Elucidating Functions of FleQ in Xanthomonas oryzae pv. oryzae by Comparative Proteomic and Phenotypic Analyses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
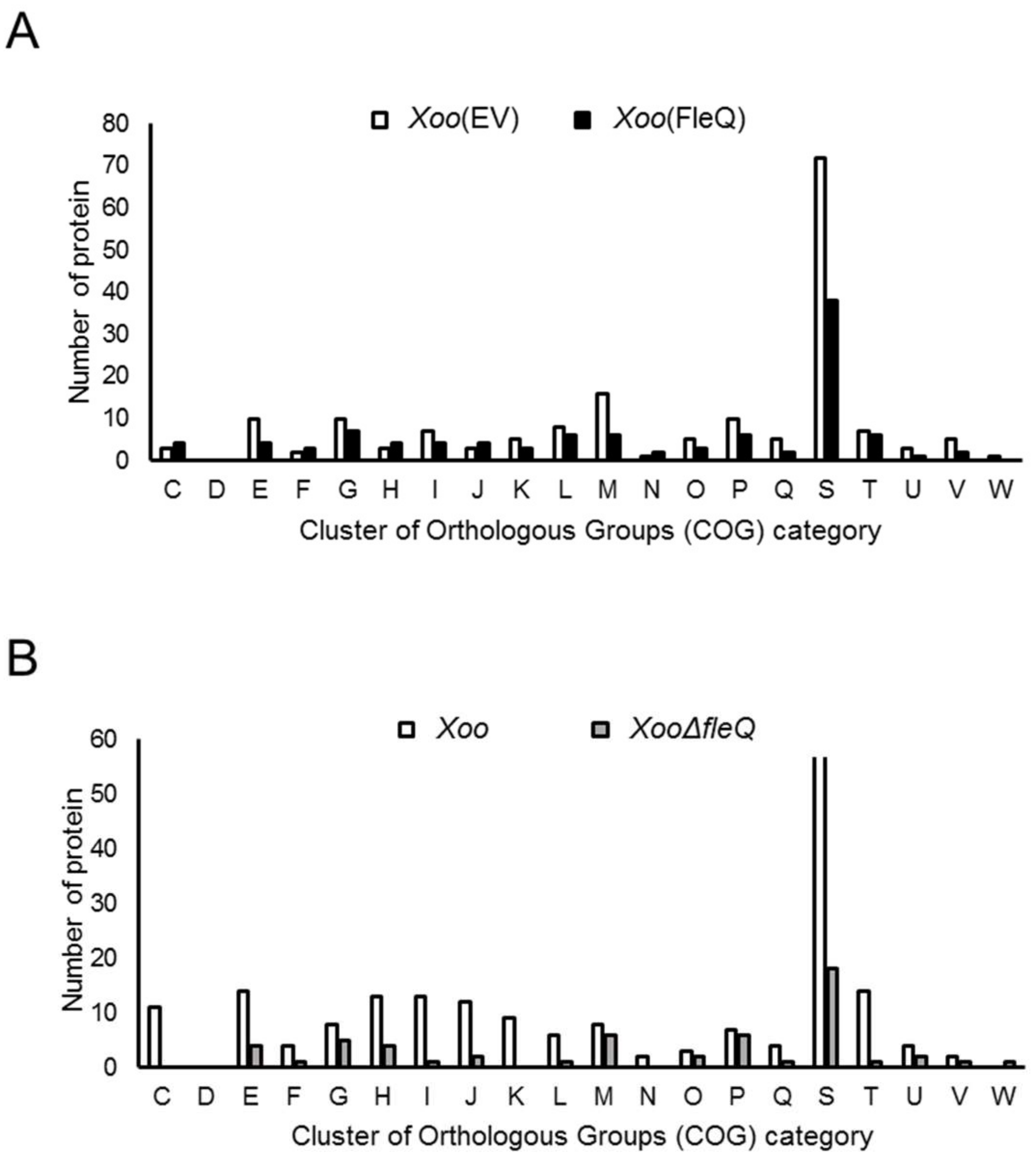
2.1. FleQ Is Involved in Controlling Expression of Genes Encoding Diverse Proteins
2.2. Overexpression of FleQ Reduced Symptom Development in Rice Plants
2.3. FleQ Affects EPS Production and Biofilm Formation
2.4. FleQ Is Crucial for Twitching and Swarming Motility
2.5. FleQ Is Involved in Siderophore Production
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Condition
4.2. Generation of Xoo Strains
4.3. Label-Free Shotgun Proteomic Analysis
4.4. Virulence Test
4.5. Motility Assay
4.6. EPS Production and Biofilm Formation
4.7. Siderophore Production
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Reddy, V.S.; Kumar, Y.N.; Raghavendra, A.; Sowjenya, G.; Kumar, S.; Ramyasree, G.; Reddy, G.R.; Kangueane, P. In silico model of DSF synthase RpfF protein from Xanthomonas oryzae pv. oryzae: A novel target for bacterial blight of rice disease. Bioinformation 2012, 8, 504–507. [Google Scholar] [PubMed]
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; Rangaraj, N.; Sonti, R.V. Multiple adhesin-like functions of Xanthomonas oryzae pv. oryzae are involved in promoting leaf attachment, entry, and virulence on rice. Mol. Plant Microbe Interact. 2009, 22, 73–85. [Google Scholar] [PubMed]
- Singh, V.B.; Kumar, A.; Isaac Kirubakaran, S.; Ayyadurai, N.; Sunish Kumar, R.; Sakthivel, N. Comparison of exopolysaccharides produced by Xanthomonas oryzae pv. oryzae strains, BXO1 and BXO8 that show varying degrees of virulence in rice (Oryza sativa L.). J. Phytopathol. 2006, 154, 410–413. [Google Scholar]
- Sinha, D.; Gupta, M.K.; Patel, H.K.; Ranjan, A.; Sonti, R.V. Cell wall degrading enzyme induced rice innate immune responses are suppressed by the type 3 secretion system effectors XopN, XopQ, XopX and XopZ of Xanthomonas oryzae pv. oryzae. PLoS ONE 2013, 8, e75867. [Google Scholar] [CrossRef] [PubMed]
- Studholme, D.J.; Dixon, R. Domain architectures of sigma54-dependent transcriptional activators. J. Bacteriol. 2003, 185, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.G.; Canals, A.; Bernues, J.; Sola, M.; Coll, M. The structure of a transcription activation subcomplex reveals how sigma(70) is recruited to PhoB promoters. EMBO J. 2011, 30, 3776–3785. [Google Scholar] [CrossRef] [PubMed]
- Burian, J.; Yim, G.; Hsing, M.; Axerio-Cilies, P.; Cherkasov, A.; Spiegelman, G.B.; Thompson, C.J. The mycobacterial antibiotic resistance determinant WhiB7 acts as a transcriptional activator by binding the primary sigma factor SigA (RpoV). Nucleic Acids Res. 2013, 41, 10062–10076. [Google Scholar] [CrossRef] [PubMed]
- Jyot, J.; Dasgupta, N.; Ramphal, R. FleQ, the major flagellar gene regulator in Pseudomonas aeruginosa, binds to enhancer sites located either upstream or atypically downstream of the RpoN binding site. J. Bacteriol. 2002, 184, 5251–5260. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Yu, C.; Li, H.; Wu, X.; Li, B.; Chen, H.; Wu, M.; He, C. Alternative sigma factor RpoN2 is required for flagellar motility and full virulence of Xanthomonas oryzae pv. oryzae. Microbiol. Res. 2015, 170, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.M.; Yang, T.C.; Yang, S.H.; Tseng, Y.H. Deduction of upstream sequences of Xanthomonas campestris flagellar genes responding to transcription activation by FleQ. Biochem. Biophys. Res. Commun. 2005, 335, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. A transcriptional activator, FleQ, regulates mucin adhesion and flagellar gene expression in Pseudomonas aeruginosa in a cascade manner. J. Bacteriol. 1997, 179, 5574–5581. [Google Scholar] [CrossRef] [PubMed]
- Hickman, J.W.; Harwood, C.S. Identification of FleQ from Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol. Microbiol. 2008, 69, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.; Wang, J.; Wang, X. FleQ regulates both the type VI secretion system and flagella in Pseudomonas putida. Biotechnol. Appl. Biochem. 2017, 65, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Millikan, D.S.; Ruby, E.G. FlrA, a sigma54-dependent transcriptional activator in Vibrio fischeri, is required for motility and symbiotic light-organ colonization. J. Bacteriol. 2003, 185, 3547–3557. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Nathawat, R.; Sinha, D.; Haque, A.S.; Sankaranarayanan, R.; Sonti, R.V. Mutations in the predicted active site of Xanthomonas oryzae pv. oryzae XopQ differentially affect virulence, suppression of host innate immunity, and induction of the HR in a nonhost plant. Mol. Plant. Microbe Interact. 2015, 28, 195–206. [Google Scholar]
- Cheong, H.; Kim, C.Y.; Jeon, J.S.; Lee, B.M.; Sun Moon, J.; Hwang, I. Xanthomonas oryzae pv. oryzae type III effector XopN targets OsVOZ2 and a putative thiamine synthase as a virulence factor in rice. PLoS ONE 2013, 8, e73346. [Google Scholar]
- Guo, X.; Zou, H.; Li, Y.; Zou, L.; Chen, G. HrpD6 gene determines Xanthomonas oryzae pv. oryzae to trigger hypersensitive response in tobacco and pathogenicity in rice. Wei Sheng Wu Xue Bao 2010, 50, 1155–1163. [Google Scholar] [PubMed]
- Wang, J.C.; So, B.H.; Kim, J.H.; Park, Y.J.; Lee, B.M.; Kang, H.W. Genome-wide identification of pathogenicity genes in Xanthomonas oryzae pv. oryzae by transposon mutagenesis. Plant Pathol. 2008, 57, 1136–1145. [Google Scholar] [CrossRef]
- Maunders, E.; Welch, M. Matrix exopolysaccharides; the sticky side of biofilm formation. FEMS Microbiol. Lett. 2017, 364, fnx120. [Google Scholar] [CrossRef] [PubMed]
- Danese, P.N.; Pratt, L.A.; Kolter, R. Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J. Bacteriol. 2000, 182, 3593–3596. [Google Scholar] [CrossRef] [PubMed]
- Tans-Kersten, J.; Huang, H.; Allen, C. Ralstonia solanacearum needs motility for invasive virulence on tomato. J. Bacteriol. 2001, 183, 3597–3605. [Google Scholar] [CrossRef] [PubMed]
- Jahn, C.E.; Willis, D.K.; Charkowski, A.O. The flagellar sigma factor FliA is required for Dickeya dadantii virulence. Mol. Plant Microbe Interact. 2008, 21, 1431–1442. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M.; Koutsoudis, M.D.; Wang, X.; von Bodman, S.B. Pantoea stewartii subsp. stewartii exhibits surface motility, which is a critical aspect of Stewart’s wilt disease development on maize. Mol. Plant Microbe Interact. 2008, 21, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Malamud, F.; Torres, P.S.; Roeschlin, R.; Rigano, L.A.; Enrique, R.; Bonomi, H.R.; Castagnaro, A.P.; Marano, M.R.; Vojnov, A.A. The Xanthomonas axonopodis pv. citri flagellum is required for mature biofilm and canker development. Microbiology 2011, 157, 819–829. [Google Scholar] [PubMed]
- Mattick, J.S. Type IV pili and twitching motility. Annu. Rev. Microbiol. 2002, 56, 289–314. [Google Scholar] [CrossRef] [PubMed]
- Russell, M.A.; Darzins, A. The pilE gene product of Pseudomonas aeruginosa, required for pilus biogenesis, shares amino acid sequence identity with the N-termini of type 4 prepilin proteins. Mol. Microbiol. 1994, 13, 973–985. [Google Scholar] [CrossRef] [PubMed]
- Wandersman, C.; Delepelaire, P. Bacterial iron sources: From siderophores to hemophores. Annu. Rev. Microbiol. 2004, 58, 611–647. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Sonti, R.V. rpfF mutants of Xanthomonas oryzae pv. oryzae are deficient for virulence and growth under low iron conditions. Mol. Plant Microbe Interact. 2002, 15, 463–471. [Google Scholar] [PubMed]
- Salzberg, S.L.; Sommer, D.D.; Schatz, M.C.; Phillippy, A.M.; Rabinowicz, P.D.; Tsuge, S.; Furutani, A.; Ochiai, H.; Delcher, A.L.; Kelley, D.; et al. Genome sequence and rapid evolution of the rice pathogen Xanthomonas oryzae pv. oryzae PXO99A. BMC Genomics 2008, 9, 204. [Google Scholar]
- Bertani, G. Lysogeny at mid-twentieth century: P1, P2, and other experimental, systems. J. Bacteriol. 2004, 186, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Park, H.J.; Bae, N.; Park, H.; Kim, D.W.; Han, S.W. Comparative proteomic analysis of three Xanthomonas spp. cultured in minimal and rich media. Proteomics 2017, 17, 1700142. [Google Scholar] [CrossRef] [PubMed]
- Bae, N.; Park, H.J.; Park, H.; Kim, M.; Han, S.W. Deciphering the functions of the outer membrane porin OprBXo involved in virulence, motility, exopolysaccharide production, biofilm formation, and stress tolerance in Xanthomonas oryzae pv. oryzae. Mol. Plant Pathol. 2018, in press. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Choi, M.; Jung, K.H.; An, G. Analysis of the early-flowering mechanisms and generation of T-DNA tagging lines in Kitaake, a model rice cultivar. J. Exp. Bot. 2013, 64, 4169–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Park, C.J.; Bae, N.; Han, S.W. Deciphering the role of tyrosine sulfation in Xanthomonas oryzae pv. oryzae using shotgun proteomic analysis. Plant. Pathol. J. 2016, 32, 266–272. [Google Scholar] [PubMed]
- Park, H.J.; Jung, H.W.; Han, S.W. Functional and proteomic analyses reveal that wxcB is involved in virulence, motility, detergent tolerance, and biofilm formation in Xanthomonas campestris pv. vesicatoria. Biochem. Biophys. Res. Commun. 2014, 452, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.P.; Park, J.; Cho, M.H.; Lee, S.W. Role of DetR in defence is critical for virulence of Xanthomonas oryzae pv. oryzae. Mol. Plant Pathol. 2016, 17, 601–613. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, N.; Park, H.-J.; Park, H.; Kim, M.; Do, E.; Han, S.-W. Elucidating Functions of FleQ in Xanthomonas oryzae pv. oryzae by Comparative Proteomic and Phenotypic Analyses. Int. J. Mol. Sci. 2018, 19, 3038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103038
Bae N, Park H-J, Park H, Kim M, Do E, Han S-W. Elucidating Functions of FleQ in Xanthomonas oryzae pv. oryzae by Comparative Proteomic and Phenotypic Analyses. International Journal of Molecular Sciences. 2018; 19(10):3038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103038
Chicago/Turabian StyleBae, Nahee, Hye-Jee Park, Hanbi Park, Minyoung Kim, Eunsoo Do, and Sang-Wook Han. 2018. "Elucidating Functions of FleQ in Xanthomonas oryzae pv. oryzae by Comparative Proteomic and Phenotypic Analyses" International Journal of Molecular Sciences 19, no. 10: 3038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103038