Comparative Proteomic Analysis during the Involvement of Nitric Oxide in Hydrogen Gas-Improved Postharvest Freshness in Cut Lilies



Abstract
:1. Introduction
2. Results
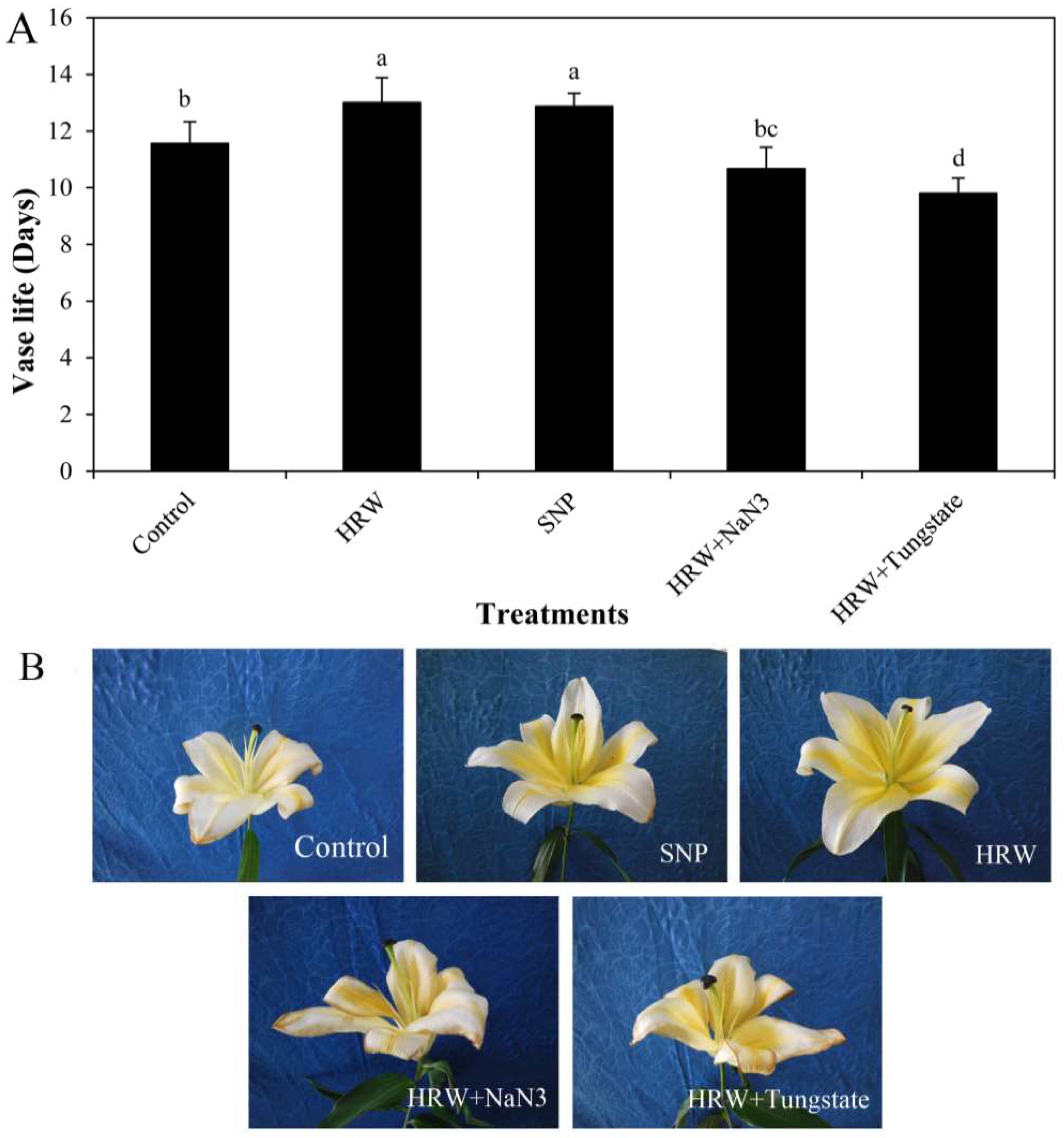
2.1. Effects of HRW, Sodium Nitroprusside (SNP), and NO Inhibitors on Vase Life
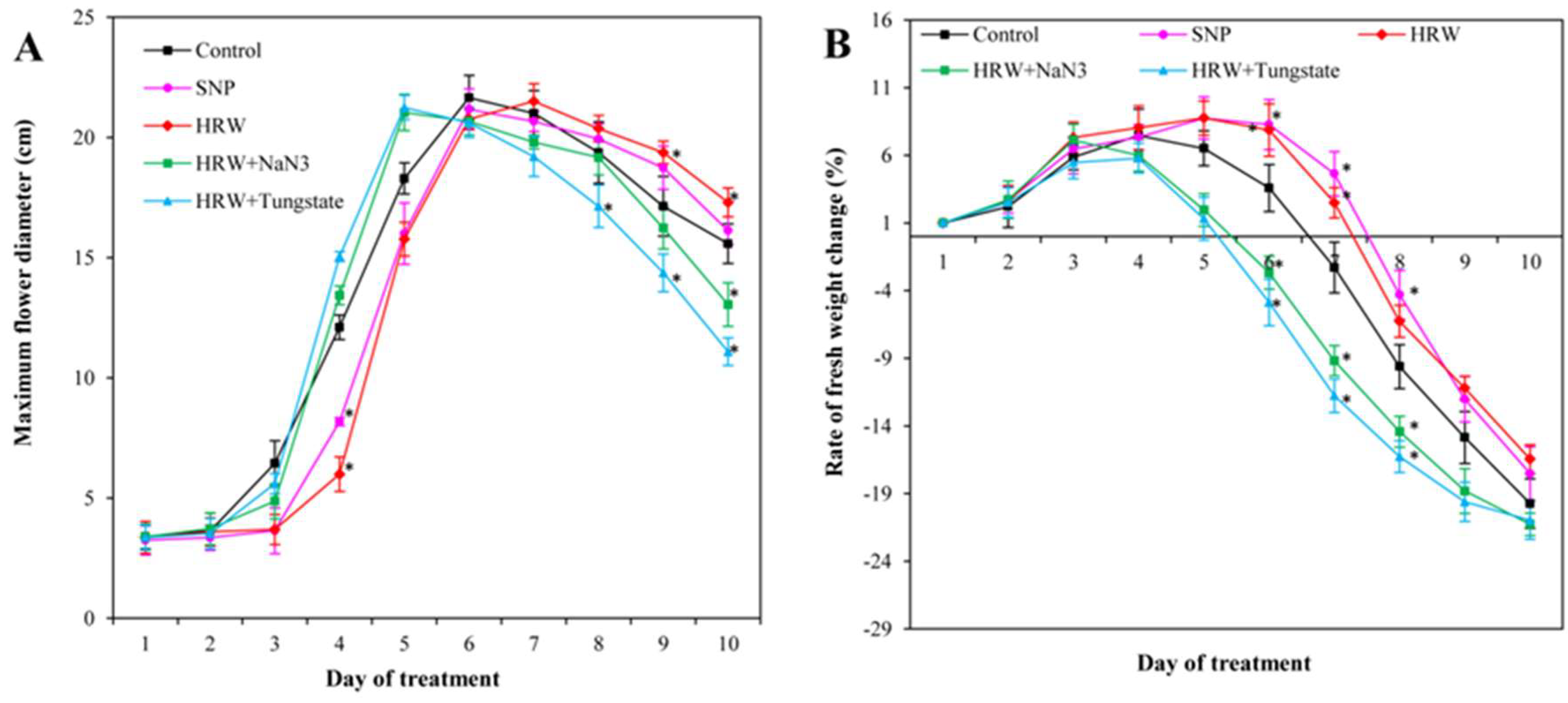
2.2. Effects of HRW, SNP, and NO Inhibitors on Maximum Flower Diameter and Rate of Fresh Weight Change
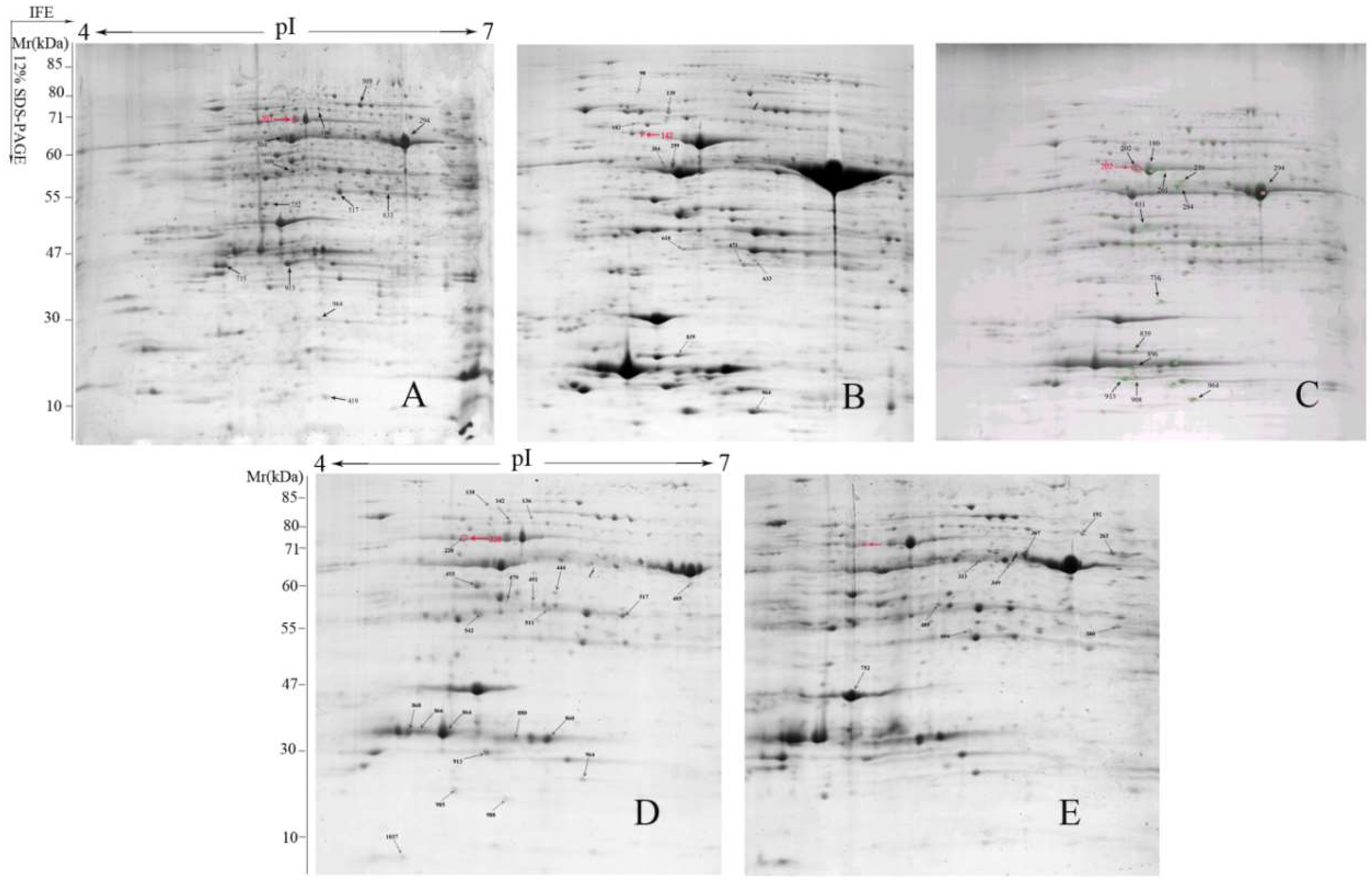
2.3. Two-Dimensional Electrophoresis Analysis and Identification of Proteins
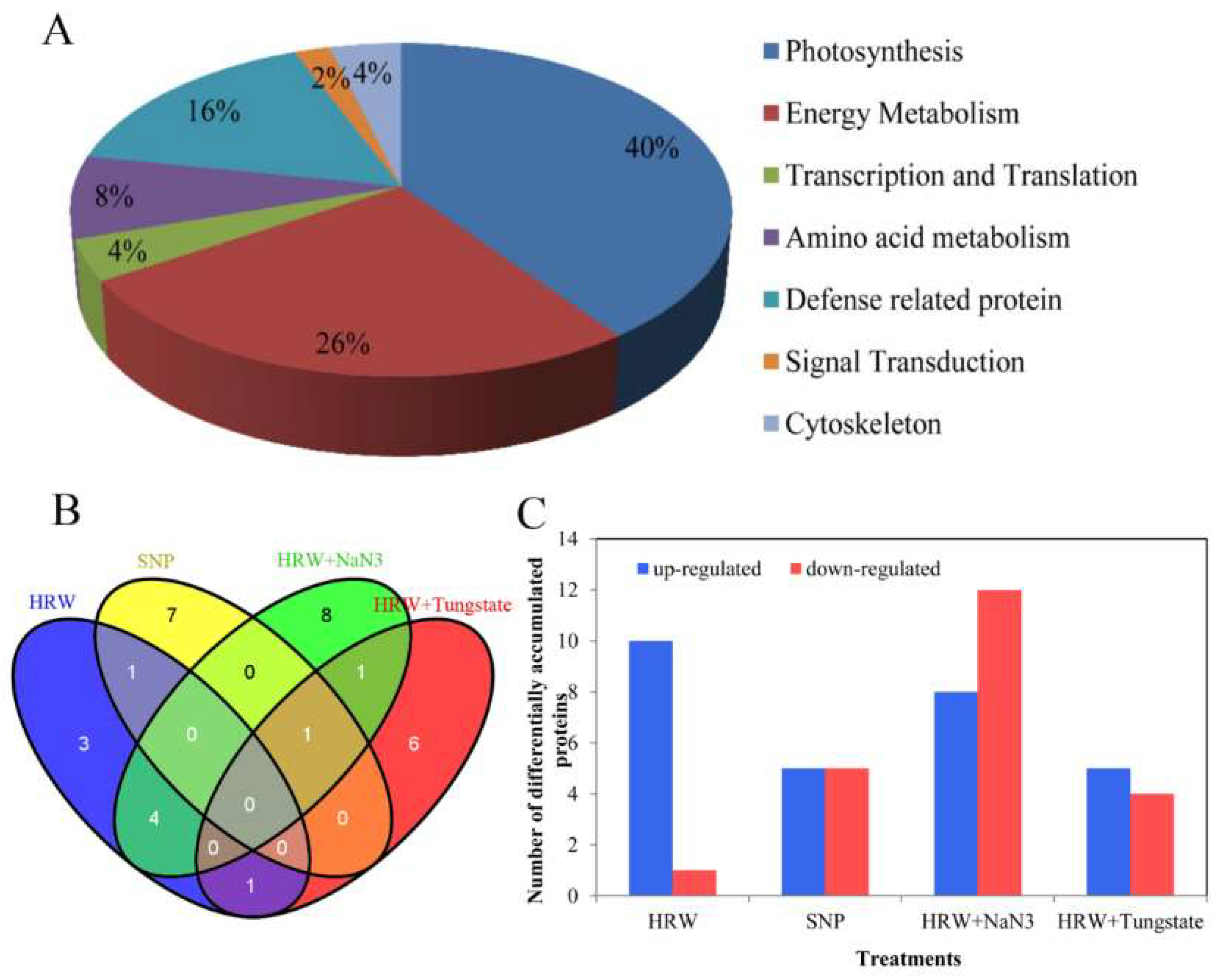
2.4. Functional Classification and Analysis of Differentially Accumulated Proteins
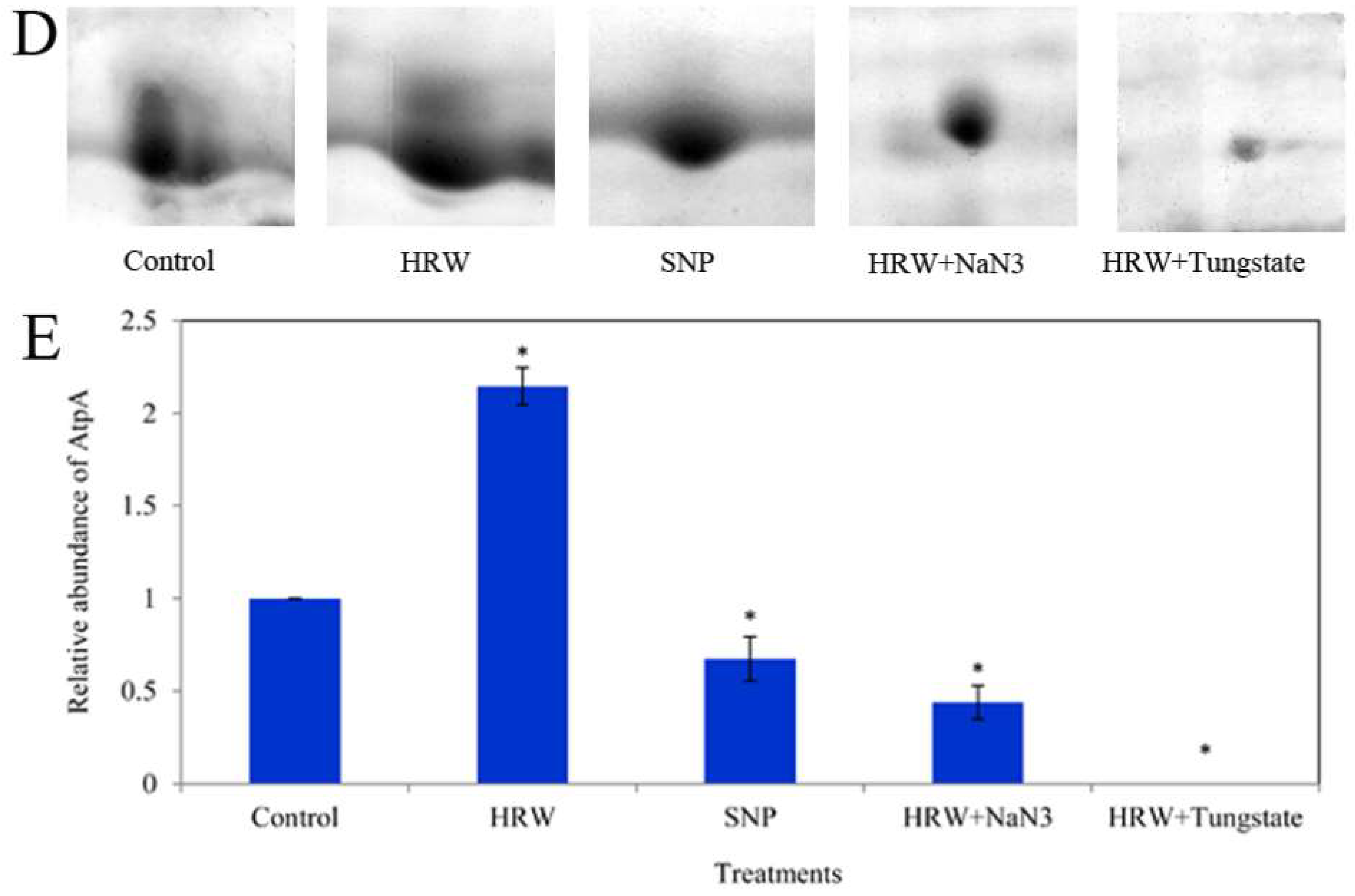
2.5. Relative Expression of LlatpA Gene and the Activity of ATP Synthase (ATPase)
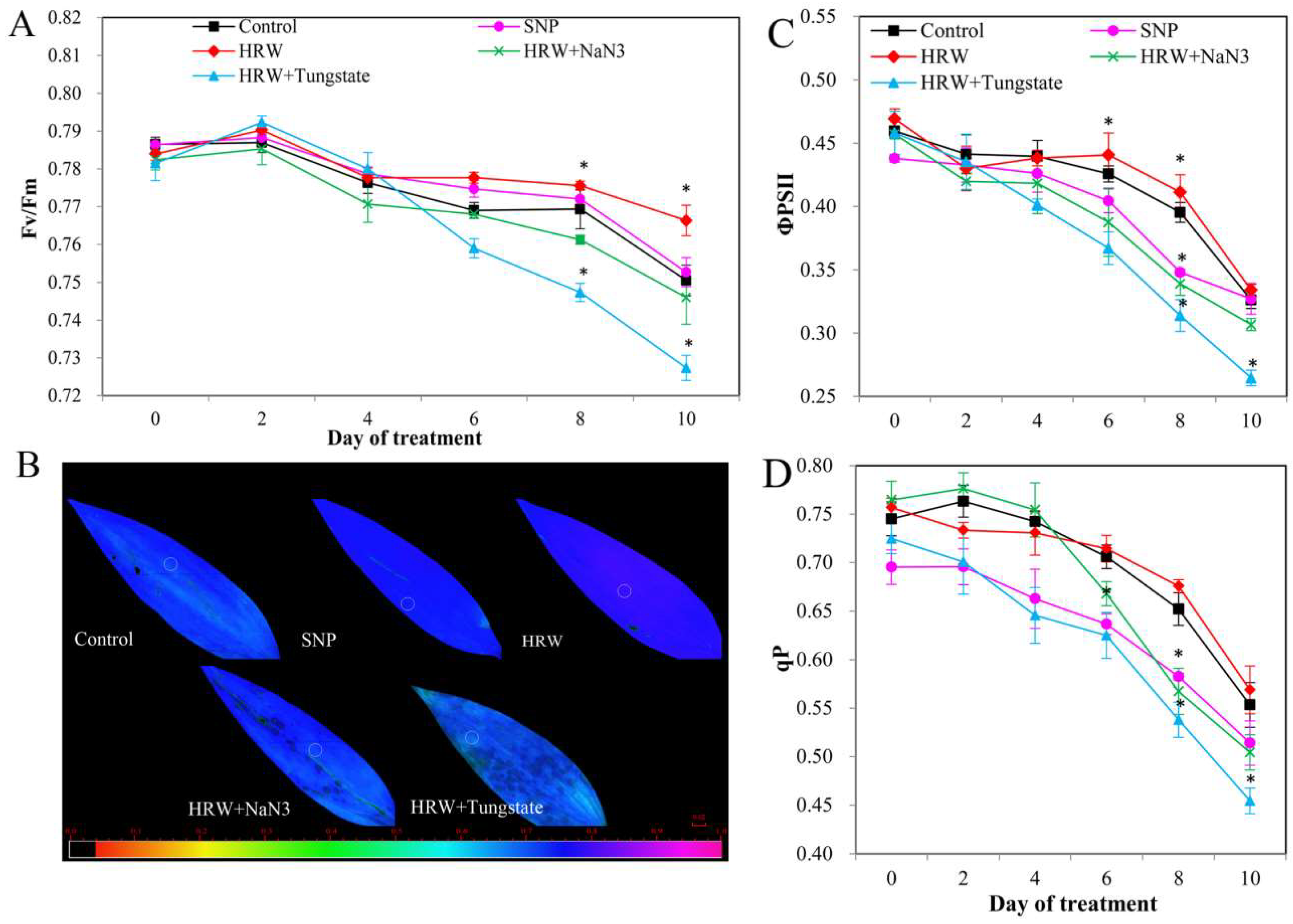
2.6. Chlorophyll Fluorescence and Photosynthetic Parameters
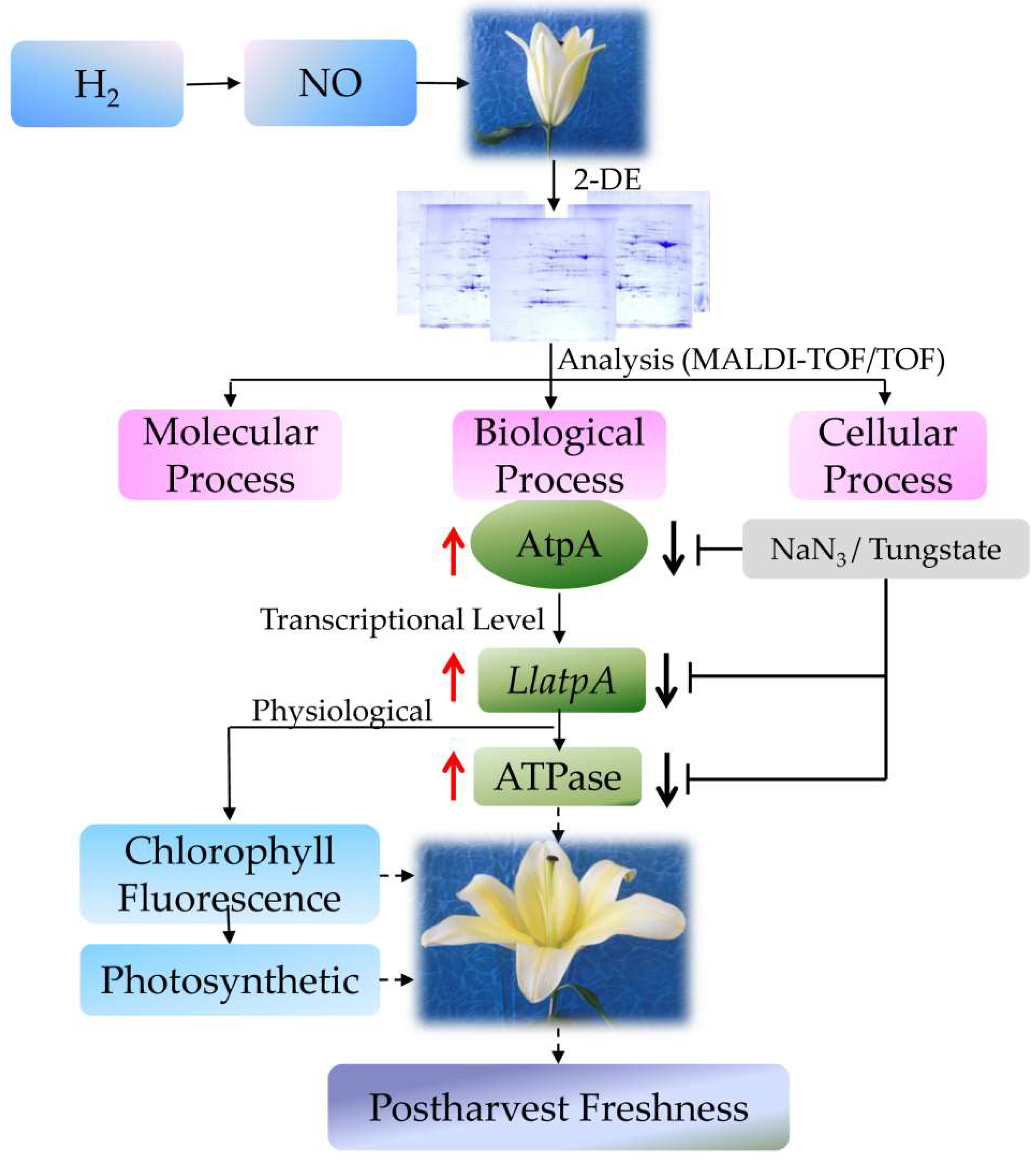
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Determination of Vase Life and Maximum Flower Diameter
4.3. Determination of the Rate of Fresh Weight Change
4.4. Protein Extraction
4.5. Two-Dimensional Electrophoresis (2-DE) and Gel Image Analysis
4.6. Protein Identification and Database Searching
4.7. Quantitative RT-PCR (qRT-PCR) Analyses
4.8. Determination of ATP Synthase Activity
4.9. Determination of Chlorophyll Fluorescence and Photosynthetic Parameters
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AtpA | ATP synthase CF1 alpha subunit (chloroplast) |
| Ci | intercellular CO2 concentration |
| DTT | DL-dithiothreitol |
| Fv/Fm | the maximum quantum yield of PSII |
| Gs | stomatal conductance |
| HRW | hydrogen-rich water |
| Pn | net photosynthetic rate |
| qP | photochemical quenching |
| SNP | sodium nitroprusside |
| TCA | trichloroacetic acid |
| Tr | transpiration rate |
| 2-DE | two-dimensional electrophoresis |
| β-ME | β-mercaptoethanol |
| ФPSII | the effective quantum yield of PSII |
References
- Li, C.X.; Gong, T.Y.; Bian, B.T.; Liao, W.B. Roles of hydrogen gas in plants: A review. Funct. Plant Biol. 2018, 45, 783–792. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Chen, Q.H.; Wang, Y.M.; Shen, Z.G.; Shen, W.B.; Xu, X.M. Hydrogen-rich water induces aluminum tolerance in maize seedlings by enhancing antioxidant capacities and nutrient homeostasis. Ecotoxicol. Environ. Saf. 2017, 144, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.T.; Fang, P.; Zhu, K.K.; Mao, Y.; Gao, C.Y.; Xie, Y.J.; Wang, J.; Shen, W.B. Hydrogen-rich water confers plant tolerance to mercury toxicity in alfalfa seedlings. Ecotoxicol. Environ. Saf. 2014, 105, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Su, N.N.; Wu, Q.; Liu, Y.Y.; Cai, J.T.; Shen, W.B.; Xia, K.; Cui, J. Hydrogen-rich water reestablishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation. J. Agr. Food Chem. 2014, 62, 6454–6462. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.Y.; Duan, X.L.; Yao, P.; Cui, W.T.; Cheng, D.; Zhang, J.; Jin, Q.J.; Chen, J.; Dai, T.S.; et al. Hydrogen gas is involved in auxin-induced lateral root formation by modulating nitric oxide synthesis. Int. J. Mol. Sci. 2017, 18, 2084. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Zhang, W.; Qi, F.; Cui, W.T.; Xie, Y.J.; Shen, W.B. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.L.; Li, P.X.; Wang, Y.N.; Gu, R.X. Hydrogen-rich water delays postharvest ripening and senescence of kiwifruit. Food Chem. 2014, 156, 100. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.J.; Jin, X.; Liao, W.B.; Wang, M.; Niu, L.J.; Li, X.P.; Zhu, Y.C. Effect of hydrogen-rich water on vase life and quality in cut lily and rose flowers. Hortic. Environ. Biotechnol. 2017, 58, 576–584. [Google Scholar] [CrossRef]
- Manjunatha1, G.; Lokesh, V.; Neelwarne, B.; Singh, Z.; Gupta, K.J. Nitric oxide applications for quality enhancement of horticulture produce. Jules Janick. Hortic. Rev. 2014, 42, 121–156. [Google Scholar] [CrossRef]
- Shabaniana, S.; Esfahania, M.N.; Karamian, R.; Tran, L.S.P. Physiological and biochemical modifications by postharvest treatment with sodium nitroprusside extend vase life of cut flowers of two gerbera cultivars. Postharvest Biol. Technol. 2018, 137, 1–8. [Google Scholar] [CrossRef]
- Dwivedic, S.K.; Aroraa, A.; Singha, V.P.; Sairama, R.; Bhattacharya, R.C. Effect of sodium nitroprusside on differential activity of antioxidants and expression of SAGs in relation to vase life of gladiolus cut flowers. Sci. Hortic. 2016, 210, 158–165. [Google Scholar] [CrossRef]
- Zeng, C.L.; Liu, L.; Xu, G.Q. The physiological responses of carnation cut flowers to exogenous nitric oxide. Sci. Hortic. 2011, 127, 424–430. [Google Scholar] [CrossRef]
- Liao, W.B.; Zhang, M.L.; Yu, J.H. Role of nitric oxide in delaying senescence of cut rose flowers and its interaction with ethylene. Sci. Hortic. 2013, 155, 30–38. [Google Scholar] [CrossRef]
- Xie, Y.J.; Mao, Y.; Zhang, W.; Lai, D.W.; Wang, Q.Y.; Shen, W.B. Reactive oxygen species-dependent nitric oxide production contributes to hydrogen-promoted stomatal closure in Arabidopsis. Plant Physiol. 2014, 165, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Liao, W.B.; Niu, L.J.; Wang, M.; Ma, Z.J. Nitric oxide is involved in hydrogen gas-induced cell cycle activation during adventitious root formation in cucumber. BMC Plant Biol. 2016, 16, 146. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Cui, W.T.; Zhu, K.K.; Xie, Y.J.; Zhang, C.H.; Shen, W.B. Hydrogen-rich water alleviates aluminum-induced inhibition of root elongation in alfalfa via decreasing nitric oxide production. J. Hazard. Mater. 2014, 267, 40–47. [Google Scholar] [CrossRef]
- Baudouin, E.; Hancock, J.T. Nitric oxide signaling in plants. Front. Plant Sci. 2014, 4, 553. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Liao, W.B. The metabolic constituent and rooting-related enzymes responses of marigold explants to hydrogen gas during adventitious root development. Theor. Exp. Plant Physiol. 2017, 29, 1–9. [Google Scholar] [CrossRef]
- Hu, H.L.; Zhao, S.P.; Li, P.X.; Shen, W.B. Hydrogen gas prolongs the shelf life of kiwifruit by decreasing ethylene biosynthesis. Postharvest Biol. Technol. 2018, 135, 123–130. [Google Scholar] [CrossRef]
- Zhu, Y.C.; Liao, W.B.; Wang, M.; Niu, L.J.; Xu, Q.Q.; Jin, X. Nitric oxide is required for hydrogen gas-induced adventitious root formation in cucumber. J. Plant Physiol. 2016, 195, 50. [Google Scholar] [CrossRef]
- Liu, R.X.; Chen, S.M.; Jiang, J.F.; Zhu, L.; Zheng, C.; Han, S.; Gu, J.; Sun, j.; Wang, H.B.; Song, A.P.; Chen, F.D. Proteomic changes in the base of chrysanthemum cuttings during adventitious root formation. BMC Genom. 2013, 14, 919. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Fan, L.; Hughes, T.; Palmer Campbell, L.; Li, L.; Li, X.H. Quantitative proteomic investigation on the effect of 1-methylcyclopropene treatments on postharvest quality of selected cut flowers. Acta Hortic. 2015, 1104, 311–318. [Google Scholar] [CrossRef]
- Dai, C.; Cui, W.T.; Pan, J.C.; Xie, Y.J.; Wang, J.; Shen, W.B. Proteomic analysis provides insights into the molecular bases of hydrogen gas-induced cadmium resistance in Medicago sativa. J. Proteom. 2017, 152, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.Y.; Zhang, L.; Jiang, L.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Effect of postharvest nitric oxide treatment on the proteome of peach fruit during ripening. Postharvest Biol. Technol. 2016, 112, 277–289. [Google Scholar] [CrossRef]
- Li, L.; Song, J.; Kalt, W.; Forney, C.; Tsao, R.; Pinto, D.; Pinto, D.; Chisholm, K.; Campbell, L.; Fillmore, S.; et al. Quantitative proteomic investigation employing stable isotope labeling by peptide dimethylation on proteins of strawberry fruit at different ripening stages. J. Proteom. 2013, 94, 219–239. [Google Scholar] [CrossRef] [PubMed]
- Li, T.T.; Zhu, H.; Wu, Q.X.; Yang, C.W.; Duan, X.W.; Qu, H.X.; Yun, Z.; Jiang, Y.M. Comparative proteomic approaches to analysis of litchi pulp senescence after harvest. Food Res. Int. 2015, 78, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Hisabori, T.; Konno, H.; Ichimura, H.; Strotmann, H.; Bald, D. Molecular devices of chloroplast F1-ATP synthase for the regulation. BBA Bioenerg. 2002, 1555, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.X.; Pan, D.Z.; Li, J.; Tan, F.L.; Hoffmann-Benning, S.; Liang, W.Y.; Chen, W. Proteomic analysis of changes in the Kandelia candel chloroplast proteins reveals pathways associated with salt tolerance. Plant Sci. 2015, 231, 159. [Google Scholar] [CrossRef]
- Lee, C.; Rhee, J.K.; Kim, D.G.; Choi, Y.E. Proteomic study reveals photosynthesis as downstreams of both MAP kinase and cAMP signaling pathways in Chlamydomonas reinhardtii. Photosynthetica 2015, 53, 625–629. [Google Scholar] [CrossRef]
- Wolfgang, J.; Nathan, N. ATP Synthase. Annu. Rev. Biochem. 2015, 84, 631–657. [Google Scholar] [CrossRef]
- Shao, H.; Cao, Q.; Tao, X.; Gu, Y.; Chang, M.; Huang, C.; Zhang, Y.; Feng, H. Cloning and characterization of ATP synthase CF1 α gene from sweet potato. Afr. J. Biotechnol. 2011, 10, 19035–19042. [Google Scholar] [CrossRef]
- Shu, S.; Yuan, Y.H.; Chen, J.; Sun, J.; Zhang, W.H.; Tang, Y.Y.; Zhong, M.; Guo, S. The role of putrescine in the regulation of proteins and fatty acids of thylakoid membranes under salt stress. Sci. Rep. 2015, 5, 14390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viale, A.; Vallejos, R.; Jagendorf, A.T. Hydrogen exchange into soluble spinach chloroplast coupling factor during heat activation of its ATPase. BBA–Bioenerg. 1981, 637, 496–503. [Google Scholar] [CrossRef]
- He, W.X.; Xu, Y.; Tang, L.; Wei, Q.; Li, J.; Chen, F. Molecular cloning regulation of chilling--repressed gene atpA in Elumus sibiricus. Prog. Biochem. Biophys. 2005, 32, 67–74. [Google Scholar]
- Zandonadi, D.B.; Santos, M.P.; Dobbss, L.B.; Olivares, F.L.; Canellas, L.P.; Binzel, M.L.; Okorokova-Façanha, A.L.; Façanha, A.R. Nitric oxide mediates humic acids-induced root development and plasma membrane H+-ATPase activation. Planta 2010, 231, 1025–1036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Han, X.X.; Chen, X.L.; Jin, H.; Cui, X.M. Exogenous nitric oxide on antioxidative system and ATPase activities from tomato seedlings under copper stress. Sci. Hortic. 2009, 123, 217–223. [Google Scholar] [CrossRef]
- Okumura, M.; Inoue, S.I.; Kuwata, K.; Kinoshita, T. Photosynthesis activates plasma membrane H+-ATPase via sugar accumulation in Arabidopsis leaves. Plant Physiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.N.; Zhao, X.Q.; Wang, Z.Q.; Shen, W.B.; Xu, X.M. Protective effects of hydrogen-rich water on the photosynthetic apparatus of maize seedlings (Zea mays L.) as a result of an increase in antioxidant enzyme activities under high light stress. Plant Growth Regul. 2015, 77, 43–56. [Google Scholar] [CrossRef]
- Cui, J.X.; Zhou, Y.H.; Ding, J.G.; Xia, X.J.; Shi, K.; Chen, S.C.; Asami, T.; Yu, J.Q. Role of nitric oxide in hydrogen peroxide-dependent induction of abiotic stress tolerance by brassinosteroids in cucumber. Plant Cell Environ. 2011, 34, 347. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.P.; Srivastava, P.K.; Prasad, S.M. Nitric oxide alleviates arsenic-induced toxic effects in ridged Luffa seedlings. Plant Physiol. Biochem. 2013, 71, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Wodala, B.; Deák, Z.; Vass, I.; Erdei, L. Nitric oxide modifies photosynthetic electron transport in pea leaves. Acta Biol. Szegediensis 2005, 49, 7–8. [Google Scholar]
- Dong, N.G.; Li, Y.F.; Qi, J.X.; Chen, Y.H.; Hao, Y.B. Nitric oxide synthase-dependent nitric oxide production enhances chilling tolerance of walnut shoots in vitro via involvement chlorophyll fluorescence and other physiological parameter levels. Sci. Hortic. 2018, 230, 68–77. [Google Scholar] [CrossRef]
- Spetea, C.; Schoefs Spetea, B. Solute transporters in plant thylakoid membranes: Key players during photosynthesis and light stress. Commun. Integr. Biol. 2010, 3, 122–129. [Google Scholar] [CrossRef]
- Wang, T.; Yang, W.H.; Xie, Y.F.; Shi, D.W.; Ma, Y.L.; Sun, X. Effects of exogenous nitric oxide on the photosynthetic characteristics of bamboo (Indocalamus barbatus McClure) seedlings under acid rain stress. Plant Growth Regul. 2017, 82, 69–78. [Google Scholar] [CrossRef]
- Chen, P.B.; Li, X.; Huo, K.; Wei, X.D.; Dai, C.C.; Lv, C.G. Promotion of photosynthesis in transgenic rice over-expressing of maize C4 phosphoenolpyruvate carboxylase gene by nitric oxide donors. J. Plant Physiol. 2014, 171, 458–466. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Gao, J.D.; Chen, Z.J.; Qiao, X.Y.; Huang, H.L.; Cui, B.Y.; Zhu, Q.F.; Dai, Z.; Wu, H.; Pan, Y.; Yang, C. Comparative proteomics reveals the physiological differences between winter tender shoots and spring tender shoots of a novel tea (Camellia sinensis L.) cultivar evergrowing in winter. BMC Plant Biol. 2017, 17, 206. [Google Scholar] [CrossRef] [PubMed]
- Sheffield, J.; Taylor, N.; Fauquet, C.; Chen, S. The cassava (Manihot esculenta Crantz) root proteome: Protein identification and differential expression. Proteomics 2006, 6, 1588–1598. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmitgen, T.D. Analysis of relative gene expression data using real-time quantification PCR and the 2–∆∆Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Protein Name | Species | Accession No. | Expressed MW (Da)/pI | Theoretical MW (Da)/pI | Peptide Count | Score | Protein Score Confidence level (C.I.%) | Up/Down |
|---|---|---|---|---|---|---|---|---|---|
| 201 | ATP synthase CF1 alpha subunit (chloroplast) | Lilium superbum | YP_009130198.1 | 55,285.2/5.41 | 55,319.38/5.41 | 21 | 962 | 100 | ↑ |
| 259 | ATP synthase alpha subunit, partial (mitochondrion) | Erythronium dens-canis | AFM91753.1 | 28,195.8/6.51 | 28,213.43/6.50 | 10 | 114 | 100 | ↑ |
| 202 | ATP synthase CF1 alpha subunit (chloroplast) | Lilium superbum | YP_009130198.1 | 55,285.2/5.41 | 55,319.38/5.41 | 22 | 936 | 100 | ↑ |
| 908 | Photosystem II oxygen evolving complex protein 2 precursor | Fritillaria agrestis | AAC04809.1 | 28,094.2/8.31 | 28,111.52/8.31 | 5 | 60 | 98.435 | ↓ |
| 284 | ATP synthase CF1 beta subunit, partial (plastid) | Lilium superbum | AEZ48850.1 | 53,576.9/5.22 | 53,610.53/5.22 | 15 | 59 | 98.118 | ↑ |
| 294 | Ribulose-1,5-bisphosphate carboxulase/oxygenase large subunit, partial (chloroplast) | Heloniopsis kawanoi | AIW53238.1 | 50,960.6/6.23 | 50,992.85/6.24 | 29 | 1180 | 100 | ↑ |
| 180 | ATP synthase CF1 alpha subunit (chloroplast) | Lilium superbum | YP_009130198.1 | 55,285.2/5.41 | 5319.38/5.41 | 21 | 865 | 100 | ↑ |
| 896 | Carbonic anhydrase | Musa acuminata subsp. | Tr|M0TL28 | 22,422.1/5.07 | 22,436.69/5.06 | 4 | 104 | 99.985 | ↑ |
| 716 | PDZ domain-containing protein | Cynara cardunculus var. | Tr|A0A118JU51 | 36,198.3/6.18 | 36,220.78/6.18 | 6 | 113 | 99.998 | ↑ |
| 913 | Chlorophyll a-b binding protein, chloroplastic | Musa acuminata subsp. | Tr|M0SBM9 | 29,718.2/8.96 | 29,737.07/8.96 | 4 | 171 | 100 | ↑ |
| 431 | Actin | Lilium davidii var. Davidii | ALO18835.1 | 41,649.0/5.31 | 41,675.77/5.31 | 15 | 447 | 100 | ↑ |
| 511 | Glutamine synthetase | Tulipa pulchella | BAM84282.1 | 38,673.4/5.64 | 38,697.60/5.64 | 5 | 90 | 99.999 | ↑ |
| 479 | Actin | Lilium regale | AFU06383.1 | 41,619.0/5.31 | 41,645.75/5.31 | 16 | 421 | 100 | ↑ |
| 492 | Monodehydroascorbate reductase | Lilium longiflorum | ADF43731.1 | 46,732.5/5.89 | 46,761.56/5.89 | 14 | 86 | 99.996 | ↑ |
| 220 | ATP synthase CF1 alpha subunit (chloroplast) | Lilium superbum | YP_009130198.1 | 55,285.2/5.41 | 55,319.38/5.41 | 19 | 730 | 100 | ↓ |
| 988 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (chloroplast) | Gagea wilczekii | AAM29162.1 | 50,739.4/5.96 | 50,771.69/5.96 | 12 | 494 | 100 | ↑ |
| 1060 | Pathogenesis-related protein 10 | Lilium regale | AHG94651.1 | 16,709.4/5.31 | 16,719.85/5.31 | 7 | 536 | 100 | ↓ |
| 136 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (chloroplast) | Trillium camschatcense | AFP48691.1 | 44,763.6/6.52 | 44,792.01/6.53 | 9 | 63 | 99.14 | ↑ |
| 985 | ATP synthase beta subunit, partial (chloroplast) | Fritillaria acmopetala | AKG96681.1 | 51,914.1/5.13 | 51,946.57/5.13 | 16 | 68 | 99.774 | ↓ |
| 142 | ATP synthase CF1 alpha subunit (chloroplast) | Ripogonum album | ANO45506.1 | 55,341.1/5.26 | 55,375.27/5.26 | 10 | 72 | 99.91 | ↓ |
| 415 | Glutamine synthetase | Erythranthe guttata | Tr|A0A022RZ30 | 39,028.6/5.40 | 39,053.05/5.39 | 7 | 393 | 100 | ↓ |
| 405 | 6-Phosphogluconate dehydrogenase, decarboxylating | Citrus sinensis | Tr|A0A067G3F9 | 53,519.6/6.38 | 53,553.34/6.38 | 13 | 437 | 100 | ↓ |
| 444 | Elongation factor Tu | Vigna angularis var. Angularis | Tr|A0A0S3RGB1 | 52,659.2/6.34 | 52,692.26/6.34 | 14 | 671 | 100 | ↓ |
| 868 | Chlorophyll a-b binding protein, chloroplastic | Kalanchoe fedtschenkoi | Tr|A0A089WZX0 | 28,226.3/5.15 | 28,244.20/5.15 | 7 | 195 | 100 | ↑ |
| 517 | Glutamine synthetase | Lolium perenne | Tr|C5IW59 | 38,973.5/5.40 | 38,998.03/5.40 | 10 | 348 | 100 | ↓ |
| 866 | Chlorophyll a-b binding protein, chloroplastic | Carya cathayensis | Tr|Q1KLZ3 | 28,296.3/5.15 | 28,314.25/5.15 | 5 | 101 | 99.969 | ↑ |
| 880 | Beta carbonic anhydrase 3 | Arabidopsis thaliana | Sp|Q9ZUC2 | 28,810.8/6.54 | 28,829.03/6.54 | 6 | 94 | 99.83 | ↑ |
| 864 | Chlorophyll a-b binding protein, chloroplastic | Kalanchoe fedtschenkoi | Tr|A0A089WZX0 | 28,226.3/5.15 | 28,244.20/5.15 | 10 | 294 | 100 | ↓ |
| 860 | Carbonic anhydrase | Zea mays | Tr|Q41729 | 71,291.9/8.93 | 71,337.55/8.93 | 10 | 101 | 99.969 | ↓ |
| 542 | Ribulose bisphosphate carboxylase/oxygenase activase | Medicago truncatula | Tr|G7JTD2 | 52,135.9/5.42 | 52,169.06/5.42 | 15 | 358 | 100 | ↓ |
| 1037 | Type II peroxiredoxin | Medicago truncatula | Tr|A0A072U4Q3 | 25,893.6/9.35 | 25,909.84/9.35 | 10 | 191 | 100 | ↓ |
| 304 | ATP synthase CF1 beta subunit, partial (plastid) | Lilium superbum | AEZ48850.1 | 53,576.9/5.22 | 53,610.53/5.22 | 28 | 1190 | 100 | ↓ |
| 964 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (chloroplast) | Cardiocrinum giganteum var. Yunnanense | AAM29161.1 | 50,201.1/6.04 | 50,233.05/6.04 | 12 | 88 | 99.998 | ↑ |
| 299 | ATP synthase CF1 beta subunit, partial (plastid) | Lilium superbum | AEZ48850.1 | 53,576.9/5.22 | 53,610.53/5.22 | 21 | 112 | 100 | ↓ |
| 130 | 70 kDa heat shock protein | Sandersonia aurantiaca | AAL85887.1 | 36,768.6/4.83 | 36,791.57/4.82 | 3 | 260 | 100 | ↓ |
| 633 | NADP-dependent alkenal double bond reductase P2 | Morus notabilis | Tr|W9SE47 | 40,693.7/6.23 | 40,719.76/6.22 | 7 | 96 | 99.89 | ↓ |
| 90 | Elongation factor G, mitochondrial | Medicago truncatula | Tr|A0A072UPP0 | 81,881.8/5.50 | 81,933.70/5.50 | 17 | 579 | 100 | ↓ |
| 671 | Cysteine synthase | Populus trichocarpa | Tr|B9HJY5 | 34,176.2/7.64 | 34,197.70/7.64 | 11 | 132 | 100 | ↑ |
| 839 | Putative L-ascorbate peroxidase 2, cytosolic-like | Solanum chacoense | Tr|A0A0V0HVQ3 | 28,638.6/5.75 | 28,656.76/5.75 | 9 | 324 | 100 | ↑ |
| 618 | Trypsin-like serine protease | Medicago truncatula | Tr|G7KIR6 | 45,774.3/6.79 | 45,802.44/6.80 | 10 | 525 | 100 | ↑ |
| 182 | FtsH-like protein Pftf | Nicotiana tabacum | Tr|Q9ZP50 | 74,335.8/6.00 | 74,382.14/6.00 | 24 | 658 | 100 | ↑ |
| 307 | ATP synthase alpha subunit, partial (mitochondrion) | Lilium lancifolium | AAR28047.1 | 41,824.8/6.47 | 41,850.81/6.47 | 13 | 397 | 100 | ↑ |
| 313 | Atpb (chloroplast) | Lilium distichum | AMT85217.1 | 53,546.9/5.22 | 53,580.50/5.22 | 17 | 113 | 100 | ↓ |
| 580 | Glyceraldehyde-3-phosphate dehydrogenase, partial | Lilium regale | AHZ94971.1 | 36,779.2/7.11 | 36,802.04/7.11 | 12 | 255 | 100 | ↑ |
| 349 | Ribulose-1,5-bisphosphate carboxylase/oxygenase large subunit, partial (chloroplast) | Cardiocrinum giganteum var. Yunnanense | AAM29161.1 | 50,201.1/6.04 | 50,233.05/6.04 | 19 | 159 | 100 | ↑ |
| 489 | Monodehydroascorbate reductase | Lilium longiflorum | ADF43731.1 | 46,732.5/5.89 | 46,761.56/5.89 | 18 | 271 | 100 | ↓ |
| 752 | Photosystem II oxygen evolving complex protein 1 precursor | Fritillaria agrestis | AAC04808.1 | 34,847.8/6.26 | 34,869.39/6.25 | 16 | 695 | 100 | ↓ |
| 263 | Dihydrolipoyl dehydrogenase | Salvia miltiorrhiza | Tr|A0A0G2SJN7 | 53,520/6.96 | 53,553.72/6.96 | 6 | 259 | 100 | ↑ |
| 192 | Malic enzyme | Phaseolus angularis | Tr|A0A0L9UG31 | 73,189.2/8.33 | 73,235.49/8.33 | 9 | 177 | 100 | ↓ |
| 604 | Fructose-bisphosphate aldolase | Oxytropis ochrocephala | Tr|A0A0K1JSG5 | 42,894.2/6.39 | 42,920.76/6.39 | 9 | 477 | 100 | ↑ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Ma, Z.; Liao, W. Comparative Proteomic Analysis during the Involvement of Nitric Oxide in Hydrogen Gas-Improved Postharvest Freshness in Cut Lilies. Int. J. Mol. Sci. 2018, 19, 3955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123955
Huo J, Huang D, Zhang J, Fang H, Wang B, Wang C, Ma Z, Liao W. Comparative Proteomic Analysis during the Involvement of Nitric Oxide in Hydrogen Gas-Improved Postharvest Freshness in Cut Lilies. International Journal of Molecular Sciences. 2018; 19(12):3955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123955
Chicago/Turabian StyleHuo, Jianqiang, Dengjing Huang, Jing Zhang, Hua Fang, Bo Wang, Chunlei Wang, Zhanjun Ma, and Weibiao Liao. 2018. "Comparative Proteomic Analysis during the Involvement of Nitric Oxide in Hydrogen Gas-Improved Postharvest Freshness in Cut Lilies" International Journal of Molecular Sciences 19, no. 12: 3955. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19123955