Evolutionary Analysis of Plastid Genomes of Seven Lonicera L. Species: Implications for Sequence Divergence and Phylogenetic Relationships

Abstract
:1. Introduction
2. Results
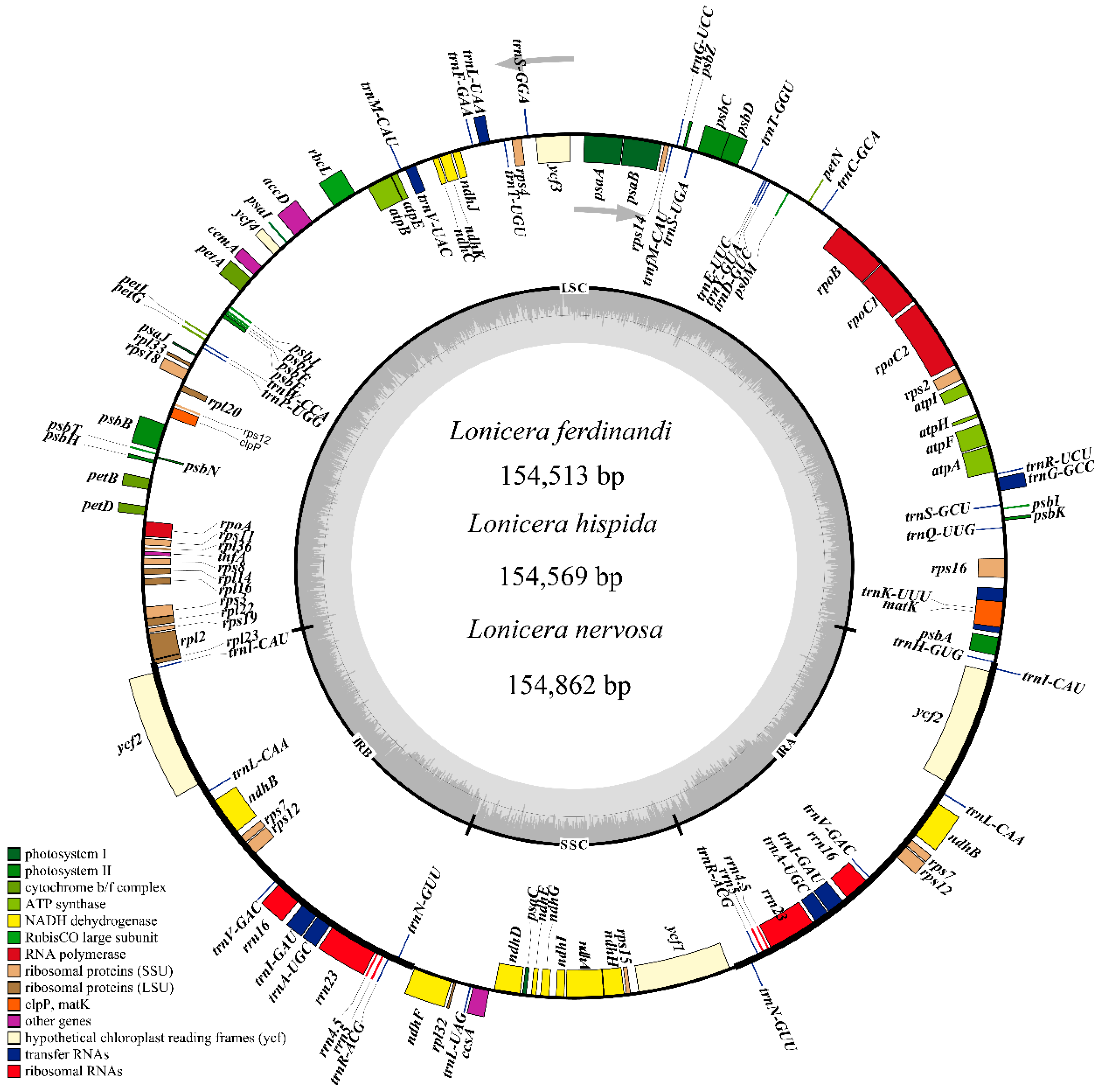
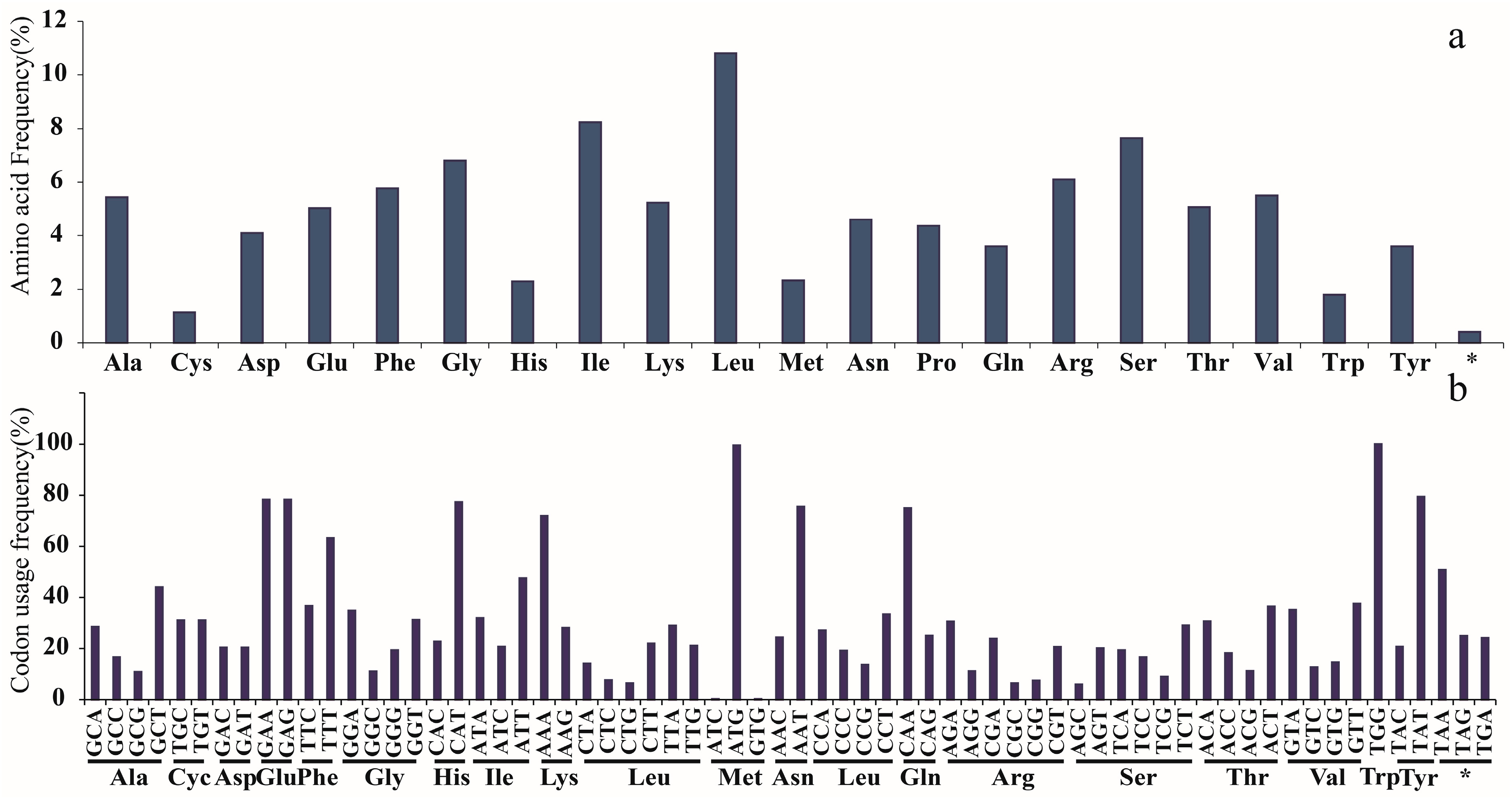
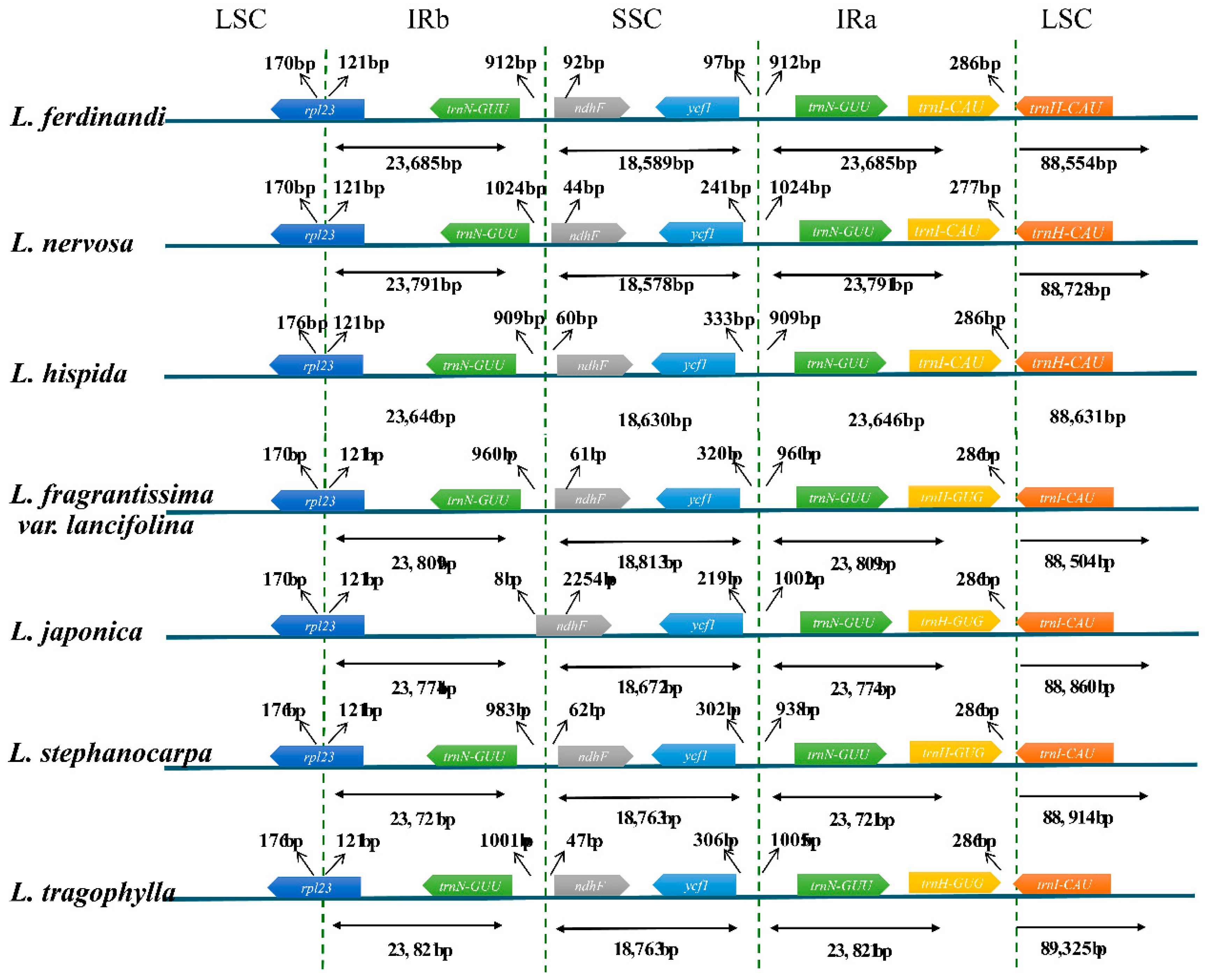
2.1. Characters of Plastid Genomes
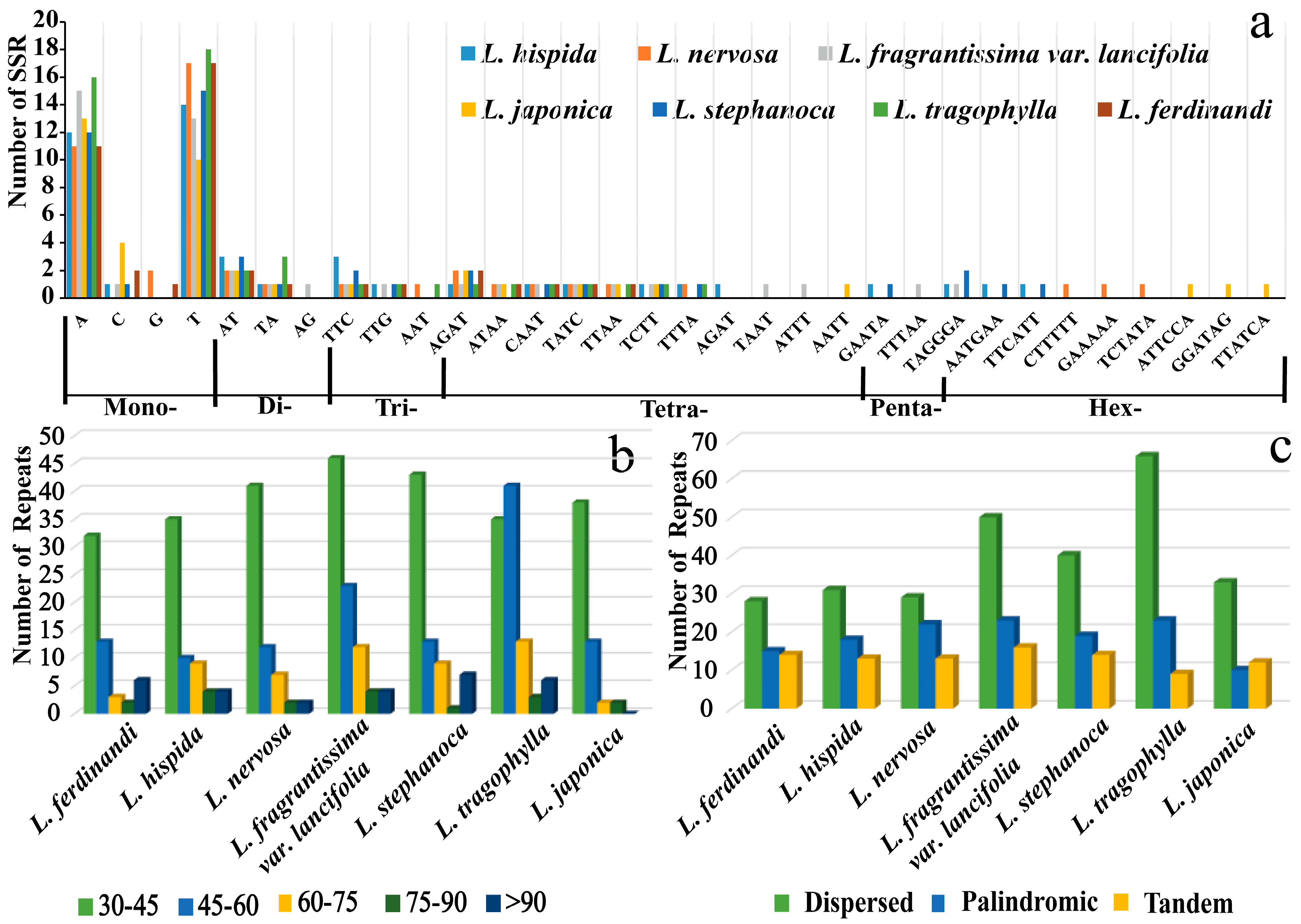
2.2. Repeat Sequences Analysis
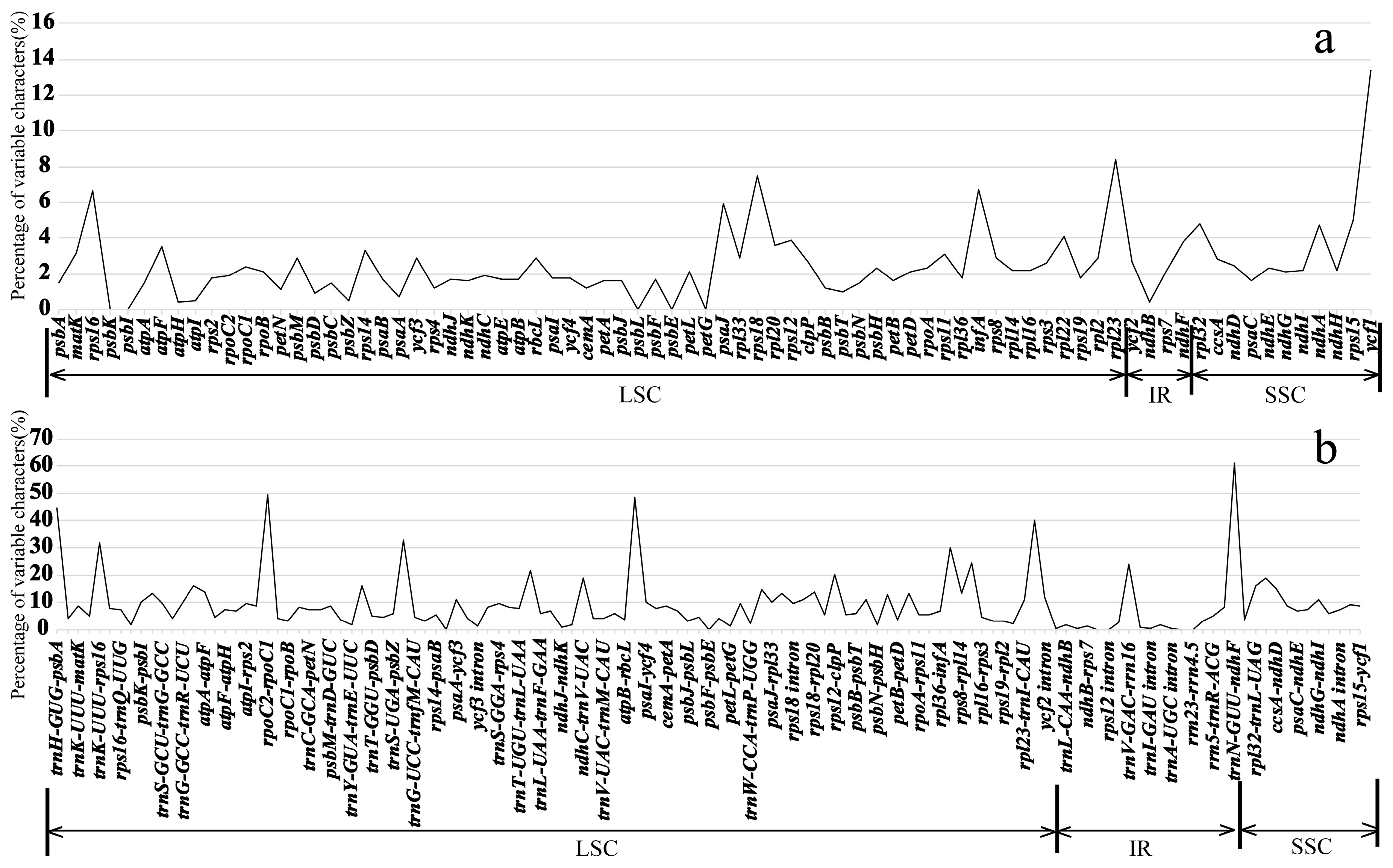
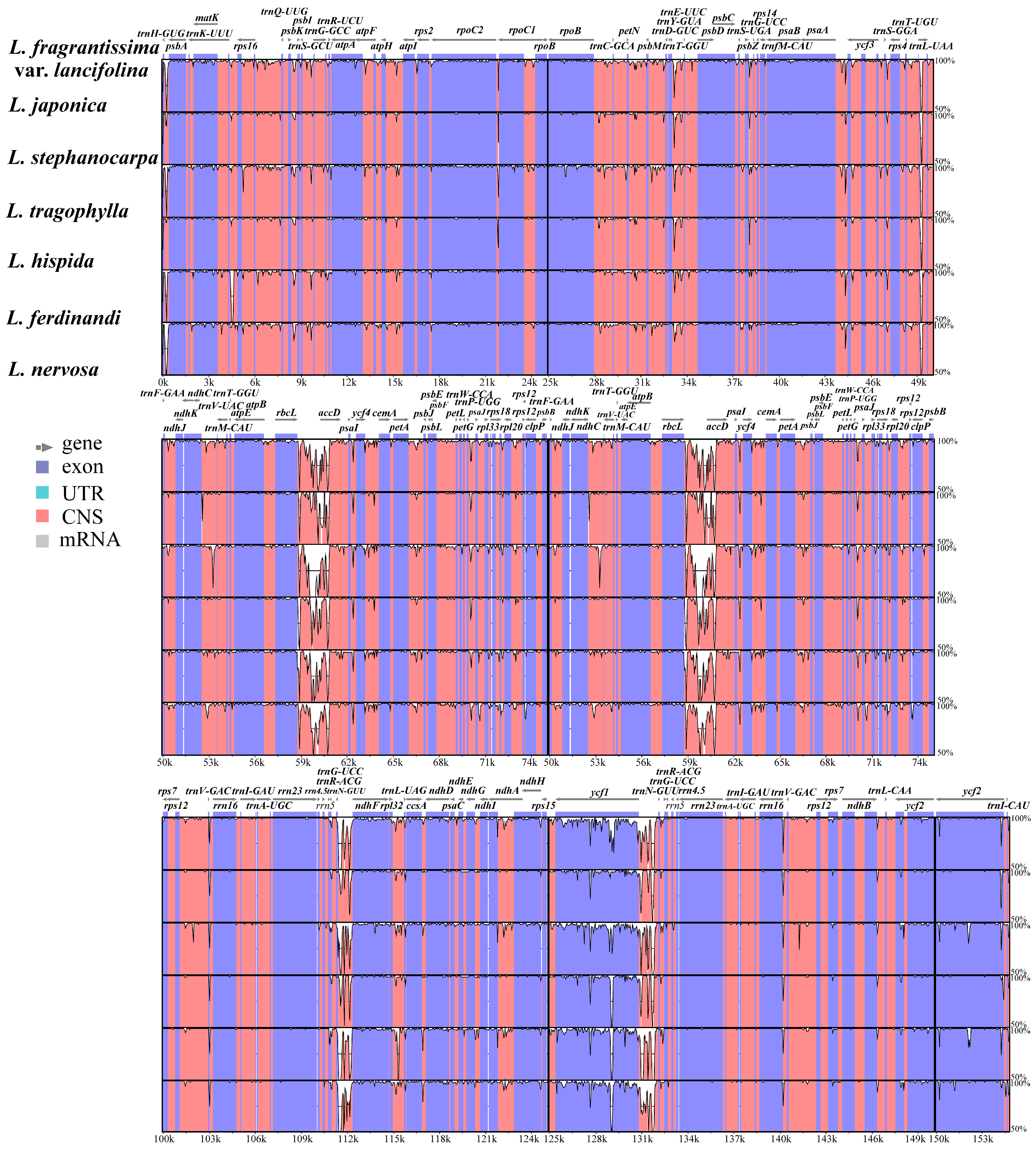
2.3. Divergence Hotspots of Plastid Genomes
2.4. Positive Selection Analysis
2.5. Phylogenetic Analysis
3. Discussion
3.1. Features of Plastid Genomes
3.2. Repeat Sequence Variations
3.3. Positive Selection Analysis
3.4. Phylogenetic Relationship
4. Method
4.1. Sampling and Sequencing
4.2. Chloroplast Genome Assembly and Annotation
4.3. Repeat Sequence Analysis
4.4. Sequence Divergence Analysis
4.5. Gene Selection Sites Analysis
4.6. Phylogenetic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Jacobs, B.; Lens, F.; Smets, E. Evolution of fruit and seed characters in the Diervilla and Lonicera clades (Caprifoliaceae, Dipsacales). Ann. Bot. 2009, 104, 253–276. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.S. A preliminary numerical taxonomy of the family Caprifoliaceae. Acta Phytotaxon. Sin. 1983, 21, 26–32. [Google Scholar]
- Li, H.J.; Li, P.; Wang, M.C.; Ye, W.C. A new secoiridoid glucoside from Lonicera japonica. Chin. J. Nat. Med. 2003, 3, 132–133. [Google Scholar]
- Shang, X.F.; Pan, H.; Li, M.X.; Miao, X.L.; Ding, H. Lonicera japonica thunb.: Ethnopharmacology, phytochemistry and pharmacology of an important traditional Chinese medicine. J. Ethnopharmacol. 2011, 138, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.T.; Chen, J.; Song, Y.; Sheng, L.S.; Li, P.; Qi, L.W. Identification and quantification of 32 bioactive compounds in Lonicera species by high performance liquid chromatography coupled with time-of-flight mass spectrometry. J. Pharm. Biomed. 2008, 48, 1351–1360. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Qi, L.W.; Li, H.J.; Li, P.; Yi, L.; Ma, H.L.; Tang, D. Simultaneous determination of iridoids, phenolic acids, flavonoids, and saponins in Flos Lonicerae and Flos Lonicerae Japonicae by HPLC-DAD-ELSD coupled with principal component analysis. J. Sep. Sci. 2007, 30, 3181–3192. [Google Scholar] [CrossRef]
- Rehder, A. Synopsis of the Genus Lonicera. Mo. Bot. Gard. Annu. Rep. 1903, 27–232. [Google Scholar] [CrossRef]
- Lin, S.L.; Yang, L.M.; Qiao, G.P. The study on greenwood cutting reproduction technology of Lonicera edulls. J. Jilin Agric. Univ. 2003, 25, 10. [Google Scholar]
- Choi, C.W.; Jung, H.A.; Kang, S.S.; Choi, J.S. Antioxidant constituents and a new triterpenoid glycoside from Flos Lonicerae. Arch. Pharm. Res. 2007, 30, 1–7. [Google Scholar] [CrossRef]
- Gao, Z.T.; Liu, Y.; Wang, X.Y.; Song, J.Y.; Chen, S.L.; Ragupathy, S.; Han, J.P.; Newmaster, S.G. Derivative technology of DNA barcoding (nucleotide signature and SNP double peak methods) detects adulterants and substitution in Chinese patent medicines. Sci. Rep. 2017, 7, 5858. [Google Scholar] [CrossRef]
- Rai, A.; Kamochi, H.; Suzuki, H.; Nakamura, M.; Takahashi, H.; Hatada, T.; Saito, K.; Yamazaki, M. De novo transcriptome assembly and characterization of nine tissues of Lonicera japonica, to identify potential candidate genes involved in chlorogenic acid, luteolosides, and secoiridoid biosynthesis pathways. J. Nat. Med. 2017, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nina, T.; Michael, J.D.; Li, J.H. Phylogenetics of the Caprifolieae and Lonicera (Dipsacales) based on nuclear and chloroplast DNA sequences. Syst. Bot. 2008, 33, 776–783. [Google Scholar]
- He, L.; Qian, J.; Li, X.W.; Sun, Z.Y.; Xu, X.L.; Chen, S.L. Complete chloroplast genome of medicinal plant Lonicera japonica: Genome rearrangement, intron gain and loss, and implications for phylogenetic studies. Molecules 2017, 22, 249. [Google Scholar] [CrossRef] [PubMed]
- Wicke, S.; Schneeweiss, G.M.; dePamphilis, C.W.; Müller, K.F.; Quandt, D. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function. Plant Mol. Biol. 2011, 76, 273–297. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.D. Plastid chromosomes: Structure and evolution. In Cell Culture and Somatic Cell Genetics of Plants; Bogorad, L., Vasil, I.K., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 5–53. [Google Scholar]
- Raubeson, L.A.; Jansen, R.K. Chloroplast genomes of plants. In Plant Diversity and Evolution: Genotypic and Phenotypic Variation in Higher Plants; Henry, R.J., Ed.; CAB International: London, UK, 2005; pp. 45–68. [Google Scholar]
- Jansen, R.K.; Ruhlman, T.A. Plastid genomes of seed plants. In Genomics of Chloroplasts and Mitochondria, Advances in Photosynthesis and Respiration; Bock, R., Knoop, V., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 103–126. [Google Scholar]
- Ruhlman, T.A.; Jansen, R.K. The plastid genomes of flowering plants. In Chloroplast Biotechnology: Methods and Protocols; Maliga, P., Ed.; Spring: New York, NY, USA, 2014; pp. 3–38. [Google Scholar]
- George, B.; Bhatt, B.S.; Awasthi, M.; George, B.; Singh, A.K. Comparative analysis of microsatellites in chloroplast genomes of lower and higher plants. Curr. Genet. 2015, 61, 665–667. [Google Scholar] [CrossRef] [PubMed]
- Tobes, R.; Pareja, E. Repetitive extragenic palindromic sequences in the Pseudomonas syringae pv. tomato DC3000 genome: Extragenic signals for genome reannotation. Res. Microbiol. 2005, 156, 424–433. [Google Scholar] [PubMed]
- Li, S.F.; Su, T.; Cheng, G.Q.; Wang, B.X.; Li, X.; Deng, C.L.; Gao, W.J. Chromosome evolution in connection with repetitive sequences and epigenetics in plants. Genes 2017, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yue, M.; Niu, C.; Ma, X.F.; Li, Z.H. Comparative analysis of the complete chloroplast genome of four endangered herbals of Notopterygium. Genes 2017, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.B.; Wu, Y.; Yang, J.; Shahzad, K.; Li, Z.H. Comparative chloroplast genomics of Dipsacales species: Insights into sequence variation, adaptive evolution, and phylogenetic relationships. Front. Plant Sci. 2018, 9, 689. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.L.; Wan, J.N.; Guo, Y.P.; Ge, S.; Rao, G.Y. Adaptive evolution of the chrysanthemyl diphosphate synthase gene involved in irregular monoterpene metabolism. BMC Evol. Biol. 2012, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Shiu, S.H.; Byrnes, J.K.; Pan, R.S.; Zhang, P.; Li, W.H. Role of positive selection in the retention of duplicate genes in mammalian genomes. Proc. Natl. Acad. Sci. USA 2006, 103, 2232–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.C.; Zhou, T.; Duan, D.; Yang, J.; Feng, L.; Zhao, G.F. Comparative analysis of the complete chloroplast genomes of five Quercus species. Front Plant. Sci. 2016, 7, 573. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Chen, J.; Wang, T. Adaptive evolution in the chloroplast gene rps4 in ferns. Bull. Bot. Res. 2010, 30, 42–50. [Google Scholar]
- Hao, D.C.; Chen, S.L.; Xiao, P.G. Molecular evolution and positive darwinian selection of the chloroplast maturase matK. J. Plant Res. 2010, 123, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, C.M.; Zhao, X.; Fei, Z.G.; Wan, K.K.; Zhang, Z.; Pang, X.M.; Yin, X.; Bai, Y.; Sun, X.Q.; et al. The jujube genome provides insights into genome evolution and the domestication of sweetness/acidity taste in fruit trees. PLoS. Genet. 2016, 12, e1006433. [Google Scholar] [CrossRef]
- Bock, R. Structure, function, and inheritance of plastid genomes. In Cell and Molecular Biology of Plastids; Bock, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 29–63. [Google Scholar]
- Prade, V.M.; Gundlach, H.; Twardziok, S.; Chapman, B.; Tan, C.; Langridge, P.; Schulman, A.H.; Stein, N.; Waugh, R.; Zhang, G.P.; et al. The pseudogenes of barley. Plant J. 2018, 93, 502. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Salmena, L.; Zhang, J.W.; Carver, B.; Haveman, W.J.; Pandolfi, P.P. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 2010, 465, 1033–1038. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.J.; Gao, A.M.; Ji, L.J.; Xu, J. Pseudogene in cancer: Real functions and promising signature. J. Med. Genet. 2015, 52, 17–24. [Google Scholar]
- Hong, C.P.; Park, J.; Lee, Y.; Lee, M.; Park, S.G.; Uhm, Y.; Kim, C.K. accD nuclear transfer of platycodon grandiflorum and the plastid of early Campanulaceae. BMC Genom. 2017, 18, 607. [Google Scholar] [CrossRef]
- Harris, M.E.; Meyer, G.; Vandergon, T.; Vandergon, V.O. Loss of the acetyl-CoA carboxylase (accD) gene in Poales. Plant Mol. Biol. Rep. 2012, 31, 21–31. [Google Scholar] [CrossRef]
- Goremykin, V.V.; Holland, B.; Hirschernst, K.I.; Hellwig, F.H. Analysis of Acorus calamus chloroplast genome and its phylogenetic implications. Mol. Biol. Evol. 2005, 22, 1813–1822. [Google Scholar] [CrossRef]
- Rousseau-Gueutin, M.; Huang, X.; Higginson, E.; Ayliffe, M.; Day, A.; Timmis, J.N. Potential functional replacement of the plastidic acetyl-CoA carboxylase subunit (accD) gene by recent transfers to the nucleus in some angiosperm lineages. Plant Physiol. 2013, 161, 1918–1929. [Google Scholar] [CrossRef]
- Guisinger, M.M.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. Genome-wide analyses of Geraniaceae plastid DNA reveal unprecedented patterns of increased nucleotide substitutions. Proc. Natl. Acad. Sci. USA 2008, 105, 18424–18429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalier-Smith, T. Chloroplast evolution: Secondary symbiogenesis and multiple losses. Curr. Biol. CB 2002, 12, R62. [Google Scholar] [CrossRef]
- Pauwels, M.; Vekemans, X.; Godé, C.; Frérot, H.; Castric, V.; Saumitou-Laprade, P. Nuclear and chloroplast DNA phylogeography reveals vicariance among European populations of the model species for the study of metal tolerance, Arabidopsis halleri (Brassicaceae). New Phytol. 2012, 193, 916–928. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhou, T.; Kanwal, N.; Zhao, Y.M.; Bai, G.Q.; Zhao, G.F. Completion of eight Gynostemma BL. (Cucurbitaceae) chloroplast genomes: Characterization, comparative analysis, and phylogenetic relationships. Front Plant. Sci. 2017, 8, 1583. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, F.; Yang, D.G.; Li, W.; Zhou, X.J.; Pei, X.Y.; Liu, Y.G.; He, K.L.; Zhang, W.S.; Ren, Z.Y.; et al. Comparative chloroplast genomics of Gossypium species: Insights into repeat sequence variations and phylogeny. Front. Plant Sci. 2018, 9, 376. [Google Scholar] [CrossRef] [PubMed]
- Erixon, P.; Oxelman, B. Whole-gene positive selection, elevated synonymous substitution rates, duplication, and indel evolution of the chloroplast clpP1 gene. PLoS ONE 2008, 3, e1386. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, Z.; Sablok, G.; Daskalova, E.; Zahmanova, G.; Apostolova, E.; Yahubyan, G.; Baev, V. Chloroplast genome analysis of resurrection tertiary relict Haberlea rhodopensis highlights genes important for desiccation stress response. Front. Plant Sci. 2017, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, M.; Ruf, S.; Bock, R. Tobacco plastid ribosomal protein S18 is essential for cell survival. Nucleic Acids. Res. 2006, 34, 4537–4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nellaeplli, S.; Ozawa, S.I.; Kuroda, H.; Takahashi, Y. The photosystem I assembly apparatus consisting of Ycf3–Y3IP1 and Ycf4 modules. Nat. Commun. 2018, 9, 2439. [Google Scholar] [CrossRef] [PubMed]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Kavanagh, T.A.; Hibberd, J.M.; Gray, J.C.; Morden, C.W.; et al. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Steane, D.A. Complete nucleotide sequence of the chloroplast genome from the Tasmanian blue gum, Eucalyptus globulus (Myrtaceae). DNA Res. 2005, 12, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Allahverdiyeva, Y.; Mamedov, F.; Mäenpää, P.; Vass, I.; Aro, E.M. Modulation of photosynthetic electron transport in the absence of terminal electron acceptors: Characterization of the rbcL deletion mutant of tobacco. BBA-Bioenergetics 2005, 1709, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Kapralov, M.V.; Filatov, D.A. Widespread positive selection in the photosynthetic Rubisco enzyme. BMC Evol. Biol. 2007, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, S.; Bédard, J.; Hirano, M.; Hirabayashi, Y.; Oishi, M.; Imai, M.; Takase, M.; Ide, T.; Nakai, M. Uncovering the protein translocon at the chloroplast inner envelope membrane. Science 2013, 339, 571–574. [Google Scholar] [CrossRef]
- Leebens-Mack, J.; Raubeson, L.A.; Cui, L.Y.; Kuehl, J.V.; Fourcade, M.H.; Chumley, T.W.; Boore, J.L.; Jansen, R.K.; dePamphilis, C.W. Identifying the basal angiosperm node in chloroplast genome phylogenies: Sampling one’s way out of the Felsenstein zone. Mol. Biol. Evol. 2005, 22, 1948–1963. [Google Scholar] [CrossRef]
- Moore, M.J.; Bell, C.D.; Soltis, P.S.; Soltis, D.E. Using plastid genome-scale data to resolve enigmatic relationships among basal angiosperms. Proc. Natl. Acad. Sci. USA 2007, 104, 19363–19368. [Google Scholar] [CrossRef] [Green Version]
- Patel, R.K.; Jain, M. NGS QC Toolkit: A toolkit for quality control of next generation sequencing data. PLoS ONE 2012, 7, e30619. [Google Scholar] [CrossRef]
- Chevreux, B.; Pfisterer, T.; Drescher, B.; Driesel, A.J.; Müller, W.E.G.; Wetter, T.; Suhai, S. Using the miraEST assembler for reliable and automated mRNA transcript assembly and SNP detection in sequenced ESTs. Genome Res. 2004, 14, 1147–1159. [Google Scholar] [CrossRef]
- Hahn, C.; Bachmann, L.; Chevreux, B. Reconstructing mitochondrial genomes directly from genomic next-generation sequencing reads—A baiting and iterative mapping approach. Nucleic Acids Res. 2013, 41, e129. [Google Scholar] [CrossRef]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schattner, P.; Brooks, A.N.; Lowe, T.M. The tRNAscan-SE, snoscan and snoGPS web servers for the detection of tRNAs and snoRNAs. Nucleic Acids Res. 2005, 33, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Kahlau, S.; Bock, R. OrganellarGenomeDRAW--a suite of tools for generating physical maps of plastid and mitochondrial genomes and visualizing expression data sets. Nucleic Acids Res. 2013, 41, W575. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Thiel, T.; Michalek, W.; Varshney, R.K.; Graner, A. Exploiting EST databases for the development and characterization of gene-derived SSR-markers in barley (Hordeum vulgare L.). Theop. Appl. Genet. 2003, 106, 411–422. [Google Scholar] [CrossRef]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.H.; Wong, W.S.W.; Nielsen, R. Bayes empirical bayes inference of amino acid sites under positive selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. Mrmodeltest v2. Program Distributed by the Author; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Swofford, D.L. Paup 4.0 for Macintosh: Phylogenetic Analysis Using Parsimony (Software and User’s Book for Macintosh); Sinauer Associates, Incorporated: Sunderland, MA, USA, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | L. ferdinandi | L. hispida | L. nervosa | L. fragrantissima var. lancifolia | L. stephanocarpa | L. tragophylla | L. japonica |
| No. of mapped reads | 180,643 | 159,034 | 158,808 | – | – | – | – |
| Average coverage | 149.1 | 419.2 | 198.4 | – | – | – | – |
| Total sequence length | 154,513 | 154,553 | 154,862 | 154,732 | 155,056 | 155,545 | 155,078 |
| Large single copy (LSC) | 88,554 | 88,631 | 88,728 | 88,504 | 88,912 | 89,299 | 88,858 |
| Inverted repeat (IR) region | 23,685 | 23,646 | 23,791 | 23,731 | 23,690 | 23,759 | 23,774 |
| Small single copy (SSC) | 18,589 | 18,630 | 18,552 | 18,766 | 18,763 | 18,728 | 18,672 |
| GC content (%) | 38.4 | 38.3 | 38.6 | 38.3 | 38.3 | 38.5 | 38.6 |
| GC content in LSC (%) | 36.9 | 36.8 | 36.9 | 36.8 | 36.8 | 37.0 | 37.1 |
| GC content in IR (%) | 43.4 | 43.4 | 43.5 | 43.3 | 43.4 | 43.5 | 43.5 |
| GC content in SSC (%) | 33.2 | 32.9 | 33.1 | 32.9 | 32.8 | 33.1 | 33.4 |
| No. of total genes | 128 | 128 | 128 | 128 | 128 | 128 | 128 |
| Protein-coding genes | 82 (4) | 82 (4) | 82 (4) | 82 (4) | 82 (4) | 82 (4) | 82 (4) |
| tRNAs genes | 37 (7) | 37 (7) | 37 (7) | 37 (7) | 37 (7) | 37 (7) | 37 (7) |
| rRNAs genes | 8 (4) | 8 (4) | 8 (4) | 8 (4) | 8 (4) | 8 (4) | 8 (4) |
| Genes with introns | 20 (4) | 20 (4) | 20 (4) | 20 (4) | 20 (4) | 20 (4) | 20 (4) |
| Gene Group | Gene Name | ||||
|---|---|---|---|---|---|
| Ribosomal RNA genes | rrn16 (a) | rrn23 (a) | rrn4.5 (a) | rrn5 (a) | – |
| Transfer RNA genes | trnI-CAU (a) | trnI-GAU (a) | trnL-UAA | trnL-CAA (a) | trnL-UAG |
| trnR-UCU | trnR-ACG (a) | trnA-UGC (a) | trnW-CCA | – | |
| trnV-UAC | trnV-GAC (a) | trnF-GAA | trnT-UGU | trnT-GGU | |
| trnP-UGG | trnfM-CAU | trnP-GGG | trnG-GCC | trnS-GGA | |
| trnS-UGA | trnS-GCU | trnD-GUC | trnC-GCA | trnN-GUU (a) | |
| trnE-UUC | trnY-GUA | trnQ-UUG | trnK-UUU | trnH-GUG | |
| Small subunit of ribosome | rps2 | rps3 | rps4 | rps7 (a) | rps8 |
| rps11 | rps12 (a) | rps14 | rps15 | rps16 | |
| rps18 | rps19 | – | – | – | |
| Large subunit of ribosome | rp12 | rp114 | rp116 | rp120 | rp122 |
| rp123 | rp132 | rp133 | rp136 | – | |
| DNA-dependent RNA polymerase | rpoA | rpoB | rpoC1 | rpoC2 | – |
| Translational initiation factor | infA | – | – | – | – |
| Subunits of photosystem I | psaA | psaB | psaC | psaI | psaJ |
| ycf3 | ycf4 | – | – | – | |
| Subunits of photosystem II | psbB | psbC | psbD | psbE | psbF |
| psbH | psbI | psbJ | psbL | psbM | |
| psbN | psbT | – | – | – | |
| NADH oxidoreductase | ndhA | ndhB (a) | ndhC, | ndhD, | ndhE, |
| ndhG, | ndhI | ndhJ | ndhK | ndhF | |
| Subunits of cytochrome | petA, | petB, | petD | petG | petL |
| petN | – | – | – | – | |
| Subunits of ATP synthase | atpA | atpB | atpE | atpF | atpH |
| atpI | – | – | – | – | |
| Large subunit of Rubisco | rbcL | – | – | – | – |
| Maturase | matk | – | – | – | – |
| Envelope membrane protein | cemA | – | – | – | – |
| Subunit of acetyl-CoA | accD (b) | – | – | – | – |
| C-type cytochrome synthesis gene | ccsA | – | – | – | – |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.-L.; Fan, W.-B.; Wang, N.; Dong, P.-B.; Zhang, T.-T.; Yue, M.; Li, Z.-H. Evolutionary Analysis of Plastid Genomes of Seven Lonicera L. Species: Implications for Sequence Divergence and Phylogenetic Relationships. Int. J. Mol. Sci. 2018, 19, 4039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124039
Liu M-L, Fan W-B, Wang N, Dong P-B, Zhang T-T, Yue M, Li Z-H. Evolutionary Analysis of Plastid Genomes of Seven Lonicera L. Species: Implications for Sequence Divergence and Phylogenetic Relationships. International Journal of Molecular Sciences. 2018; 19(12):4039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124039
Chicago/Turabian StyleLiu, Mi-Li, Wei-Bing Fan, Ning Wang, Peng-Bin Dong, Ting-Ting Zhang, Ming Yue, and Zhong-Hu Li. 2018. "Evolutionary Analysis of Plastid Genomes of Seven Lonicera L. Species: Implications for Sequence Divergence and Phylogenetic Relationships" International Journal of Molecular Sciences 19, no. 12: 4039. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124039