S100A10 and Cancer Hallmarks: Structure, Functions, and its Emerging Role in Ovarian Cancer


Abstract
:1. Introduction
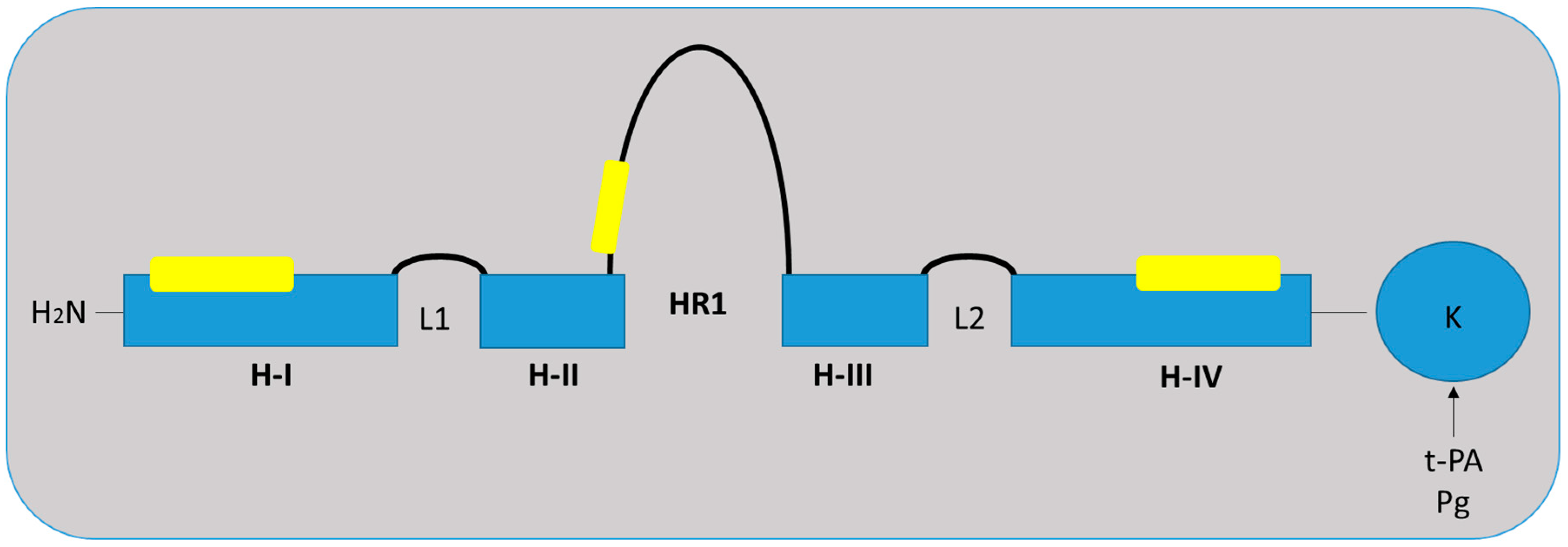
2. S100A10 Structure and Function
3. S100A10 Interaction with Annexin A2
4. Interaction of S100A10 with Other Proteins
5. S100A10 Expression in Cancers
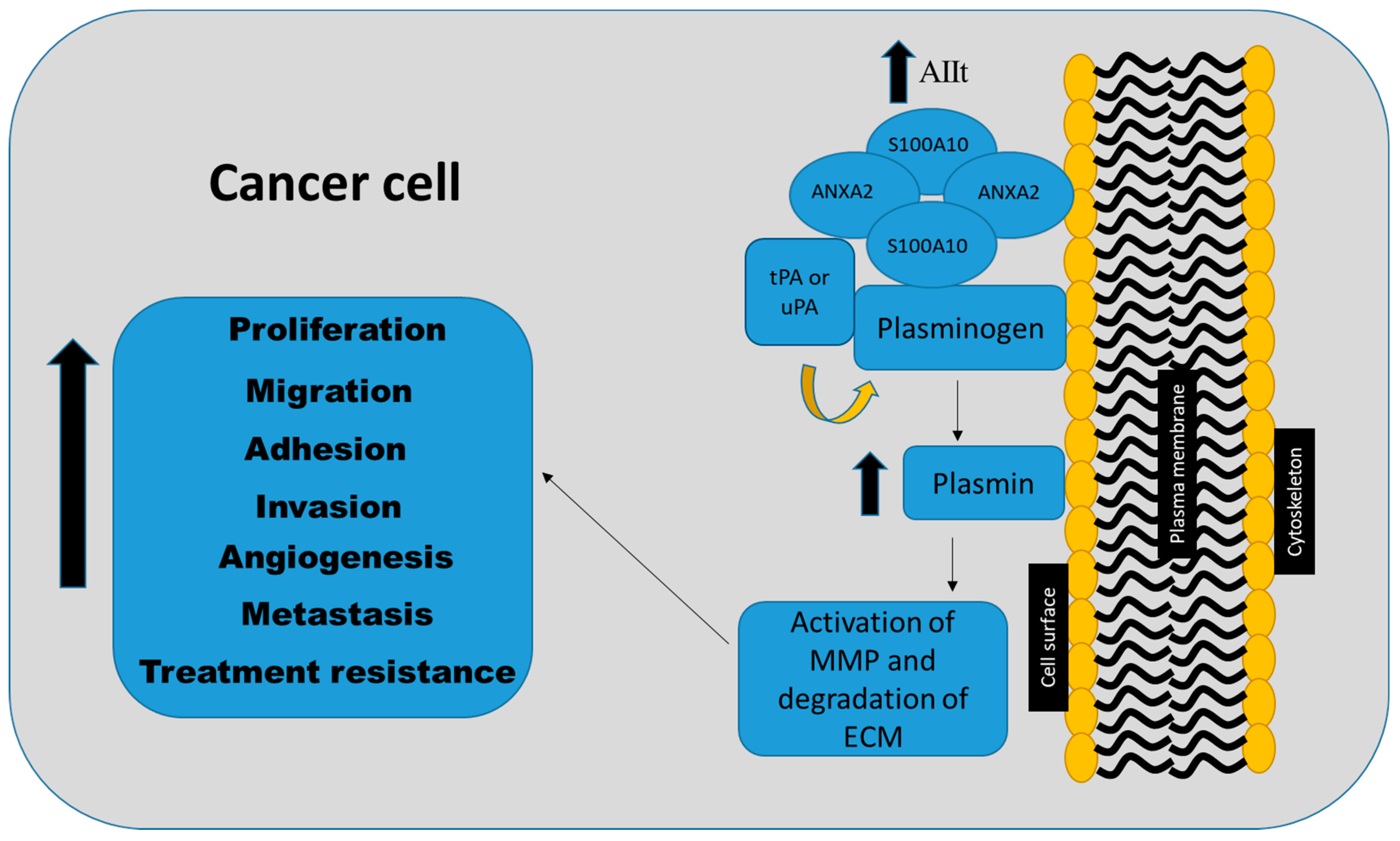
6. Functional Role of S100A10 in Cancer
6.1. Proliferation
6.2. Adhesion
6.3. Migration
6.4. Invasion
6.5. Angiogenesis
6.6. Metastasis
6.7. Therapy Resistance
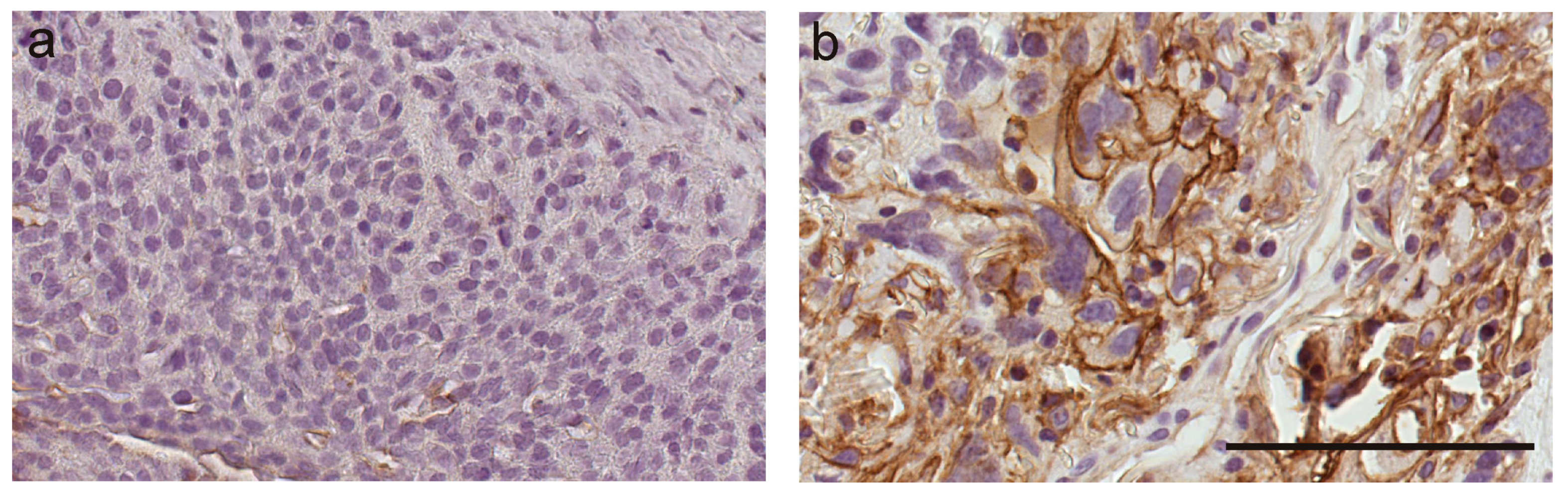
7. Role of S100A10 in Ovarian Cancer and Chemotherapy Resistance
8. Strategies to Target S100A10 in Cancer Cells
9. Summary and Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AIIt | S100A10-annexin A2 heterotetramer |
| ALL | acute lymphoblastic leukemia |
| APL | acute promyelotic leukemia |
| ABC | ATP-binding cassette |
| ATRA | all-trans retinoic acid |
| BAD | Bcl-2-associated death promoter |
| BEAS-2B | bronchial epithelial cells |
| CHO | Chinese hamster ovary |
| CRA | Chemoresponse assay |
| DLCI | data-link connection identifier |
| ECM | extracellular matrix |
| LLC | Lewis Lung Carcinoma |
| MMP | matrix metalloproteinases |
| NSCLC | non-small cell lung cancer |
| PFI | platinum-free interval |
| PFS | progression-free survival |
| Pg | plasminogen |
| PML | promyelocytic leukemia |
| OS | overall survival |
| RhoGAP | Rho GTPase-activating protein |
| t-PA | tissue-type plasminogen activator |
| uPA | urokinase-type plasminogen activator |
References
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Cannistra, S.A. Cancer of the ovary. N. Engl. J. Med. 2004, 351, 2519–2529. [Google Scholar] [CrossRef] [PubMed]
- Colombo, P.E.; Fabbro, M.; Theillet, C.; Bibeau, F.; Rouanet, P.; Ray-Coquard, I. Sensitivity and resistance to treatment in the primary management of epithelial ovarian cancer. Crit. Rev. Oncol. Hematol. 2014, 89, 207–216. [Google Scholar] [CrossRef] [PubMed]
- El Bairi, K.; Amrani, M.; Kandhro, A.H.; Afqir, S. Prediction of therapy response in ovarian cancer: Where are we now? Crit. Rev. Clin. Lab. Sci. 2017, 54, 233–266. [Google Scholar] [CrossRef] [PubMed]
- van Zyl, B.; Tang, D.; Bowden, N.A. Biomarkers of platinum resistance in ovarian cancer: What can we use to improve treatment. Endocr. Relat. Cancer 2018, 25, R303–R318. [Google Scholar] [CrossRef] [PubMed]
- Ween, M.P.; Armstrong, M.A.; Oehler, M.K.; Ricciardelli, C. The role of ABC transporters in ovarian cancer progression and chemoresistance. Crit. Rev. Oncol. Hematol. 2015, 96, 220–256. [Google Scholar] [CrossRef]
- Helleman, J.; Jansen, M.P.; Burger, C.; van der Burg, M.E.; Berns, E.M. Integrated genomics of chemotherapy resistant ovarian cancer: A role for extracellular matrix, TGFbeta and regulating microRNAs. Int. J. Biochem. Cell Biol. 2010, 42, 25–30. [Google Scholar] [CrossRef]
- Chien, J.; Kuang, R.; Landen, C.; Shridhar, V. Platinum-sensitive recurrence in ovarian cancer: The role of tumor microenvironment. Front. Oncol. 2013, 3, 251. [Google Scholar] [CrossRef]
- Rutherford, T.; Orr, J., Jr.; Grendys, E., Jr.; Edwards, R.; Krivak, T.C.; Holloway, R.; Moore, R.G.; Puls, L.; Tillmanns, T.; Schink, J.C.; et al. A prospective study evaluating the clinical relevance of a chemoresponse assay for treatment of patients with persistent or recurrent ovarian cancer. Gynecol. Oncol. 2013, 131, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Sargent, D.J.; Krivak, T.C.; Powell, M.A.; Gabrin, M.J.; Brower, S.L.; Coleman, R.L. Evaluation of a chemoresponse assay as a predictive marker in the treatment of recurrent ovarian cancer: Further analysis of a prospective study. Br. J. Cancer 2014, 111, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Krivak, T.C.; Lele, S.; Richard, S.; Secord, A.A.; Leath, C.A., 3rd; Brower, S.L.; Tian, C.; Moore, R.G. A chemoresponse assay for prediction of platinum resistance in primary ovarian cancer. Am. J. Obstet. Gynecol. 2014, 211, 68.e1–68.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plamadeala, V.; Kelley, J.L.; Chan, J.K.; Krivak, T.C.; Gabrin, M.J.; Brower, S.L.; Powell, M.A.; Rutherford, T.J.; Coleman, R.L. A cost-effectiveness analysis of a chemoresponse assay for treatment of patients with recurrent epithelial ovarian cancer. Gynecol. Oncol. 2015, 136, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, J.P.; Gottesman, M.M. Overcoming multidrug resistance in cancer: 35 years after the discovery of ABCB1. Drug Resist. Updat. 2012, 15, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Nymoen, D.A.; Hetland Falkenthal, T.E.; Holth, A.; Ow, G.S.; Ivshina, A.V.; Trope, C.G.; Kuznetsov, V.A.; Staff, A.C.; Davidson, B. Expression and clinical role of chemoresponse-associated genes in ovarian serous carcinoma. Gynecol. Oncol. 2015, 139, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Lokman, N.A.; Pyragius, C.E.; Ruszkiewicz, A.; Oehler, M.K.; Ricciardelli, C. Annexin A2 and S100A10 are independent predictors of serous ovarian cancer outcome. Transl. Res. 2016, 171, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Gerke, V. S100A10/p11: Family, friends and functions. Pflugers Arch. 2008, 455, 575–582. [Google Scholar] [CrossRef]
- Chen, H.; Xu, C.; Jin, Q.; Liu, Z. S100 protein family in human cancer. Am. J. Cancer Res. 2014, 4, 89–115. [Google Scholar]
- Bresnick, A.R.; Weber, D.J.; Zimmer, D.B. S100 proteins in cancer. Nat. Rev. Cancer 2015, 15, 96–109. [Google Scholar] [CrossRef] [Green Version]
- Santamaria-Kisiel, L.; Rintala-Dempsey, A.C.; Shaw, G.S. Calcium-dependent and -independent interactions of the S100 protein family. Biochem. J. 2006, 396, 201–214. [Google Scholar] [CrossRef]
- Liu, Y.; Myrvang, H.K.; Dekker, L.V. Annexin A2 complexes with S100 proteins: Structure, function and pharmacological manipulation. Br. J. Pharmacol. 2015, 172, 1664–1676. [Google Scholar] [CrossRef] [PubMed]
- Surette, A.P.; Madureira, P.A.; Phipps, K.D.; Miller, V.A.; Svenningsson, P.; Waisman, D.M. Regulation of fibrinolysis by S100A10 in vivo. Blood 2011, 118, 3172–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madureira, P.A.; Surette, A.P.; Phipps, K.D.; Taboski, M.A.; Miller, V.A.; Waisman, D.M. The role of the annexin A2 heterotetramer in vascular fibrinolysis. Blood 2011, 118, 4789–4797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, P.A.; Surette, A.P.; Liwski, R.S.; Svenningsson, P.; Waisman, D.M. S100A10 regulates plasminogen-dependent macrophage invasion. Blood 2010, 116, 1136–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessner, F.; Dlugos, C.P.; Chehab, T.; Schaefer, C.; Homey, B.; Gerke, V.; Weide, T.; Pavenstadt, H.; Rescher, U. CC chemokine receptor 10 cell surface presentation in melanocytes is regulated by the novel interaction partner S100A10. Sci. Rep. 2016, 6, 22649. [Google Scholar] [CrossRef] [PubMed]
- Bissonnette, L.; Drissennek, L.; Antoine, Y.; Tiers, L.; Hirtz, C.; Lehmann, S.; Perrochia, H.; Bissonnette, F.; Kadoch, I.J.; Haouzi, D.; et al. Human S100A10 plays a crucial role in the acquisition of the endometrial receptivity phenotype. Cell Adhes. Migr. 2016, 10, 282–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teijeiro, J.M.; Roldan, M.L.; Marini, P.E. Annexin A2 and S100A10 in the mammalian oviduct. Cell Tissue Res. 2016, 363, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aleem, S.A.; Dekker, L.V. Assessment of the cellular localisation of the annexin A2/S100A10 complex in human placenta. J. Mol. Histol. 2018, 49, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Kim, Y.; Warner-Schmidt, J.; Oh, Y.S.; Greengard, P. p11 and its role in depression and therapeutic responses to antidepressants. Nat. Rev. Neurosci. 2013, 14, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.W.; Westin, L.; Kim, J.; Chang, J.C.; Oh, Y.S.; Amreen, B.; Gresack, J.; Flajolet, M.; Kim, D.; Aperia, A.; et al. Alteration by p11 of mGluR5 localization regulates depression-like behaviors. Mol. Psychiatry 2015, 20, 1546–1556. [Google Scholar] [CrossRef] [Green Version]
- Meschin, P.; Demion, M.; Cazorla, O.; Finan, A.; Thireau, J.; Richard, S.; Lacampagne, A. p11 modulates calcium handling through 5-HT(4)R pathway in rat ventricular cardiomyocytes. Cell Calcium. 2015, 58, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Madureira, P.A.; O’Connell, P.A.; Surette, A.P.; Miller, V.A.; Waisman, D.M. The biochemistry and regulation of S100A10: A multifunctional plasminogen receptor involved in oncogenesis. J. Biomed. Biotechnol. 2012, 2012, 353687. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, P.A.; Waisman, D.M. Regulation of plasmin generation by the annexin A2 heterotetramer: A shift in perspective. Future Oncol. 2012, 8, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Deora, A.B.; Kreitzer, G.; Jacovina, A.T.; Hajjar, K.A. An annexin 2 phosphorylation switch mediates p11-dependent translocation of annexin 2 to the cell surface. J. Biol. Chem. 2004, 279, 43411–43418. [Google Scholar] [CrossRef]
- Lokman, N.A.; Ween, M.P.; Oehler, M.K.; Ricciardelli, C. The role of annexin A2 in tumorigenesis and cancer progression. Cancer Microenviron. 2011, 4, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, A.; Bydoun, M.; Holloway, R.; Waisman, D. Annexin A2 heterotetramer: Structure and function. Int. J. Mol. Sci. 2013, 14, 6259–6305. [Google Scholar] [CrossRef]
- Kumari, S.; Malla, R. New Insight on the Role of Plasminogen Receptor in Cancer Progression. Cancer Growth Metastasis 2015, 8, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Cell surface remodeling by plasmin: A new function for an old enzyme. J. Biomed. Biotechnol. 2012, 2012, 564259. [Google Scholar] [CrossRef]
- Rety, S.; Sopkova, J.; Renouard, M.; Osterloh, D.; Gerke, V.; Tabaries, S.; Russo-Marie, F.; Lewit-Bentley, A. The crystal structure of a complex of p11 with the annexin II N-terminal peptide. Nat. Struct. Biol. 1999, 6, 89–95. [Google Scholar]
- Semov, A.; Moreno, M.J.; Onichtchenko, A.; Abulrob, A.; Ball, M.; Ekiel, I.; Pietrzynski, G.; Stanimirovic, D.; Alakhov, V. Metastasis-associated protein S100A4 induces angiogenesis through interaction with Annexin II and accelerated plasmin formation. J. Biol. Chem. 2005, 280, 20833–20841. [Google Scholar] [CrossRef]
- He, K.L.; Deora, A.B.; Xiong, H.; Ling, Q.; Weksler, B.B.; Niesvizky, R.; Hajjar, K.A. Endothelial cell annexin A2 regulates polyubiquitination and degradation of its binding partner S100A10/p11. J. Biol. Chem. 2008, 283, 19192–19200. [Google Scholar] [CrossRef] [PubMed]
- Valapala, M.; Vishwanatha, J.K. Lipid raft endocytosis and exosomal transport facilitate extracellular trafficking of annexin A2. J. Biol. Chem. 2011, 286, 30911–30925. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Foley, K.; Huang, L.; Leubner, A.; Mo, G.; Olino, K.; Edil, B.H.; Mizuma, M.; Sharma, R.; Le, D.T.; et al. Tyrosine 23 phosphorylation-dependent cell-surface localization of annexin A2 is required for invasion and metastases of pancreatic cancer. PLoS ONE 2011, 6, e19390. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.T.; Lin, C.F.; Wang, C.Y.; Anderson, R.; Lin, Y.S. Interferon-gamma stimulates p11-dependent surface expression of annexin A2 in lung epithelial cells to enhance phagocytosis. J. Cell Physiol. 2012, 227, 2775–2787. [Google Scholar] [CrossRef] [PubMed]
- Hajjar, K.A.; Krishnan, S. Annexin II: A mediator of the plasmin/plasminogen activator system. Trends Cardiovasc. Med. 1999, 9, 128–138. [Google Scholar] [CrossRef]
- O’Connell, P.A.; Madureira, P.A.; Berman, J.N.; Liwski, R.S.; Waisman, D.M. Regulation of S100A10 by the PML-RAR-alpha oncoprotein. Blood 2011, 117, 4095–4105. [Google Scholar] [CrossRef] [PubMed]
- Bydoun, M.; Waisman, D.M. On the contribution of S100A10 and annexin A2 to plasminogen activation and oncogenesis: An enduring ambiguity. Future Oncol. 2014, 10, 2469–2479. [Google Scholar] [CrossRef]
- Kwon, M.; MacLeod, T.J.; Zhang, Y.; Waisman, D.M. S100A10, annexin A2, and annexin a2 heterotetramer as candidate plasminogen receptors. Front. Biosci. 2005, 10, 300–325. [Google Scholar] [CrossRef]
- Hou, Y.; Yang, L.; Mou, M.; Hou, Y.; Zhang, A.; Pan, N.; Qiang, R.; Wei, L.; Zhang, N. Annexin A2 regulates the levels of plasmin, S100A10 and Fascin in L5178Y cells. Cancer Investig. 2008, 26, 809–815. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, B.; Zhang, Y.; Cao, J.; Chen, T. Silencing of the annexin II gene down-regulates the levels of S100A10, c-Myc, and plasmin and inhibits breast cancer cell proliferation and invasion. Saudi Med. J. 2010, 31, 374–381. [Google Scholar]
- Suzuki, S.; Tanigawara, Y. Forced expression of S100A10 reduces sensitivity to oxaliplatin in colorectal cancer cells. Proteome Sci. 2014, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Okuse, K.; Malik-Hall, M.; Baker, M.D.; Poon, W.Y.; Kong, H.; Chao, M.V.; Wood, J.N. Annexin II light chain regulates sensory neuron-specific sodium channel expression. Nature 2002, 417, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Girard, C.; Tinel, N.; Terrenoire, C.; Romey, G.; Lazdunski, M.; Borsotto, M. p11, an annexin II subunit, an auxiliary protein associated with the background K+ channel, TASK-1. EMBO J. 2002, 21, 4439–4448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Graaf, S.F.; Hoenderop, J.G.; Gkika, D.; Lamers, D.; Prenen, J.; Rescher, U.; Gerke, V.; Staub, O.; Nilius, B.; Bindels, R.J. Functional expression of the epithelial Ca2+ channels (TRPV5 and TRPV6) requires association of the S100A10-annexin 2 complex. EMBO J. 2003, 22, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Donier, E.; Rugiero, F.; Okuse, K.; Wood, J.N. Annexin II light chain p11 promotes functional expression of acid-sensing ion channel ASIC1a. J. Biol. Chem. 2005, 280, 38666–38672. [Google Scholar] [CrossRef]
- Svenningsson, P.; Chergui, K.; Rachleff, I.; Flajolet, M.; Zhang, X.; El Yacoubi, M.; Vaugeois, J.M.; Nomikos, G.G.; Greengard, P. Alterations in 5-HT1B receptor function by p11 in depression-like states. Science 2006, 311, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Rezvanpour, A.; Santamaria-Kisiel, L.; Shaw, G.S. The S100A10-annexin A2 complex provides a novel asymmetric platform for membrane repair. J. Biol. Chem. 2011, 286, 40174–40183. [Google Scholar] [CrossRef] [PubMed]
- Ozorowski, G.; Milton, S.; Luecke, H. Structure of a C-terminal AHNAK peptide in a 1:2:2 complex with S100A10 and an acetylated N-terminal peptide of annexin A2. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 92–104. [Google Scholar] [CrossRef]
- Oh, Y.S.; Gao, P.; Lee, K.W.; Ceglia, I.; Seo, J.S.; Zhang, X.; Ahn, J.H.; Chait, B.T.; Patel, D.J.; Kim, Y.; et al. SMARCA3, a chromatin-remodeling factor, is required for p11-dependent antidepressant action. Cell 2013, 152, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, S.; Asano, E.; Ito, S.; Ohno, K.; Hamaguchi, M.; Senga, T. S100A10 is required for the organization of actin stress fibers and promotion of cell spreading. Mol. Cell. Biochem. 2013, 374, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Degos, L.; Wang, Z.Y. All trans retinoic acid in acute promyelocytic leukemia. Oncogene 2001, 20, 7140–7145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menell, J.S.; Cesarman, G.M.; Jacovina, A.T.; McLaughlin, M.A.; Lev, E.A.; Hajjar, K.A. Annexin II and bleeding in acute promyelocytic leukemia. N. Engl. J. Med. 1999, 340, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yang, Y.; Sun, J.; Dong, X.; Wang, J.; Liu, H.; Lu, C.; Chen, X.; Shao, J.; Yan, J. Annexin A2-S100A10 heterotetramer is upregulated by PML/RARalpha fusion protein and promotes plasminogen-dependent fibrinolysis and matrix invasion in acute promyelocytic leukemia. Front. Med. 2017, 11, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnapillai, A.; Kolb, E.A.; Dhanan, P.; Mason, R.W.; Napper, A.; Barwe, S.P. Disruption of Annexin II /p11 Interaction Suppresses Leukemia Cell Binding, Homing and Engraftment, and Sensitizes the Leukemia Cells to Chemotherapy. PLoS ONE 2015, 10, e0140564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhu, M.; Cheng, W.; Xing, R.; Li, W.; Zhao, M.; Xu, L.; Li, E.; Luo, G.; Lu, Y. Downregulation of 425G>a variant of calcium-binding protein S100A14 associated with poor differentiation and prognosis in gastric cancer. J. Cancer Res. Clin. Oncol. 2015, 141, 691–703. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Fei, F.; Qu, J.; Wang, X.; Zhao, Y.; Li, Y.; Zhang, S. iTRAQ-based proteomic analysis of DMH-induced colorectal cancer in mice reveals the expressions of beta-catenin, decorin, septin-7, and S100A10 expression in 53 cases of human hereditary polyposis colorectal cancer. Clin. Transl. Oncol. 2018. [Google Scholar] [CrossRef]
- Li, C.M.; Hou, Y.H.; Zhang, J.; Zhang, L. The expressions and roles of S100A6 and S100A10 in gastric cancer. Biomed. Res. India 2017, 28, 2131–2138. [Google Scholar]
- Rust, R.; Visser, L.; van der Leij, J.; Harms, G.; Blokzijl, T.; Deloulme, J.C.; van der Vlies, P.; Kamps, W.; Kok, K.; Lim, M.; et al. High expression of calcium-binding proteins, S100A10, S100A11 and CALM2 in anaplastic large cell lymphoma. Br. J. Haematol. 2005, 131, 596–608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tingfeng, X.; Fuqiang, P.; Dong, L. Expression and clinical significance of S100 family genes in patients with melanoma. Melanoma Res. 2018. [Google Scholar] [CrossRef]
- McKiernan, E.; McDermott, E.W.; Evoy, D.; Crown, J.; Duffy, M.J. The role of S100 genes in breast cancer progression. Tumour Biol. 2011, 32, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Domoto, T.; Miyama, Y.; Suzuki, H.; Teratani, T.; Arai, K.; Sugiyama, T.; Takayama, T.; Mugiya, S.; Ozono, S.; Nozawa, R. Evaluation of S100A10, annexin II and B-FABP expression as markers for renal cell carcinoma. Cancer Sci. 2007, 98, 77–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teratani, T.; Watanabe, T.; Kuwahara, F.; Kumagai, H.; Kobayashi, S.; Aoki, U.; Ishikawa, A.; Arai, K.; Nozawa, R. Induced transcriptional expression of calcium-binding protein S100A1 and S100A10 genes in human renal cell carcinoma. Cancer Lett. 2002, 175, 71–77. [Google Scholar] [CrossRef]
- El-Rifai, W.; Moskaluk, C.A.; Abdrabbo, M.K.; Harper, J.; Yoshida, C.; Riggins, G.J.; Frierson, H.F., Jr.; Powell, S.M. Gastric cancers overexpress S100A calcium-binding proteins. Cancer Res. 2002, 62, 6823–6826. [Google Scholar] [PubMed]
- Chetcuti, A.; Margan, S.H.; Russell, P.; Mann, S.; Millar, D.S.; Clark, S.J.; Rogers, J.; Handelsman, D.J.; Dong, Q. Loss of annexin II heavy and light chains in prostate cancer and its precursors. Cancer Res. 2001, 61, 6331–6334. [Google Scholar] [PubMed]
- Martinez-Aguilar, J.; Clifton-Bligh, R.; Molloy, M.P. A multiplexed, targeted mass spectrometry assay of the S100 protein family uncovers the isoform-specific expression in thyroid tumours. BMC Cancer 2015, 15, 199. [Google Scholar] [CrossRef]
- Ito, Y.; Arai, K.; Nozawa, R.; Yoshida, H.; Higashiyama, T.; Takamura, Y.; Miya, A.; Kobayashi, K.; Kuma, K.; Miyauchi, A. S100A10 expression in thyroid neoplasms originating from the follicular epithelium: Contribution to the aggressive characteristic of anaplastic carcinoma. Anticancer Res. 2007, 27, 2679–2683. [Google Scholar]
- Tan, Y.; Ma, S.Y.; Wang, F.Q.; Meng, H.P.; Mei, C.Z.; Liu, A.G.; Wu, H.R. Proteomic-based analysis for identification of potential serum biomarkers in gallbladder cancer. Oncol. Rep. 2011, 26, 853–859. [Google Scholar]
- Sato, K.; Saiki, Y.; Arai, K.; Ishizawa, K.; Fukushige, S.; Aoki, K.; Abe, J.; Takahashi, S.; Sato, I.; Sakurada, A.; et al. S100A10 upregulation associates with poor prognosis in lung squamous cell carcinoma. Biochem. Biophys. Res. Commun. 2018, 505, 466–470. [Google Scholar] [CrossRef]
- Katono, K.; Sato, Y.; Jiang, S.X.; Kobayashi, M.; Saito, K.; Nagashio, R.; Ryuge, S.; Satoh, Y.; Saegusa, M.; Masuda, N. Clinicopathological Significance of S100A10 Expression in Lung Adenocarcinomas. Asian Pac. J. Cancer Prev. 2016, 17, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Bydoun, M.; Sterea, A.; Liptay, H.; Uzans, A.; Huang, W.Y.; Rodrigues, G.J.; Weaver, I.C.G.; Gu, H.; Waisman, D.M. S100A10, a novel biomarker in pancreatic ductal adenocarcinoma. Mol. Oncol. 2018, 12, 1895–1916. [Google Scholar] [CrossRef]
- Gocheva, V.; Naba, A.; Bhutkar, A.; Guardia, T.; Miller, K.M.; Li, C.M.; Dayton, T.L.; Sanchez-Rivera, F.J.; Kim-Kiselak, C.; Jailkhani, N.; et al. Quantitative proteomics identify Tenascin-C as a promoter of lung cancer progression and contributor to a signature prognostic of patient survival. Proc. Natl. Acad. Sci. USA 2017, 114, E5625–E5634. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Zhang, Z.; Song, W.; Zhou, B.; Zhang, Y.; Li, G.; Qiu, S. S100A10 as a novel biomarker in colorectal cancer. Tumour Biol. 2013, 34, 3785–3790. [Google Scholar] [CrossRef] [PubMed]
- Giraldez, M.D.; Lozano, J.J.; Cuatrecasas, M.; Alonso-Espinaco, V.; Maurel, J.; Marmol, M.; Horndler, C.; Ortego, J.; Alonso, V.; Escudero, P.; et al. Gene-expression signature of tumor recurrence in patients with stage II and III colon cancer treated with 5′fluoruracil-based adjuvant chemotherapy. Int. J. Cancer 2013, 132, 1090–1097. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cui, J.; Tang, Y.L.; Huang, L.; Zhou, C.Y.; Xu, J.X. Prognostic Roles of mRNA Expression of S100 in Non-Small-Cell Lung Cancer. BioMed Res. Int. 2018, 2018, 9815806. [Google Scholar] [CrossRef] [PubMed]
- Hatoum, D.; Yagoub, D.; Ahadi, A.; Nassif, N.T.; McGowan, E.M. Annexin/S100A Protein Family Regulation through p14ARF-p53 Activation: A Role in Cell Survival and Predicting Treatment Outcomes in Breast Cancer. PLoS ONE 2017, 12, e0169925. [Google Scholar] [CrossRef] [PubMed]
- Phipps, K.D.; Surette, A.P.; O’Connell, P.A.; Waisman, D.M. Plasminogen receptor S100A10 is essential for the migration of tumor-promoting macrophages into tumor sites. Cancer Res. 2011, 71, 6676–6683. [Google Scholar] [CrossRef]
- Yang, X.; Popescu, N.C.; Zimonjic, D.B. DLC1 interaction with S100A10 mediates inhibition of in vitro cell invasion and tumorigenicity of lung cancer cells through a RhoGAP-independent mechanism. Cancer Res. 2011, 71, 2916–2925. [Google Scholar] [CrossRef]
- Shan, X.; Miao, Y.; Fan, R.; Qian, H.; Chen, P.; Liu, H.; Yan, X.; Li, J.; Zhou, F. MiR-590-5P inhibits growth of HepG2 cells via decrease of S100A10 expression and Inhibition of the Wnt pathway. Int. J. Mol. Sci. 2013, 14, 8556–8569. [Google Scholar] [CrossRef]
- D’Souza, S.; Kurihara, N.; Shiozawa, Y.; Joseph, J.; Taichman, R.; Galson, D.L.; Roodman, G.D. Annexin II interactions with the annexin II receptor enhance multiple myeloma cell adhesion and growth in the bone marrow microenvironment. Blood 2012, 119, 1888–1896. [Google Scholar] [CrossRef] [Green Version]
- Shiozawa, Y.; Havens, A.M.; Jung, Y.; Ziegler, A.M.; Pedersen, E.A.; Wang, J.; Wang, J.; Lu, G.; Roodman, G.D.; Loberg, R.D.; et al. Annexin II/annexin II receptor axis regulates adhesion, migration, homing, and growth of prostate cancer. J. Cell. Biochem. 2008, 105, 370–380. [Google Scholar] [CrossRef]
- Myrvang, H.K.; Guo, X.; Li, C.; Dekker, L.V. Protein interactions between surface annexin A2 and S100A10 mediate adhesion of breast cancer cells to microvascular endothelial cells. FEBS Lett. 2013, 587, 3210–3215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Fogg, D.K.; Waisman, D.M. RNA interference-mediated silencing of the S100A10 gene attenuates plasmin generation and invasiveness of Colo 222 colorectal cancer cells. J. Biol. Chem. 2004, 279, 2053–2062. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.S.; Fogg, D.K.; Yoon, C.S.; Waisman, D.M. p11 regulates extracellular plasmin production and invasiveness of HT1080 fibrosarcoma cells. FASEB J. 2003, 17, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Madureira, P.A.; Bharadwaj, A.G.; Bydoun, M.; Garant, K.; O’Connell, P.; Lee, P.; Waisman, D.M. Cell surface protease activation during RAS transformation: Critical role of the plasminogen receptor, S100A10. Oncotarget 2016, 7, 47720–47737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Bardia, A.; Wittner, B.S.; Stott, S.L.; Smas, M.E.; Ting, D.T.; Isakoff, S.J.; Ciciliano, J.C.; Wells, M.N.; Shah, A.M.; et al. Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science 2013, 339, 580–584. [Google Scholar] [CrossRef]
- Spijkers-Hagelstein, J.A.; Mimoso Pinhancos, S.; Schneider, P.; Pieters, R.; Stam, R.W. Src kinase-induced phosphorylation of annexin A2 mediates glucocorticoid resistance in MLL-rearranged infant acute lymphoblastic leukemia. Leukemia 2013, 27, 1063–1071. [Google Scholar] [CrossRef]
- Browne, B.C.; Hochgrafe, F.; Wu, J.; Millar, E.K.; Barraclough, J.; Stone, A.; McCloy, R.A.; Lee, C.S.; Roberts, C.; Ali, N.A.; et al. Global characterization of signalling networks associated with tamoxifen resistance in breast cancer. FEBS J. 2013, 280, 5237–5257. [Google Scholar] [CrossRef] [Green Version]
- Johansson, H.J.; Sanchez, B.C.; Forshed, J.; Stal, O.; Fohlin, H.; Lewensohn, R.; Hall, P.; Bergh, J.; Lehtio, J.; Linderholm, B.K. Proteomics profiling identify CAPS as a potential predictive marker of tamoxifen resistance in estrogen receptor positive breast cancer. Clin. Proteom. 2015, 12, 8. [Google Scholar] [CrossRef] [Green Version]
- Hsu, S.Y.; Kaipia, A.; Zhu, L.; Hsueh, A.J. Interference of BAD (Bcl-xL/Bcl-2-associated death promoter)-induced apoptosis in mammalian cells by 14-3-3 isoforms and P11. Mol. Endocrinol. 1997, 11, 1858–1867. [Google Scholar]
- Gillet, J.P.; Calcagno, A.M.; Varma, S.; Davidson, B.; Bunkholt Elstrand, M.; Ganapathi, R.; Kamat, A.A.; Sood, A.K.; Ambudkar, S.V.; Seiden, M.V.; et al. Multidrug resistance-linked gene signature predicts overall survival of patients with primary ovarian serous carcinoma. Clin. Cancer Res. 2012, 18, 3197–3206. [Google Scholar] [CrossRef]
- Li, C.; Reddy, T.R.; Fischer, P.M.; Dekker, L.V. A Cy5-labeled S100A10 tracer used to identify inhibitors of the protein interaction with annexin A2. Assay Drug Dev. Technol. 2010, 8, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.R.; Li, C.; Guo, X.; Myrvang, H.K.; Fischer, P.M.; Dekker, L.V. Design, synthesis, and structure-activity relationship exploration of 1-substituted 4-aroyl-3-hydroxy-5-phenyl-1H-pyrrol-2(5H)-one analogues as inhibitors of the annexin A2-S100A10 protein interaction. J. Med. Chem. 2011, 54, 2080–2094. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.R.; Li, C.; Fischer, P.M.; Dekker, L.V. Three-dimensional pharmacophore design and biochemical screening identifies substituted 1,2,4-triazoles as inhibitors of the annexin A2-S100A10 protein interaction. ChemMedChem 2012, 7, 1435–1446. [Google Scholar] [CrossRef] [PubMed]
- Reddy, T.R.; Li, C.; Guo, X.; Fischer, P.M.; Dekker, L.V. Design, synthesis and SAR exploration of tri-substituted 1,2,4-triazoles as inhibitors of the annexin A2-S100A10 protein interaction. Bioorg. Med. Chem. 2014, 22, 5378–5391. [Google Scholar] [CrossRef]
- Lo-Coco, F.; Avvisati, G.; Vignetti, M.; Thiede, C.; Orlando, S.M.; Iacobelli, S.; Ferrara, F.; Fazi, P.; Cicconi, L.; Di Bona, E.; et al. Retinoic acid and arsenic trioxide for acute promyelocytic leukemia. N. Engl. J. Med. 2013, 369, 111–121. [Google Scholar] [CrossRef]
- Olwill, S.A.; McGlynn, H.; Gilmore, W.S.; Alexander, H.D. All-trans retinoic acid-induced downregulation of annexin II expression in myeloid leukaemia cell lines is not confined to acute promyelocytic leukaemia. Br. J. Haematol. 2005, 131, 258–264. [Google Scholar] [CrossRef] [Green Version]
- Olwill, S.A.; McGlynn, H.; Gilmore, W.S.; Alexander, H.D. Annexin II cell surface and mRNA expression in human acute myeloid leukaemia cell lines. Thromb. Res. 2005, 115, 109–114. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, H.; Wang, J.; Yang, L.; Hu, Y.; Shen, G.; Guo, P.; Qiao, Z.; Song, S. Arsenic trioxide, retinoic acid and Ara-c regulated the expression of annexin II on the surface of APL cells, a novel co-receptor for plasminogen/tissue plasminogen activator. Thromb. Res. 2002, 106, 63–70. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Jiang, M.; Dai, L.; Zhang, W.; Wu, D.; Ruan, C. The expression of annexin II and its role in the fibrinolytic activity in acute promyelocytic leukemia. Leuk. Res. 2011, 35, 879–884. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Yao, X.L.; Cowan, M.; Huang, X.L.; Schneider, R.; Grant, L.R.; Logun, C.; Shelhamer, J.H. Retinoic acid reduces p11 protein levels in bronchial epithelial cells by a posttranslational mechanism. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L1103–L1109. [Google Scholar] [CrossRef]
- Holloway, R.W.; Thomas, M.L.; Cohen, A.M.; Bharadwaj, A.G.; Rahman, M.; Marcato, P.; Marignani, P.A.; Waisman, D.M. Regulation of cell surface protease receptor S100A10 by retinoic acid therapy in acute promyelocytic leukemia (APL). Cell Death Dis. 2018, 9, 920. [Google Scholar] [CrossRef] [PubMed]
- Purev, E.; Soprano, D.R.; Soprano, K.J. PP2A interaction with Rb2/p130 mediates translocation of Rb2/p130 into the nucleus in all-trans retinoic acid-treated ovarian carcinoma cells. J. Cell. Physiol. 2011, 226, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Ricciardelli, C.; Lokman, N.A.; Sabit, I.; Gunasegaran, K.; Bonner, W.M.; Pyragius, C.E.; Macpherson, A.M.; Oehler, M.K. Novel ex vivo ovarian cancer tissue explant assay for prediction of chemosensitivity and response to novel therapeutics. Cancer Lett. 2018, 421, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Escudier, S.M.; Kantarjian, H.M.; Estey, E.H. Thrombosis in patients with acute promyelocytic leukemia treated with and without all-trans retinoic acid. Leuk. Lymphoma 1996, 20, 435–439. [Google Scholar] [CrossRef] [PubMed]
- de-Medeiros, B.C.; Strapasson, E.; Pasquini, R.; de-Medeiros, C.R. Effect of all-trans retinoic acid on newly diagnosed acute promyelocytic leukemia patients: Results of a Brazilian center. Braz. J. Med. Biol. Res. 1998, 31, 1537–1543. [Google Scholar] [CrossRef] [PubMed]
- Mandelli, F.; Diverio, D.; Avvisati, G.; Luciano, A.; Barbui, T.; Bernasconi, C.; Broccia, G.; Cerri, R.; Falda, M.; Fioritoni, G.; et al. Molecular remission in PML/RAR alpha-positive acute promyelocytic leukemia by combined all-trans retinoic acid and idarubicin (AIDA) therapy. Gruppo Italiano-Malattie Ematologiche Maligne dell’Adulto and Associazione Italiana di Ematologia ed Oncologia Pediatrica Cooperative Groups. Blood 1997, 90, 1014–1021. [Google Scholar] [PubMed]
- Tallman, M.S.; Andersen, J.W.; Schiffer, C.A.; Appelbaum, F.R.; Feusner, J.H.; Ogden, A.; Shepherd, L.; Willman, C.; Bloomfield, C.D.; Rowe, J.M.; et al. All-trans-retinoic acid in acute promyelocytic leukemia. N. Engl. J. Med. 1997, 337, 1021–1028. [Google Scholar] [CrossRef]
- De Stefano, V.; Sora, F.; Rossi, E.; Chiusolo, P.; Laurenti, L.; Fianchi, L.; Zini, G.; Pagano, L.; Sica, S.; Leone, G. The risk of thrombosis in patients with acute leukemia: Occurrence of thrombosis at diagnosis and during treatment. J. Thromb. Haemost. 2005, 3, 1985–1992. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cancer | S100A10 Expression | Ref. |
|---|---|---|
| Blood | S100A10 expression is increased on the surface of leukemia cells | [46,63] |
| S100A10 mRNA and protein is upregulated in B-cell acute lymphoblastic leukemia | [64] | |
| Breast | S100A10 expression is upregulated in basal-type breast cancer | [70] |
| Colorectal | S100A10 is increased in hereditary polyposis colorectal cancer | [66] |
| Increased S100A10 expression is associated with poor prognosis and reduced overall survival in colorectal cancer | [82] | |
| S100A10 gene expression is associated with tumor recurrence in colon cancer | [83] | |
| Gallbladder | High cytoplasmic S100A10 expression is associated with poor prognosis | [77] |
| Kidney | S100A10 expression is 2.5-fold higher in renal cell carcinoma compared with normal kidney tissue | [71] |
| S100A10 is expressed in renal cell carcinoma and absent in non-cancerous renal tumors | [72] | |
| Lung | Overexpression of S100A10 is associated with poor prognosis | [78,79,84] |
| Lymphatic | S100A10 is overexpressed in anaplastic large cell lymphoma | [68] |
| Pancreas | S100A10 mRNA and protein is overexpressed in pancreatic cancer and predicts patient outcome | [80] |
| Prostate | S100A10 expression is lost in prostate cancer tissues | [74] |
| Skin | S100A10 expression is increased in melanoma compared with normal skin | [69] |
| Stomach | S100A10 is overexpressed in gastric cancer | [65,73] |
| S100A10 expression is upregulated in pre-cancerous lesions and associated with gastric cancer metastasis to the lymph node | [67] | |
| Thyroid | S100A10 is overexpressed in anaplastic thyroid carcinomas compared with normal tissues | [76] |
| S100A10 expression is decreased in follicular adenomas and thyroid carcinomas | [75] |
| Function | Observation | Ref. |
|---|---|---|
| Proliferation | S100A10 is upregulated by p53 activation in breast cancer cells | [85] |
| Lewis Lung carcinoma and T241 fibrosarcoma proliferation is inhibited in S100A10 knockout mice | [86] | |
| S100A10 is downregulated by a knockdown of annexin A2, which decreases the proliferation of breast cancer cell lines | [50] | |
| The displacement of S100A10 from annexin A2 attenuates plasminogen activation, impairing colony formation and growth of lung cancer cells | [87] | |
| Cell growth inhibition by the microRNA miR-590-5P in hepatic carcinoma cells is mediated via S100A10 | [88] | |
| Annexin A2/S100A10 activates the ERK1/2 and AKT pathways in multiple myeloma cells to enhance cell growth | [89] | |
| S100A10 knockdown reduces proliferation of colon cancer cells | [82] | |
| S100A10 knockdown inhibits growth of pancreatic cancer cells PANC-1 in immunocompromised NOD/SCID mice | [80] | |
| Adhesion | Annexin A2/S100A10 regulates adhesion of leukemia cells and prostate cancer cells to osteoblasts | [64,90] |
| Cell surface S100A10 expression promotes adhesion of breast and prostate cancer cells to endothelial cells in vitro | [90,91] | |
| Migration | S100A10 expression is associated with the recruitment and migration of macrophages | [24,70,86] |
| The displacement of S100A10 from annexin A2 attenuates plasminogen activation and impairs the migration of A549 lung cancer cells | [87] | |
| S100A10 knockdown reduces the migration of colon cancer cells | [82] | |
| Invasion | S100A10 in colon cancer cells is crucial for promoting plasmin production and cell invasiveness | [92] |
| S100A10 antibodies inhibit the invasion of acute promyelocytic leukemia cells | [63] | |
| S100A10 expression in fibrosarcoma cells increases plasmin production and cell invasiveness | [93] | |
| The displacement of S100A10 from annexin A2 attenuates plasminogen activation and impairs invasion of lung cancer cells | [87] | |
| Macrophages from S100A10 knockout mice have reduced plasmin-dependent invasion | [24] | |
| S100A10 depletion in RAS-transformed cell lines (HEK293, NIH-3T3) results in a loss of plasmin production and reduced cell invasiveness | [94] | |
| siRNA S100A10 reduces invasion of HCT-116 and DLD-1 colon cancer cell lines | [82] | |
| Angiogenesis | S100A10-null mice have reduced blood vessel density compared to wild-type mice | [86] |
| Metastasis | Loss of S100A10 reduces metastatic burden in the HT1080 fibrosarcoma mouse model | [93] |
| Overexpression of S100A10 increases the metastatic burden in the HT1080 fibrosarcoma mouse model | [93] | |
| S100A10 is one of 170 genes activated during the process of intravasation in breast cancer cells | [95] | |
| Treatment resistance | Disruption of both annexin A2 and S100A10 interactions sensitize leukemia cells to chemotherapy | [64] |
| Overexpression of S100A10 reduces the sensitivity of colorectal cancer cells to oxaliplatin | [51] | |
| Knockdown of S100A10 inhibits annexin A2 phosphorylation and increases sensitivity of acute lymphoblastic leukemia cells to prednisolone | [96] | |
| S100A10 protein expression is increased in tamoxifen-resistant MCF-7 breast cancer cells and breast cancer tissues | [97,98] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noye, T.M.; Lokman, N.A.; Oehler, M.K.; Ricciardelli, C. S100A10 and Cancer Hallmarks: Structure, Functions, and its Emerging Role in Ovarian Cancer. Int. J. Mol. Sci. 2018, 19, 4122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124122
Noye TM, Lokman NA, Oehler MK, Ricciardelli C. S100A10 and Cancer Hallmarks: Structure, Functions, and its Emerging Role in Ovarian Cancer. International Journal of Molecular Sciences. 2018; 19(12):4122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124122
Chicago/Turabian StyleNoye, Tannith M., Noor A. Lokman, Martin K. Oehler, and Carmela Ricciardelli. 2018. "S100A10 and Cancer Hallmarks: Structure, Functions, and its Emerging Role in Ovarian Cancer" International Journal of Molecular Sciences 19, no. 12: 4122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19124122