GC-MS Metabolomics to Evaluate the Composition of Plant Cuticular Waxes for Four Triticum aestivum Cultivars

 ,
,
Abstract
:
1. Introduction
2. Results
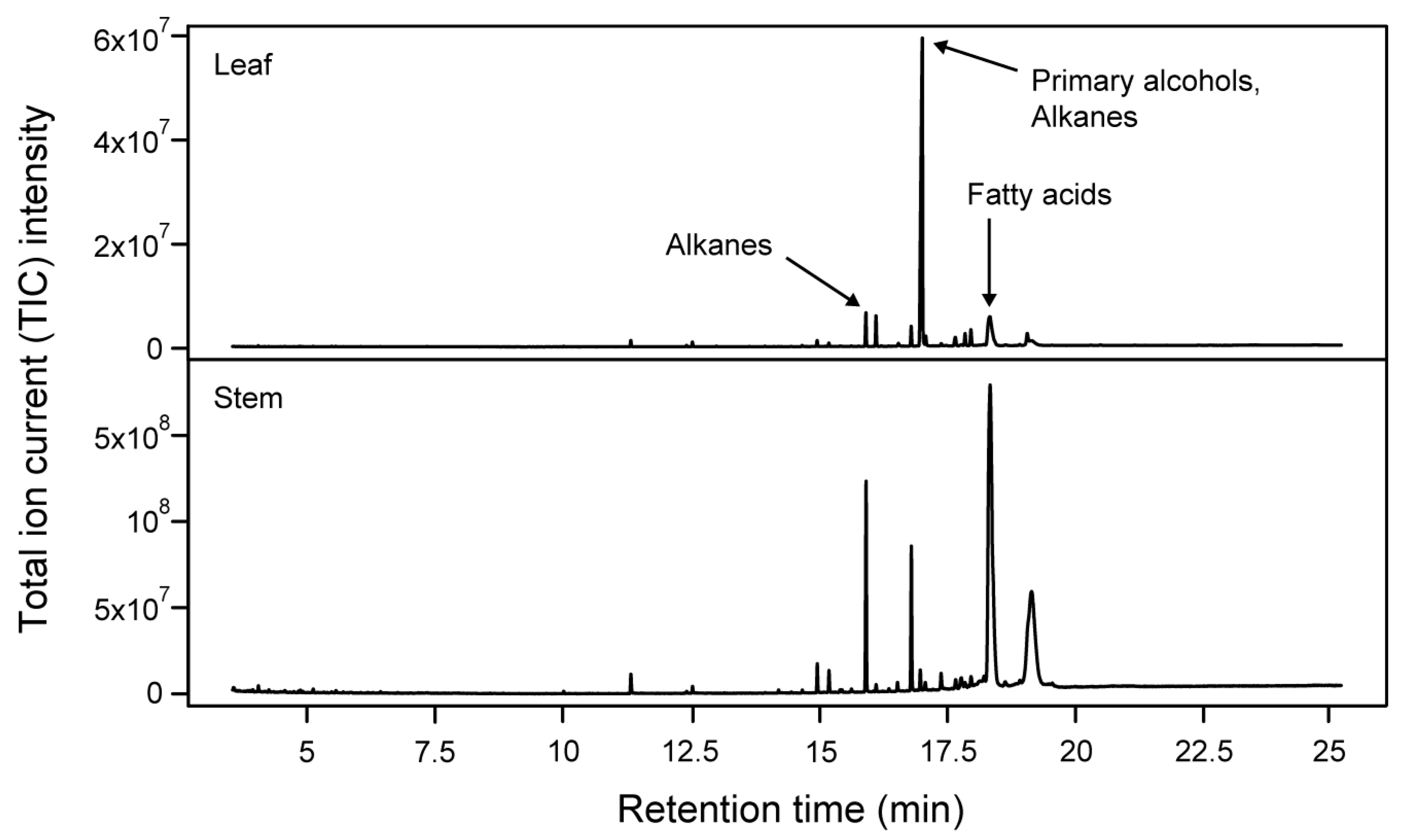
2.1. Cuticular Waxes Detected on Leaf and Stem Cuticles of Wheat
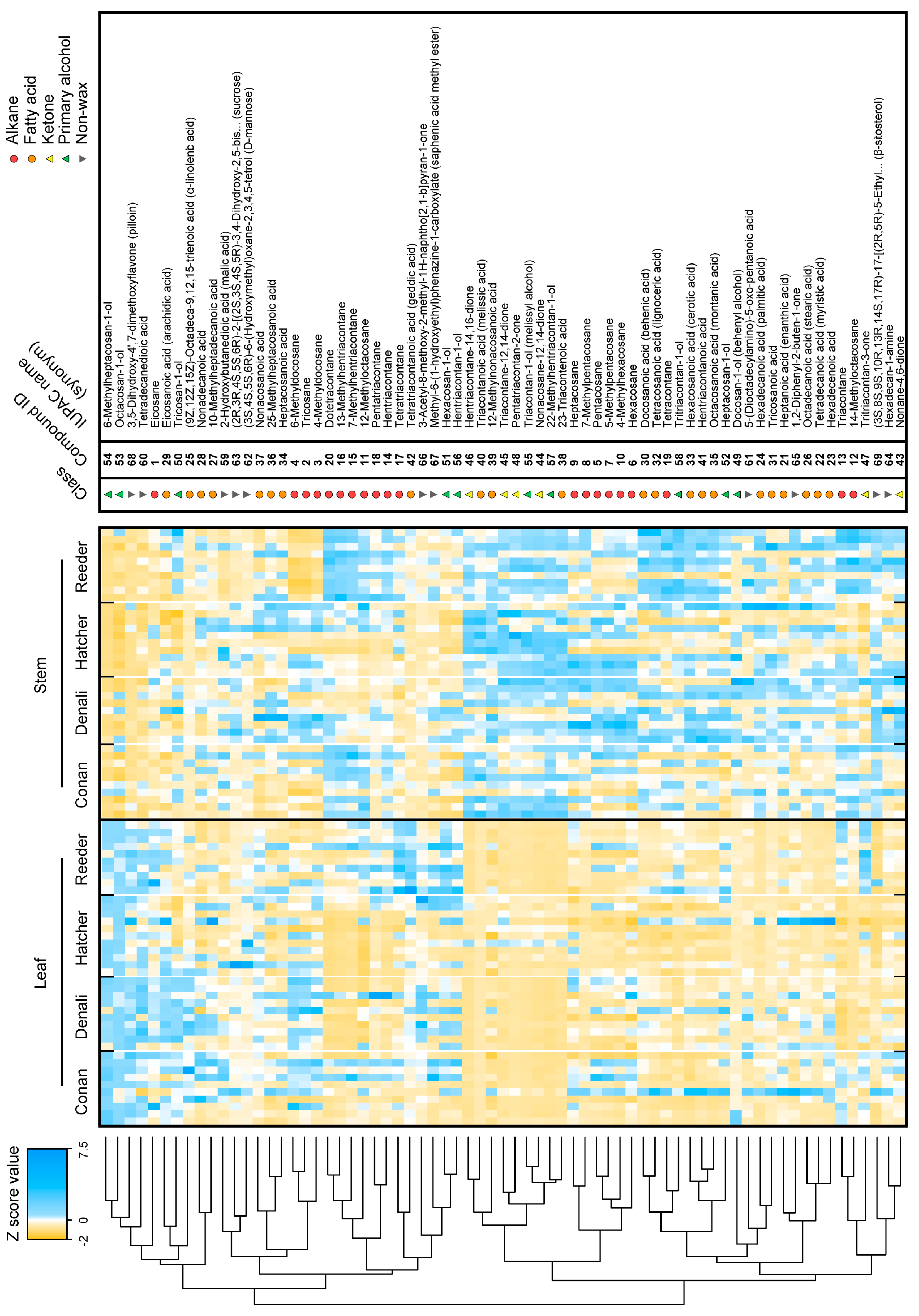
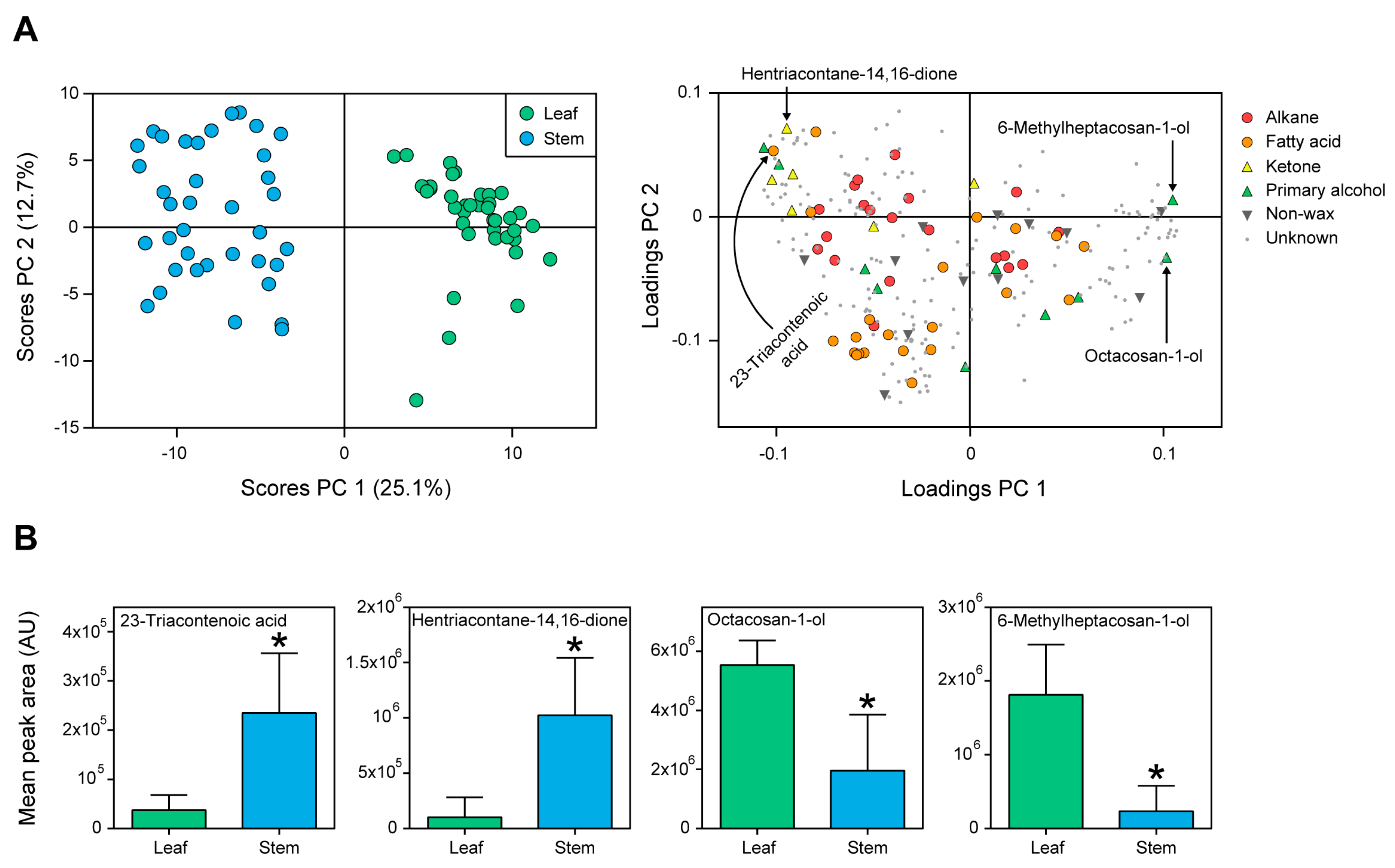
2.2. Cuticular Wax Composition in Leaves and Stems of Wheat
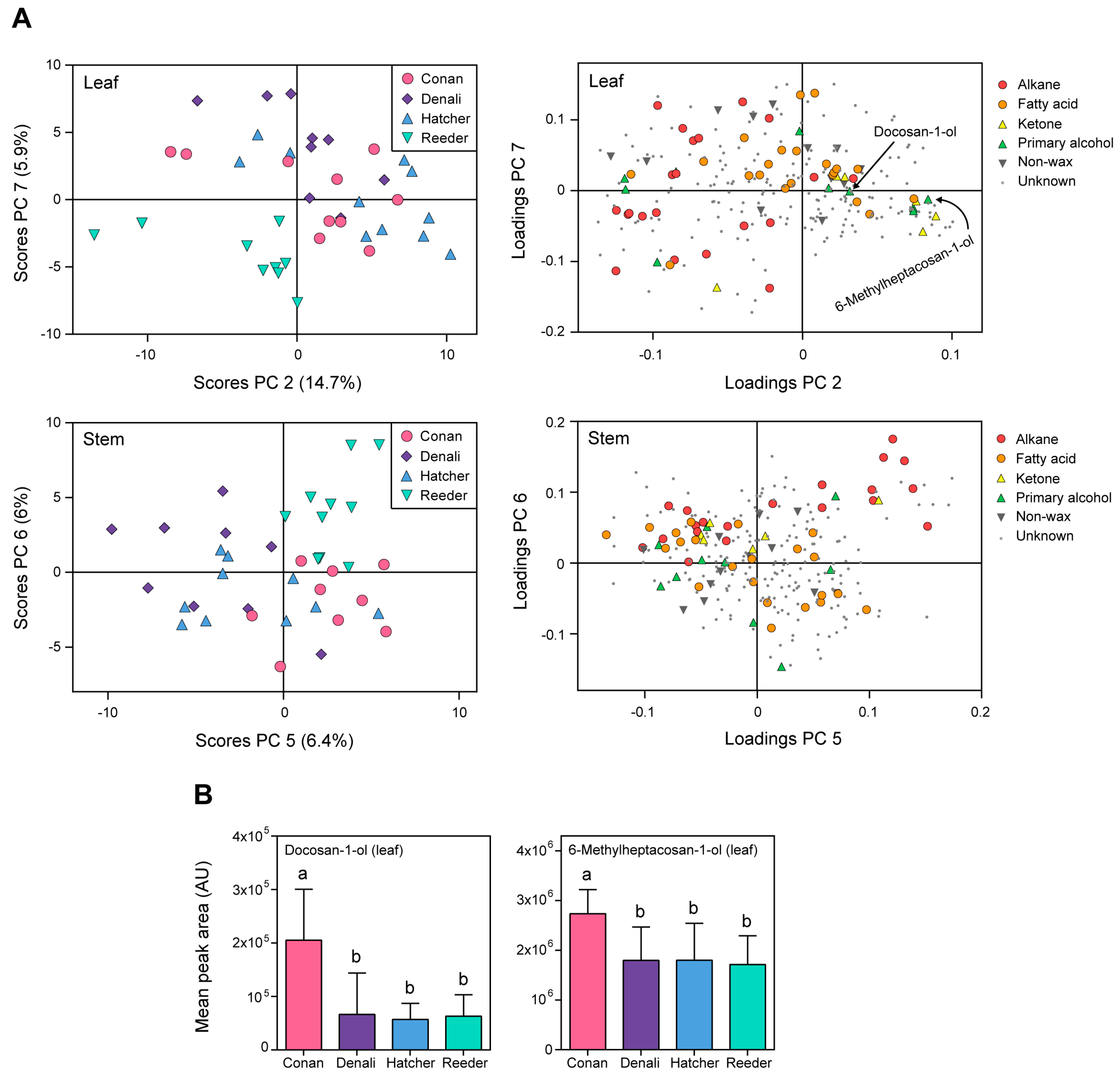
2.3. Wheat Cuticular Wax Composition among Cultivars
2.4. Association of Epicuticular Wax Content and Crystal Microstructure in Wheat
3. Discussion
3.1. Many of the Detected Cuticular Waxes Have Specialized Biological Functions
3.2. Cuticular Waxes Differed between Leaves and Stems
3.3. Cuticular Waxes Varied in Composition among the Four Cultivars
3.4. Cuticular Wax Composition Was Associated with Epicuticular Wax Crystal Microstructure
3.5. A GC-MS Metabolomics Workflow Was Applicable to Assess Epicuticular Wax Variation in Wheat
3.6. Conclusions
4. Materials and Methods
4.1. Plant Material
4.2. Leaf Photographs for Glaucousness
4.3. Metabolite Extraction, Detection by GC-MS, Data Processing and Annotation
4.4. Scanning Electron Microscopy
4.5. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of variance |
| FDR | False discovery rate |
| GC-MS | Gas chromatography-mass spectrometry |
| HSD | Honest significant difference |
| PC | Principal component |
| PCA | Principal component analysis |
| RI | Retention index |
| SEM | Scanning electron microscope |
| TIC | Total ion current |
References
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef] [PubMed]
- Riederer, M.; Schreiber, L. Protecting against water loss: Analysis of the barrier properties of plant cuticles. J. Exp. Bot. 2001, 52, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Luang, S.; Li, Y.; Bazanova, N.; Morran, S.; Song, Z.; Perera, M.A.; Hrmova, M.; Borisjuk, N.; Lopato, S. Identification and characterization of wheat drought-responsive MYB transcription factors involved in the regulation of cuticle biosynthesis. J. Exp. Bot. 2016, 67, 5363–5380. [Google Scholar] [CrossRef] [PubMed]
- Baur, P.; Buchholz, A.; Schönherr, J. Diffusion in plant cuticles as affected by temperature and size of organic solutes: Similarity and diversity among species. Plant Cell Environ. 1997, 20, 982–994. [Google Scholar] [CrossRef]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L.; Samuels, L. Plant cuticles shine: Advances in wax biosynthesis and export. Curr. Opin. Plant Biol. 2009, 12, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Metraux, J.P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef] [PubMed]
- Eigenbrode, S.D.; Espelie, K.E.; Shelton, A.M. Behavior of neonate Diamondback moth larvae [Plutella xylostella (L.)] on leaves and on extracted leaf waxes of resistant and susceptible cabbages. J. Chem. Ecol. 1991, 17, 1691–1704. [Google Scholar] [CrossRef] [PubMed]
- Krauss, P.; Markstadter, C.; Riederer, M. Attenuation of UV radiation by plant cuticles from woody species. Plant Cell Environ. 1997, 20, 1079–1085. [Google Scholar] [CrossRef]
- Cervantes, D.E.; Eigenbrode, S.D.; Ding, H.-J.; Bosque-Pérez, N.A. Oviposition responses by Hessian fly, Mayetiola destructor, to wheats varying in surfaces waxes. J. Chem. Ecol. 2002, 28, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Jenks, M.A.; Gaston, C.H.; Goodwin, M.S.; Keith, J.A.; Teusink, R.S. Seasonal variation in cuticular waxes on Hosta genotypes differing in leaf surface glaucousness. HortScience 2002, 37, 673–677. [Google Scholar]
- Von Wettstein-Knowles, P. Plant Waxes; John Wiley & Sons, Ltd.: Chichester, UK, 2012. [Google Scholar]
- Wang, Y.; Wang, J.; Chai, G.; Li, C.; Hu, Y.; Chen, X.; Wang, Z. Developmental changes in composition and morphology of cuticular waxes on leaves and spikes of glossy and glaucous wheat (Triticum aestivum L.). PLoS ONE 2015, 10, e0141239. [Google Scholar] [CrossRef] [PubMed]
- Hen-Avivi, S.; Savin, O.; Racovita, R.C.; Lee, W.S.; Adamski, N.M.; Malitsky, S.; Almekias-Siegl, E.; Levy, M.; Vautrin, S.; Berges, H.; et al. A metabolic gene cluster in the wheat W1 and the barley Cer-cqu loci determines β-diketone biosynthesis and glaucousness. Plant Cell 2016, 28, 1440–1460. [Google Scholar] [CrossRef] [PubMed]
- Barthlott, W.; Neinhuis, C.; Cutler, D.; Ditsch, F.; Meusel, I.; Theisen, I.; Wilhelmi, H. Classification and terminology of plant epicuticular waxes. Bot. J. Linn. Soc. 1998, 126, 237–260. [Google Scholar] [CrossRef]
- Chambers, T.C.; Ritchie, I.M.; Booth, M.A. Chemical models for plant wax morphogenesis. New Phytol. 1976, 77, 43–49. [Google Scholar] [CrossRef]
- Adamski, N.M.; Bush, M.S.; Simmonds, J.; Turner, A.S.; Mugford, S.G.; Jones, A.; Findlay, K.; Pedentchouk, N.; von Wettstein-Knowles, P.; Uauy, C. The Inhibitor of wax 1 (Iw1) prevents formation of β- and OH-β-diketones in wheat cuticular waxes and maps to a sub-cM interval on chromosome arm 2BS. Plant J. 2013, 74, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Blenn, B.; Bandoly, M.; Küffne, A.; Otte, T.; Geiselhardt, S.; Fatouros, N.E.; Hilker, M. Insect egg deposition induces indirect defense and epicuticular wax changes in Arabidopsis thaliana. J. Chem. Ecol. 2012, 38, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, S.Z.; Simas, N.K.; Arruda, R.C.O.; Sato, A. Composition of epicuticular wax layer of two species of Mandevilla (Apocynoideae, Apocynaceae) from Rio de Janeiro, Brazil. Biochem. Syst. Ecol. 2011, 39, 198–202. [Google Scholar] [CrossRef]
- Still, G.G.; Davis, D.G.; Zander, G.L. Plant epicuticular lipids: Alteration by herbicidal carbamates. Plant Physiol. 1970, 46, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Athukorala, Y.; Mazza, G. Supercritical carbon dioxide and hexane extraction of wax from triticale straw: Content, composition and thermal properties. Ind. Crops Prod. 2010, 31, 550–556. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Broeckling, C.D.; Blancaflor, E.B.; Sledge, M.K.; Sumner, L.W.; Wang, Z.Y. Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa). Plant J. 2005, 42, 689–707. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Broeckling, C.D.; Sumner, L.W.; Wang, Z.Y. Heterologous expression of two Medicago truncatula putative ERF transcription factor genes, WXP1 and WXP1, in Arabidopsis led to increased leaf wax accumulation and improved drought tolerance, but differential response in freezing tolerance. Plant Mol. Biol. 2007, 64, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Bush, R.T.; McInerney, F.A. Leaf wax n-alkane distributions in and across modern plants: Implications for paleoecology and chemotaxonomy. Geochim. Cosmochim. Acta 2013, 117, 161–179. [Google Scholar] [CrossRef]
- Loneman, D.M.; Peddicord, L.; Al-Rashid, A.; Nikolau, B.J.; Lauter, N.; Yandeau-Nelson, M.D. A robust and efficient method for the extraction of plant extracellular surface lipids as applied to the analysis of silks and seedling leaves of maize. PLoS ONE 2017, 12, e0180850. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, B.; Zhang, J.; Wang, C.; Liu, C.; Liu, Y.; Zhu, X.; Ren, X. Relationships between cuticular waxes and skin greasiness of apples during storage. Postharvest Biol. Technol. 2017, 131, 55–67. [Google Scholar] [CrossRef]
- Osorio, S.; Thi Do, P.; Fernie, A.R. Profiling primary metabolites of tomato fruit with gas chromatography/mass spectrometry. In Plant Metabolomics: Methods and Protocols; Hardy, N.W., Hall, R.D., Eds.; Springer/Humana Press: New York, NY, USA, 2012; pp. 101–109. ISBN 978-1-61779-594-7. [Google Scholar]
- Racovita, R.C.; Hen-Avivi, S.; Fernandez-Moreno, J.P.; Granell, A.; Aharoni, A.; Jetter, R. Composition of cuticular waxes coating flag leaf blades and peduncles of Triticum aestivum cv. Bethlehem. Phytochemistry 2016, 130, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Steinbauer, M.J.; Davies, N.W.; Gaertner, C.; Derridj, S. Epicuticular waxes and plant primary metabolites on the surfaces of juvenile Eucalyptus globulus and E. nitens (Myrtaceae) leaves. Aust. J. Bot. 2009, 57, 474–485. [Google Scholar] [CrossRef]
- Weaver, D.K.; Buteler, M.; Hofland, M.L.; Runyon, J.B.; Nansen, C.; Talbert, L.E.; Lamb, P.; Carlson, G.R. Cultivar preferences of ovipositing Wheat stem sawflies as influenced by the amount of volatile attractant. J. Econ. Entomol. 2009, 102, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Aretz, I.; Meierhofer, D. Advantages and pitfalls of mass spectrometry based metabolome profiling in systems biology. Int. J. Mol. Sci. 2016, 17, 632. [Google Scholar] [CrossRef] [PubMed]
- Heuberger, A.L.; Robison, F.M.; Lyons, S.M.A.; Broeckling, C.D.; Prenni, J.E. Evaluating plant immunity using mass spectrometry-based metabolomics workflows. Front. Plant Sci. 2014, 5, 291. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, M.P.; Heuberger, A.L.; Blanco, F.; Forrellad, M.; Taibo, C.; Klepp, L.; Sabio García, J.; Nikel, P.I.; Jackson, M.; Bigi, F. Metabolic profile of Mycobacterium smegmatis reveals Mce4 proteins are relevant for cell wall lipid homeostasis. Metabolomics 2016, 12, 97. [Google Scholar] [CrossRef]
- Abdel-Ghany, S.E.; Day, I.; Heuberger, A.L.; Broeckling, C.D.; Reddy, A.S.N. Metabolic engineering of Arabidopsis for butanetriol production using bacterial genes. Metab. Eng. 2013, 20, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Heo, H.Y.; Blake, N.; Stougaard, R.N.; Kephart, K.D.; Wichman, D.M.; Carr, P.; Briar, S.; Miller, J.; Reddy, G.V.P.; Lamb, P.; et al. Performance Evaluation and Recommendations for Spring Wheat. Unpublished work.
- Gorb, E.; Bohm, S.; Jacky, N.; Maier, L.P.; Dening, K.; Pechook, S.; Pokroy, B.; Gorb, S. Insect attachment on crystalline bioinspired wax surfaces formed by alkanes of varying chain lengths. Beilstein J. Nanotechnol. 2014, 5, 1031–1041. [Google Scholar] [CrossRef] [PubMed]
- Agoramoorthy, G.; Chandrasekaran, M.; Venkatesalu, V.; Hsu, M.J. Antibacterial and antifungal activities of fatty acid methyl esters of the blind-your-eye mangrove from India. Braz. J. Microbiol. 2007, 38, 739–742. [Google Scholar] [CrossRef]
- Vanhercke, T.; Wood, C.C.; Stymne, S.; Singh, S.P.; Green, A.G. Metabolic engineering of plant oils and waxes for use as industrial feedstocks. Plant Biotechnol. J. 2013, 11, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Leszczynski, B.; Goławska, S.; Matok, H. Allelopathic action of Triticale allochemicals towards grain aphid. In Allelopathy: New Concepts and Methodology; Fujii, Y., Hiradate, S., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 353–364. ISBN 9781578084463. [Google Scholar]
- Karmakar, A.; Barik, A. Solena aplexicaulis (Cucurbitaceae) flower surface wax influencing attraction of a generalist insect herbivore, Aulacophora foveicollis (Coleoptera: Chrysomelidae). Int. J. Trop. Insect Sci. 2016, 36, 70–81. [Google Scholar] [CrossRef]
- Dyer, J.M.; Stymne, S.; Green, A.G.; Carlsson, A.S. High-value oils from plants. Plant J. 2008, 54, 640–655. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jung, J.H.; Lee, S.B.; Go, Y.S.; Kim, H.J.; Cahoon, R.; Markham, J.E.; Cahoon, E.B.; Suh, M.C. Arabidopsis 3-ketoacyl-coenzyme A synthase9 is involved in the synthesis of tetracosanoic acids as precursors of cuticular waxes, suberins, sphingolipids, and phospholipids. Plant Physiol. 2013, 162, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Broz, A.K.; Broeckling, C.D.; De-la-Peña, C.; Lewis, M.R.; Greene, E.; Callaway, R.M.; Sumner, L.W.; Vivanco, J.M. Plant neighbor identity influences plant biochemistry and physiology related to defense. BMC Plant Biol. 2010, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.; Knights, B.A.; Math, V.B.; Ritchie, A.L. β-diketones in Rhododendron waxes. Phytochemistry 1975, 14, 2447–2451. [Google Scholar] [CrossRef]
- Zhang, Z.; Wei, W.; Zhu, H.; Challa, G.S.; Bi, C.; Trick, H.N.; Li, W. W3 is a new wax locus that is essential for biosynthesis of β-diketone, development of glaucousness, and reduction of cuticle permeability in common wheat. PLoS ONE 2015, 10, e0140524. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.A.; Richards, R.A.; Turner, N.C. Yield, water relations, gas exchange, and surface reflectances of near-isogenic wheat lines differing in glaucousness. Crop Sci. 1983, 23, 318–325. [Google Scholar] [CrossRef]
- Wu, H.; Qin, J.; Han, J.; Zhao, X.; Ouyang, S.; Liang, Y.; Zhang, D.; Wang, Z.; Wu, Q.; Xie, J.; et al. Comparative high-resolution mapping of the wax inhibitors Iw1 and Iw2 in hexaploid wheat. PLoS ONE 2013, 8, e84691. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, Y.; Wu, H.; Xu, J.; Li, T.; Hegebarth, D.; Jetter, R.; Chen, L.; Wang, Z. Three TaFAR genes function in the biosynthesis of primary alcohols and the response to abiotic stresses in Triticum aestivum. Sci. Rep. 2016, 6, 25008. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Luang, S.; Li, Y.; Bazanova, N.; Borisjuk, N.; Hrmova, M.; Lopato, S. Wheat drought-responsive WXPL transcription factors regulate cuticle biosynthesis genes. Plant Mol. Biol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Tulloch, A.P.; Hoffman, L.L. Epicuticular waxes of Secale cereale and Triticale hexaploide leaves. Phytochemistry 1974, 13, 2535–2540. [Google Scholar] [CrossRef]
- Richards, R.A.; Rawson, H.M.; Johnson, D.A. Glaucousness in wheat: Its development and effect on water-use efficiency, gas exchange and photosynthetic tissue temperatures. Aust. J. Plant Physiol. 1986, 13, 465–473. [Google Scholar]
- Barber, H.N.; Jackson, W.D. Natural selection in action in Eucalyptus. Nature 1957, 179, 1267–1269. [Google Scholar] [CrossRef]
- Gorb, E.; Haas, K.; Henrich, A.; Enders, S.; Barbakadze, N.; Gorb, S. Composite structure of the crystalline epicuticular wax layer of the slippery zone in the pitchers of the carnivorous plant Nepenthes alata and its effect on insect attachment. J. Exp. Biol. 2005, 208, 4651–4662. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Guo, Y.J.; Guo, Y.J.; Han, L.; Tang, H.; Conyers, M. Leaf cuticular waxes and physiological parameters in alfalfa leaves as influenced by drought. Photosynthetica 2012, 50, 458–466. [Google Scholar] [CrossRef]
- Damon, S.J.; Groves, R.L.; Havey, M.J. Variation for epicuticular waxes on onion foliage and impacts on numbers of onion thrips. J. Am. Soc. Hortic. Sci. 2014, 139, 495–501. [Google Scholar]
- Cowlishaw, M.G.; Bickerstaffe, R.; Connor, H.E. Intraspecific variation in the epicuticular wax composition of four species of Chionochloa. Biochem. Syst. Ecol. 1983, 11, 247–259. [Google Scholar] [CrossRef]
- Eigenbrode, S.D.; Jetter, R. Attachment to plant surface waxes by an insect predator. Integr. Comp. Biol. 2002, 42, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Holloway, P.J.; Jeffree, C.E. Epicuticular waxes. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Thomas, B., Murphy, D.J., Murray, B.G., Eds.; Academic Press: Cambridge, MA, USA, 2017; Volume 2, pp. 374–386. ISBN 9780123948083. [Google Scholar]
- Broeckling, C.D.; Afsar, F.A.; Neumann, S.; Ben-Hur, A.; Prenni, J.E. RAMClust: A novel feature clustering method enables spectral-matching-based annotation for metabolomics data. Anal. Chem. 2014, 86, 6812–6817. [Google Scholar] [CrossRef] [PubMed]
- Mahieu, N.G.; Genenbacher, J.L.; Patti, G.J. A roadmap for the XCMS family of software solutions in metabolomics. Curr. Opin. Chem. Biol. 2016, 30, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Kosma, D.K.; Nemacheck, J.A.; Jenks, M.A.; Williams, C.E. Changes in properties of wheat leaf cuticle during interactions with Hessian fly. Plant J. 2010, 63, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Barthlott, W.; Koch, S.; Hommes, A.; Wandelt, K.; Mamdouh, W.; De-Feyter, S.; Broekmann, P. Structural analysis of wheat wax (Triticum aestivum, c.v. ‘Naturastar’ L.): From the molecular level to three dimensional crystals. Planta 2006, 223, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Haley, S.D.; Quick, J.S.; Johnson, J.J.; Peairs, F.B.; Stromberger, J.A.; Clayshulte, S.R.; Clifford, B.L.; Rudolf, J.B.; Seabourn, B.W.; Chung, O.K.; et al. Registration of ‘Hatcher’ wheat. Crop Sci. 2005, 45, 2654–2656. [Google Scholar] [CrossRef]
- Haley, S.D.; Johnson, J.J.; Peairs, F.B.; Stromberger, J.A.; Hudson, E.E.; Seifert, S.A.; Kottke, R.A.; Valdez, V.A.; Rudolph, J.B.; Martin, T.J.; et al. Registration of ‘Denali’ wheat. J. Plant Regist. 2012, 6, 311–314. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Kopka, J.; Schauer, N.; Krueger, S.; Birkemeyer, C.; Usadel, B.; Bergmüller, E.; Dörmann, P.; Weckwerth, W.; Gibon, Y.; Stitt, M.; et al. [email protected]: the Golm metabolome database. Bioinformatics 2005, 21, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Golm Metabolome Database. Available online: http://gmd.mpimp-golm.mpg.de/ (accessed on 15 September 2015).
- National Institute of Standards and Technology. Available online: http://www.nist.gov/ (accessed on 15 September 2015).
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | IUPAC Name (Synonym) | Retention Index | ANOVA p-Value † | Association | Reference | |
|---|---|---|---|---|---|---|
| Tissue | Cultivar | |||||
| Alkanes | ||||||
| 1 | Eicosane | 2000 | ‡ | 0.6 | Plant–insect int. | [10] |
| 2 | Tricosane | 2300 | 0.13 | ‡ | Chemotaxonomy | [24] |
| 3 | 4-Methyldocosane | 2334 | 0.33 | ‡ | ||
| 4 | 6-Methyldocosane | 2336 | ‡ | ‡ | ||
| 5 | Pentacosane | 2500 | ‡ | 0.77 | Chemotaxonomy | [24] |
| 6 | Hexacosane | 2600 | ‡ | 0.17 | Chemotaxonomy | [24] |
| 7 | 5-Methylpentacosane | 2641 | ‡ | 0.63 | ||
| 8 | 7-Methylpentacosane | 2645 | ‡ | 0.63 | ||
| 9 | Heptacosane | 2700 | ‡ | 0.56 | Chemotaxonomy | [24] |
| 10 | 4-Methylhexacosane | 2727 | ‡ | 0.4 | ||
| 11 | 12-Methyloctacosane | 2912 | ‡ | ‡ | ||
| 12 | 14-Methyloctacosane | 2915 | ‡ | ‡ | ||
| 13 | Triacontane | 3000 | ‡ | ‡ | Chemotaxonomy | [24] |
| 14 | Hentriacontane | 3100 | 0.12 | 0.05 | Plant–insect int. | [10] |
| 15 | 7-Methylhentriacontane | 3111 | ‡ | ‡ | ||
| 16 | 13-Methylhentriacontane | 3124 | ‡ | ‡ | ||
| 17 | Tetratriacontane | 3400 | 0.27 | ‡ | Chemotaxonomy | [24] |
| 18 | Pentatriacontane | 3500 | 0.61 | 0.16 | Chemotaxonomy | [24] |
| 19 | Tetracontane | 4000 | ‡ | ‡ | Plant–insect int. | [36] |
| 20 | Dotetracontane | 4200 | ‡ | ‡ | ||
| Fatty acids | ||||||
| 21 | Heptanoic acid (enanthic acid) | 1044 | 0.25 | 0.3 | ||
| 22 | Tetradecanoic acid (myristic acid) | 1499 | ‡ | 0.68 | Antibiotic | [37] |
| 23 | Hexadecenoic acid | 1602 | 0.51 | 0.57 | ||
| 24 | Hexadecanoic acid (palmitic acid) | 1623 | ‡ | 0.9 | Plant–insect int. | [10] |
| 25 | (9Z,12Z,15Z)-Octadeca-9,12,15-trienoic acid (α-linolenic acid) | 1755 | ‡ | 0.13 | Oil production | [38] |
| 26 | Octadecanoic acid (stearic acid) | 1765 | 0.25 | 0.77 | Plant–insect int. | [39] |
| 27 | 10-Methylheptadecanoic acid | 1768 | 0.12 | 0.61 | ||
| 28 | Nonadecanoic acid | 1825 | 0.67 | 0.3 | Plant–insect int. | [40] |
| 29 | Eicosanoic acid (arachidic acid) | 1911 | ‡ | 0.52 | ||
| 30 | Docosanoic acid (behenic acid) | 2103 | ‡ | 0.05 | Oil production | [41] |
| 31 | Tricosanoic acid | 2166 | ‡ | 0.77 | ||
| 32 | Tetracosanoic acid (lignoceric acid) | 2246 | ‡ | 0.09 | Major wax precursor | [42] |
| 33 | Hexacosanoic acid (cerotic acid) | 2404 | ‡ | 0.06 | Allelopathy | [43] |
| 34 | Heptacosanoic acid | 2497 | 0.07 | 0.17 | ||
| 35 | Octacosanoic acid (montanic acid) | 2563 | ‡ | 0.46 | ||
| 36 | 25-Methylheptacosanoic acid | 2569 | 0.07 | 0.17 | ||
| 37 | Nonacosanoic acid | 2647 | 0.61 | 0.25 | ||
| 38 | 23-Triacontenoic acid | 2728 | ‡ | 0.54 | ||
| 39 | 12-Methylnonacosanoic acid | 2731 | ‡ | 0.77 | ||
| 40 | Triacontanoic acid (melissic acid) | 2741 | ‡ | 0.32 | ||
| 41 | Hentriacontanoic acid | 2796 | ‡ | 0.32 | ||
| 42 | Tetratriacontanoic acid (geddic acid) | 3037 | ‡ | ‡ | Plant–insect int. | [18] |
| Ketones | ||||||
| 43 | Nonane-4,6-dione | 1124 | ‡ | 0.33 | ||
| 44 | Nonacosane-12,14-dione | 2689 | ‡ | 0.41 | Major leaf wax | [44] |
| 45 | Triacontane-12,14-dione | 2698 | ‡ | 0.21 | ||
| 46 | Hentriacontane-14,16-dione | 2735 | ‡ | 0.39 | Glaucousness | [45] |
| 47 | Tritriacontan-3-one | 2839 | 0.93 | ‡ | ||
| 48 | Pentatriacontan-2-one | 2881 | ‡ | 0.54 | ||
| Primary alcohols | ||||||
| 49 | Docosan-1-ol (behenyl alcohol) | 2044 | ‡ | ‡ | ||
| 50 | Tricosan-1-ol | 2112 | 0.79 | ‡ | ||
| 51 | Hexacosan-1-ol | 2325 | ‡ | ‡ | Plant–insect int. | [10] |
| 52 | Heptacosan-1-ol | 2404 | 0.9 | ‡ | ||
| 53 | Octacosan-1-ol | 2467 | ‡ | 0.77 | Plant–insect int. | [10] |
| 54 | 6-Methylheptacosan-1-ol | 2498 | ‡ | ‡ | ||
| 55 | Triacontan-1-ol (melissyl alcohol) | 2668 | ‡ | 0.63 | ||
| 56 | Hentriacontan-1-ol | 2703 | ‡ | 0.25 | ||
| 57 | 22-Methylhentriacontan-1-ol | 2780 | ‡ | 0.41 | ||
| 58 | Tritriacontan-1-ol | 2855 | ‡ | ‡ | ||
| # | Class | IUPAC Name (Synonym) | RI | ANOVA p-Value † | |
|---|---|---|---|---|---|
| Tissue | Cultivar | ||||
| 59 | Carboxylic acid | 2-Hydroxybutanedioic acid (malic acid) | 1192 | 0.19 | ‡ |
| 60 | Tetradecanedioic acid | 1471 | 0.14 | 0.34 | |
| 61 | 5-(Dioctadecylamino)-5-oxo-pentanoic acid | 2589 | ‡ | 0.3 | |
| 62 | Carbohydrate | (3 S,4S,5S,6R)-6-(Hydroxymethyl)oxane-2,3,4,5-tetrol (d-mannose) | 1602 | 0.23 | ‡ |
| 63 | (2 R,3R,4S,5S,6R)-2-[(2S,3S,4S,5R)-3,4-Dihydroxy-2,5-bis(hydroxymethyl)Oxolan-2-yl]oxy-6-(hydroxymethyl)oxane-3,4,5-triol (sucrose) | 2140 | 0.89 | 0.3 | |
| 64 | Primary amine | Hexadecan-1-amine | 854 | ‡ | 0.61 |
| 65 | Polycyclic ketone | 1,2-Diphenyl-2-buten-1-one | 2184 | 0.15 | 0.57 |
| 66 | 3-Acetyl-8-methoxy-2-methyl-1H-naphtho[2,1-b]pyran-1-one | 2389 | ‡ | 0.14 | |
| 67 | FAME | Methyl-6-(1-hydroxyethyl)phenazine-1-carboxylate(saphenic acid methyl ester) | 2885 | ‡ | 0.24 |
| 68 | Flavonoid | 3,5-Dihydroxy-4′′,7-dimethoxyflavone (pilloin) | 2287 | ‡ | 0.92 |
| 69 | Sterol | (3 S,8S,9S,10R,13R,14S,17R)-17-[(2R,5R)-5-Ethyl-6-methylheptan-2-yl]-10,13-dimethyl-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-ol (β-sitosterol) | 2801 | ‡ | 0.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lavergne, F.D.; Broeckling, C.D.; Cockrell, D.M.; Haley, S.D.; Peairs, F.B.; Jahn, C.E.; Heuberger, A.L. GC-MS Metabolomics to Evaluate the Composition of Plant Cuticular Waxes for Four Triticum aestivum Cultivars. Int. J. Mol. Sci. 2018, 19, 249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020249
Lavergne FD, Broeckling CD, Cockrell DM, Haley SD, Peairs FB, Jahn CE, Heuberger AL. GC-MS Metabolomics to Evaluate the Composition of Plant Cuticular Waxes for Four Triticum aestivum Cultivars. International Journal of Molecular Sciences. 2018; 19(2):249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020249
Chicago/Turabian StyleLavergne, Florent D., Corey D. Broeckling, Darren M. Cockrell, Scott D. Haley, Frank B. Peairs, Courtney E. Jahn, and Adam L. Heuberger. 2018. "GC-MS Metabolomics to Evaluate the Composition of Plant Cuticular Waxes for Four Triticum aestivum Cultivars" International Journal of Molecular Sciences 19, no. 2: 249. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020249