Floral Scent Emission from Nectaries in the Adaxial Side of the Innermost and Middle Petals in Chimonanthus praecox

 ,
,
Abstract
:1. Introduction
2. Results
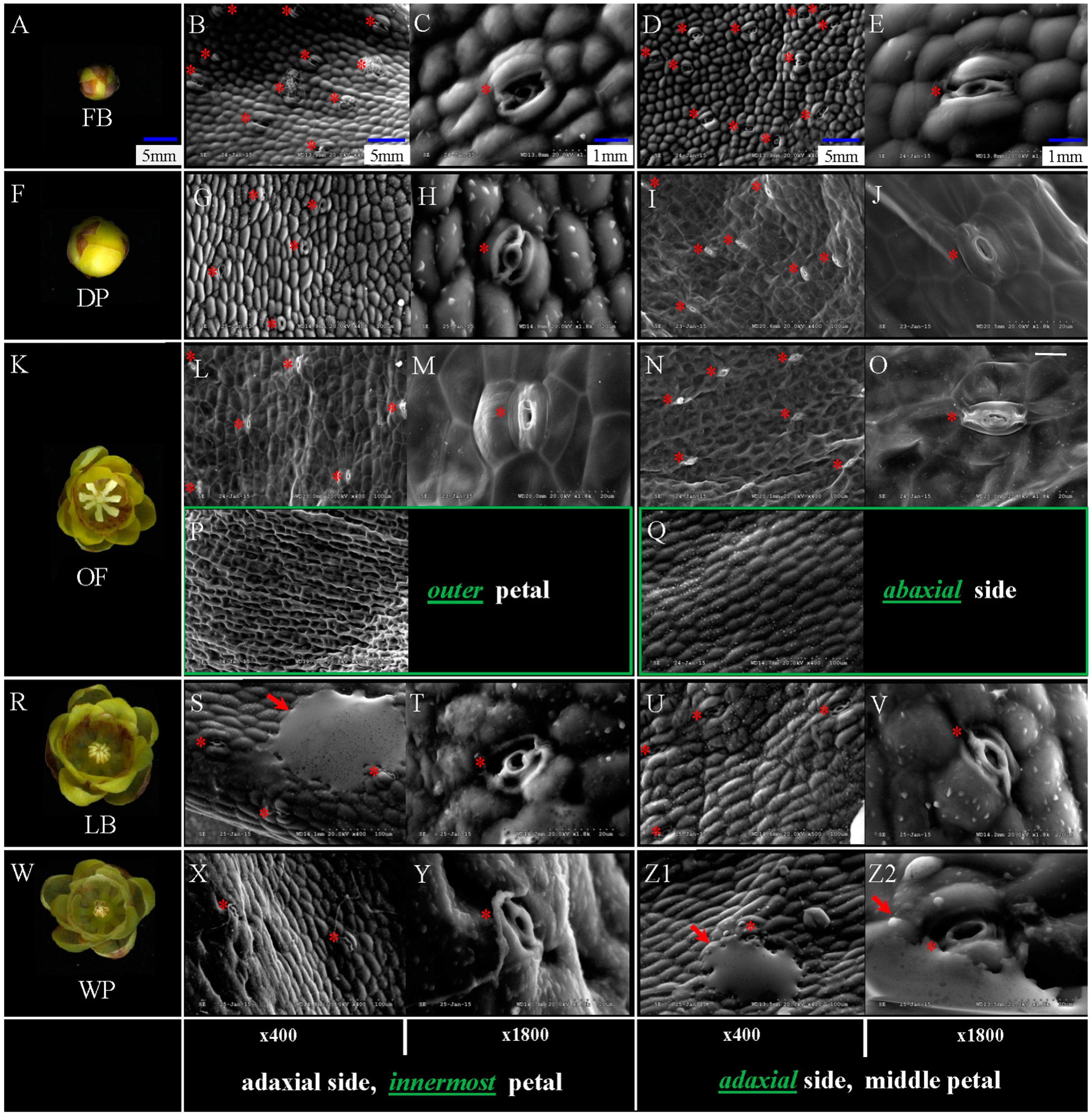
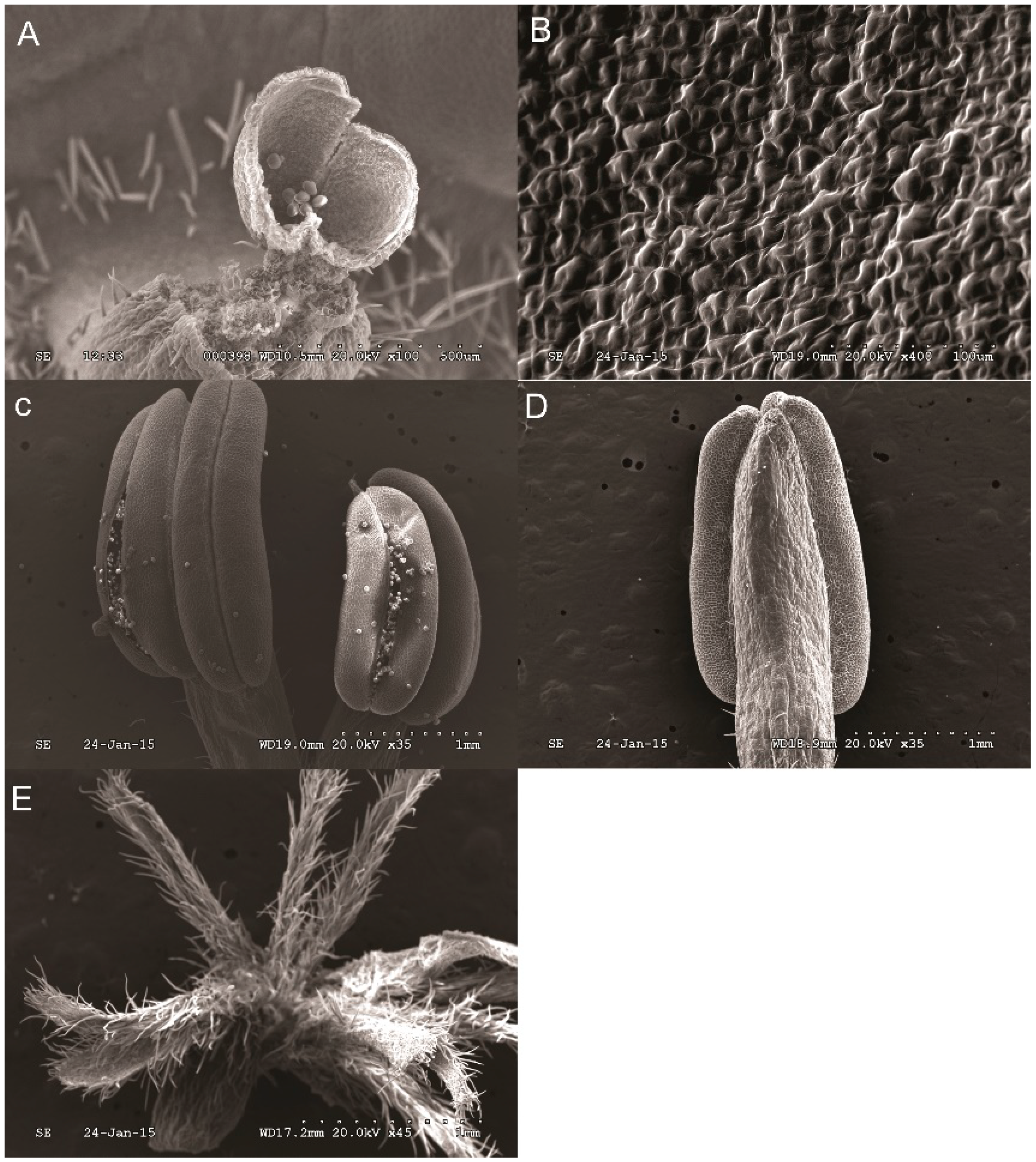
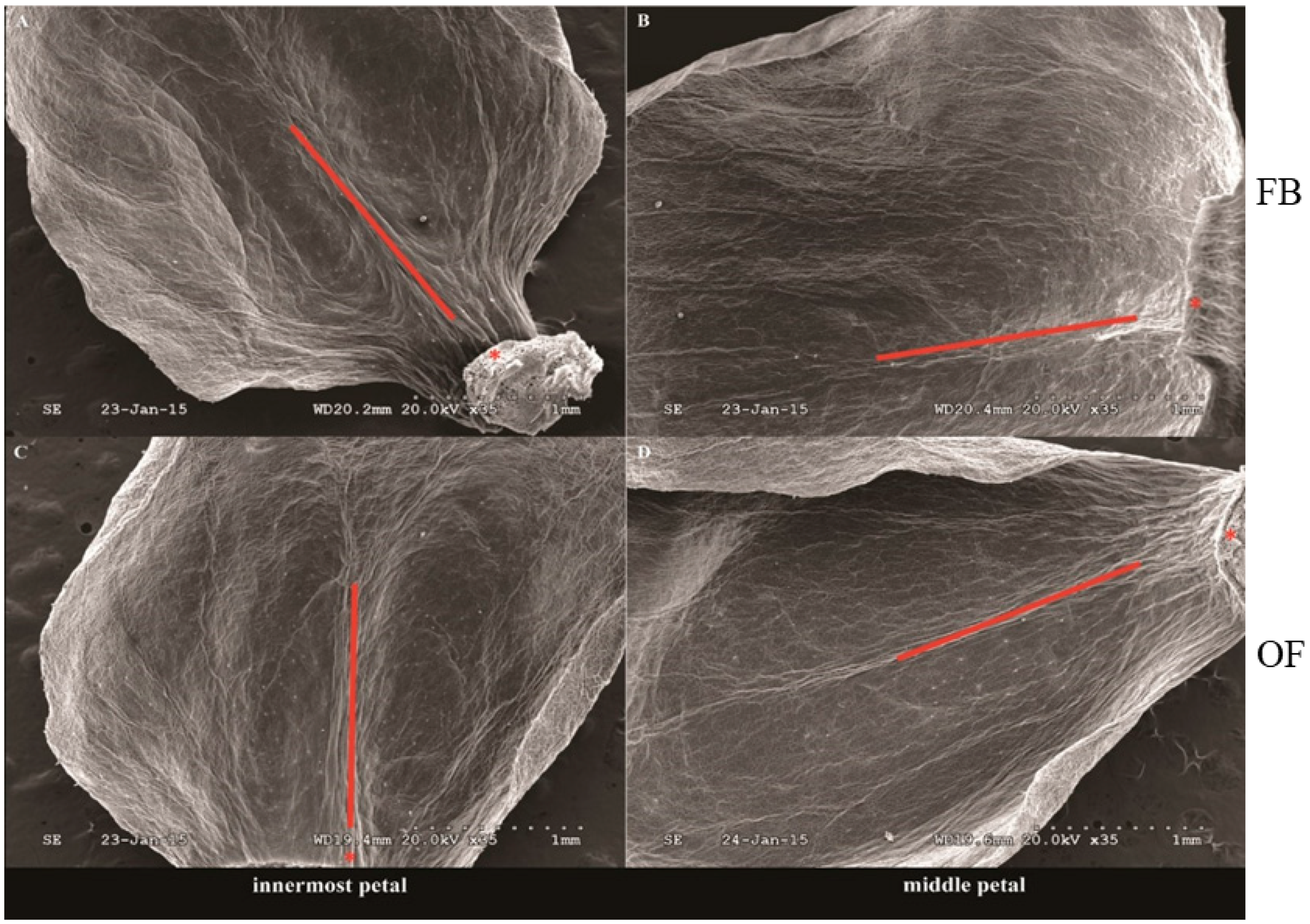
2.1. Distribution Characteristics of Nectary on Petals of Different Stages
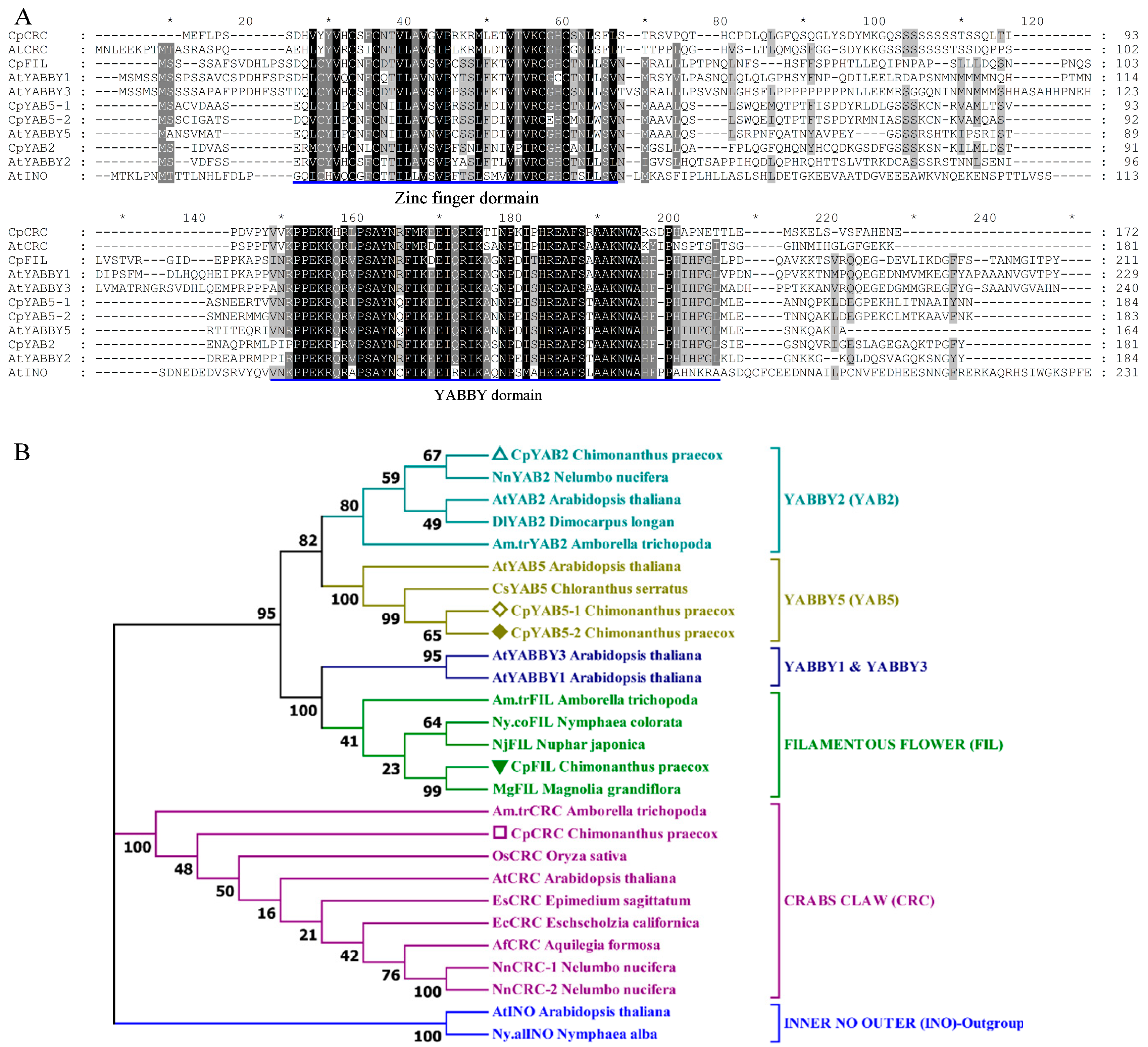
2.2. Sequence Alignment and Phylogenetic Analysis
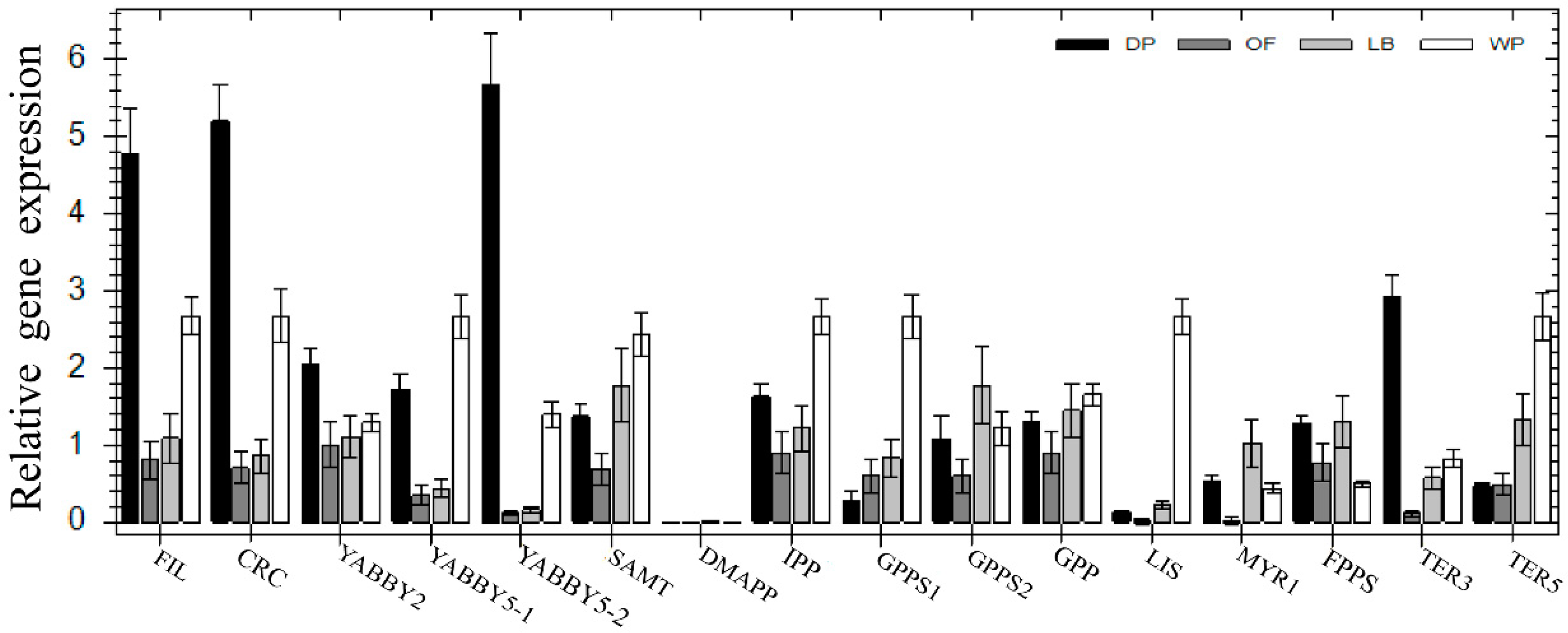
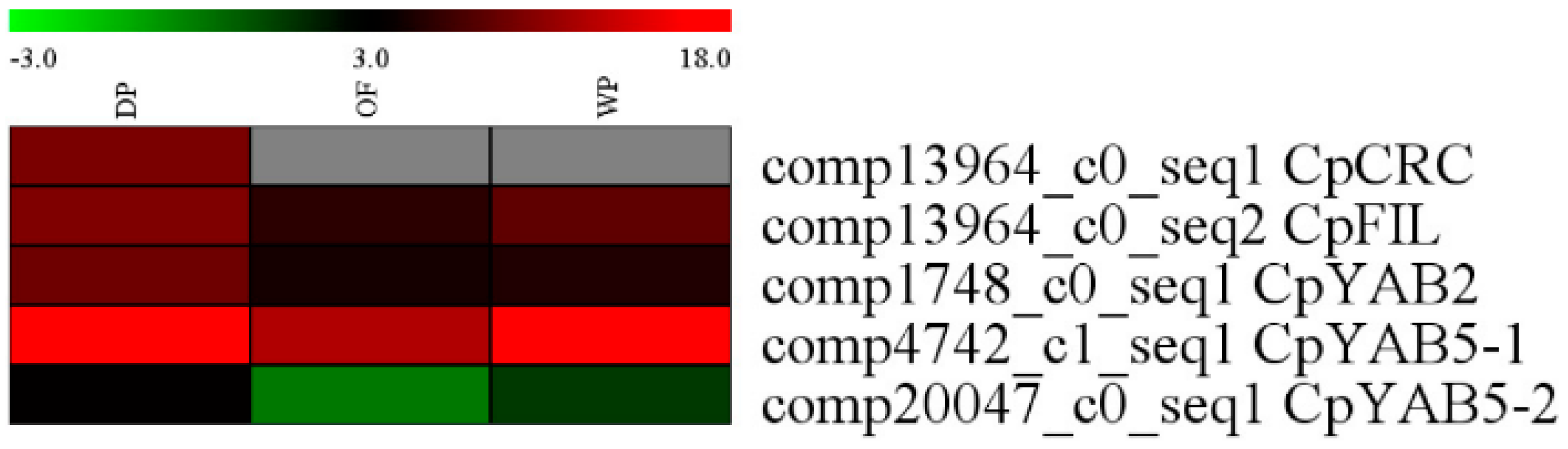
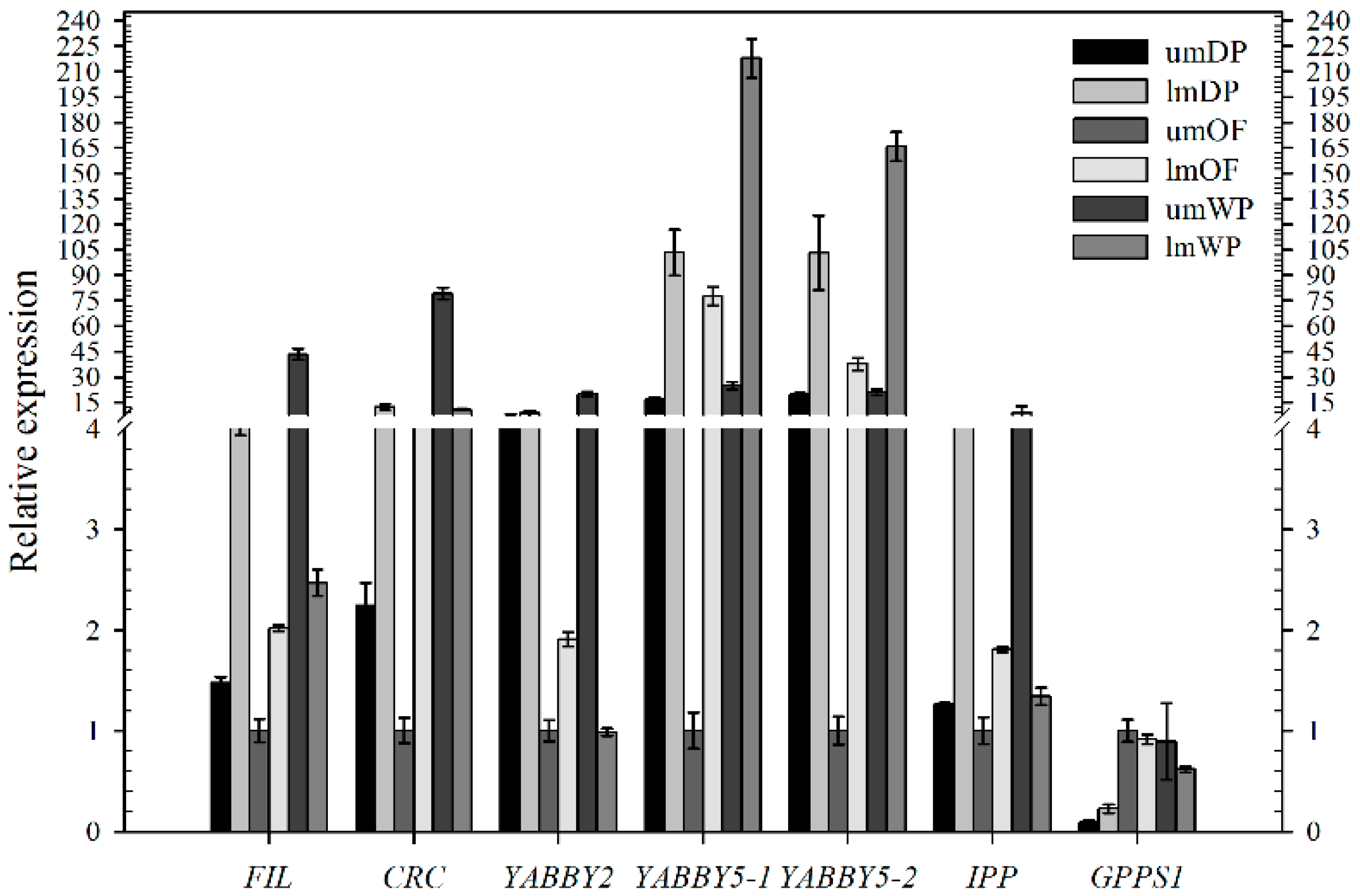
2.3. Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Scanning Electron Microscope (SEM)
4.3. Sequence Alignment and Phylogenetic Analysis
4.4. Gene Expression Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| SEM | scanning electron microscope |
| qRT-PCR | quantitative Reverse Transcript-Polymerase Chain Reaction |
| CRC | CRABS CLAW |
| FIL | FILAMENTOUS FLOWER |
| INO | INNER NO OUTER |
| YAB | YABBY |
| SAMT | S-adenosyl-L-methionine: salicylic acid carboxyl methyltransferase |
| DMAPP | dimethylallyl pyrophosphate |
| IPP | isopentenyl pyrophosphate |
| GPPS | geranyl diphosphate synthase |
| GPP | geranyl pyrophosphate |
| LIS | S-linalool synthase |
| MYR1 | myrcenesynthase |
| FPPS | farnesyl pyrophosphonate synthase |
| TER | α-terpineol synthase |
References
- Schmid, R. Reproductive versus extra-reproductive nectaries-historical perspective and terminological recommendations. Bot. Rev. 1988, 54, 179–227. [Google Scholar] [CrossRef]
- Lee, J.Y.; Baum, S.F.; Alvarez, J.; Patel, A.; Chitwood, D.H.; Bowman, J.L. Activation of CRABS CLAW in the Nectaries and Carpels of Arabidopsis. Plant Cell 2005, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Baum, S.F.; Eshed, Y.; Bowman, J.L. The Arabidopsis nectary is an ABC-independent floral structure. Development 2001, 128, 4657–4667. [Google Scholar] [PubMed]
- Bowman, J.L. The YABBY gene family and abaxial cell fate. Curr. Opin. Plant Biol. 2000, 3, 17–22. [Google Scholar] [CrossRef]
- Gross, T.; Broholm, S.; Becker, A. CRABS CLAW Acts as a Bifunctional Transcription Factor in Flower Development. Front. Plant Sci. 2018, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Goldshmidt, A.; Alvarez, J.P.; Bowman, J.L.; Eshed, Y. Signals derived from YABBY gene activities in organ primordia regulate growth and partitioning of Arabidopsis shoot apical meristems. Plant Cell 2008, 20, 1217–1230. [Google Scholar] [CrossRef] [PubMed]
- Bartholmes, C.; Hidalgo, O.; Gleissberg, S. Evolution of the YABBY gene family with emphasis on the basal eudicot Eschscholzia californica (Papaveraceae). Plant Biol. 2012, 14, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, K.R.; Eshed, Y.; Baum, S.F.; Otsuga, D.; Drews, G.N.; Bowman, J.L. Members of the YABBY gene family specify abaxial cell fate in Arabidopsis. Development 1999, 126, 4117–4128. [Google Scholar] [PubMed]
- Eshed, Y.; Baum, S.F.; Bowman, J.L. Distinct mechanisms promote polarity establishment in carpels of Arabidopsis. Cell 1999, 99, 199–209. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Nagasawa, N.; Kawasaki, S.; Matsuoka, M.; Nagato, Y.; Hirano, H.Y. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell 2004, 16, 500–509. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Xu, Y.Y.; Xu, Z.H.; Chong, K. A rice YABBY gene, OsYABBY4, preferentially expresses in developing vascular tissue. Dev. Genes Evol. 2007, 217, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, Y.; Abiko, M.; Horibata, A.; Hirano, H.Y. A transposon, Ping, is integrated into intron 4 of the DROOPING LEAF gene of rice, weakly reducing its expression and causing a mild drooping leaf phenotype. Plant Cell Physiol. 2008, 49, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, Y.; Toriba, T.; Nakamura, H.; Ichikawa, H.; Hirano, H.Y. Temporal and spatial regulation of DROOPING LEAF gene expression that promotes midrib formation in rice. Plant J. 2011, 65, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, W.; Toriba, T.; Ohmori, Y.; Yoshida, A.; Kawai, A.; Mayama-Tsuchida, T.; Ichikawa, H.; Mitsuda, N.; Ohme-Takagi, M.; Hirano, H.Y. The YABBY Gene TONGARI-BOUSHI1 Is Involved in Lateral Organ Development and Maintenance of Meristem Organization in the Rice Spikelet. Plant Cell 2012, 24, 80–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Baum, S.F.; Oh, S.H.; Jiang, C.Z.; Chen, J.C.; Bowman, J.L. Recruitment of CRABS CLAW to promote nectary development within the eudicot clade. Development 2005, 132, 5021–5032. [Google Scholar] [CrossRef] [PubMed]
- Morel, P.; Heijmans, K.; Ament, K.; Chopy, M.; Trehin, C.; Chambrier, P.; Rodrigues Bento, S.; Bimbo, A.; Vandenbussche, M. The Floral C-Lineage Genes Trigger Nectary Development in Petunia and Arabidopsis. Plant Cell 2018, 30, 2020–2037. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Smyth, D.R. CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 1999, 126, 2387–2396. [Google Scholar] [PubMed]
- Brown, W.H. The bearing of nectaries on the phylogeny of flowering plants. Proc. Am. Phil. Soc. 1938, 79, 549–595. [Google Scholar]
- Endress, P.K. The Flowers in Extant Basal Angiosperms and Inferences on Ancestral Flowers. Int. J. Plant Sci. 2001, 162, 1111–1140. [Google Scholar] [CrossRef]
- Fahn, A. The topography of the nectary in the flower and its phylogenetic trend. Phytomorphology 1953, 3, 424–426. [Google Scholar]
- Kozomara, B.; Vinterhalter, B.; Radojevic, L.; Vinterhalter, D. In vitro propagation of Chimonanthus praecox (L.), a winter flowering ornamental shrub. In Vitro Cell Dev. Biol. Plant 2008, 44, 142–147. [Google Scholar] [CrossRef]
- Xiang, L.; Zhao, K.; Chen, L. Molecular cloning and expression of Chimonanthus praecox farnesyl pyrophosphate synthase gene and its possible involvement in the biosynthesis of floral volatile sesquiterpenoids. Plant Physiol. Biochem. 2010, 48, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.G.; Zhou, M.Q.; Chen, L.Q.; Zhang, D.L.; Robert, G.W. Genetic diversity and discrimination of Chimonanthus praecox (L.) link germplasm using ISSR and RAPD markers. Hortscience 2007, 42, 1144–1148. [Google Scholar]
- Vainstein, A.; Lewinsohn, E.; Pichersky, E.; Weiss, D. Floral Fragrance. New Inroads into an Old Commodity. Plant Physiol. 2001, 127, 1383–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kram, B.W.; Carter, C.J. Arabidopsis thaliana as a model for functional nectary analysis. Sex. Plant Reprod. 2009, 22, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, H.F.; Peng, C.C.; Chen, Z.Z.; Long, Z.F. Cloning of SAMT gene cDNA from Chimonanthus praecox and its expression in Escherichia coli. Agric. Sci. Technol. 2012, 13, 82–87. [Google Scholar]
- Liu, D.; Sui, S.; Ma, J.; Li, Z.; Guo, Y.; Luo, D.; Yang, J.; Li, M. Transcriptomic analysis of flower development in wintersweet (Chimonanthus praecox). PLoS ONE 2014, 9, e86976. [Google Scholar] [CrossRef] [PubMed]
- Xue, B.; Shao, Y.Y.; Saunders, R.M.; Tan, Y.H. Alphonsea glandulosa (Annonaceae), a New Species from Yunnan, China. PLoS ONE 2017, 12, e0170107. [Google Scholar] [CrossRef] [PubMed]
- Nepi, M. Nectary structure and ultrastructure. In Nectaries and Nectar; Nicolson, S.W., Nepi, M., Pacini, E., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 129–166. [Google Scholar]
- Elias, T.S. Extrafloral nectaries: Their structure and distribution. In The Biology of Nectaries; Bentley, B., Elias, T.S., Eds.; Columbia University Press: New York, NY, USA, 1983; pp. 174–203. [Google Scholar]
- Bremer, B.; Bremer, K.; Chase, M.W.; Fay, M.F.; Reveal, J.L.; Soltis, D.E.; Soltis, P.S.; Stevens, P.F.; Anderberg, A.A.; Moore, M.J.; et al. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [Green Version]
- Deng, C.; Song, G.; Hu, Y. Rapid determination of volatile compounds emitted from Chimonanthus praecox flowers by HS-SPME-GC-MS. Z. Naturforsch. C 2004, 59, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.R.; Taylor, G.W.; Eskew, M.H. The volatile oil of calycanthus floridus. J. Am. Chem. Soc. 1914, 36, 2182–2185. [Google Scholar] [CrossRef]
- Azuma, H.T.M.; Asakawa, Y. Floral Scent Chemistry and Stamen Movement of Chimonanthus praecox (L.) Link (Calycanthaceae). Acta Phytotaxon. Geobot. 2005, 56, 197–201. [Google Scholar]
- Juarez, M.T.; Twigg, R.W.; Timmermans, M.C. Specification of adaxial cell fate during maize leaf development. Development 2004, 131, 4533–4544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Yeung, K.Y.; Fraley, C.; Murua, A.; Raftery, A.E.; Ruzzo, W.L. Model-based clustering and data transformations for gene expression data. Bioinformatics 2001, 17, 977–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer Sequence | Reverse Primer Sequence |
|---|---|---|
| Actin | AGGCTAAGATTCAAGACAAGG | TTGGTCGCAGCTGATTGCTG |
| CpFIL | AATCCCGACATAACCCACAGAGAG | TCCTGTTGGCGCACGCTAGTT |
| CpCRC | CCTCCCGTCACCTTACAAACTACAG | CTGCTACAAGGAACACTGACCGC |
| CpYABBY2 | CCATTGTCAAGATAAAGGTAGCGATT | CTGGTGGTGGTATAGGTAGCATTCG |
| CpYABBY5-1 | TCTCCCTCTCTATTTATCCTCGTTT | GTAAAAGGCTAAAGCAGGATCATG |
| CpYABBY5-2 | TTTTGAACACTGGAAACTTCGTCTT | GATGCAGCTCGACATCTCACTATCT |
| SAMT | ACCATTTTCACATCATTGCCAGAC | CTTCCTCTTTTACCATCAAGTGCTG |
| DMAPP | ATCGGAGAAGAAAGTGAGCGAGAGT | GCCGTGTATCGAAGCAGCAGT |
| IPP | CAGACCATCTCTTTCTCCCACTTTC | GGTCGGAGAGAAGGTGGTAGAGGTA |
| GPPS1 | GTTAGCCAACTTTCCATACCATTTC | GAGTGACAACATCATCAAAGAAGGG |
| GPPS2 | ATGAAGATGATTAGATTTCGAGTCCAAG | ATAACCAATTTACAACCCCTGACCC |
| GPP | TCTACAGAAAATGGGAGAAAACGAT | TATCTGTTTCTGTCACCAAATCCAC |
| LIS | GGCCAAAGTTAATGAAGTGAGATCC | CGTATATGCCATCGTTGCTGCC |
| MYR1 | TTTCACAAAAATTGCCTTCAACCTT | CAAGGTGATGGAGAACTAAAACAAAAC |
| FPPS | TCTTTGTCCAGTTCTTCCAGCGTT | ATCAGTGAAATCAAAGGCGGAATCT |
| TER3 | AGAGTTGAATTGCACAGGGTGATAG | GCAGTGGATGTTGTTGATCAGCTC |
| TER5 | CTCTCCCTCAGTCTCTTCTCCCTTT | ATCTCCATGCAACATTGGCTACAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Jiang, Y.; Liu, D.; Ma, J.; Li, J.; Li, M.; Sui, S. Floral Scent Emission from Nectaries in the Adaxial Side of the Innermost and Middle Petals in Chimonanthus praecox. Int. J. Mol. Sci. 2018, 19, 3278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103278
Li Z, Jiang Y, Liu D, Ma J, Li J, Li M, Sui S. Floral Scent Emission from Nectaries in the Adaxial Side of the Innermost and Middle Petals in Chimonanthus praecox. International Journal of Molecular Sciences. 2018; 19(10):3278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103278
Chicago/Turabian StyleLi, Zhineng, Yingjie Jiang, Daofeng Liu, Jing Ma, Jing Li, Mingyang Li, and Shunzhao Sui. 2018. "Floral Scent Emission from Nectaries in the Adaxial Side of the Innermost and Middle Petals in Chimonanthus praecox" International Journal of Molecular Sciences 19, no. 10: 3278. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103278