Dysregulated Marginal Zone B Cell Compartment in a Mouse Model of Sjögren’s Syndrome with Ocular Inflammation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
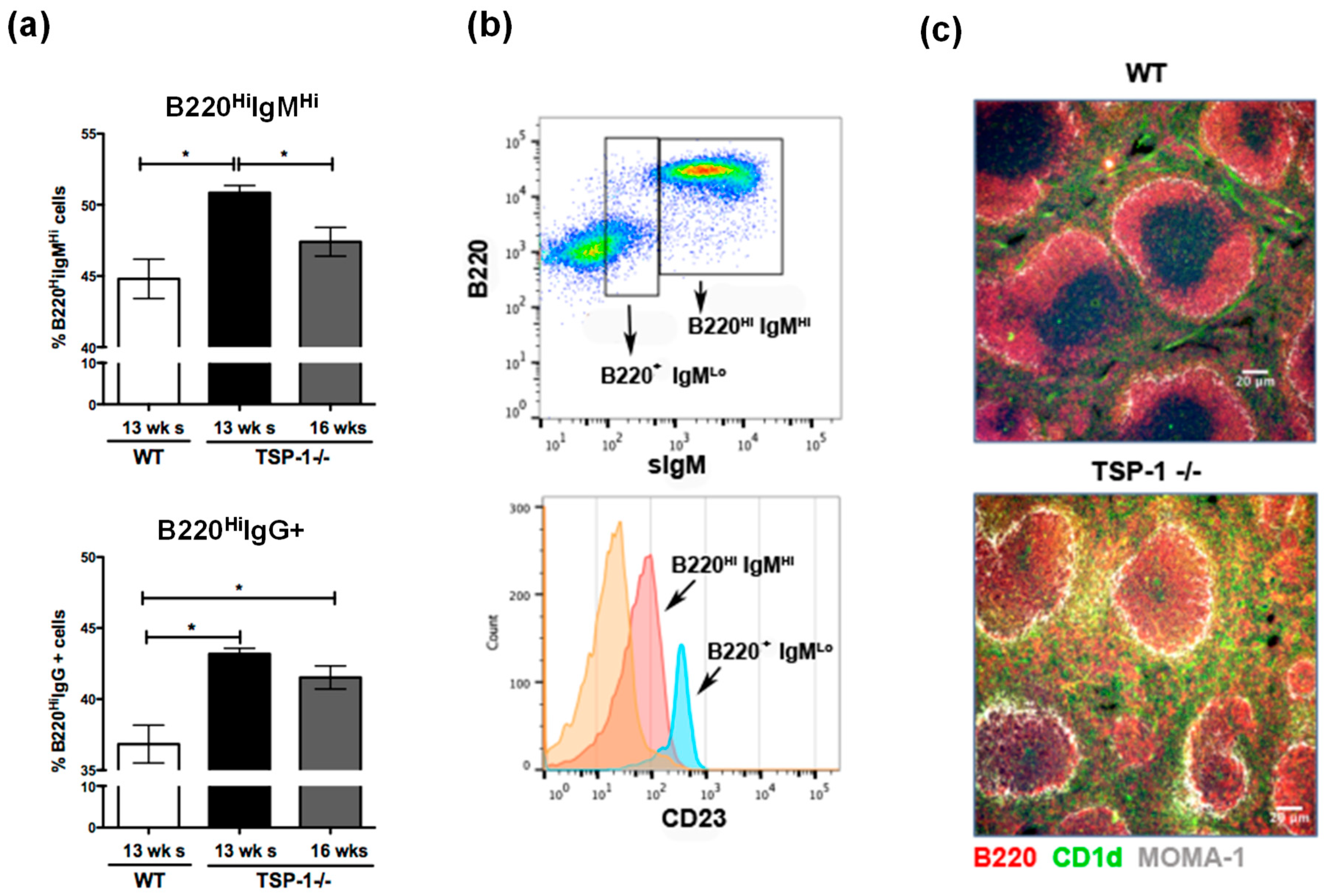
2.1. Splenic Marginal Zone Compartment Is Altered in TSP1−/− Mice
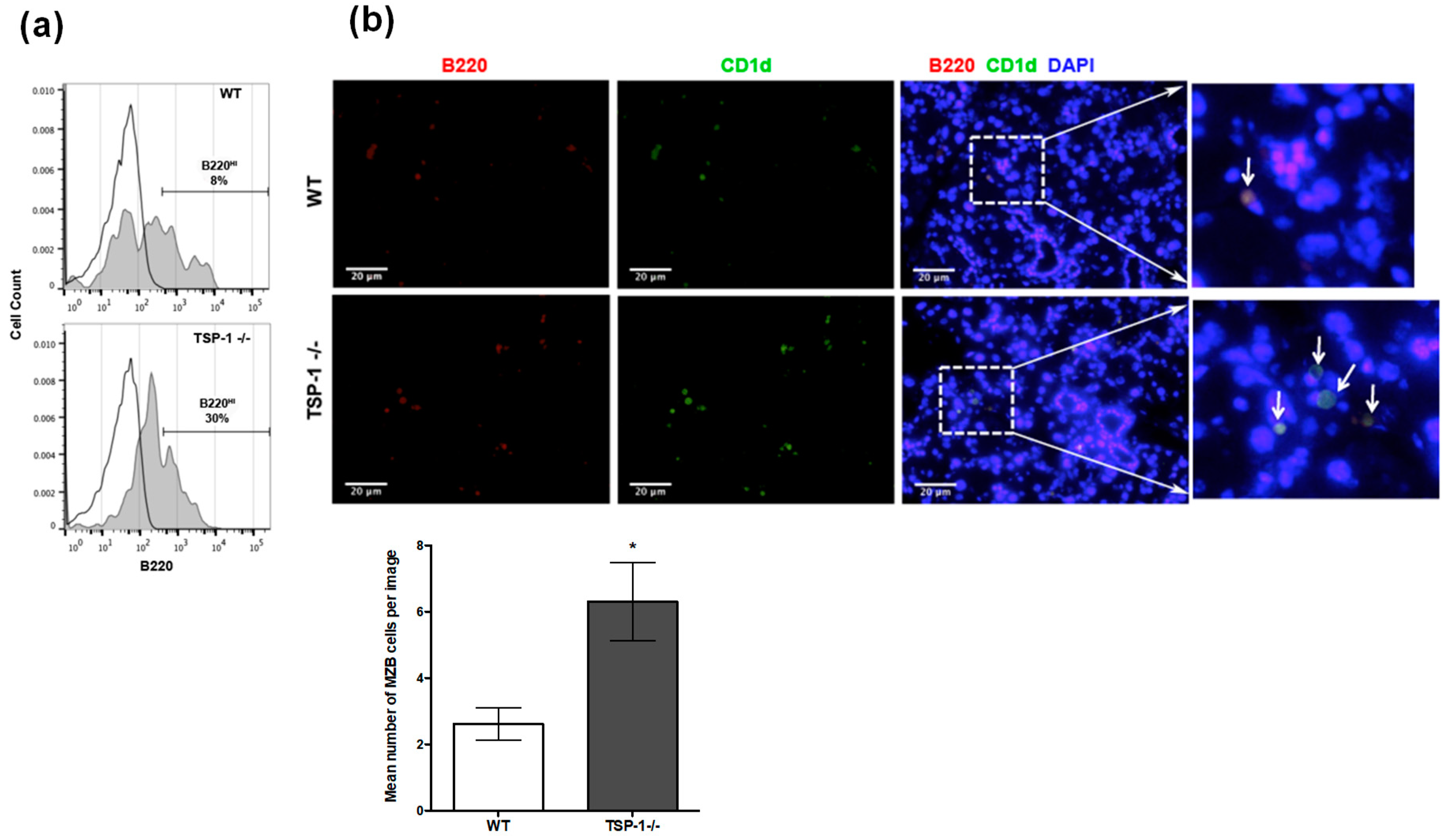
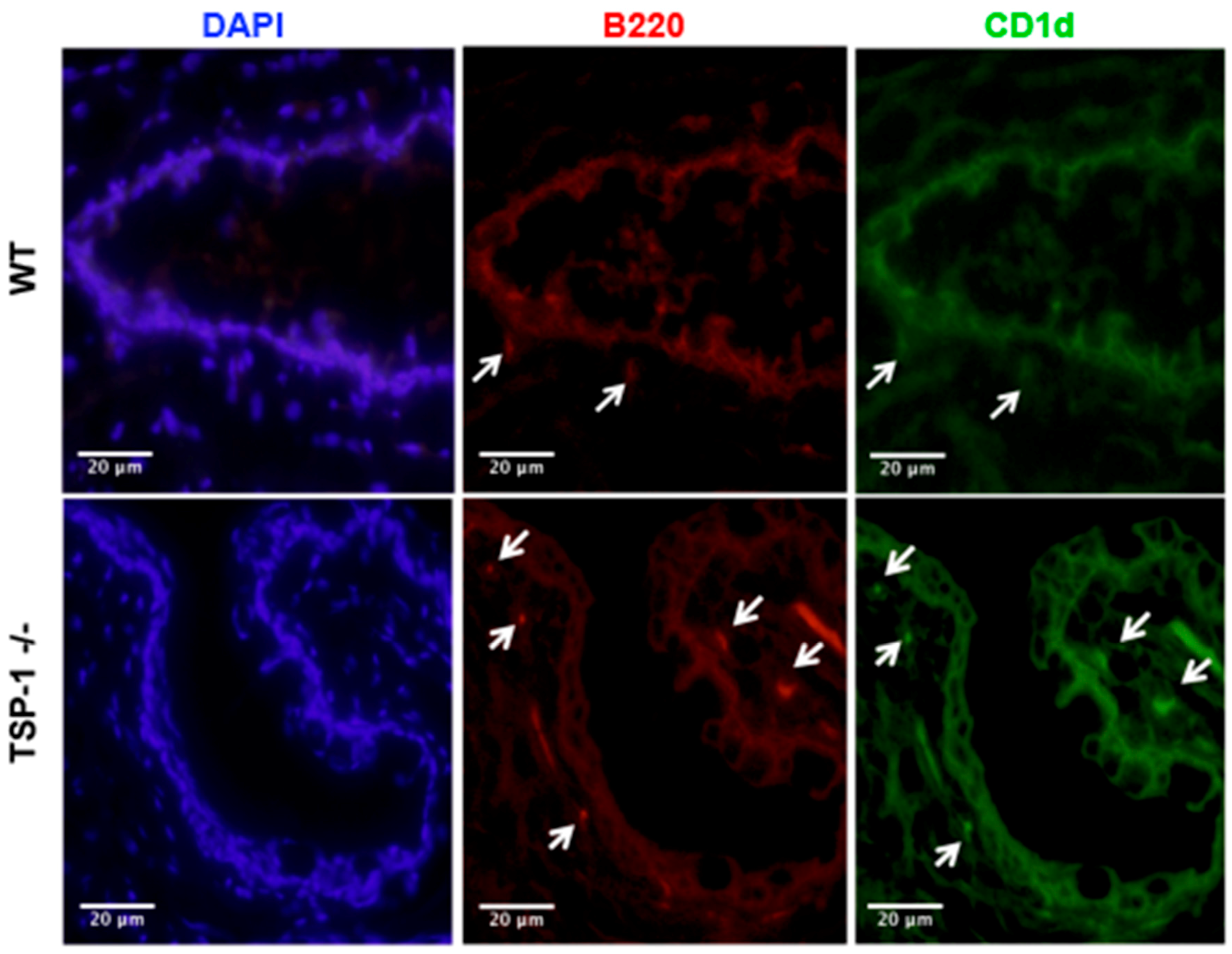
2.2. Frequency of MZB Cells Is Increased in the Lacrimal Glands of TSP1−/− Mice
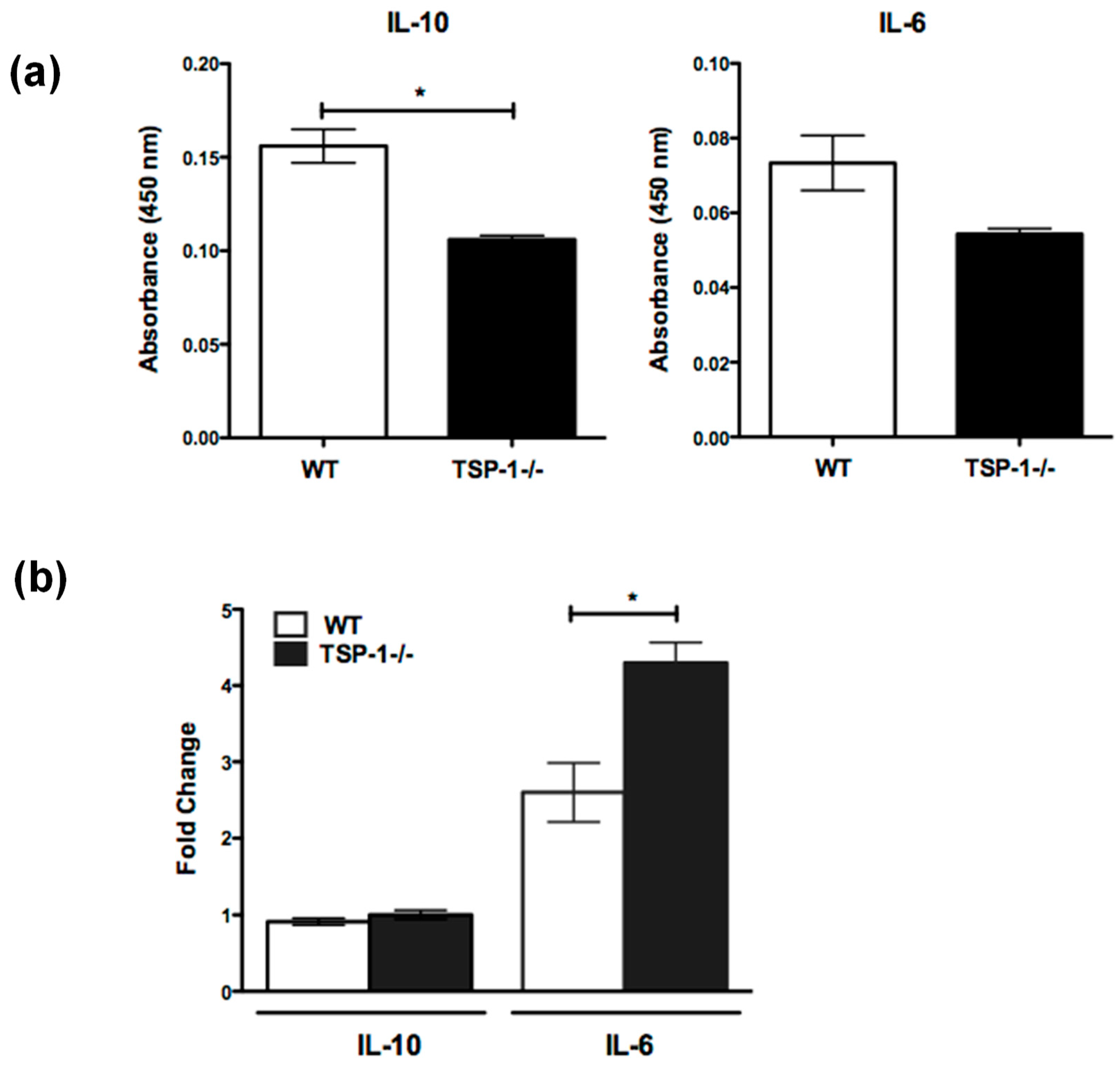
2.3. Cytokine Secretion by MZB Cells Is Altered in TSP1−/− Mice
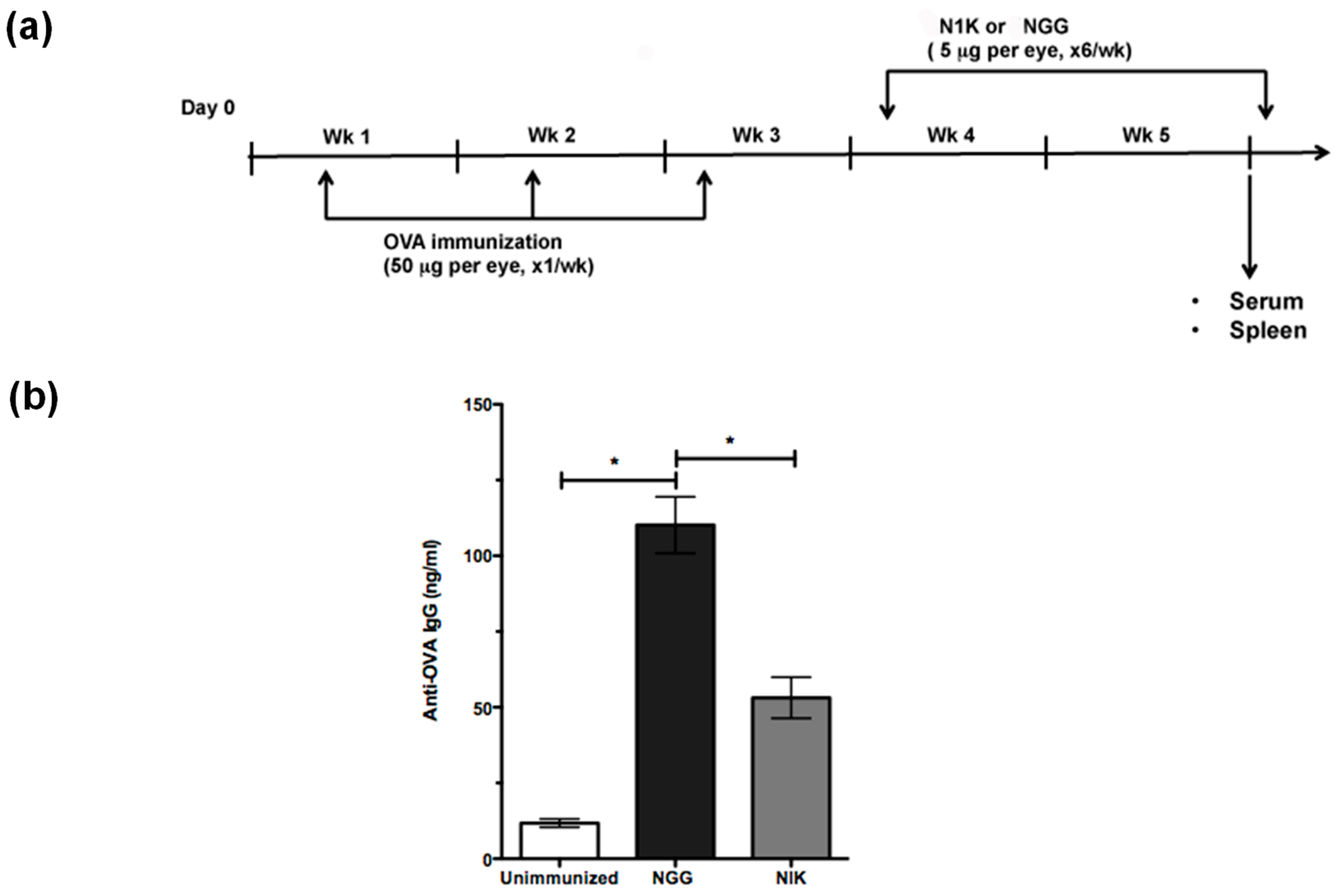
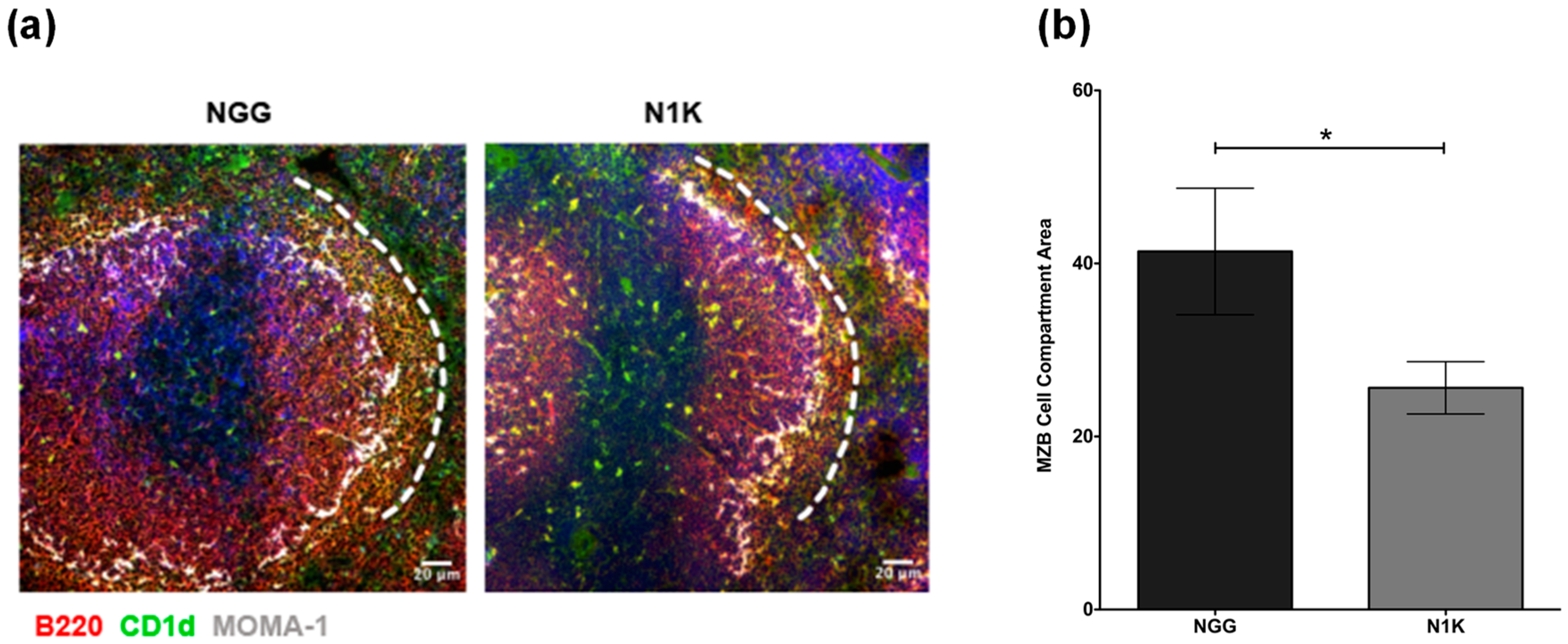
2.4. Thrombospondin-Derived Peptide Reduces Antibody Response and Splenic MZB Compartment Size in Immunized TSP1−/− Mice
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Antibodies and Reagents
4.3. Immunization and Peptide Treatment
4.4. Immunohistochemistry
4.5. Flow Cytometry
4.6. Marginal Zone B Cell Assay
4.7. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SS | Sjögren’s Syndrome |
| MZB | Marginal zone B cells |
| TSP | Thrombospondin |
| Th17 | T helper 17 |
References
- Hansen, A.; Lipsky, P.E.; Dorner, T. B cells in Sjogren’s syndrome: Indications for disturbed selection and differentiation in ectopic lymphoid tissue. Arthritis Res. Ther. 2007, 9, 218. [Google Scholar] [CrossRef] [PubMed]
- Voulgarelis, M.; Tzioufas, A.G. Current Aspects of Pathogenesis in Sjogren’s Syndrome. Ther. Adv. Musculoskelet Dis. 2010, 2, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Royer, B.; Cazals-Hatem, D.; Sibilia, J.; Agbalika, F.; Cayuela, J.M.; Soussi, T.; Maloisel, F.; Clauvel, J.P.; Brouet, J.C.; Mariette, X. Lymphomas in patients with Sjogren’s syndrome are marginal zone B-cell neoplasms, arise in diverse extranodal and nodal sites, and are not associated with viruses. Blood 1997, 90, 766–775. [Google Scholar] [PubMed]
- Daridon, C.; Pers, J.O.; Devauchelle, V.; Martins-Carvalho, C.; Hutin, P.; Pennec, Y.L.; Saraux, A.; Youinou, P. Identification of transitional type II B cells in the salivary glands of patients with Sjogren’s syndrome. Arthritis Rheum. 2006, 54, 2280–2288. [Google Scholar] [CrossRef] [PubMed]
- Zouali, M.; Richard, Y. Marginal zone B-cells, a gatekeeper of innate immunity. Front. Immunol. 2011, 2, 63. [Google Scholar] [CrossRef] [PubMed]
- Attanavanich, K.; Kearney, J.F. Marginal zone, but not follicular B cells, are potent activators of naive CD4 T cells. J. Immunol. 2004, 172, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Carvalho, T.; Kearney, J.F. Development and selection of marginal zone B cells. Immunol. Rev. 2004, 197, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Cerutti, A.; Cols, M.; Puga, I. Marginal zone B cells: Virtues of innate-like antibody-producing lymphocytes. Nat. Rev. Immunol. 2013, 13, 118–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesta, M.F. Normal structure, function, and histology of mucosa-associated lymphoid tissue. Toxicol. Pathol. 2006, 34, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Weber, J.C.; Levallois, H.; Labouret, N.; Soley, A.; Koenig, S.; Korganow, A.S.; Pasquali, J.L. Salivary gland lymphomas in patients with Sjogren’s syndrome may frequently develop from rheumatoid factor B cells. Arthritis Rheum. 2000, 43, 908–916. [Google Scholar] [CrossRef]
- Batten, M.; Groom, J.; Cachero, T.G.; Qian, F.; Schneider, P.; Tschopp, J.; Browning, J.L.; Mackay, F. BAFF mediates survival of peripheral immature B lymphocytes. J. Exp. Med. 2000, 192, 1453–1466. [Google Scholar] [CrossRef] [PubMed]
- Wellmann, U.; Werner, A.; Winkler, T.H. Altered selection processes of B lymphocytes in autoimmune NZB/W mice, despite intact central tolerance against DNA. Eur. J. Immunol. 2001, 31, 2800–2810. [Google Scholar] [CrossRef] [Green Version]
- Marino, E.; Batten, M.; Groom, J.; Walters, S.; Liuwantara, D.; Mackay, F.; Grey, S.T. Marginal-zone B-cells of nonobese diabetic mice expand with diabetes onset, invade the pancreatic lymph nodes, and present autoantigen to diabetogenic T-cells. Diabetes 2008, 57, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, C.A.; Sutherland, A.P.; Groom, J.R.; Batten, M.L.; Ng, L.G.; Gommerman, J.; Mackay, F. Development of nephritis but not sialadenitis in autoimmune-prone BAFF transgenic mice lacking marginal zone B cells. Eur. J. Immunol. 2006, 36, 2504–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; Gao, C.; Suresh, L.; Xian, Z.; Song, N.; Chaves, L.D.; Yu, M.; Ambrus, J.L., Jr. Central role for marginal zone B cells in an animal model of Sjogren’s syndrome. Clin. Immunol. 2016, 168, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Turpie, B.; Yoshimura, T.; Gulati, A.; Rios, J.D.; Dartt, D.A.; Masli, S. Sjogren’s syndrome-like ocular surface disease in thrombospondin-1 deficient mice. Am. J. Pathol. 2009, 175, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Ruiz, L.; Regenfuss, B.; Mir, F.A.; Kearns, J.; Masli, S. Conjunctival inflammation in thrombospondin-1 deficient mouse model of Sjogren’s syndrome. PLoS ONE 2013, 8, e75937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contreras Ruiz, L.; Mir, F.A.; Turpie, B.; Masli, S. Thrombospondin-derived peptide attenuates Sjogren’s syndrome-associated ocular surface inflammation in mice. Clin. Exp. Immunol. 2017, 188, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Pillai, S.; Cariappa, A.; Moran, S.T. Marginal zone B cells. Annu. Rev. Immunol. 2005, 23, 161–196. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Niu, H.; Zheng, Y.Y.; Morel, L. Autoreactive marginal zone B cells enter the follicles and interact with CD4+ T cells in lupus-prone mice. BMC Immunol. 2011, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Lal, G.; Kulkarni, N.; Nakayama, Y.; Singh, A.K.; Sethi, A.; Burrell, B.E.; Brinkman, C.C.; Iwami, D.; Zhang, T.; Hehlgans, T.; et al. IL-10 from marginal zone precursor B cells controls the differentiation of Th17, Tfh and Tfr cells in transplantation tolerance. Immunol. Lett. 2016, 170, 52–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barr, T.A.; Shen, P.; Brown, S.; Lampropoulou, V.; Roch, T.; Lawrie, S.; Fan, B.; O’Connor, R.A.; Anderton, S.M.; Bar-Or, A.; et al. B cell depletion therapy ameliorates autoimmune disease through ablation of IL-6-producing B cells. J. Exp. Med. 2012, 209, 1001–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, K.; Heaney, J.; Sibinska, Z.; Salter, D.; Savill, J.; Gray, D.; Gray, M. A tolerogenic role for Toll-like receptor 9 is revealed by B-cell interaction with DNA complexes expressed on apoptotic cells. Proc. Natl. Acad. Sci. USA 2012, 109, 887–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herlands, R.A.; Christensen, S.R.; Sweet, R.A.; Hershberg, U.; Shlomchik, M.J. T cell-independent and toll-like receptor-dependent antigen-driven activation of autoreactive B cells. Immunity 2008, 29, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.; Odendahl, M.; Reiter, K.; Jacobi, A.M.; Feist, E.; Scholze, J.; Burmester, G.R.; Lipsky, P.E.; Dorner, T. Diminished peripheral blood memory B cells and accumulation of memory B cells in the salivary glands of patients with Sjogren’s syndrome. Arthritis Rheum. 2002, 46, 2160–2171. [Google Scholar] [CrossRef] [PubMed]
- Ambrus, J.L.; Suresh, L.; Peck, A. Multiple Roles for B-Lymphocytes in Sjogren’s Syndrome. J. Clin. Med. 2016, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.P.; Katagiri, S.; Takeuchi, T.; Tsubota, K. Biopsy of labial salivary glands and lacrimal glands in the diagnosis of Sjogren’s syndrome. J. Rheumatol. 1996, 23, 76–82. [Google Scholar] [PubMed]
- Parkin, B.; Chew, J.B.; White, V.A.; Garcia-Briones, G.; Chhanabhai, M.; Rootman, J. Lymphocytic infiltration and enlargement of the lacrimal glands: A new subtype of primary Sjogren’s syndrome? Ophthalmology 2005, 112, 2040–2047. [Google Scholar] [CrossRef] [PubMed]
- Groom, J.R.; Fletcher, C.A.; Walters, S.N.; Grey, S.T.; Watt, S.V.; Sweet, M.J.; Smyth, M.J.; Mackay, C.R.; Mackay, F. BAFF and MyD88 signals promote a lupuslike disease independent of T cells. J. Exp. Med. 2007, 204, 1959–1971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, L.; Chen, Y.; Masaki, Y.; Okazaki, T.; Umehara, H. Possible Mechanisms of Lymphoma Development in Sjogren’s Syndrome. Curr. Immunol. Rev. 2013, 9, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mitsdoerffer, M.; Lee, Y.; Jager, A.; Kim, H.J.; Korn, T.; Kolls, J.K.; Cantor, H.; Bettelli, E.; Kuchroo, V.K. Proinflammatory T helper type 17 cells are effective B-cell helpers. Proc. Natl. Acad. Sci. USA 2010, 107, 14292–14297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.C.; Yang, P.; Wang, J.; Wu, Q.; Myers, R.; Chen, J.; Yi, J.; Guentert, T.; Tousson, A.; Stanus, A.L.; et al. Interleukin 17-producing T helper cells and interleukin 17 orchestrate autoreactive germinal center development in autoimmune BXD2 mice. Nat. Immunol. 2008, 9, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Baglaenko, Y.; Manion, K.P.; Chang, N.H.; Gracey, E.; Loh, C.; Wither, J.E. IL-10 Production Is Critical for Sustaining the Expansion of CD5+ B and NKT Cells and Restraining Autoantibody Production in Congenic Lupus-Prone Mice. PLoS ONE 2016, 11, e0150515. [Google Scholar] [CrossRef] [PubMed]
- Sang, A.; Zheng, Y.Y.; Yin, Y.; Dozmorov, I.; Li, H.; Hsu, H.C.; Mountz, J.D.; Morel, L. Dysregulated cytokine production by dendritic cells modulates B cell responses in the NZM2410 mouse model of lupus. PLoS ONE 2014, 9, e102151. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, N.; Chin, I.; Gabriel, P.; Blaum, E.; Masli, S. Dysregulated Marginal Zone B Cell Compartment in a Mouse Model of Sjögren’s Syndrome with Ocular Inflammation. Int. J. Mol. Sci. 2018, 19, 3117. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103117
Singh N, Chin I, Gabriel P, Blaum E, Masli S. Dysregulated Marginal Zone B Cell Compartment in a Mouse Model of Sjögren’s Syndrome with Ocular Inflammation. International Journal of Molecular Sciences. 2018; 19(10):3117. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103117
Chicago/Turabian StyleSingh, Niharika, Ian Chin, Paul Gabriel, Emily Blaum, and Sharmila Masli. 2018. "Dysregulated Marginal Zone B Cell Compartment in a Mouse Model of Sjögren’s Syndrome with Ocular Inflammation" International Journal of Molecular Sciences 19, no. 10: 3117. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103117