implications for models of hypoxia signal transduction. Blood 1993, 82, 3610–3615. [Google Scholar] [PubMed]
- Hu, Y.; Hu, D.; Yu, H.; Xu, W.; Fu, R. Hypoxia-inducible factor 1a and ROCK1 regulate proliferation and collagen synthesis in hepatic stellate cells under hypoxia. Mol. Med. Rep. 2018. [Google Scholar] [CrossRef] [PubMed]
- Raykhel, I.; Moafi, F.; Myllymäki, S.M.; Greciano, P.G.; Matlin, K.S.; Moyano, J.V.; Manninen, A.; Myllyharju, J. BAMBI is a novel HIF1-dependent modulator of TGFβ-mediated disruption of cell polarity during hypoxia. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed]
- Shah, K.M.; Orton, P.; Mani, N.; Wilkinson, J.M.; Gartland, A. Osteocyte physiology and response to fluid shear stress are impaired following exposure to cobalt and chromium: Implications for bone health following joint replacement. J. Orthop. Res. 2017, 35, 1716–1723. [Google Scholar] [CrossRef] [PubMed]
- Wutzl, A.; Brozek, W.; Lernbass, I.; Rauner, M.; Hofbauer, G.; Schopper, C.; Watzinger, F.; Peterlik, M.; Pietschmann, P. Bone morphogenetic proteins 5 and 6 stimulate osteoclast generation. J. Biomed. Mater. Res. A 2006, 77, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Sabokbar, A.; Mahoney, D.J.; Hemingway, F.; Athanasou, N.A. Non-Canonical (RANKL-Independent) Pathways of Osteoclast Differentiation and Their Role in Musculoskeletal Diseases. Clin. Rev. Allergy Immunol. 2016, 51, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, H.M. Effects of cobalt chloride on phenotypes of normal human saphenous vein smooth muscle cells. Int. J. Clin. Exp. Med. 2014, 7, 4933–4941. [Google Scholar] [PubMed]
- Poole, K.; Khairy, K.; Friedrichs, J.; Franz, C.; Cisneros, D.A.; Howard, J.; Mueller, D. Molecular-scale topographic cues induce the orientation and directional movement of fibroblasts on two-dimensional collagen surfaces. J. Mol. Biol. 2005, 349, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Goel, S.A.; Guo, L.W.; Shi, X.D.; Kundi, R.; Sovinski, G.; Seedial, S.; Liu, B.; Kent, K.C. Preferential secretion of collagen type 3 versus type 1 from adventitial fibroblasts stimulated by TGF-β/Smad3-treated medial smooth muscle cells. Cell. Signal. 2013, 25, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silver, M.H.; Foidart, J.M.; Pratt, R.M. Distribution of fibronectin and collagen during mouse limb and palate development. Differentiation 1981, 18, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Maehata, Y.; Takamizawa, S.; Ozawa, S.; Izukuri, K.; Kato, Y.; Sato, S.; Lee, M.C.; Kimura, A.; Hata, R. Type III collagen is essential for growth acceleration of human osteoblastic cells by ascorbic acid 2-phosphate, a long-acting vitamin C derivative. Matrix Biol. 2007, 26, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.D.; Dusevich, V.; Feng, J.Q.; Manolagas, S.C.; Jilka, R.L. Extracellular matrix made by bone marrow cells facilitates expansion of marrow-derived mesenchymal progenitor cells and prevents their differentiation into osteoblasts. J. Bone Miner. Res. 2007, 22, 1943–1956. [Google Scholar] [CrossRef] [PubMed]
- Kivirikko, S.; Mauviel, A.; Pihlajaniemi, T.; Uitto, J. Cytokine modulation of type XV collagen gene expression in human dermal fibroblast cultures. Exp. Dermatol. 1999, 8, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Mabilleau, G.; Gill, H.S.; Sabokbar, A. Cobalt and chromium ions affect human osteoclast and human osteoblast physiology in vitro. In Vitro 2012, 1, 219. [Google Scholar] [CrossRef]
- Catelas, I.; Petit, A.; Vali, H.; Fragiskatos, C.; Meilleur, R.; Zukor, D.J.; Antoniou, J.; Huk, O.L. Quantitative analysis of macrophage apoptosis vs. necrosis induced by cobalt and chromium ions in vitro. Biomaterials 2007, 26, 2441–2453. [Google Scholar] [CrossRef] [PubMed]
- Fleury, C.; Petit, A.; Mwale, F.; Antoniou, J.; Zukor, D.J.; Tabrizian, M.; Huk, O.L. Effect of cobalt and chromium ions on human MG-63 osteoblasts in vitro: Morphology, cytotoxicity, and oxidative stress. Biomaterials 2006, 27, 3351–3360. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, R.T.; James, W.M.; Zerlauth, G. 1 alpha, 25-dihydroxyvitamin D3 specific regulation of growth, morphology, and fibronectin in a human osteosarcoma cell line. J. Cell Physiol. 1985, 123, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Pautke, C.; Schieker, M.; Tischer, T.; Kolk, A.; Neth, P.; Mutschler, W.; Milz, S. Characterization of osteosarcoma cell lines MG-63, Saos-2 and U-2 OS in comparison to human osteoblasts. Anticancer Res. 2004, 24, 3743–3748. [Google Scholar] [PubMed]
- Schwartz, Z.; Lohmann, C.H.; Oefinger, J.; Bonewald, L.F.; Dean, D.D.; Boyan, B.D. Implant surface characteristics modulate differentiation behavior of cells in the osteoblastic lineage. Adv. Dent. Res. 1999, 13, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Boyan, B.D.; Schwartz, Z.; Bonewald, L.F.; Swain, L.D. Localization of 1,25-(OH)2D3-responsive alkaline phosphatase in osteoblast-like cells (ROS 17/2.8, MG 63, and MC 3T3) and growth cartilage cells in culture. J. Biol. Chem. 1989, 264, 11879–11886. [Google Scholar] [PubMed]
- Boyan, B.D.; Schwartz, Z.; Lohmann, C.H.; Sylvia, V.L.; Cochran, D.L.; Dean, D.D.; Puzas, J.E. Pretreatment of bone with osteoclasts affects phenotypic expression of osteoblast-like cells. J. Orthop. Res. 2003, 21, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Lohmann, C.H.; Dean, D.D.; Köster, G.; Casasola, D.; Buchhorn, G.H.; Fink, U.; Schwartz, Z.; Boyan, B.D. Ceramic and PMMA particles differentially affect osteoblast phenotype. Biomaterials 2002, 23, 1855–1863. [Google Scholar] [CrossRef]
- Buchhorn, G.H.; Willert, H.G.; Semlitsch, M.; Schön, S.; Steinemann, S.; Schmidt, M. Preparation, characterization, and animal testing for biocompatibility of metal particles of iron-, cobalt-, and titanium-based implant alloys. In Particulate Debris from Medical Implants: Mechanisms of Formation and Histological Consequences; John, K.R.S., Ed.; ASTM International: West Conshohocken, PA, USA, 1992; Volume 1144, pp. 177–188. [Google Scholar]
- Katzer, A.; Hockertz, S.; Buchhorn, G.H.; Loehr, J.F. In vitro toxicity and mutagenicity of CoCrMo and TiAl wear particles. Toxicology 2003, 190, 145–154. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Fold Regulation | Comments | p-Value |
|---|---|---|---|
| BGN | −2.12 | 3.45254 × 10−5 | |
| BMP4 | −2.81 | 3.30439 × 10−5 | |
| BMP5 | −21.52 | A | 1.83701 × 10−5 |
| BMPR2 | −2.15 | 0.003576291 | |
| CDH11 | −4.14 | 7.55543 × 10−7 | |
| CHRD | −2.22 | 0.025290803 | |
| COL14A1 | −6.54 | 0.000723157 | |
| COL15A1 | −6.74 | A | 4.58304 × 10−7 |
| COL1A1 | −2.52 | 0.000370615 | |
| COL1A2 | −3.23 | 7.20666 × 10−6 | |
| COL3A1 | −10.78 | 1.96377 × 10−5 | |
| COL5A1 | −2.03 | 0.000462367 | |
| COMP | −2.65 | 0.000687369 | |
| IGF1R | −2.85 | 0.001021934 | |
| ITGA1 | −2.13 | 0.037993599 | |
| ITGA3 | −2.12 | 0.018352525 | |
| MMP2 | −2.7 | 6.30662 × 10−7 | |
| RUNX2 | −2.51 | 3.16733 × 10−6 | |
| SMAD1 | −2.93 | 1.23015 × 10−5 | |
| SMAD2 | −2.06 | 0.002343501 | |
| SOX9 | −3.18 | 2.62519 × 10−6 | |
| TGFB2 | −7.4 | A | 4.4503 × 10−7 |
| TGFB3 | −2.27 | 8.28135 × 10−5 | |
| TWIST1 | −2.32 | 0.00077027 | |
| VEGFB | −2.1 | 0.002973727 |
| Gene Symbol | Fold Regulation | Comments | p-Value |
|---|---|---|---|
| BMP6 | 4 | 0.015432431 | |
| EGF | 3.89 | 0.00777753 | |
| FGF1 | 2.11 | 0.010384109 | |
| PDGFA | 3.15 | A | 0.000170489 |
| Gene Symbol | Fold Regulation | Comments | p-Value |
|---|---|---|---|
| BMP5 | −2.16 | 0.0008533 | |
| BMP6 | −2.01 | 0.0209783 | |
| COL14A1 | −2.01 | 0.0197225 | |
| COL15A1 | −2.12 | A | 5.71 × 10−5 |
| PDGFA | −2.13 | 0.0004219 | |
| SMAD1 | −2.25 | 7.161 × 10−5 | |
| SMAD4 | −2.18 | 0.1233691 | |
| VDR | −2.08 | 0.0001971 |
| Gene Symbol | Fold Regulation | Comments | p-Value |
|---|---|---|---|
| FGF1 | 2.06 | B | 0.0666043 |
| GDF10 | 3.15 | B | 0.0609019 |
| NOG | 2.39 | A | 0.0058095 |
| VEGFA | 2.17 | 0.0046092 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drynda, A.; Drynda, S.; Kekow, J.; Lohmann, C.H.; Bertrand, J. Differential Effect of Cobalt and Chromium Ions as Well as CoCr Particles on the Expression of Osteogenic Markers and Osteoblast Function. Int. J. Mol. Sci. 2018, 19, 3034. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103034
Drynda A, Drynda S, Kekow J, Lohmann CH, Bertrand J. Differential Effect of Cobalt and Chromium Ions as Well as CoCr Particles on the Expression of Osteogenic Markers and Osteoblast Function. International Journal of Molecular Sciences. 2018; 19(10):3034. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103034
Chicago/Turabian StyleDrynda, Andreas, Susanne Drynda, Jörn Kekow, Christoph Hubertus Lohmann, and Jessica Bertrand. 2018. "Differential Effect of Cobalt and Chromium Ions as Well as CoCr Particles on the Expression of Osteogenic Markers and Osteoblast Function" International Journal of Molecular Sciences 19, no. 10: 3034. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103034