A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells


Abstract
:1. Introduction
2. Results and Discussions
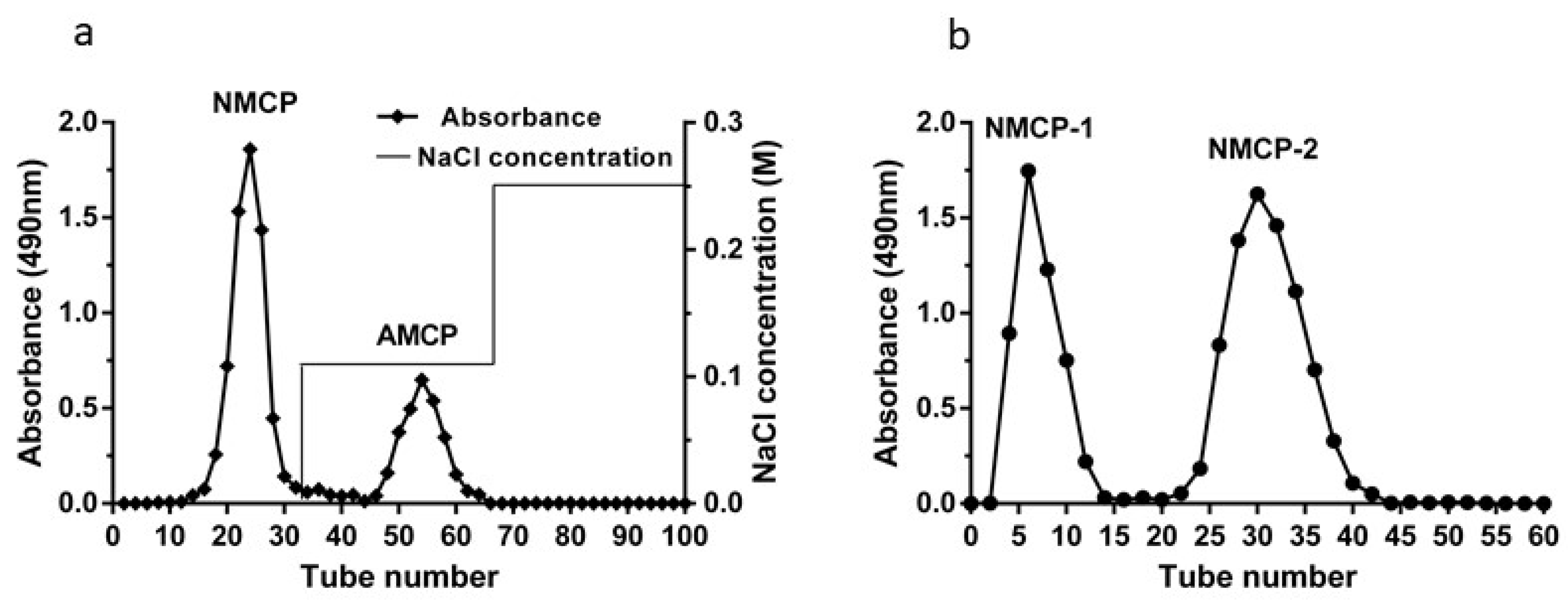
2.1. Purification of Crude MCP
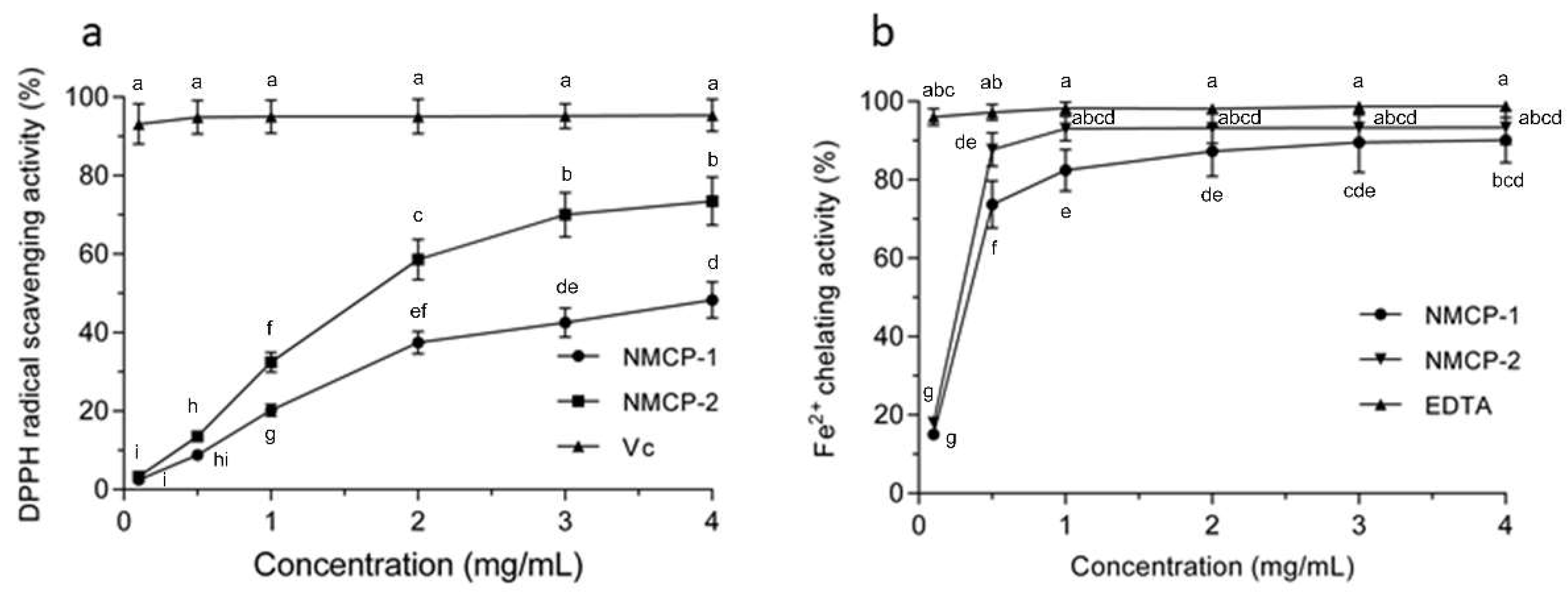
2.2. DPPH (2,2-diphenyl-1-picrylhydrazyl) Scavenging Effect and Ferrous Ion Chelating Ability of NMCP-1 and NMCP-2
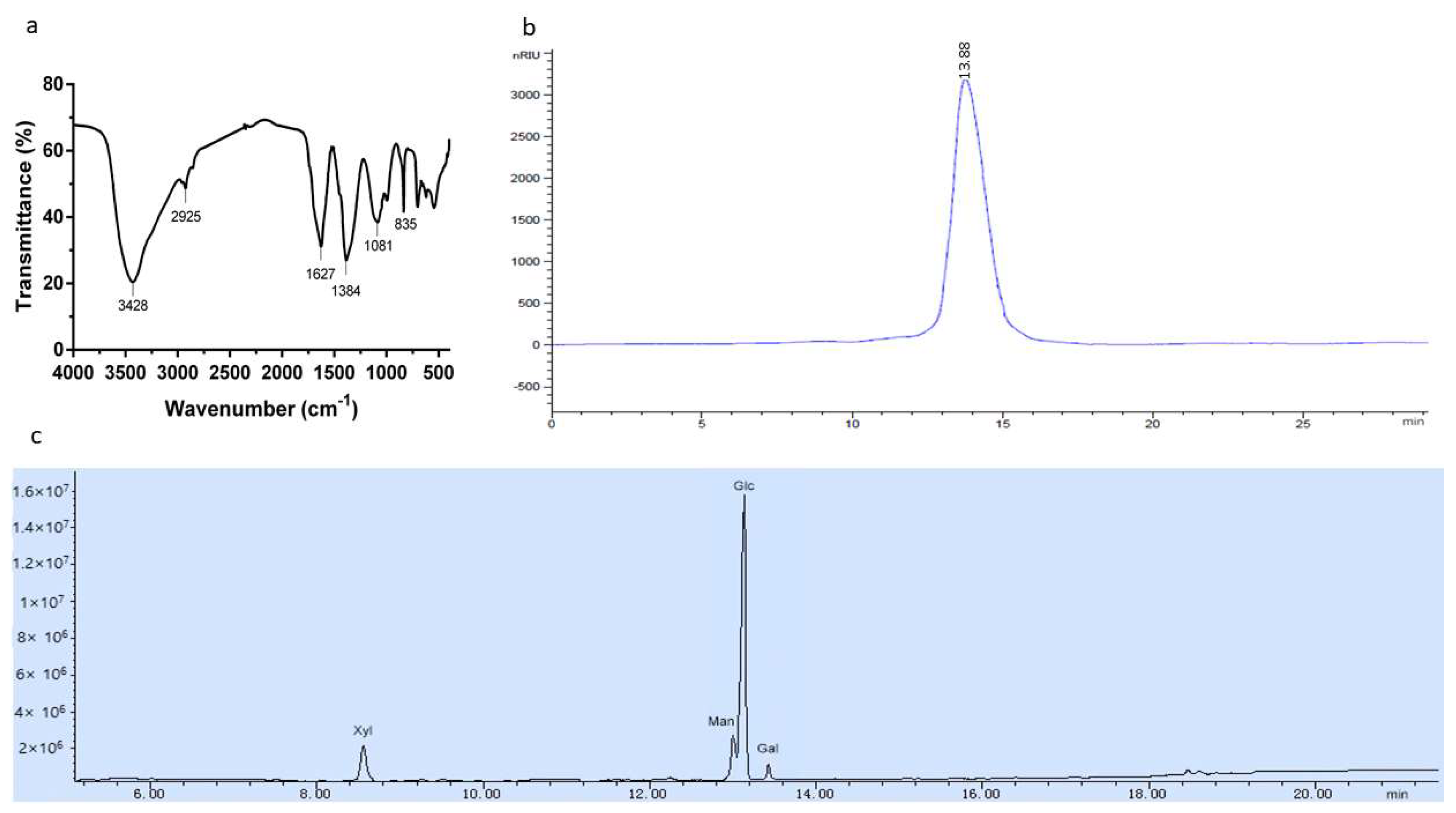
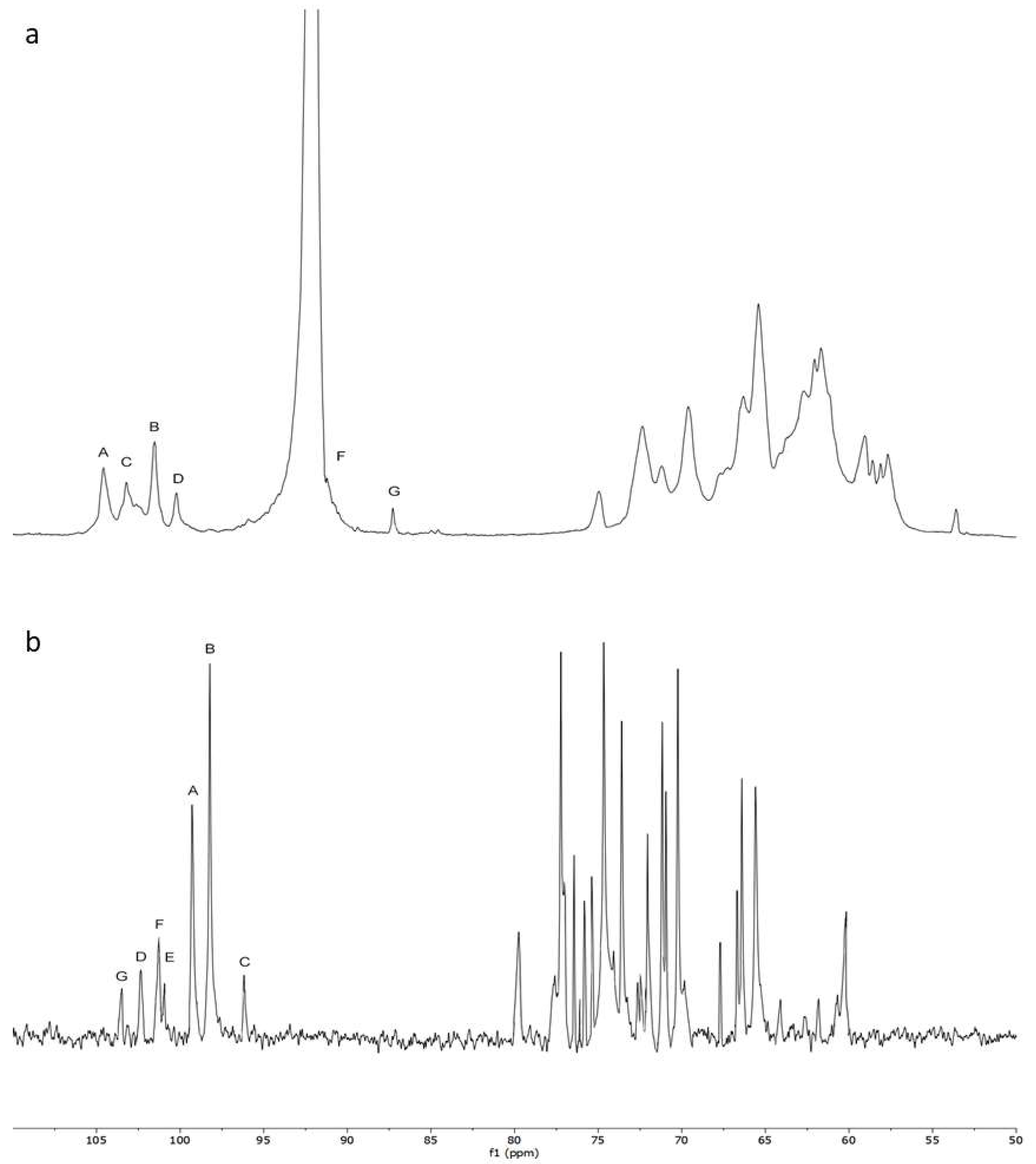
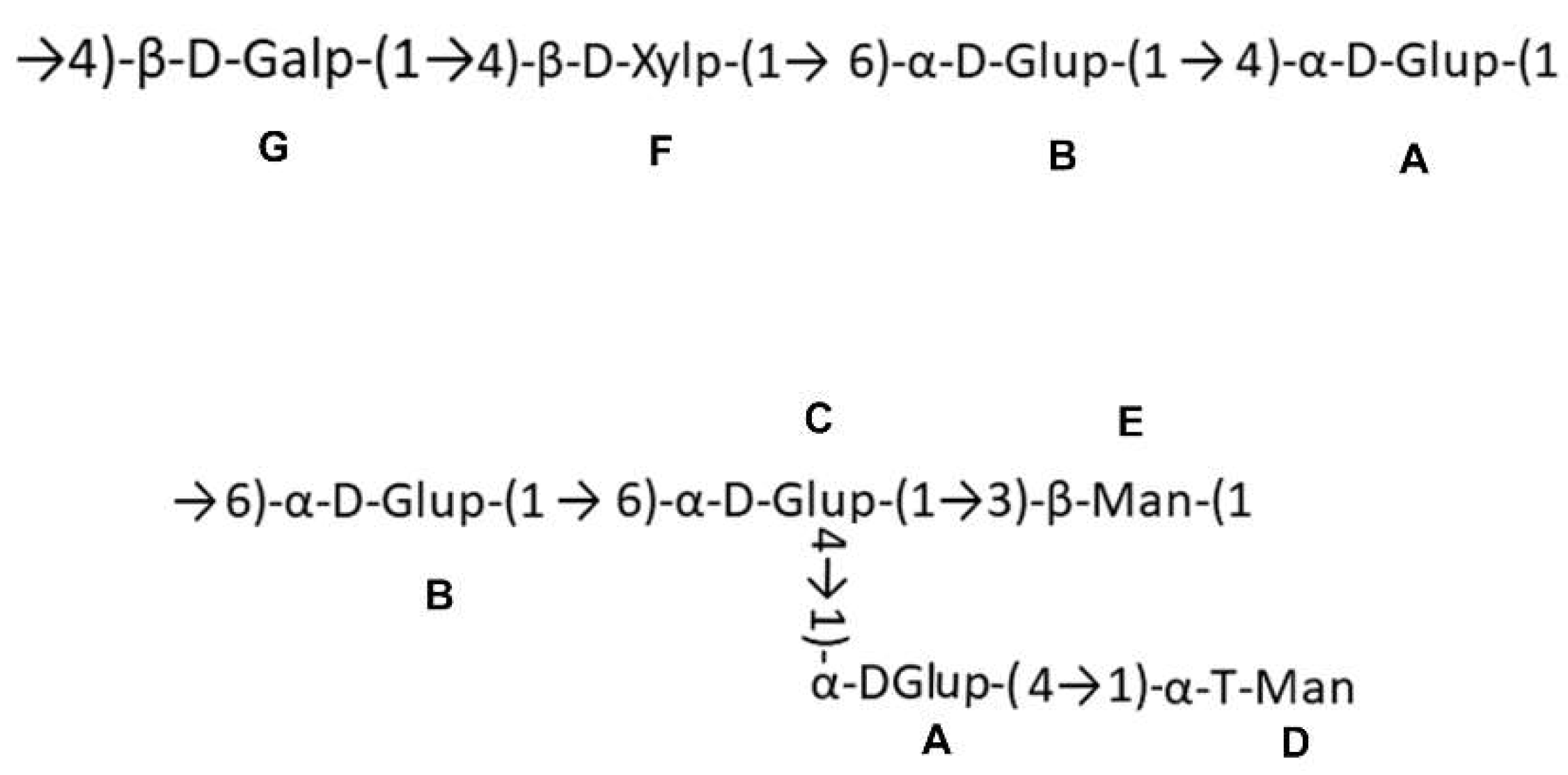
2.3. Chemical Characters of the Polysaccharide
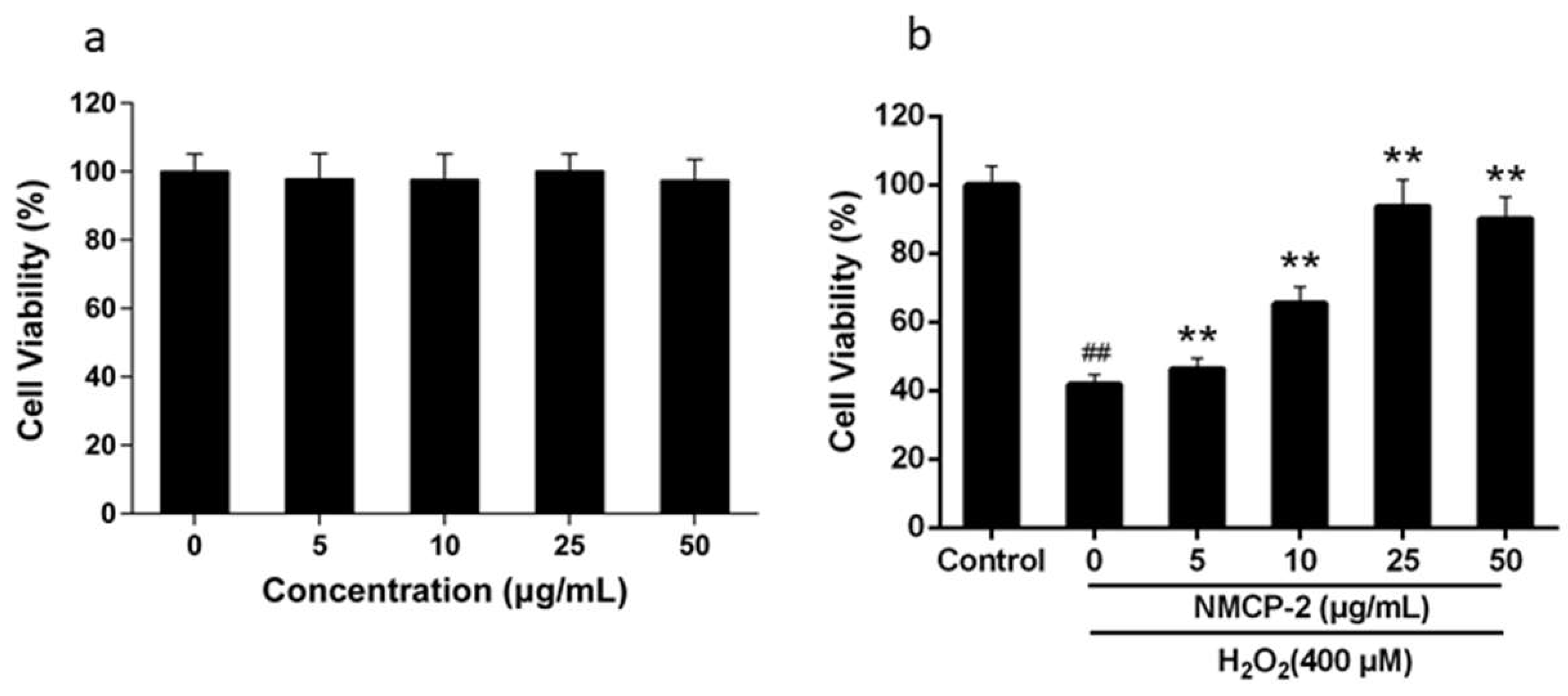
2.4. Effects of NMCP-2 on the Viability of HEK 293T Cells
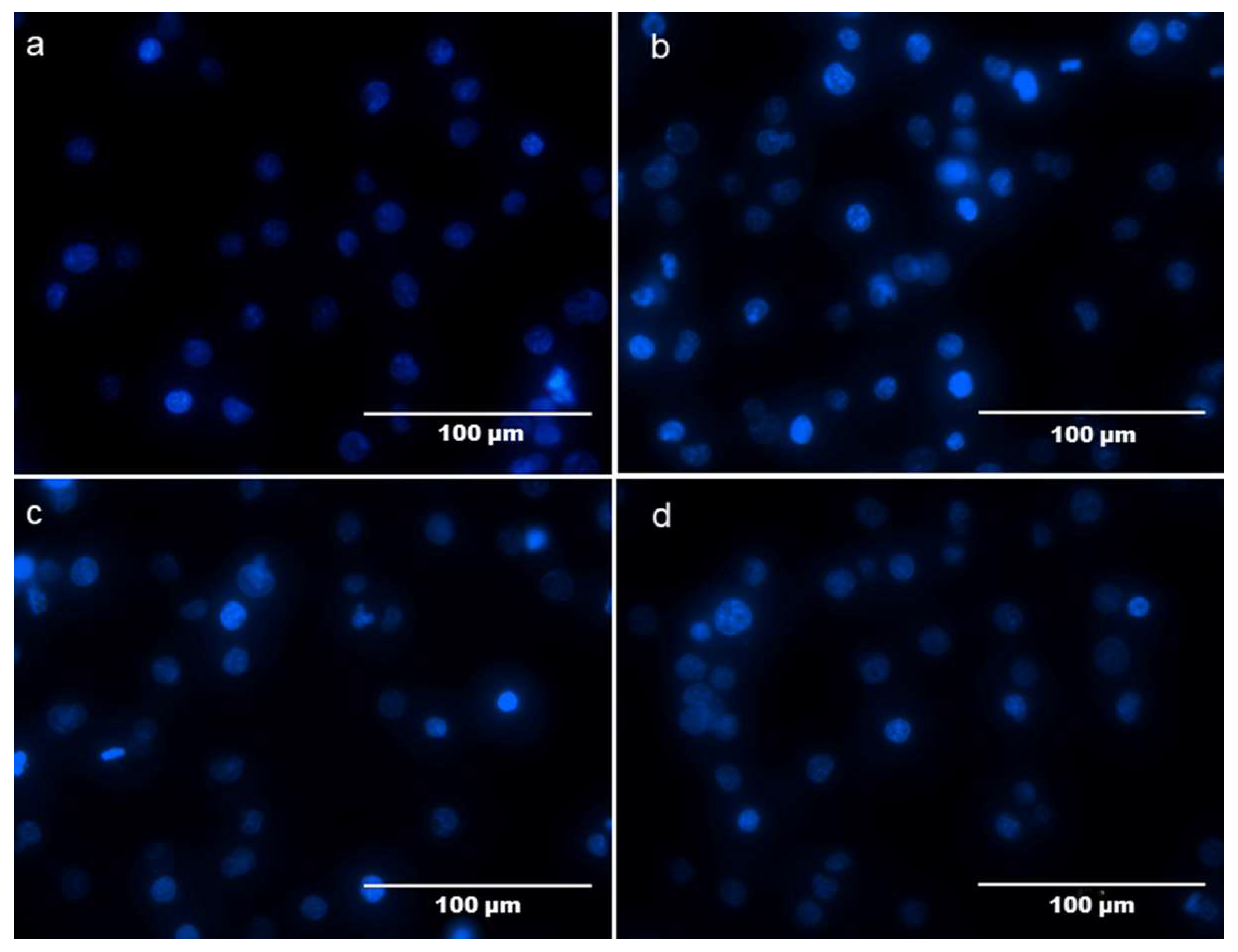
2.5. Protective Effect of NMCP-2 against H2O2-Induced HEK 293T Cells Apoptosis
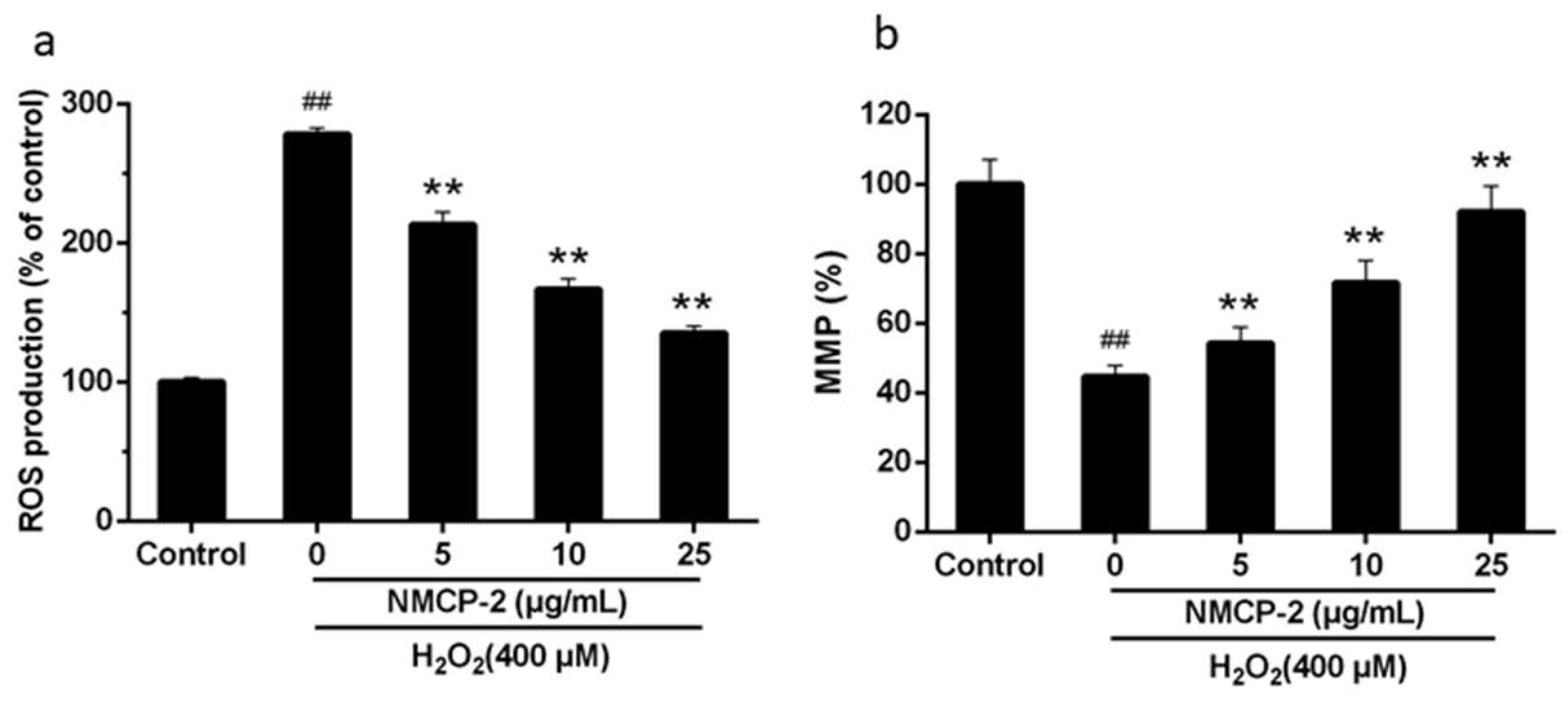
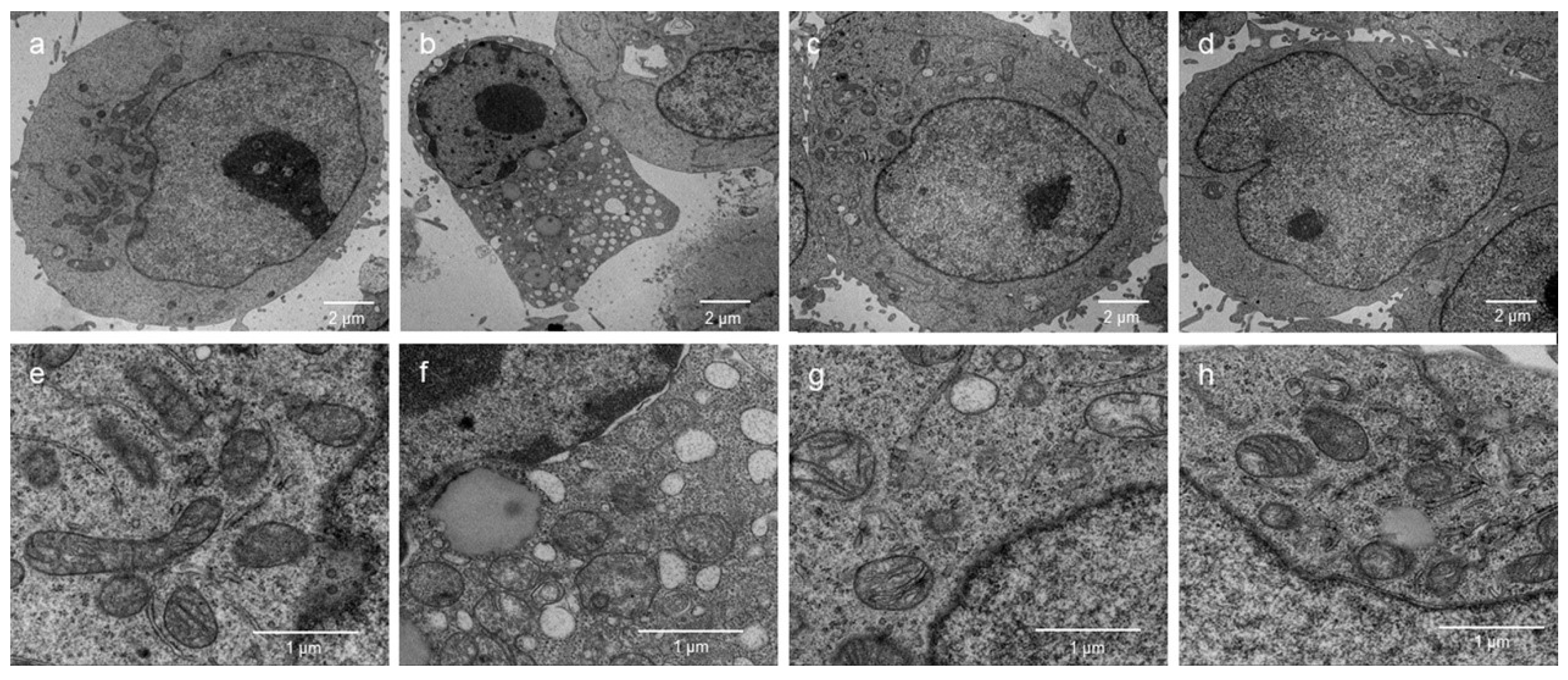
2.6. Effects of the NMCP-2 on Ultrastructure Changes Caused by H2O2
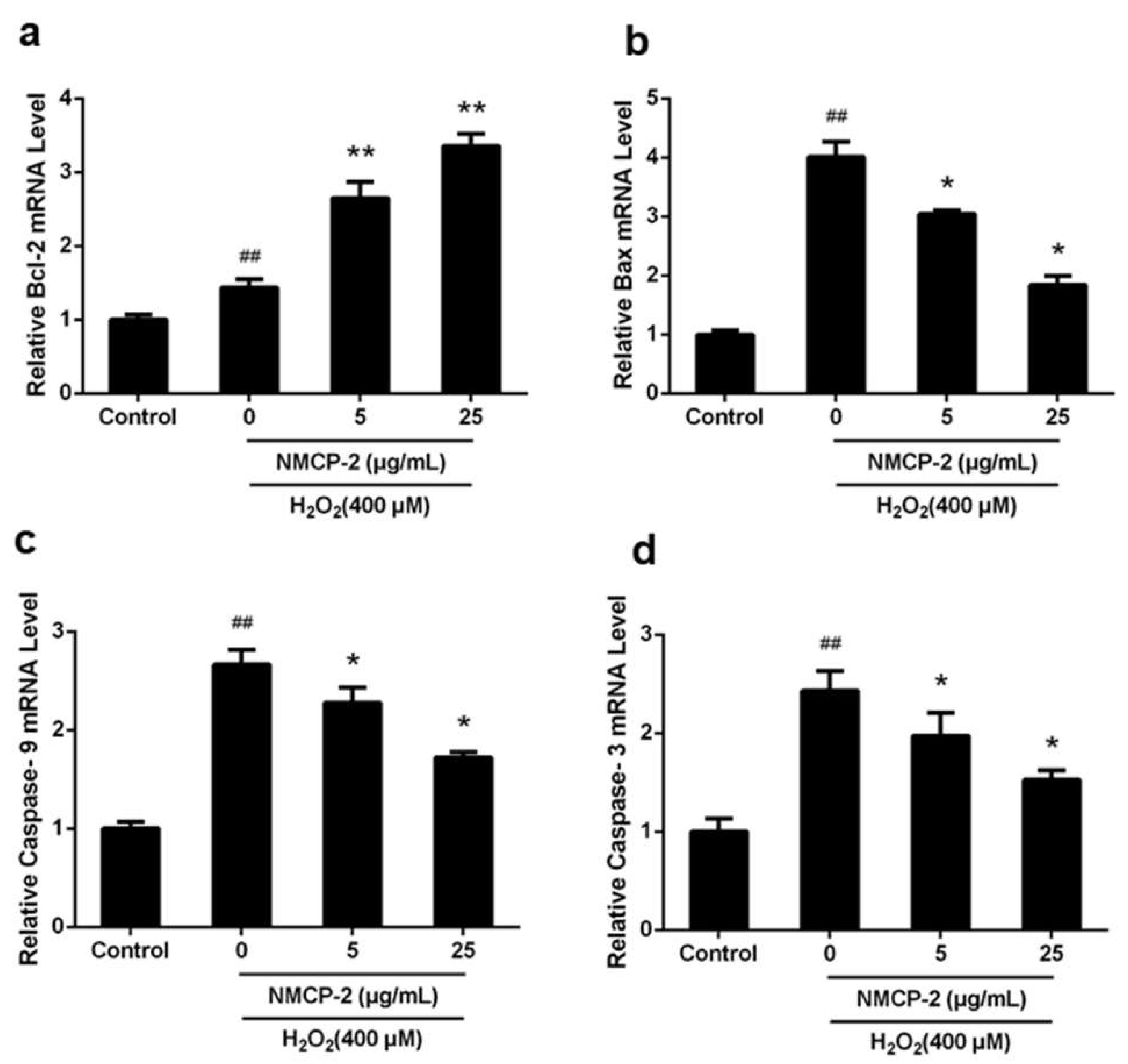
2.7. mRNA Expression of Apoptosis Related Genes after NMCP-2 Exposure
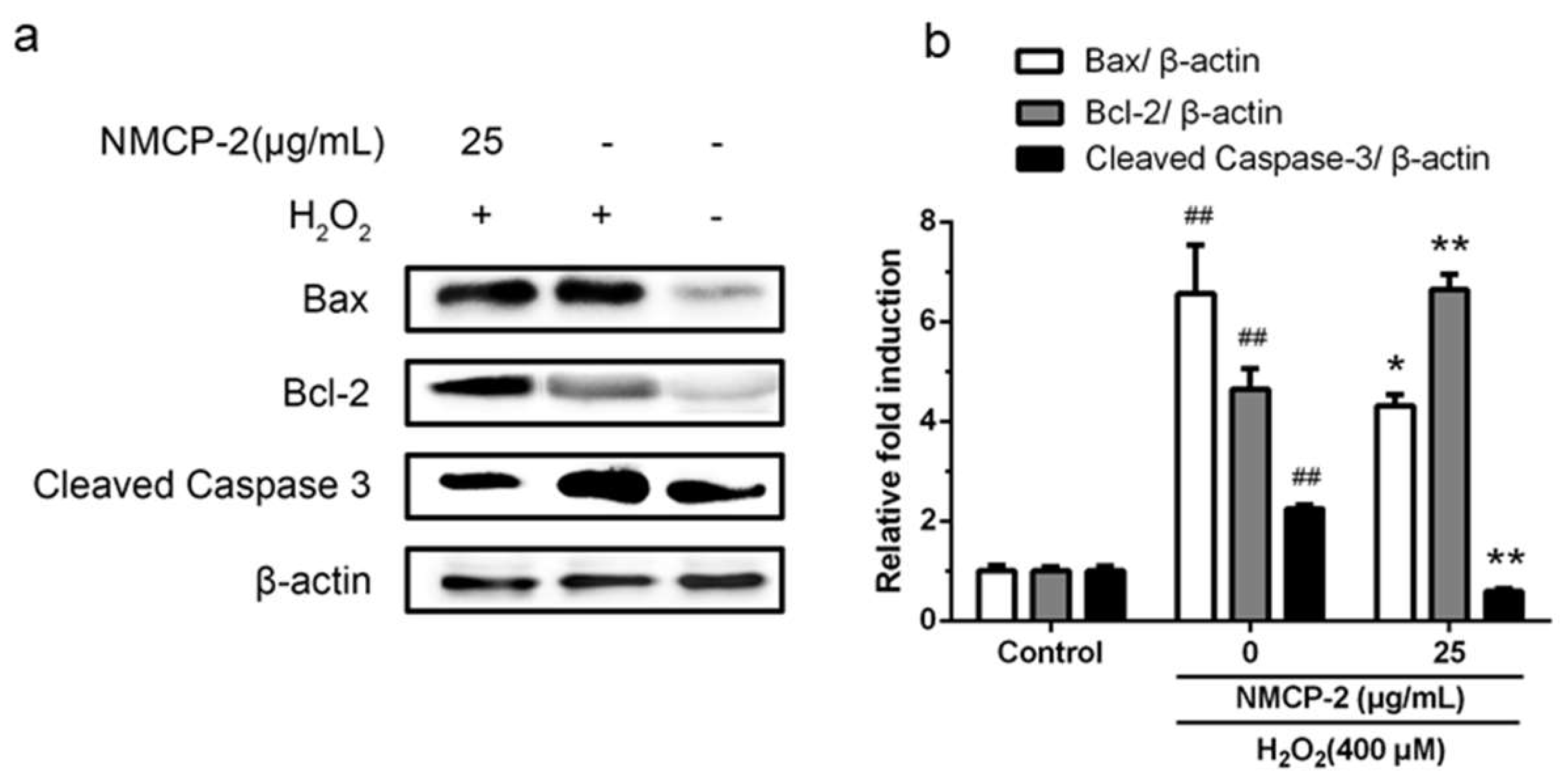
2.8. Effect of NMCP-2 on Cell Signals Related to Apoptosis Determined by Western Blot
3. Materials and Methods
3.1. Materials and Reagents
3.2. Extraction and Purification of MCP
3.3. Radical Scavenging Activity and Metal Chelating In Vitro
3.3.1. DPPH Radical Scavenging Assay
3.3.2. Ferrous Chelating Activity
3.4. Preliminary Characterization of NMCP-2
3.4.1. Fourier-Transform infrared (FT-IR) Spectroscopy Assay
3.4.2. Measurement of Molecular Weights
3.4.3. Monosaccharide Composition Analysis
3.4.4. Nuclear Magnetic Resonance (NMR) Spectroscopy Analysis
3.5. Culture of HEK 293T Cell Lines
3.6. Cell Viability Assay
3.7. Measurement of Intracellular ROS Production
3.8. Determination of Mitochondrial Membrane Potential (MMP)
3.9. Fluorescent Microscopy Measurements
3.10. Transmission Electron Microscopy (TEM) Analyses
3.11. Quantitative Real-Time Polymerase Chain Reaction (q-PCR)
3.12. Western Blotting Analysis
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wang, Z.J.; Xie, J.H.; Kan, L.J.; Wang, J.Q.; Shen, M.Y.; Li, W.J.; Nie, S.P.; Xie, M.Y. Sulfated polysaccharides from Cyclocarya paliurus reduce H2O2-induced oxidative stress in RAW264.7 cells. Int. J. Biol. Macromol. 2015, 80, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhou, C.; Huang, S.; Jiang, C. Selenium polysaccharide SPMP-2a from Pleurotus geesteranus alleviates H2O2-induced oxidative damage in HaCaT cells. Biomed. Res. Int. 2017, 4940384. [Google Scholar] [CrossRef]
- Xiong, C.; Li, Q.; Chen, C.; Chen, Z.; Huang, W. Neuroprotective effect of crude polysaccharide isolated from the fruiting bodies of Morchella importuna against H2O2-induced PC12 cell cytotoxicity by reducing oxidative stress. Biomed. pharmacother. 2016, 83, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.C.; Xu, L.; Chen, X.; Wu, J.Y. Molecular properties and antioxidant activities of polysaccharides isolated from alkaline extract of wild Armillaria ostoyae mushrooms. Carbohydr. Polym. 2016, 137, 739–746. [Google Scholar] [CrossRef]
- Zhu, W.; Xue, X.; Zhang, Z. Structural, physicochemical, antioxidant and antitumor property of an acidic polysaccharide from Polygonum multiflorum. Int. J. Biol. Macromol. 2017, 96, 494–500. [Google Scholar] [CrossRef]
- Krichen, F.; Karoud, W.; Sila, A.; Abdelmalek, S.E.; Ghorbel, R.; Ellouz-Chaabouni, S.; Bougatef, A. Extraction, characterization and antimicrobial activity of sulfated polysaccharides from fish skins. Int. J. Biol. Macromol. 2015, 75, 283–289. [Google Scholar] [CrossRef]
- Mao, G.H.; Ren, Y.; Li, Q.; Wu, H.Y.; Jin, D.; Zhao, T.; Xu, C.Q.; Zhang, D.H.; Jia, Q.D.; Bai, Y.P.; et al. Anti-tumor and immunomodulatory activity of selenium (Se)-polysaccharide from Se-enriched Grifola frondosa. Int. J. Biol. Macromol. 2016, 82, 607–613. [Google Scholar] [CrossRef]
- Stachowiak, B.; Regula, J. Health-promoting potential of edible macromycetes under special consideration of polysaccharides: A review. Eur. Food Res. Technol. 2012, 234, 369–380. [Google Scholar] [CrossRef]
- Cai, L.; Zou, S.; Liang, D.; Luan, L. Structural characterization, antioxidant and hepatoprotective activities of polysaccharides from Sophorae tonkinensis Radix. Carbohydr. Polym. 2018, 184, 354–365. [Google Scholar] [CrossRef]
- Su, C.A.; Xu, X.Y.; Liu, D.Y.; Wu, M.; Zeng, F.Q.; Zeng, M.Y.; Wei, W.; Jiang, N.; Luo, X. Isolation and characterization of exopolysaccharide with immunomodulatory activity from fermentation broth of Morchella conica. DARU J. Pharm. Sci. 2013, 21, 5. [Google Scholar] [CrossRef] [PubMed]
- Turkoglu, A.; Kivrak, I.; Mercan, N.; Duru, M.E.; Gezer, K.; Turkoglu, H. Antioxidant and antimicrobial activities of Morchella conica Pers. Afr. J. Biotechnol. 2006, 5, 1146–1150. [Google Scholar] [CrossRef]
- Wu, G.H.; Hu, T.; Li, Z.Y.; Huang, Z.L.; Jiang, J.G. In vitro antioxidant activities of the polysaccharides from Pleurotus tuber-regium (Fr.) Sing. Food Chem. 2014, 148, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.T.; Ma, X.M.; Liu, S.T.; Liao, Y.L.; Zhao, G.Q. Isolation, purification and antioxidant activities of polysaccharides from Grifola frondosa. Carbohydr. Polym. 2012, 89, 61–66. [Google Scholar] [CrossRef]
- Sun, H.H.; Mao, W.J.; Chen, Y.; Guo, S.D.; Li, H.Y.; Qi, X.H.; Chen, Y.L.; Xu, J. Isolation, chemical characteristics and antioxidant properties of the polysaccharides from marine fungus Penicillium sp. F23-2. Carbohydr. Polym. 2009, 78, 117–124. [Google Scholar] [CrossRef]
- Ghazala, I.; Sila, A.; Frikha, F.; Driss, D.; Ellouz-Chaabouni, S.; Haddar, A. Antioxidant and antimicrobial properties of water soluble polysaccharide extracted from carrot peels by-products. J. Food Sci. Technol. 2015, 52, 6953–6965. [Google Scholar] [CrossRef]
- Tong, H.B.; Liu, X.M.; Tian, D.; Sun, X. Purification, chemical characterization and radical scavenging activities of alkali-extracted polysaccharide fractions isolated from the fruit bodies of Tricholoma matsutake. World J. Microbiol. Biotechnol. 2013, 29, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, X.Y.; Wang, M.Z.; Jiao, L.L. Characterization and antioxidant activities of acidic polysaccharides from Gynostemma pentaphyllum (Thunb.) Markino. Carbohydr. Polym. 2015, 127, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Lv, G.Y.; Song, T.T.; Jin, Q.L.; Huang, J.B.; Fan, L.F.; Cai, W.M. Comparison of the preliminary characterizations and antioxidant properties of polysaccharides obtained from Phellinus baumii growth on different culture substrates. Carbohydr. Polym. 2015, 132, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yu, N.; Wang, Y.; Sun, Y.; Lu, K.; Guan, W. Extraction optimization of Bruguiera gymnorrhiza polysaccharides with radical scavenging activities. Carbohydr. Polym. 2013, 96, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Wan, Y.; Xu, J.Y.; Wu, G.H.; Li, L.; Yao, X.H. Ultrasound extraction of polysaccharides from mulberry leaves and their effect on enhancing antioxidant activity. Carbohydr. Polym. 2016, 137, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Vida, C.; Toda, I.M.; de Cruces, J.; Garrido, A.; Gonzalez-Sanchez, M.; De la Fuente, M. Role of macrophages in age-related oxidative stress and lipofuscin accumulation in mice. Redox Biol. 2017, 12, 423–437. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Inoguchi, T.; Yamasaki, T.; Yamato, M.; Ide, M.; Sonoda, N.; Yamada, K.; Takayanagi, R. A novel DPP-4 inhibitor teneligliptin scavenges hydroxyl radicals: In vitro study evaluated by electron spin resonance spectroscopy and in vivo study using DPP-4 deficient rats. Metabolism 2016, 65, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Han, S.M.; Kim, J.M.; Park, K.K.; Chang, Y.C.; Pak, S.C. Neuroprotective effects of melittin on hydrogen peroxide-induced apoptotic cell death in neuroblastoma SH-SY5Y cells, Neuroprotective effects of melittin on hydrogen peroxide-induced apoptotic cell death in neuroblastoma SH-SY5Y cells. BMC Complement. Altern. Med. 2014, 14, 286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.S.; Li, W.J.; Zhang, X.Y.; Yan, Y.X.; Nie, S.P.; Gong, D.M.; Tang, X.F.; He, M.; Xie, M.Y. Ganoderma atrum polysaccharide ameliorates anoxia/reoxygenation-mediated oxidative stress and apoptosis in human umbilical vein endothelial cells. Int. J. Biol. Macromol. 2017, 98, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Wang, H.; Liu, M.; Li, X.; Huang, M.; Zhou, H.; Lin, S.; Lin, Z.; Yang, B. Ganoderma lucidum polysaccharide peptide prevents renal ischemia reperfusion injury via counteracting oxidative stress. Sci. Rep. 2015, 5, 16910. [Google Scholar] [CrossRef] [PubMed]
- Zahedifard, M.; Faraj, F.L.; Paydar, M.; Yeng Looi, C.; Hajrezaei, M.; Hasanpourghadi, M.; Kamalidehghan, B.; Abdul Majid, N.; Mohd Ali, H.; Ameen Abdulla, M. Synthesis, characterization and apoptotic activity of quinazolinone Schiff base derivatives toward MCF-7 cells via intrinsic and extrinsic apoptosis pathways. Sci. Rep. 2015, 5, 11544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derakhshan, A.; Chen, Z.; Van Waes, C. Therapeutic small molecules target inhibitor of apoptosis proteins in cancers with deregulation of extrinsic and intrinsic cell death pathways. Clin. Cancer Res. 2017, 23, 1379–1387. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Ichim, G.; Tait, S.W. A fate worse than death: Apoptosis as an oncogenic process. Nat. Rev. Cancer 2016, 16, 539–548. [Google Scholar] [CrossRef]
- Wang, X.; Miao, J.; Yan, C.; Ge, R.; Liang, T.; Liu, E.; Li, Q. Chitosan attenuates dibutyltin-induced apoptosis in PC12 cells through inhibition of the mitochondria-dependent pathway. Carbohydr. Polym. 2016, 151, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Kepp, O.; Kroemer, G. Caspase-3 and prostaglandins signal for tumor regrowth in cancer therapy. Oncogene 2012, 31, 2805–2808. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Sun, Y.H.; Guo, X.L.; Liu, C.; Mao, Q.; Hou, J.M. Optimization of ultrasonic-microwave synergistic extraction of polysaccharides from Morchella conica. J. Food Process. Pres. 2018, 42, e13423. [Google Scholar] [CrossRef]
- Bilos, M.S. Antioxidant determinations by the use of a stable free radical. Nature 1958, 181, 1199–1200. [Google Scholar] [CrossRef]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Liu, C.; Sun, Y.H.; Mao, Q.; Guo, X.L.; Li, P.; Xu, N. Characteristics and Antitumor Activity of Morchella esculenta Polysaccharide Extracted by Pulsed Electric Field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef] [PubMed]
- Albersheim, P.; Nevins, D.J.; English, P.D.; Karr, A. A method for the analysis of sugars in plant cell-wall polysaccharides by gas-liquid chromatography. Carbohydr. Res 1967, 5, 340–345. [Google Scholar] [CrossRef]
- Peng, Y.F.; Han, B.Q.; Liu, W.S.; Zhou, R.X. Deproteinization and structural characterization of bioactive exopolysaccharides from Ganoderma sinense mycelium. Sep. Sci. Technol. 2016, 51, 359–369. [Google Scholar] [CrossRef]
- Yan, X.Y.; Yang, X.; Hao, X.H.; Ren, Q.R.; Gao, J.P.; Wang, Y.; Chang, N.; Qiu, Y.L.; Song, G.H. Sodium Fluoride Induces Apoptosis in H9c2 Cardiomyocytes by Altering Mitochondrial Membrane Potential and Intracellular ROS Level. Biol. Trace Elem. Res. 2015, 166, 210–215. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Shen, J.J.; Chen, Y.Y.; Liu, H.Z.; Zhou, H.; Bai, Z.B.; Hu, Z.M.; Guo, X.M. PINK1 protects against oxidative stress induced senescence of human nucleus pulposus cells via regulating mitophagy. Biochem. Biophys. Res. Commun. 2018, 504, 406–414. [Google Scholar] [CrossRef]
- Mehmood, T.; Maryam, A.; Zhang, H.; Li, Y.M.; Khan, M.; Ma, T.H. Deoxyelephantopin induces apoptosis in HepG2 cells via oxidative stress, NF-κB inhibition and mitochondrial dysfunction. IUBMB 2016, 43, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, L.; Owen, M.D.; Albert, L.; Jones, M.D. Epithelial cell specialization within human peyer’s patches: An ultrastructrual study of intestinal lymphiod follicles. Gastroenterology 1974, 66, 189–203. [Google Scholar] [CrossRef]
- Wang, Y.; Lei, T.; Yuan, J.T.; Wu, Y.G.; Shen, X.Y.; Gao, J.L.; Feng, W.; Lu, Z.B. GCN2 deficiency ameliorates doxorubicin-induced cardiotoxicity by decreasing cardiomyocyte apoptosis and myocardial oxidative stress. Redox Biol. 2018, 17, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.Y.; Wang, B.; Wang, L.; Abraham, N.; Tao, K.; Huang, L.; Shi, W.; Dong, Y.S.; Qu, Y. Pre-ischemia melatonin treatment alleviated acute neuronal injury after ischemic stroke by inhibiting endoplasmic reticulum stress-dependent autophagy via PERK and IRE1 signalings. J. Pineal Res. 2017, 62, e12395. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residues | Sugar linkage | 1 | 2 | 3 | 4 | 5 | 6 | |
|---|---|---|---|---|---|---|---|---|
| A | →4)-α-D-Glup-(1→ | H | 5.12 | 3.59 | 3.93 | 3.81 | 3.56 | 3.96 |
| C | 99.30 | 71.16 | 71.90 | 76.48 | 75.31 | 61.02 | ||
| B | →6)-α-D-Glup-(1→ | H | 5.02 | 3.60 | 3.73 | 3.56 | 3.96 | 4.05 |
| C | 98.22 | 77.26 | 74.15 | 70.35 | 71.16 | 66.34 | ||
| C | →4, 6)-α-D-Glup-(1→ | H | 5.08 | - | 3.96 | 3.69 | - | 3.69 |
| C | 96.24 | 72.54 | 74.15 | 77.73 | - | 63.33 | ||
| D | α-T-Manp | H | 4.98 | 4.00 | 3.72 | 3.57 | 3.69 | 3.69 |
| C | 102.22 | 70.05 | 69.87 | 66.86 | 73.29 | 60.86 | ||
| E | →3)-β-Manp-(1→ | H | - | 4.13 | 3.96 | 3.69 | 3.42 | 3.81 |
| C | 101.15 | 71.16 | 74.31 | 67.82 | 76.08 | 61.55 | ||
| F | →4)-β-D-Xylp-(1→ | H | 4.67 | 3.56 | 3.69 | 3.85 | 4.13 | - |
| C | 101.30 | 79.61 | 75.69 | 77.01 | 66.86 | - | ||
| G | →4)-β-D-Galp-(1→ | H | 4.54 | 3.33 | 3.56 | 3.72 | - | - |
| C | 103.43 | 73.29 | 76.67 | 76.38 | - | - |
| Name | Primers | Sequence (5’-3’) |
|---|---|---|
| CASPASE-3 | CASPASE-3F | AACTGGACTGTGGCATTGAG |
| CASPASE-3R | ACAAAGCGACTGGATGAACC | |
| CASPASE-9 | CASPASE-9F | GGAAGAGGGACAGATGAATG |
| CASPASE-9R | TTGTTTGGCACCACTCAG | |
| BAX | BAX-F | CCTTTTCTACTTTGCCAGCAAAC |
| BAX-R | GAGGCCGTCCCAACCAC | |
| BCL-2 | BCL-2F | CGGTTCAGGTACTCAGTCATC |
| BCL-2R | CGGTGGGGTCATGTGTGTG | |
| GAPDH | GAPDH-F | GTCAACGGATTTGGTCGTA |
| GAPDH-R | GTAGTTGAGGTCAATGAAG |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, N.; Lu, Y.; Hou, J.; Liu, C.; Sun, Y. A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells. Int. J. Mol. Sci. 2018, 19, 4027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124027
Xu N, Lu Y, Hou J, Liu C, Sun Y. A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells. International Journal of Molecular Sciences. 2018; 19(12):4027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124027
Chicago/Turabian StyleXu, Na, Yi Lu, Jumin Hou, Chao Liu, and Yonghai Sun. 2018. "A Polysaccharide Purified from Morchella conica Pers. Prevents Oxidative Stress Induced by H2O2 in Human Embryonic Kidney (HEK) 293T Cells" International Journal of Molecular Sciences 19, no. 12: 4027. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124027