Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice


Abstract
:1. Introduction
2. Results
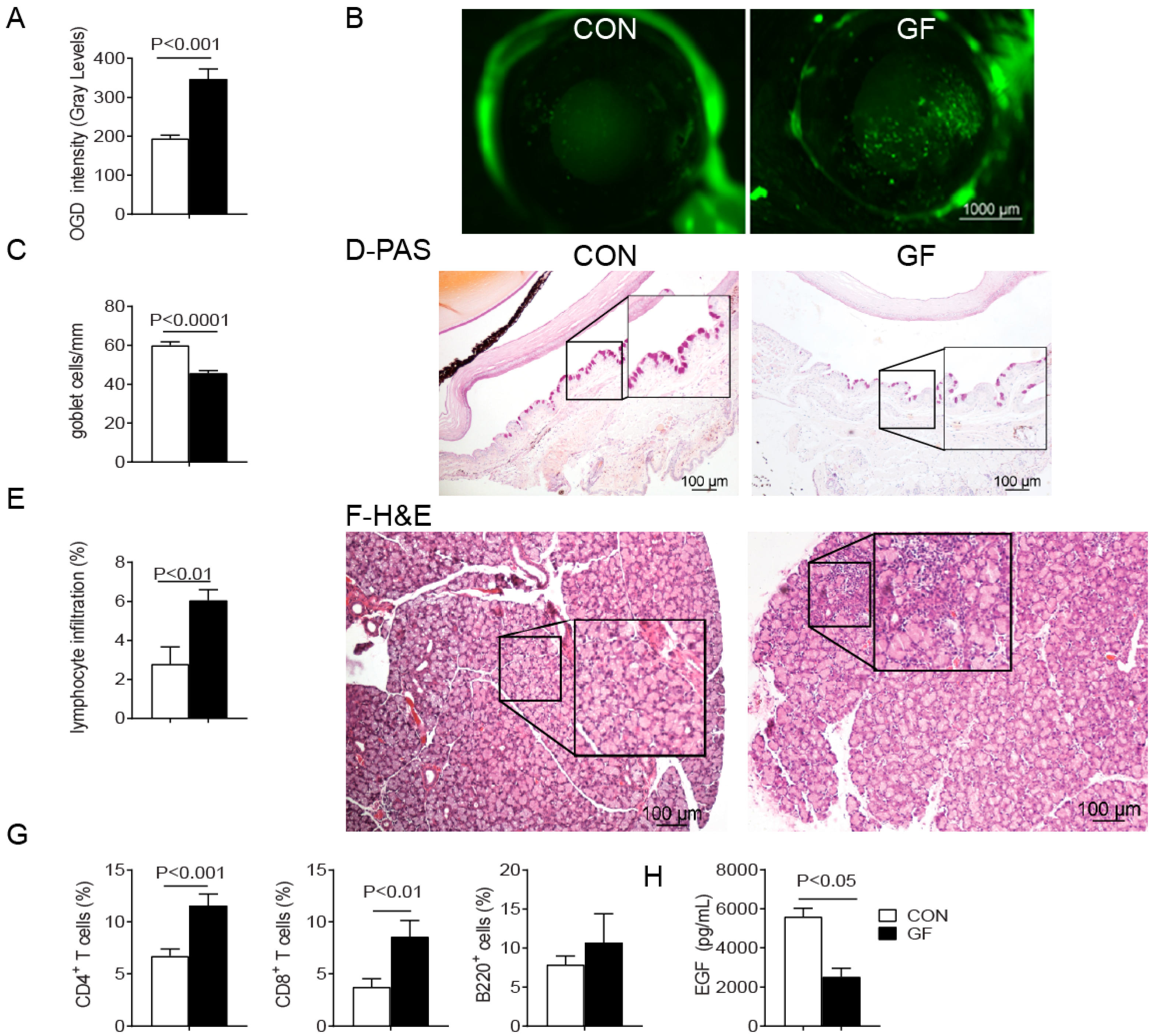
2.1. Spontaneous Sjögren-Like Lacrimal Keratoconjunctivitis
2.2. Sex Differences in Ocular Surface Manifestations
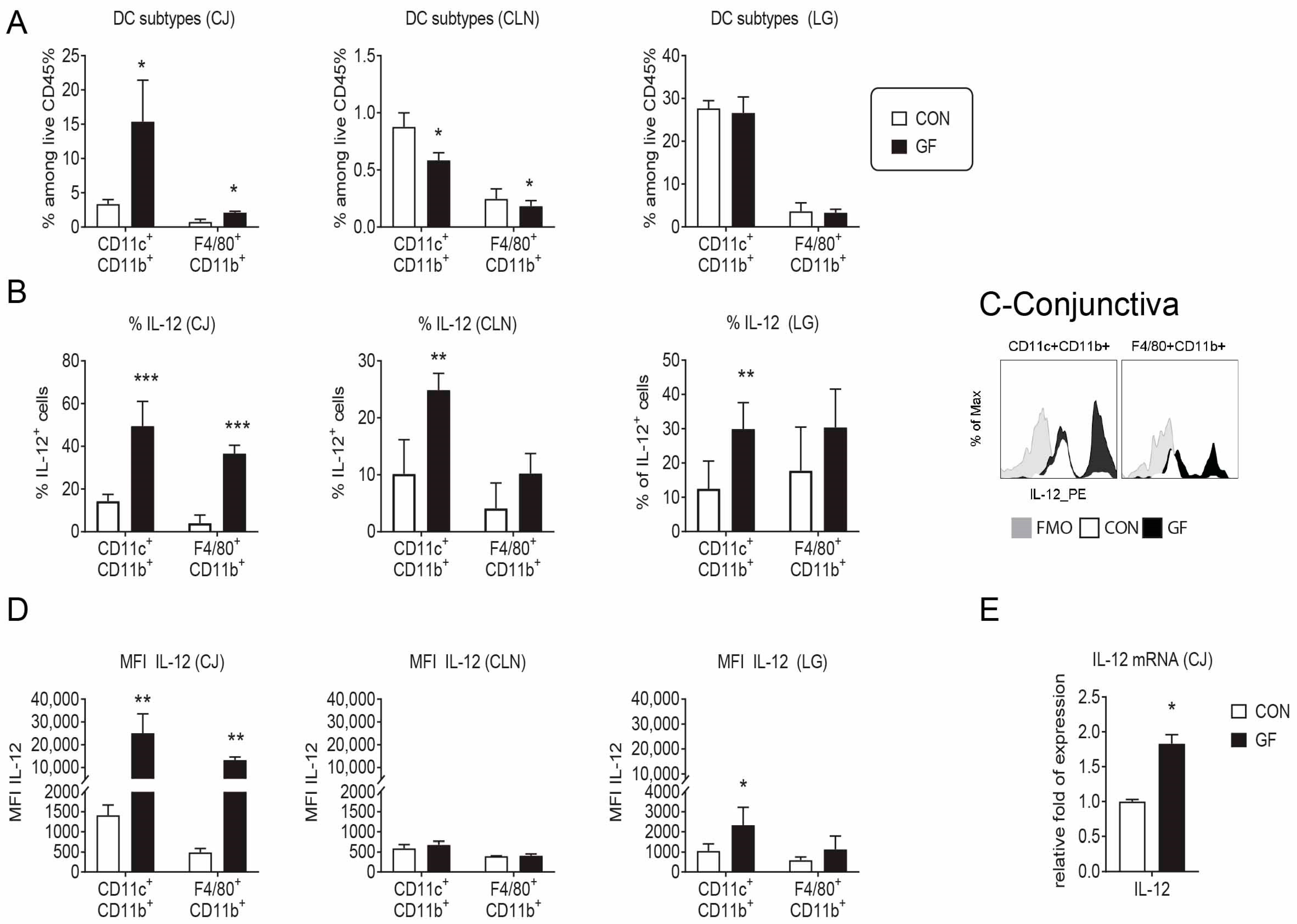
2.3. Changes in the Ocular Environment in Germ-Free Mice Predispose to Th-1 Cells
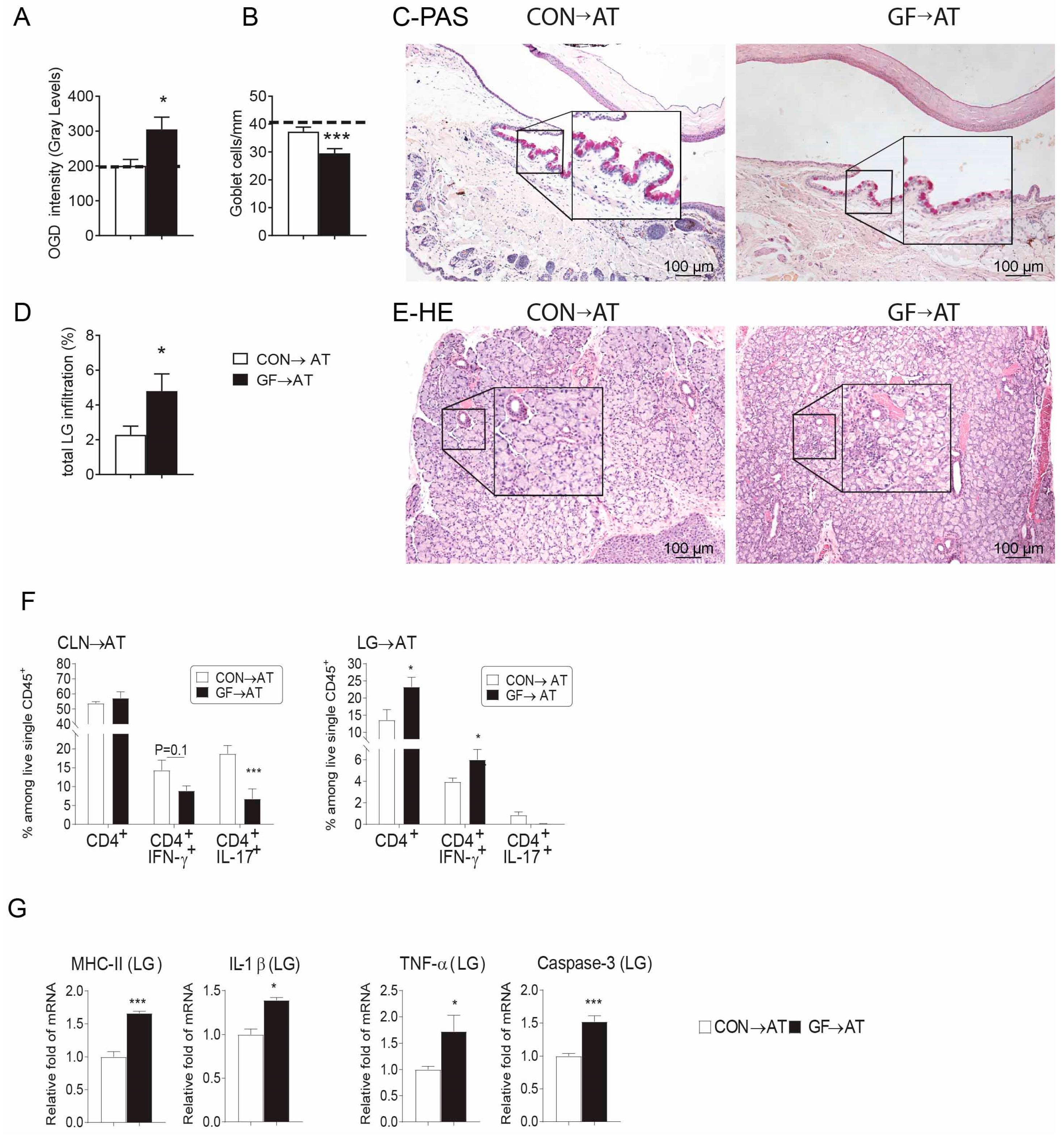
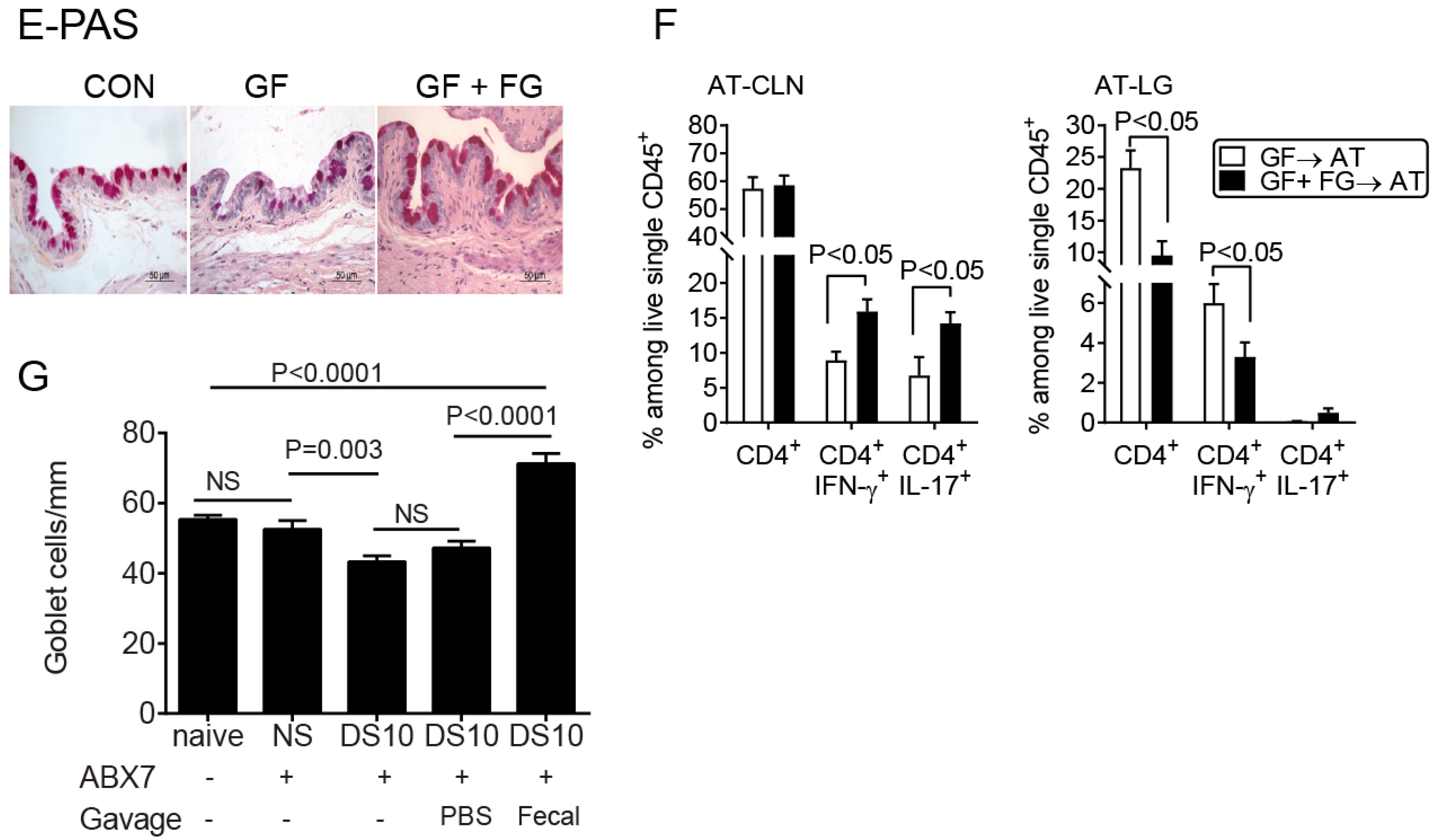
2.4. CD4+ T Cells from Germ-Free Mice Induce Sjögren-Like Lacrimal Keratoconjunctivitis
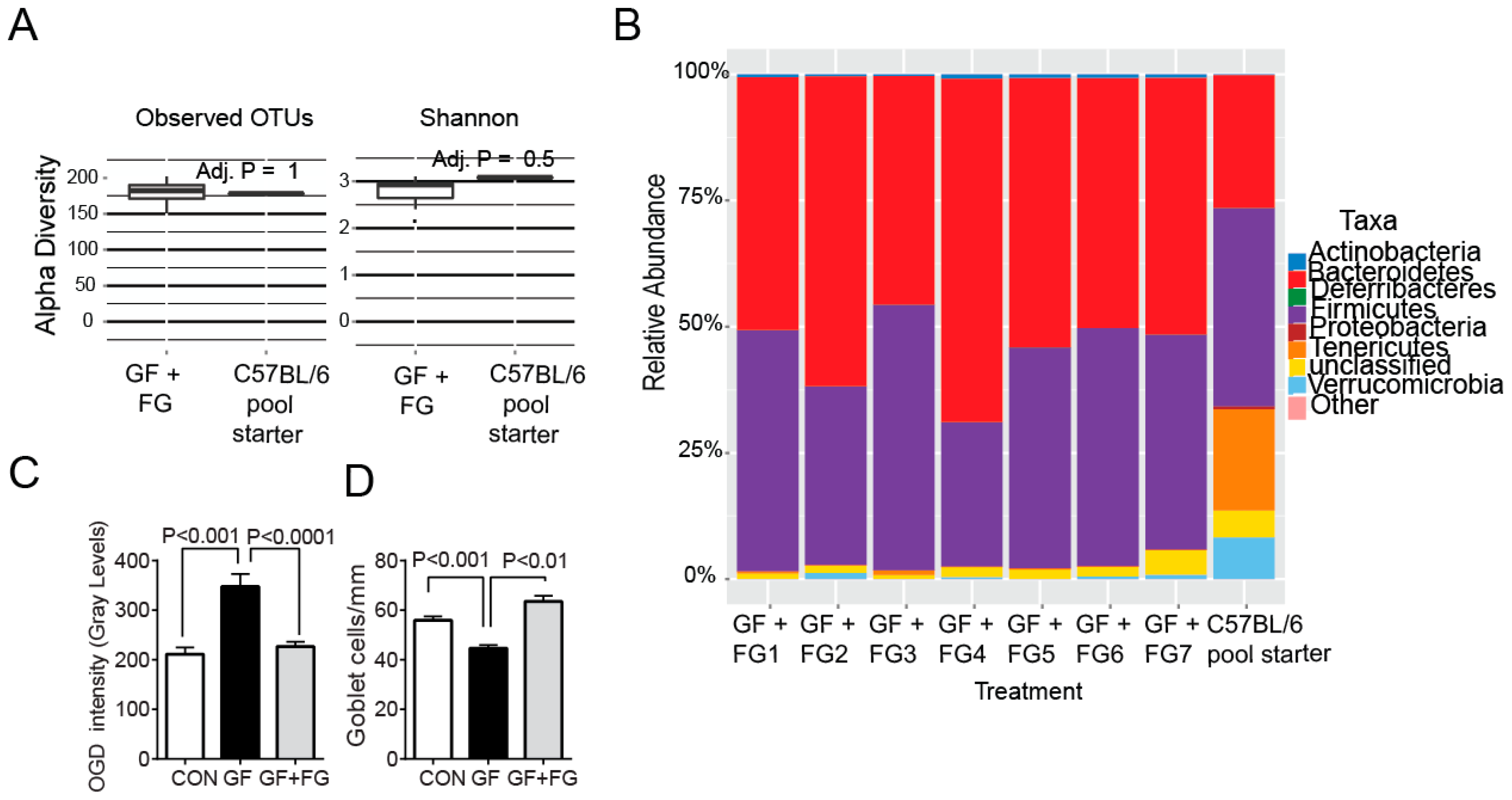
2.5. Fecal Microbiota Transplant Reversed Lacrimal Keratoconjunctivitis in Germ-Free Mice
3. Discussion
4. Methods
4.1. Animals
4.2. Corneal Permeability
4.3. Histology and PAS Staining
4.4. Measurement of Lacrimal Gland Infiltration
4.5. Flow Cytometry Analysis of Infiltrating Cells
4.6. Tear Washings and EGF Enzyme-Linked Immunosorbent Assay
4.7. Caspase-3 Activation Fluorometric Assays
4.8. RNA Isolation and Real-Time PCR
4.9. Isolation of Murine CD4+ T Cells and Adoptive Transfer
4.10. Standard Desiccating Stress Model
4.11. Antibiotic Treatment and Desiccating Stress
4.12. Fecal Microbiota Transplant
4.13. DNA Extraction from Mouse Fecal Samples, 16S rRNA Gene Amplication, and Sequencing
4.14. Microbial Community Analysis
4.15. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nolan, J. Evaluation of conjunctival and nasal bacterial cultures before intra-ocular operations. Br. J. Ophthalmol. 1967, 51, 483–485. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, J.A.; Moller, S.; Bentzon, M.W. Bacterial flora in relation to cataract extraction. II. Peroperative flora. Acta Ophthalmol. 1975, 53, 476–494. [Google Scholar] [CrossRef]
- Schabereiter-Gurtner, C.; Maca, S.; Rolleke, S.; Nigl, K.; Lukas, J.; Hirschl, A.; Lubitz, W.; Barisani-Asenbauer, T. 16S rDNA-based identification of bacteria from conjunctival swabs by PCR and DGGE fingerprinting. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1164–1171. [Google Scholar]
- Narayanan, S.; Miller, W.L.; McDermott, A.M. Expression of human β-defensins in conjunctival epithelium: Relevance to dry eye disease. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3795–3801. [Google Scholar] [CrossRef]
- Zhou, L.; Huang, L.Q.; Beuerman, R.W.; Grigg, M.E.; Li, S.F.; Chew, F.T.; Ang, L.; Stern, M.E.; Tan, D. Proteomic analysis of human tears: Defensin expression after ocular surface surgery. J. Proteome Res. 2004, 3, 410–416. [Google Scholar] [CrossRef] [PubMed]
- McDermott, A.M. Antimicrobial compounds in tears. Exp. Eye Res. 2013, 117, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Jensen, O.L.; Gluud, B.S.; Birgens, H.S. The concentration of lactoferrin in tears of normals and of diabetics. Acta Ophthalmol. 1986, 64, 83–87. [Google Scholar] [CrossRef]
- Vinding, T.; Eriksen, J.S.; Nielsen, N.V. The concentration of lysozyme and secretory IgA in tears from healthy persons with and without contact lens use. Acta Ophthalmol. 1987, 65, 23–26. [Google Scholar] [CrossRef]
- Haynes, R.J.; Tighe, P.J.; Dua, H.S. Antimicrobial defensin peptides of the human ocular surface. Br. J. Ophthalmol. 1999, 83, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Willcox, M.D. Characterization of the normal microbiota of the ocular surface. Exp. Eye Res. 2013, 117, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Zegans, M.E.; van Gelder, R.N. Considerations in understanding the ocular surface microbiome. Am. J. Ophthalmol. 2014, 158, 420–422. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, C.S.; Jones, D.B.; Stern, M.E.; Bian, F.; Moore, Q.L.; Corbiere, S.; Streckfus, C.F.; Hutchinson, D.S.; Ajami, N.J.; Petrosino, J.F.; et al. Altered Mucosal Microbiome Diversity and Disease Severity in Sjogren Syndrome. Sci. Rep. 2016, 6, 23561–23571. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, B.; Li, W. Defining the normal “core microbiome” of conjunctival microbial communities. Clin. Microbiol. Infect. 2016, 22, 643. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Moore, J.E.; Jiru, X.; Moore, J.E.; Goodall, E.A.; Dooley, J.S.; Hayes, V.E.; Dartt, D.A.; Downes, C.S.; Moore, T.C. Ocular pathogen or commensal: A PCR-based study of surface bacterial flora in normal and dry eyes. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5616–5623. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, J.; Nielsen, S.; Diez-Vives, C.; Coroneo, M.; Thomas, T.; Willcox, M. Temporal Stability and Composition of the Ocular Surface Microbiome. Sci. Rep. 2017, 7, 9880. [Google Scholar] [CrossRef] [PubMed]
- St Leger, A.J.; Desai, J.V.; Drummond, R.A.; Kugadas, A.; Almaghrabi, F.; Silver, P.; Raychaudhuri, K.; Gadjeva, M.; Iwakura, Y.; Lionakis, M.S.; et al. An Ocular Commensal Protects against Corneal Infection by Driving an Interleukin-17 Response from Mucosal γδ T Cells. Immunity 2017, 47, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Kugadas, A.; Christiansen, S.H.; Sankaranarayanan, S.; Surana, N.K.; Gauguet, S.; Kunz, R.; Fichorova, R.; Vorup-Jensen, T.; Gadjeva, M. Impact of Microbiota on Resistance to Ocular Pseudomonas aeruginosa-Induced Keratitis. PLoS Pathog. 2016, 12, e1005855. [Google Scholar] [CrossRef] [PubMed]
- Kugadas, A.; Gadjeva, M. Impact of Microbiome on Ocular Health. Ocul. Surf. 2016, 14, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Kugadas, A.; Wright, Q.; Geddes-McAlister, J.; Gadjeva, M. Role of Microbiota in Strengthening Ocular Mucosal Barrier Function through Secretory IgA. Investig. Ophthalmol. Vis. Sci. 2017, 58, 4593–4600. [Google Scholar] [CrossRef] [PubMed]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed]
- Bourassa, M.W.; Alim, I.; Bultman, S.J.; Ratan, R.R. Butyrate, neuroepigenetics and the gut microbiome: Can a high fiber diet improve brain health? Neurosci. Lett. 2016, 625, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome-gut-brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol. Ther. 2015, 158, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; Hall, J.C.; Wang, L.; Mo, X.; Yu, Z.; Popovich, P.G. Gut dysbiosis impairs recovery after spinal cord injury. J. Exp. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Luczynski, P.; McVey Neufeld, K.A.; Oriach, C.S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Growing up in a Bubble: Using Germ-Free Animals to Assess the Influence of the Gut Microbiota on Brain and Behavior. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef] [PubMed]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Dowds, C.M.; Blumberg, R.S.; Zeissig, S. Control of intestinal homeostasis through crosstalk between natural killer T cells and the intestinal microbiota. Clin. Immunol. 2015, 159, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, R.; Powrie, F. Microbiota, disease, and back to health: A metastable journey. Sci. Transl. Med. 2012, 4, 137rv7. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.E.; Priori, R.; Valesini, G.; Fairweather, D. Sex differences in Sjogren’s syndrome: A comprehensive review of immune mechanisms. Biol. Sex Differ. 2015, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Schaumberg, D.A.; Sullivan, D.A.; Buring, J.E.; Dana, M.R. Prevalence of dry eye syndrome among US women. Am. J. Ophthalmol. 2003, 136, 318–326. [Google Scholar] [CrossRef]
- McClellan, A.J.; Volpe, E.A.; Zhang, X.; Darlington, G.J.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Ocular Surface Disease and Dacryoadenitis in Aging C57BL/6 Mice. Am. J. Pathol. 2014, 184, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Bian, F.; Barbosa, F.L.; Corrales, R.M.; Pelegrino, F.S.; Volpe, E.A.; Pflugfelder, S.C.; de Paiva, C.S. Altered balance of interleukin-13/interferon-γ contributes to lacrimal gland destruction and secretory dysfunction in CD25 knockout model of Sjogren’s syndrome. Arthritis Res. Ther. 2015, 17, 53. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, K.; Monroy, D.; Pflugfelder, S.C. Cholinergic stimulation of lactoferrin and epidermal growth factor secretion by the human lacrimal gland. Cornea 1996, 15, 617–621. [Google Scholar] [CrossRef] [PubMed]
- Pelegrino, F.S.; Volpe, E.A.; Gandhi, N.B.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Deletion of interferon-γ delays onset and severity of dacryoadenitis in CD25KO mice. Arthritis Res. Ther. 2012, 14, R234. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Nagashima, H.; Brown, V.B.; Peck, A.B.; Humphreys-Beher, M.G. Two NOD Idd-associated intervals contribute synergistically to the development of autoimmune exocrinopathy (Sjogren’s syndrome) on a healthy murine background. Arthritis Rheum. 2002, 46, 1390–1398. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Peck, A.B.; Humphreys-Beher, M.G. Progress in understanding autoimmune exocrinopathy using the non-obese diabetic mouse: An update. Crit. Rev. Oral Biol. Med. 2002, 13, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Coursey, T.G.; Li, D.Q.; de Paiva, C.S.; Tukler Henriksson, J.; Pflugfelder, S.C. Conjunctival Goblet Cells modulate dendritic cell maturation and retinoic acid producing capacity. Investig. Ophthalmol. Vis. Sci. 2016, 57, 426. [Google Scholar]
- Contreras-Ruiz, L.; Masli, S. Immunomodulatory cross-talk between conjunctival goblet cells and dendritic cells. PLoS ONE 2015, 10, e0120284. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, F.L.; Xiao, Y.; Bian, F.; Coursey, T.G.; Ko, B.Y.; Clevers, H.; de Paiva, C.S.; Pflugfelder, S.C. Goblet Cells Contribute to Ocular Surface Immune Tolerance-Implications for Dry Eye Disease. Int. J. Mol. Sci. 2017, 18, 978. [Google Scholar] [CrossRef] [PubMed]
- Marko, C.K.; Menon, B.B.; Chen, G.; Whitsett, J.A.; Clevers, H.; Gipson, I.K. Spdef null mice lack conjunctival goblet cells and provide a model of dry eye. Am. J. Pathol. 2013, 183, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Coursey, T.G.; Henriksson, J.T.; Barbosa, F.L.; de Paiva, C.S.; Pflugfelder, S.C. Interferon-γ-Induced Unfolded Protein Response in Conjunctival Goblet Cells as a Cause of Mucin Deficiency in Sjogren Syndrome. Am. J. Pathol. 2016, 186, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Posadas, L.; Hodges, R.R.; Li, D.; Shatos, M.A.; Storr-Paulsen, T.; Diebold, Y.; Dartt, D.A. Interaction of IFN-γ with cholinergic agonists to modulate rat and human goblet cell function. Mucosal Immunol. 2016, 9, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Niederkorn, J.Y.; Stern, M.E.; Pflugfelder, S.C.; de Paiva, C.S.; Corrales, R.M.; Gao, J.; Siemasko, K. Desiccating Stress Induces T Cell-Mediated Sjogren’s Syndrome-Like Lacrimal Keratoconjunctivitis. J. Immunol. 2006, 176, 3950–3957. [Google Scholar] [CrossRef] [PubMed]
- Coursey, T.G.; Gandhi, N.B.; Volpe, E.A.; Pflugfelder, S.C.; de Paiva, C.S. Chemokine receptors CCR6 and CXCR3 are necessary for CD4+ T cell mediated ocular surface disease in experimental dry eye disease. PLoS ONE 2013, 8, e78508. [Google Scholar] [CrossRef] [PubMed]
- Coursey, T.G.; Bian, F.; Zaheer, M.; Pflugfelder, S.C.; Volpe, E.A.; de Paiva, C.S. Age-related spontaneous lacrimal keratoconjunctivitis is accompanied by dysfunctional T regulatory cells. Mucosal Immunol. 2016, 10, 743. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, C.S.; Villarreal, A.L.; Corrales, R.M.; Rahman, H.T.; Chang, V.Y.; Farley, W.J.; Stern, M.E.; Niederkorn, J.Y.; Li, D.Q.; Pflugfelder, S.C. Dry Eye-Induced Conjunctival Epithelial Squamous Metaplasia Is Modulated by Interferon-γ. Investig. Ophthalmol. Vis. Sci. 2007, 48, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, C.S.; Raince, J.K.; McClellan, A.J.; Shanmugam, K.P.; Pangelinan, S.B.; Volpe, E.A.; Corrales, R.M.; Farley, W.J.; Corry, D.B.; Li, D.Q.; et al. Homeostatic control of conjunctival mucosal goblet cells by NKT-derived IL-13. Mucosal Immunol. 2011, 4, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Tazume, S.; Umehara, K.; Matsuzawa, H.; Yoshida, T.; Hashimoto, K.; Sasaki, S. Immunological function of food-restricted germfree and specific pathogen-free mice. Exp. Anim. 1991, 40, 523–528. [Google Scholar] [CrossRef]
- Al-Asmakh, M.; Zadjali, F. Use of Germ-Free Animal Models in Microbiota-Related Research. J. Microbiol. Biotechnol. 2015, 25, 1583–1588. [Google Scholar] [CrossRef] [PubMed]
- Kandori, H.; Hirayama, K.; Takeda, M.; Doi, K. Histochemical, lectin-histochemical and morphometrical characteristics of intestinal goblet cells of germfree and conventional mice. Exp. Anim. 1996, 45, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, M.; Tanaka, A.; Maehara, T.; Furukawa, S.; Nakashima, H.; Nakamura, S. T helper subsets in Sjogren’s syndrome and IgG4-related dacryoadenitis and sialoadenitis: A critical review. J. Autoimmun. 2014, 51, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, T.; Letterio, J.J.; Geiser, A.G.; Kong, L.; Ogawa, N.; Zhao, W.; Koike, T.; Fernandes, G.; Dang, H.; Talal, N. Up-regulation of cytokine mRNA, adhesion molecule proteins, and MHC class II proteins in salivary glands of TGF-β1 knockout mice: MHC class II is a factor in the pathogenesis of TGF-β1 knockout mice. J. Immunol. 1997, 158, 5527–5535. [Google Scholar] [PubMed]
- Hayashi, T.; Shimoyama, N.; Mizuno, T. Destruction of salivary and lacrimal glands by Th1-polarized reaction in a model of secondary Sjogren’s syndrome in lupus-prone female NZB× NZWF1 mice. Inflammation 2012, 35, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.; Brayer, J.; Gao, J.; Brown, V.; Killedar, S.; Yasunari, U.; Peck, A.B. A dual role for interferon-γ in the pathogenesis of Sjogren’s syndrome-like autoimmune exocrinopathy in the nonobese diabetic mouse. Scand. J. Immunol. 2004, 60, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Saito, I.; Terauchi, K.; Shimuta, M.; Nishiimura, S.; Yoshino, K.; Takeuchi, T.; Tsubota, K.; Miyasaka, N. Expression of cell adhesion molecules in the salivary and lacrimal glands of Sjogren’s syndrome. J. Clin. Lab. Anal. 1993, 7, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, K.; Fukagawa, K.; Fujihara, T.; Shimmura, S.; Saito, I.; Saito, K.; Takeuchi, T. Regulation of human leukocyte antigen expression in human conjunctival epithelium. Investig. Ophthalmol. Vis. Sci. 1999, 40, 28–34. [Google Scholar]
- Zhang, X.; Chen, W.; de Paiva, C.S.; Corrales, R.M.; Volpe, E.A.; McClellan, A.J.; Farley, W.J.; Li, D.Q.; Pflugfelder, S.C. Interferon-γ exacerbates dry eye-induced apoptosis in conjunctiva through dual apoptotic pathways. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6279–6285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, W.; de Paiva, C.S.; Volpe, E.A.; Gandhi, N.B.; Farley, W.J.; Li, D.Q.; Niederkorn, J.Y.; Stern, M.E.; Pflugfelder, S.C. Desiccating Stress Induces CD4+ T-Cell-Mediated Sjogren’s Syndrome-Like Corneal Epithelial Apoptosis via Activation of the Extrinsic Apoptotic Pathway by Interferon-γ. Am. J. Pathol. 2011, 179, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; de Paiva, C.S.; Su, Z.; Volpe, E.A.; Li, D.Q.; Pflugfelder, S.C. Topical interferon-γ neutralization prevents conjunctival goblet cell loss in experimental murine dry eye. Exp. Eye Res. 2014, 118, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Youinou, P.; Pers, J.O. Disturbance of cytokine networks in Sjogren’s syndrome. Arthritis Res. Ther. 2011, 13, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roescher, N.; Tak, P.P.; Illei, G.G. Cytokines in Sjogren’s syndrome: Potential therapeutic targets. Ann. Rheum. Dis. 2010, 69, 945–948. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Kim, J.; Lee, J.H.; Sim, J.H.; Cho, D.H.; Bae, I.H.; Lee, H.; Seol, M.A.; Shin, H.M.; Kim, T.J.; et al. Modulation of gut microbiota and delayed immunosenescence as a result of syringaresinol consumption in middle-aged mice. Sci. Rep. 2016, 6, 39026. [Google Scholar] [CrossRef] [PubMed]
- Brookes, S.M.; Price, E.J.; Venables, P.J.; Maini, R.N. Interferon-γ and epithelial cell activation in Sjogren’s syndrome. Br. J. Rheumatol. 1995, 34, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Bulosan, M.; Pauley, K.M.; Yo, K.; Chan, E.K.; Katz, J.; Peck, A.B.; Cha, S. Inflammatory caspases are critical for enhanced cell death in the target tissue of Sjogren’s syndrome before disease onset. Immunol. Cell Biol. 2008, 87, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A.; Lamey, P.J.; Jarrett, R.F.; Onions, D.E. A model to study viral and cytokine involvement in Sjogren’s syndrome. Autoimmunity 1994, 18, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Daniels, P.J.; Gustafson, S.A.; French, D.; Wang, Y.; DePond, W.; McArthur, C.P. Interferon-mediated block in cell cycle and altered integrin expression in a differentiated salivary gland cell line (HSG) cultured on Matrigel. J. Interferon Cytokine Res. 2000, 20, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Fox, R.I.; Kang, H.I. Pathogenesis of Sjogren’s syndrome. Rheum. Dis. Clin. N. Am. 1992, 18, 517–538. [Google Scholar]
- Jin, J.O.; Kawai, T.; Cha, S.; Yu, Q. Interleukin-7 enhances the Th1 response to promote the development of Sjogren’s syndrome-like autoimmune exocrinopathy in mice. Arthritis Rheum. 2013, 65, 2132–2142. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, N.; Ping, L.; Zhenjun, L.; Takada, Y.; Sugai, S. Involvement of the interferon-γ-induced T cell-attracting chemokines, interferon-γ-inducible 10-kd protein (CXCL10) and monokine induced by interferon-γ (CXCL9), in the salivary gland lesions of patients with Sjogren’s syndrome. Arthritis Rheum. 2002, 46, 2730–2741. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.J.; Chen, Z.J.; Tsokos, M.; O’Connell, B.C.; Ambudkar, I.S.; Baum, B.J. Interferon-γ induced cell death in a cultured human salivary gland cell line. J. Cell Physiol. 1996, 167, 297–304. [Google Scholar] [CrossRef]
- Rahimy, E.; Pitcher, J.D., III; Pangelinan, S.B.; Chen, W.; Farley, W.J.; Niederkorn, J.Y.; Stern, M.E.; Li, D.Q.; Pflugfelder, S.C.; de Paiva, C.S. Spontaneous autoimmune dacryoadenitis in aged CD25KO mice. Am. J. Pathol. 2010, 177, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Aroniadis, O.C.; Brandt, L.J.; Kelly, C.; Freeman, S.; Surawicz, C.; Broussard, E.; Stollman, N.; Giovanelli, A.; Smith, B.; et al. The Long-term Efficacy and Safety of Fecal Microbiota Transplant for Recurrent, Severe, and Complicated Clostridium difficile Infection in 146 Elderly Individuals. J. Clin. Gastroenterol. 2015, 50, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Colman, R.J.; Rubin, D.T. Fecal microbiota transplantation as therapy for inflammatory bowel disease: A systematic review and meta-analysis. J. Crohn’s Colitis 2014, 8, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- Kelly, C.R.; Ihunnah, C.; Fischer, M.; Khoruts, A.; Surawicz, C.; Afzali, A.; Aroniadis, O.; Barto, A.; Borody, T.; Giovanelli, A.; et al. Fecal microbiota transplant for treatment of Clostridium difficile infection in immunocompromised patients. Am. J. Gastroenterol. 2014, 109, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- de Paiva, C.S.; Chotikavanich, S.; Pangelinan, S.B.; Pitcher, J.D., III; Fang, B.; Zheng, X.; Ma, P.; Farley, W.J.; Siemasko, K.S.; Niederkorn, J.Y.; et al. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009, 2, 243–253. [Google Scholar] [CrossRef] [PubMed]
- You, I.C.; Bian, F.; Volpe, E.A.; de Paiva, C.S.; Pflugfelder, S.C. Age-related conjunctival disease in the C57BL/6.NOD-Aec1Aec2 Mouse Model of Sjogren Syndrome develops independent of lacrimal dysfunction. Investig. Ophthalmol. Vis. Sci. 2015, 56, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.A.; Siracusa, M.C.; Abt, M.C.; Kim, B.S.; Kobuley, D.; Kubo, M.; Kambayashi, T.; Larosa, D.F.; Renner, E.D.; Orange, J.S.; et al. Commensal bacteria-derived signals regulate basophil hematopoiesis and allergic inflammation. Nat. Med. 2012, 18, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Britton, R.A.; Irwin, R.; Quach, D.; Schaefer, L.; Zhang, J.; Lee, T.; Parameswaran, N.; McCabe, L.R. Probiotic L. reuteri treatment prevents bone loss in a menopausal ovariectomized mouse model. J. Cell. Physiol. 2014, 229, 1822–1830. [Google Scholar] [CrossRef] [PubMed]
- Auchtung, J.M.; Robinson, C.D.; Britton, R.A. Cultivation of stable, reproducible microbial communities from different fecal donors using minibioreactor arrays (MBRAs). Microbiome 2015, 3, 42. [Google Scholar] [CrossRef] [PubMed]
- Collins, J.; Auchtung, J.M.; Schaefer, L.; Eaton, K.A.; Britton, R.A. Humanized microbiota mice as a model of recurrent Clostridium difficile disease. Microbiome 2015, 3, 35. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the MiSeq Illumina sequencing platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Sex | CON | GF | p Value | ||||
|---|---|---|---|---|---|---|---|---|
| F | M | F | M | CON vs. GF | F vs. M (GF) | |||
| Ocular surface | OGD intensity (gray levels) | 241 ± 75.6 | 228.2 ± 71.5 | 344 ± 118 | 224 ± 147 | ↑, p < 0.001 (F) | F > M (p < 0.01) | |
| Goblet cell (cells/mm) | 56.5 ± 8.8 | 53.2 ± 09.5 | 46.2 ± 7.4 | 43.0 ± 7.0 | ↓, p < 0.001 (F,M) | |||
| EGF (pg/mL) | 4282 ± 1288 | 6667 ± 1787 | 2250 ± 750 | 2257 ± 167 | ↓, p < 0.05 (F,M) | |||
| LG | Total LG infiltration (%) * | 3.5 ± 3.0 | 1.38 ± 0.4 | 6.36 ± 1.7 | 4.07 ± 2.2 | ↑, p < 0.05 (F,M) | F > M (p < 0.05) | |
| CD4+ T cells (%) (flow cytometry) | 6.0 ± 2.8 | 7.5 ± 3.7 | 10.9 ± 4.2 | 11.8 ± 4.8 | ↑, p < 0.05 (F,M) | |||
| CD8+ T cells (%) (flow cytometry) | 4.63 ± 1.2 | 3.1 ± 0.9 | 10.1 ± 0.6 | 7.8 ± 2.3 | ↑, p < 0.001 (F,M) | |||
| PCR (fold) | IFN-γ | 1.00 ± 0.25 | 0.96 ± 0.11 | 2.84 ± 0.57 | 1.31 ± 0.23 | ↑, p < 0.001 (F) | F > M (p < 0.05) | |
| MHC II | 1.00 ± 0.09 | 0.92 ± 0.16 | 1.57 ± 0.10 | 0.89 ± 0.30 | ↑, p < 0.001 (F) | F > M (p < 0.05) | ||
| Caspase 3 | 1.00 ± 0.04 | 0.57 ± 0.07 | 1.51 ± 0.10 | 1.00 ± 0.08 | ↑, p < 0.001 (F) | F > M (p < 0.001) | ||
| ↑, p < 0.01 (M) | ||||||||
| IL-1β | 1.00 ± 0.21 | 0.93 ± 0.03 | 0.76 ± 0.18 | 0.61 ± 0.14 | ↓, p < 0.05 (F,M) | |||
| IL-12 | 1.00 ± 0.08 | 1.00 ± 0.07 | 2.51 ± 0.7 | 2.28 ± 0.04 | ↑, p < 0.05 (F,M) | F > M (p < 0.01) | ||
| TNF-α | 1.00 ± 0.04 | 0.29 ± 0.21 | 0.80 ± 0.10 | 0.55 ± 0.12 | Ø | Ø | ||
| Caspase 3 activity (OD) | 2355 ± 871 | 2754 ± 687 | 7499 ± 693 | 4110 ± 1201 | ↑, p < 0.001 (F) | F > M (p < 0.001) | ||
| ↑, p < 0.01 (M) | ||||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Zaheer, M.; Bian, F.; Quach, D.; Swennes, A.G.; Britton, R.A.; Pflugfelder, S.C.; De Paiva, C.S. Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice. Int. J. Mol. Sci. 2018, 19, 565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020565
Wang C, Zaheer M, Bian F, Quach D, Swennes AG, Britton RA, Pflugfelder SC, De Paiva CS. Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice. International Journal of Molecular Sciences. 2018; 19(2):565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020565
Chicago/Turabian StyleWang, Changjun, Mahira Zaheer, Fang Bian, Darin Quach, Alton G. Swennes, Robert A. Britton, Stephen C. Pflugfelder, and Cintia S. De Paiva. 2018. "Sjögren-Like Lacrimal Keratoconjunctivitis in Germ-Free Mice" International Journal of Molecular Sciences 19, no. 2: 565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19020565