Cold and Heat Stress Diversely Alter Both Cauliflower Respiration and Distinct Mitochondrial Proteins Including OXPHOS Components and Matrix Enzymes

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Proteome Maps of Cauliflower Mitochondria under Stress Conditions
2.2. Identification of Variable Protein Spots
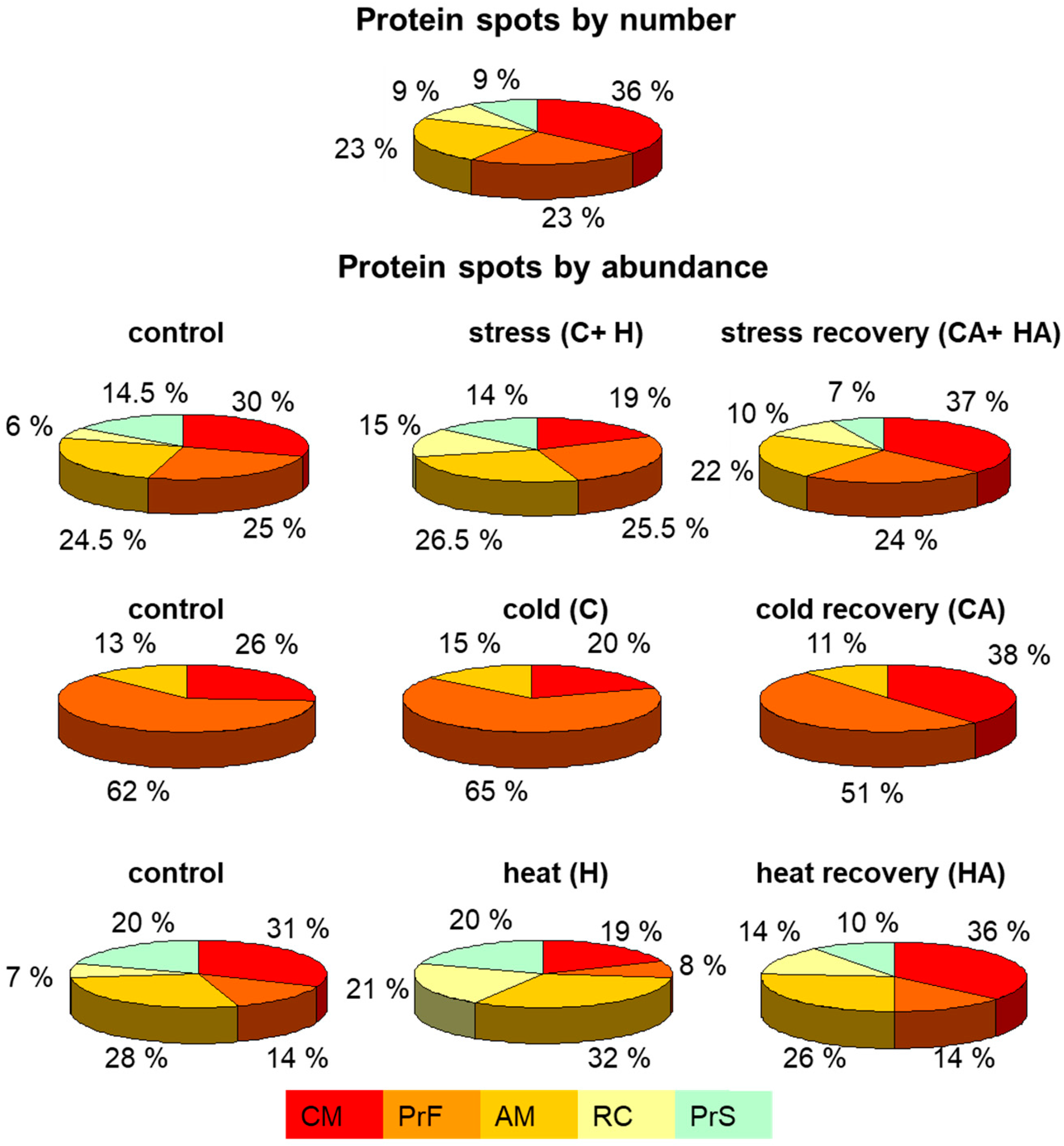
2.3. Functional Categorization of Identified Proteins
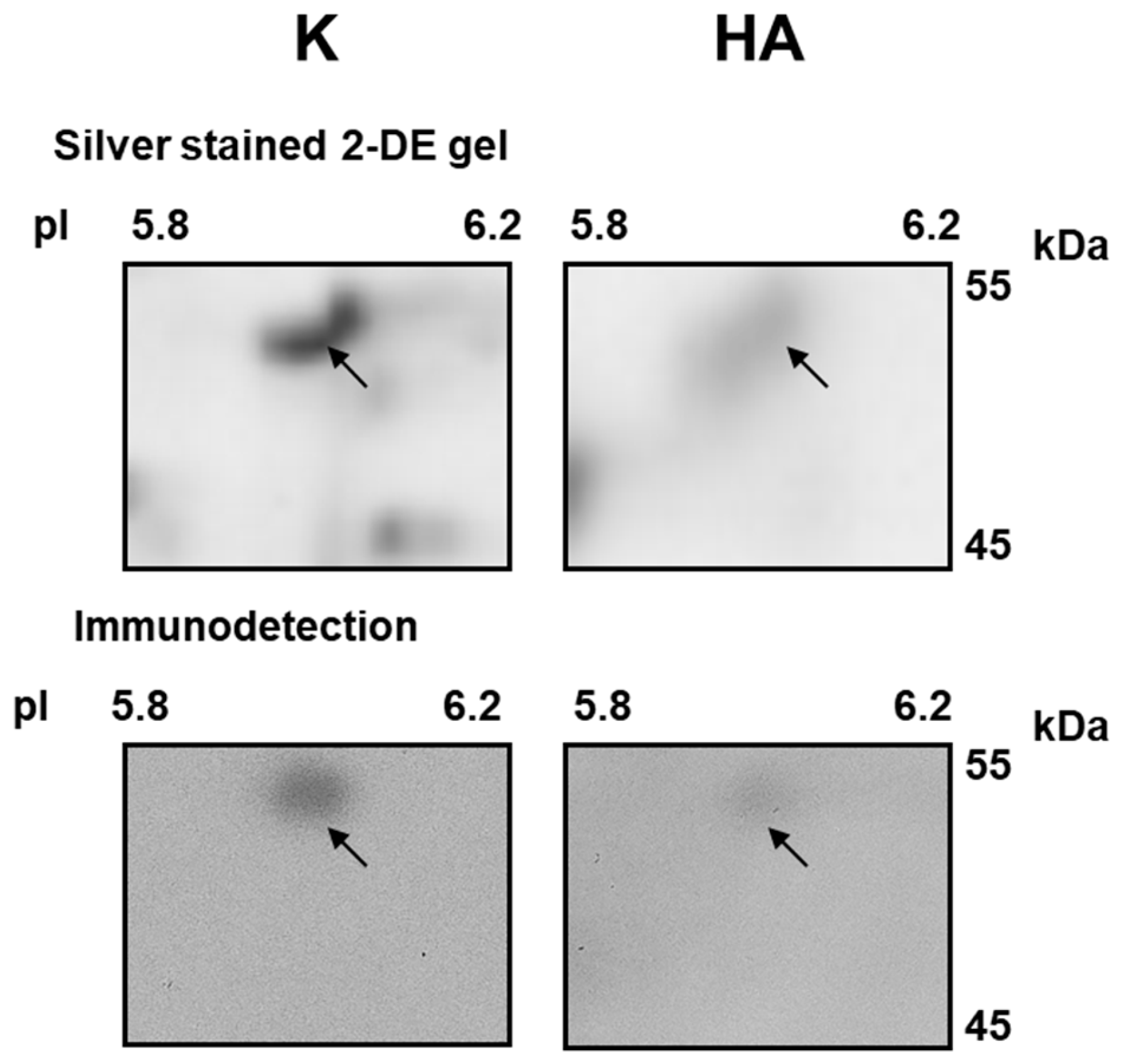
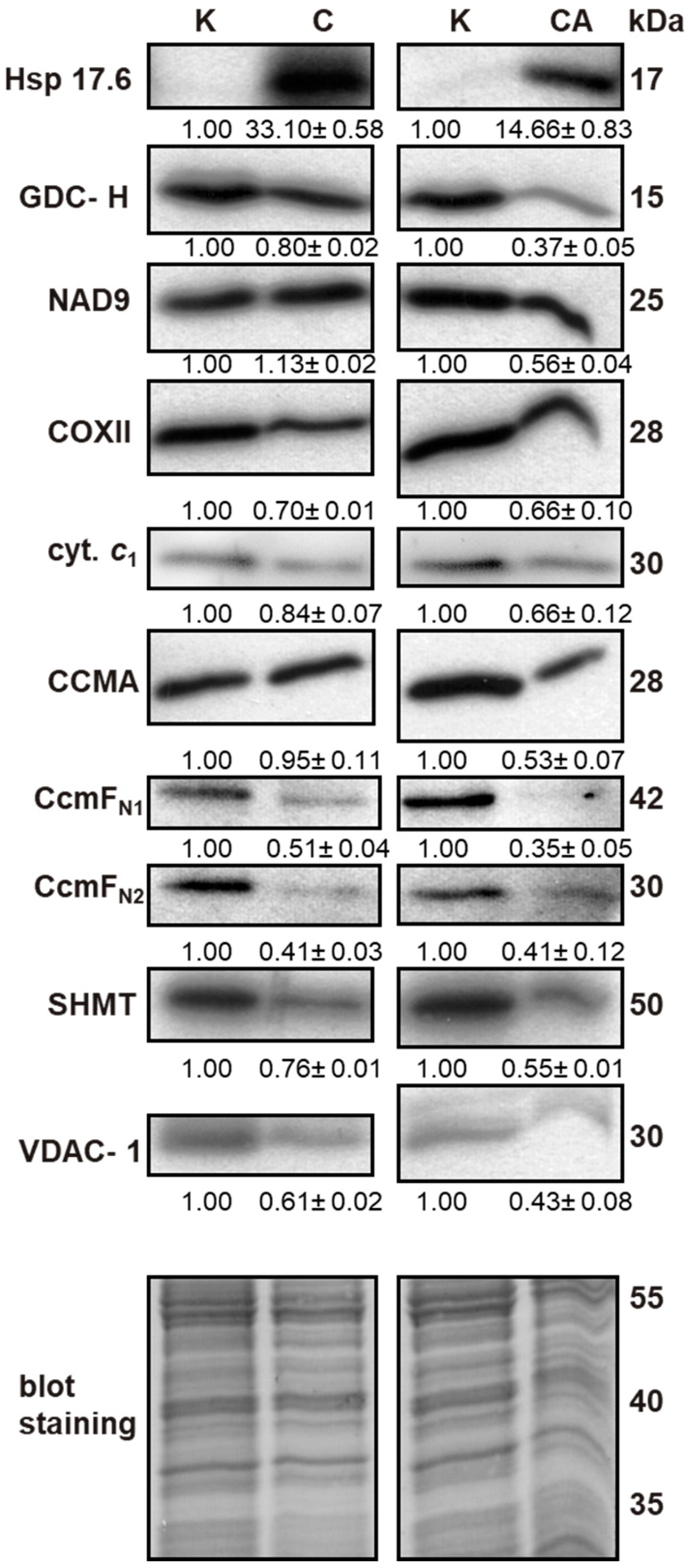
2.4. Effect of Cold Stress on Abundance of Additional Mitochondrial Proteins
2.5. Association between Protein and Transcript Level
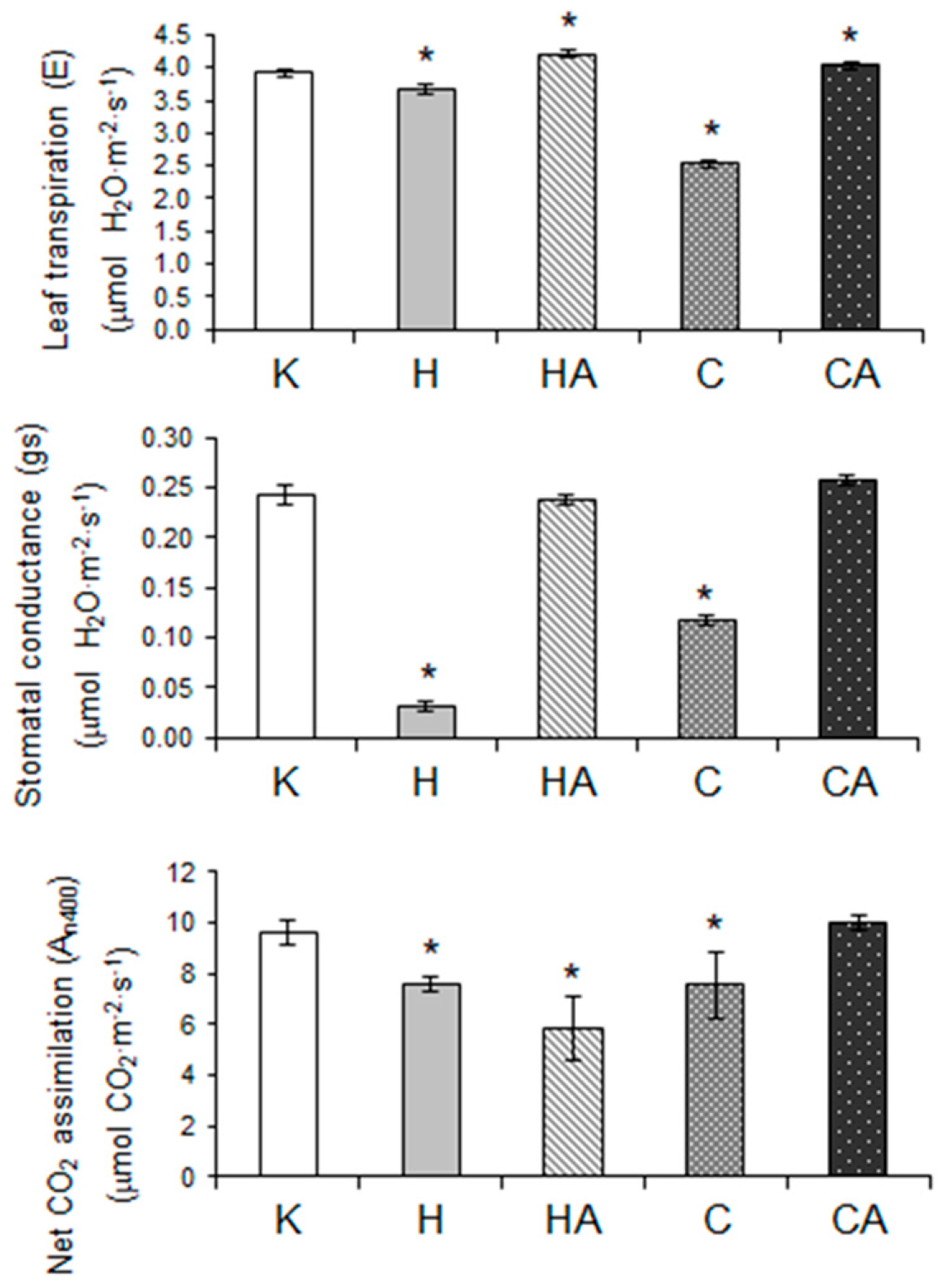
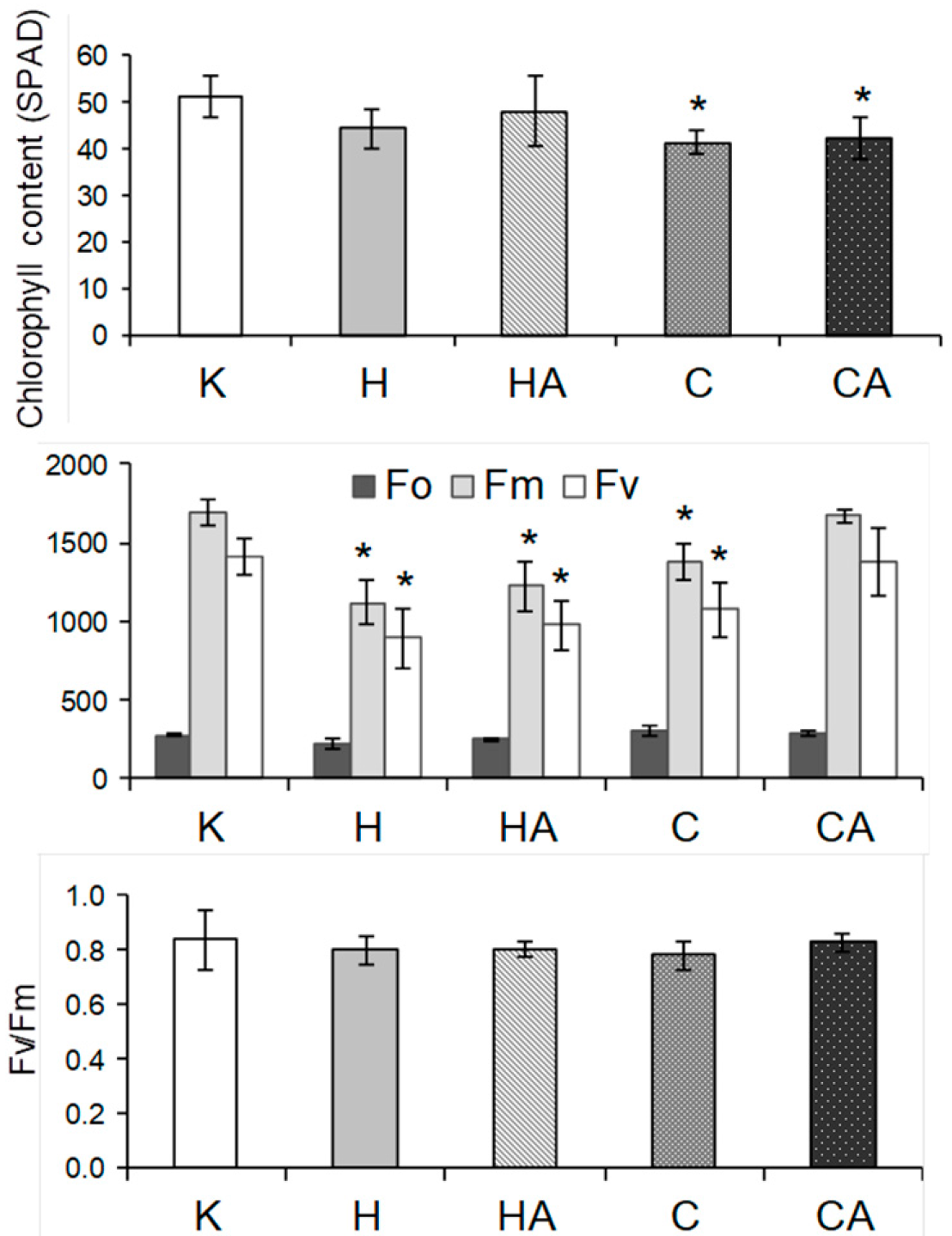
2.6. Cauliflower Physiological Responses to Cold and Heat Stress, and after Stress Recovery
3. Discussion
3.1. Identification of Cauliflower Stress-Responsive Proteins by MS Analysis
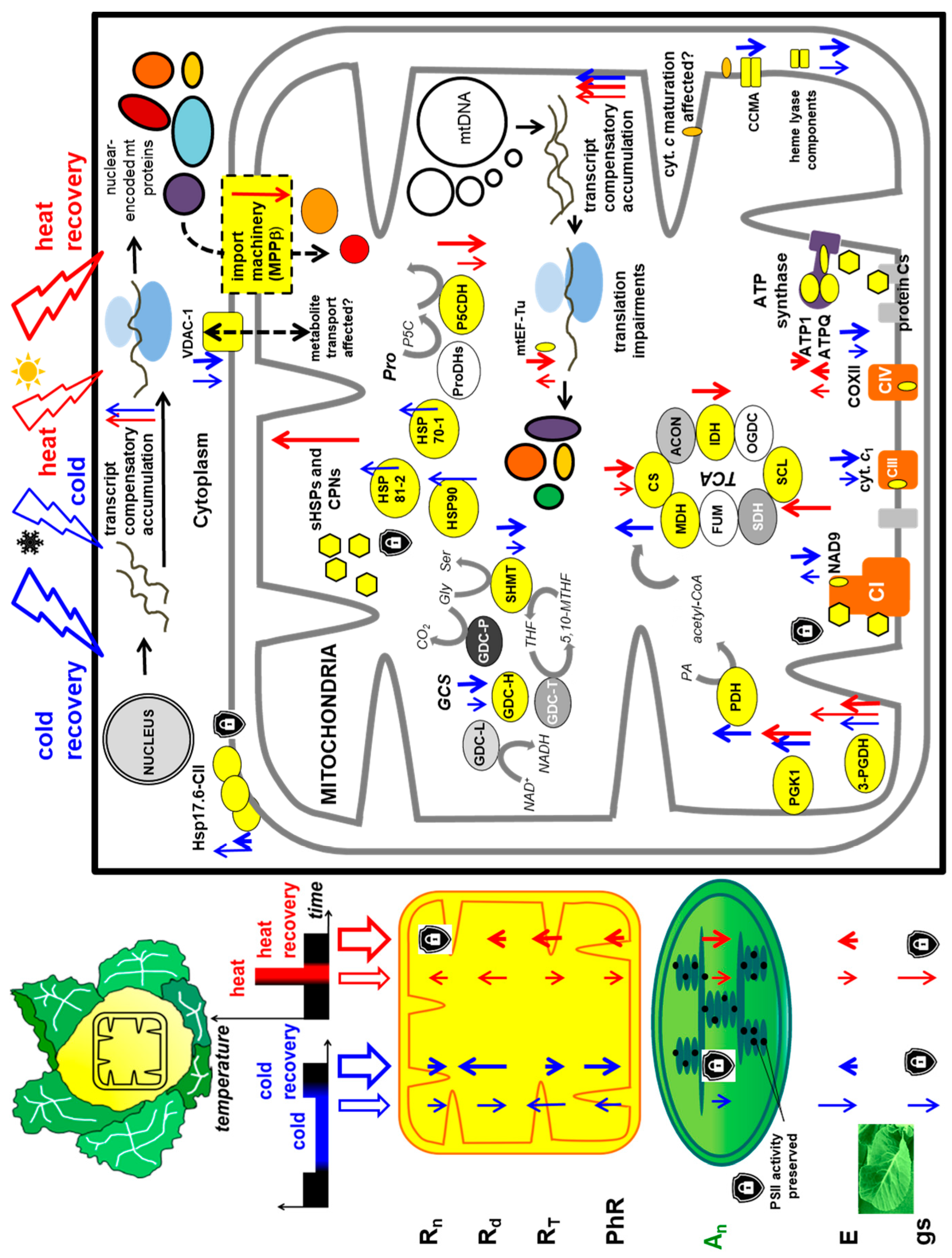
3.2. Variations in Pattern of Cauliflower Mitochondrial Proteome in Stress and Stress Recovery, and Their Metabolic Relevance
3.3. Cauliflower Leaf Respiratory Responses to Cold and Heat Stress
4. Materials and Methods
4.1. Plant Material, Growth Conditions and Stress Treatment
4.2. Gas Exchange Measurements
4.3. Chlorophyll Content and Fluorescence Measurements
4.4. Preparation of Mitochondria
4.5. Control Assays
4.6. Preparation of Samples for Two-Dimensional Electrophoresis (2D SDS-PAGE)
4.7. 2D SDS-PAGE
4.8. Statistical Analysis of 2D Protein Pattern Variations
4.9. Protein Identification by MS
4.10. SDS-PAGE and Immunoblotting
4.11. RNA Isolation and RT-semiqPCR
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| An | net CO2 assimilation rate |
| Ag | total CO2 assimilation rate |
| ANOVA | analysis of variance |
| AOX | alternative oxidase |
| ATP1 | ATP synthase subunit α |
| BSA | bovine serum albumin |
| CBB | Coomassie Brilliant Blue |
| Ccm/CCM | cytochrome c maturation |
| CAPS | 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate |
| Ci | intercellular CO2 concentration |
| COX | cytochrome c oxidase |
| CPN | chaperonin |
| CS | citrate synthase |
| 2D PAGE | two-dimensional gel electrophoresis |
| DTT | dithiothreitol |
| E | transpiration |
| EDTA | ethylenediaminetetraacetic acid |
| EGTA | ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| EF | elongation factor |
| Fm | maximal fluorescence |
| Fo | minimal fluorescence |
| FunCat | functional categorization |
| Fv | variable fluorescence |
| GDC | glycine decarboxylase |
| gs | stomatal conductance |
| HSD | honest significant difference |
| HSP | heat shock protein |
| IDH | isocitrate dehydrogenase |
| IEF | isoelectrofocusing |
| iTRAQ | isobaric tags for the absolute quantification |
| LC-MS/MS | liquid chromatography-tandem mass spectrometry |
| MDH | malate dehydrogenase |
| MIPS | Munich Information Center for Protein Sequences |
| MPP | mitochondrial processing peptidase |
| NAD | complex I subunit (mitochondrially-encoded) |
| NCBI | National Center for Biotechnology Information |
| OXPHOS | oxidative phosphorylation |
| P5CDH | Δ-1-pyrroline-5-carboxylate dehydrogenase |
| PDH | pyruvate dehydrogenase |
| PGDH | 3-phosphoglycerate dehydrogenase |
| PGK | phosphoglycerate kinase |
| PhR | photorespiration rate |
| PPFD | photosynthetic photon flux density |
| PRODH | proline dehydrogenase |
| PS | photosystem |
| PTM | posttranslational protein modification |
| RC | respiratory chain |
| Rd | respiration in the light (day respiration) rate |
| RH | relative humidity |
| Rn | respiration in the dark (night respiration) rate |
| ROS | reactive oxygen species |
| RT | total respiration rate |
| SCL | succinyl-CoA ligase |
| SHMT | serine hydroxy-methyl aminotransferase |
| TCA | tricarboxylic acid |
| VDAC | voltage-dependent anion channel |
References
- Bray, E.A.; Bailey-Serres, J.; Weretilnyk, E. Responses to abiotic stresses. In Biochemistry and Molecular Biology of Plants, 1st ed.; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; pp. 158–1249. ISBN1 13 978-0943088372. ISBN2 10 0943088372. [Google Scholar]
- Bohnert, H.J.; Nelson, D.E.; Jensen, R.G. Adaptations to Environmental Stresses. Plant Cell 1995, 7, 1099–1111. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Pereira, A. Integrative approaches for mining transcriptional regulatory programs in Arabidopsis. Brief. Funct. Genom. Proteom. 2008, 7, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Carbo, M.; Aroca, R.; Gonzàlez-Meler, M.A.; Irigoyen, J.J.; Sánchez-Díaz, M. The Electron Partitioning between the Cytochrome and Alternative Respiratory Pathways during Chilling Recovery in Two Cultivars of Maize Differing in Chilling Sensitivity. Plant Physiol. 2000, 122, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Kurimoto, K.; Millar, A.H.; Lambers, H.; Day, D.A.; Noguchi, K. Maintenance of Growth Rate at Low Temperature in Rice and Wheat Cultivars with a High Degree of Respiratory Homeostasis is Associated with a High Efficiency of Respiratory ATP Production. Plant Cell Physiol. 2004, 45, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.F.; Logan, D.C.; Tobin, A.K.; O’Toole, P.; Atkin, O.K. Heterogeneity of plant mitochondrial responses underpinning respiratory acclimation to the cold in Arabidopsis thaliana leaves. Plant Cell Environ. 2006, 29, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.-T.; Chen, S.-L.; Lin, C.-Y.; Chen, Y.-M. Chilling stress suppresses chloroplast development and nuclear gene expression in leaves of mung bean seedlings. Planta 2005, 221, 374–385. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Grimm, B.; Tripathy, B.C. Light and dark modulation of chlorophyll biosynthetic genes in response to temperature. Planta 2006, 224, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Mohanty, S.; Tripathy, B.C. Role of Temperature Stress on Chloroplast Biogenesis and Protein Import in Pea. Plant Physiol. 2009, 150, 1050–1061. [Google Scholar] [CrossRef] [PubMed]
- Lütz, C. Cell physiology of plants growing in cold environments. Protoplasma 2010, 244, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.R. Chilling-enhanced photooxidation: The production, action and study of reactive oxygen species produced during chilling in the light. Photosynth. Res. 1995, 45, 79–97. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.F.; Badger, M.R.; Day, D.A.; Barthet, M.M.; Smith, P.M.C.; Millar, A.H.; Whelan, J.; Atkin, O.K. Dynamic changes in the mitochondrial electron transport chain underpinning cold acclimation of leaf respiration. Plant Cell Environ. 2008, 31, 1156–1169. [Google Scholar] [CrossRef] [PubMed]
- Talts, P.; Pärnik, T.; Gardeström, P.; Keerberg, O. Respiratory acclimation in Arabidopsis thaliana leaves at low temperature. J. Plant Physiol. 2004, 161, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Díaz, M.; de Haro, V.; Muñoz, R.; Quiles, M.J. Chlororespiration is involved in the adaptation of Brassica plants to heat and high light intensity. Plant Cell Environ. 2007, 30, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Pastenes, C.; Horton, P. Effect of High Temperature on Photosynthesis in Beans (I. Oxygen Evolution and Chlorophyll Fluorescence). Plant Physiol. 1996, 112, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Short-term responses of Photosystem I to heat stress. Photosynth. Res. 1996, 47, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Bukhov, N.G.; Samson, G.; Carpentier, R. Nonphotosynthetic Reduction of the Intersystem Electron Transport Chain of Chloroplasts Following Heat Stress. Steady-State Rate. Photochem. Photobiol. 2000, 72, 351–357. [Google Scholar] [CrossRef]
- Bukhov, N.G.; Wiese, C.; Neimanis, S.; Heber, U. Heat sensitivity of chloroplasts and leaves: Leakage of protons from thylakoids and reversible activation of cyclic electron transport. Photosynth. Res. 1999, 59, 81–93. [Google Scholar] [CrossRef]
- Hossain, Z.; Nouri, M.-Z.; Komatsu, S. Plant cell organelle proteomics in response to abiotic stress. J. Proteome Res. 2012, 11, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Bardel, J.; Louwagie, M.; Jaquinod, M.; Jourdain, A.; Luche, S.; Rabilloud, T.; Macherel, D.; Garin, J.; Bourguignon, J. A survey of the plant mitochondrial proteome in relation to development. Proteomics 2002, 2, 880–898. [Google Scholar] [CrossRef]
- Heazlewood, J.L.; Howell, K.A.; Whelan, J.; Millar, A.H. Towards an Analysis of the Rice Mitochondrial Proteome. Plant Physiol. 2003, 132, 230–242. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, T.; Rurek, M.; Janicka, S.; Raczynska, K.D.; Augustyniak, H. Preliminary analysis of the cauliflower mitochondrial proteome. Acta Physiol. Plant. 2005, 27, 275–281. [Google Scholar] [CrossRef]
- Schmidt, U.G.; Endler, A.; Schelbert, S.; Brunner, A.; Schnell, M.; Neuhaus, H.E.; Marty-Mazars, D.; Marty, F.; Baginsky, S.; Martinoia, E. Novel Tonoplast Transporters Identified Using a Proteomic Approach with Vacuoles Isolated from Cauliflower Buds. Plant Physiol. 2007, 145, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Donnart, T.; Nouri, M.-Z.; Komatsu, S. Tissue-Specific Defense and Thermo-Adaptive Mechanisms of Soybean Seedlings under Heat Stress Revealed by Proteomic Approach. J. Proteome Res. 2010, 9, 4189–4204. [Google Scholar] [CrossRef] [PubMed]
- Koehler, G.; Wilson, R.C.; Goodpaster, J.V.; Sønsteby, A.; Lai, X.; Witzmann, F.A.; You, J.-S.; Rohloff, J.; Randall, S.K.; Alsheikh, M. Proteomic Study of Low-Temperature Responses in Strawberry Cultivars (Fragaria × ananassa) That Differ in Cold tolerance. Plant Physiol. 2012, 159, 1787–1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Lee, H.; Xiong, L.; Zhu, J.-K. A Mitochondrial Complex I Defect Impairs Cold-Regulated Nuclear Gene Expression. Plant Cell 2002, 14, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Heazlewood, J.L.; Kristensen, B.K.; Braun, H.-P.; Møller, I.M. The plant mitochondrial proteome. Trends Plant Sci. 2005, 10, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Van Aken, O.; Zhang, B.; Carrie, C.; Uggalla, V.; Paynter, E.; Giraud, E.; Whelan, J. Defining the Mitochondrial Stress Response in Arabidopsis thaliana. Mol. Plant 2009, 2, 1310–1324. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Liu, J.; Li, Y.; Shi, T. Integrative Identification of Arabidopsis Mitochondrial Proteome and Its Function Exploitation through Protein Interaction Network. PLoS ONE 2011, 6, e16022. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Tan, Y.-F.; Jacoby, R.P.; Millar, A.H. Abiotic environmental stress induced changes in the Arabidopsis thaliana chloroplast, mitochondria and peroxisome proteomes. J. Proteom. 2009, 72, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Ndimba, B.K.; Chivasa, S.; Simon, W.J.; Slabas, A.R. Identification of Arabidopsis salt and osmotic stress responsive proteins using two-dimensional difference gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 4185–4196. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, B.; Harris, N.S.; Deyholos, M.K. Comparative proteomic analysis of NaCl stress-responsive proteins in Arabidopsis roots. J. Exp. Bot. 2007, 58, 3591–3607. [Google Scholar] [CrossRef] [PubMed]
- Kosová, K.; Vítámvás, P.; Prášil, I.T.; Renaut, J. Plant proteome changes under abiotic stress—Contribution of proteomics studies to understanding plant stress response. J. Proteom. 2011, 74, 1301–1322. [Google Scholar] [CrossRef] [PubMed]
- Dunkley, T.P.J.; Hester, S.; Shadforth, I.P.; Runions, J.; Weimar, T.; Hanton, S.L.; Griffin, J.L.; Bessant, C.; Brandizzi, F.; Hawes, C.; et al. Mapping the Arabidopsis organelle proteome. Proc. Natl. Acad. Sci. USA 2006, 103, 6518–6523. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Dai, S.; McClung, S.; Yan, X.; Chen, S. Functional Differentiation of Brassica napus Guard Cells and Mesophyll Cells Revealed by Comparative Proteomics. Mol. Cell. Proteom. 2009, 8, 752–766. [Google Scholar] [CrossRef] [PubMed]
- Gammulla, C.G.; Pascovici, D.; Atwell, B.J.; Haynes, P.A. Differential proteomic response of rice (Oryza sativa) leaves exposed to high- and low-temperature stress. Proteomics 2011, 11, 2839–2850. [Google Scholar] [CrossRef] [PubMed]
- Neilson, K.A.; Mariani, M.; Haynes, P.A. Quantitative proteomic analysis of cold-responsive proteins in rice. Proteomics 2011, 11, 1696–1706. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.-F.; Millar, A.H.; Taylor, N.L. Components of Mitochondrial Oxidative Phosphorylation Vary in Abundance Following Exposure to Cold and Chemical Stresses. J. Proteome Res. 2012, 11, 3860–3879. [Google Scholar] [CrossRef] [PubMed]
- Rurek, M. Plant mitochondria under a variety of temperature stress conditions. Mitochondrion 2014, 19, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Rurek, M.; Woyda-Ploszczyca, A.M.; Jarmuszkiewicz, W. Biogenesis of mitochondria in cauliflower (Brassica oleracea var. botrytis) curds subjected to temperature stress and recovery involves regulation of the complexome, respiratory chain activity, organellar translation and ultrastructure. Biochi. Biophys. Acta-Bioenerg. 2015, 1847, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Heidarvand, L.; Millar, A.H.; Taylor, N.L. Responses of the Mitochondrial Respiratory System to Low Temperature in Plants. Crit. Rev. Plant Sci. 2017, 36, 217–240. [Google Scholar] [CrossRef]
- Giegé, P.; Sweetlove, L.J.; Cognat, V.; Leaver, C.J. Coordination of Nuclear and Mitochondrial Genome Expression during Mitochondrial Biogenesis in Arabidopsis. Plant Cell 2005, 17, 1497–1512. [Google Scholar] [CrossRef] [PubMed]
- Millenaar, F.F.; Lambers, H. The Alternative Oxidase: In vivo Regulation and Function. Plant Biol. 2003, 5, 2–15. [Google Scholar] [CrossRef]
- Lambers, H.; Robinson, S.A.; Ribas-Carbo, M. Plant Respiration: From Cell to Ecosystem. In Advances in Photosynthesis and Respiration Series, 1st ed.; Lambers, H., Ribas-Carbo, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 18, pp. 1–15. ISBN 9781402035883. [Google Scholar]
- Vanlerberghe, G.C.; Cvetkovska, M.; Wang, J. Is the maintenance of homeostatic mitochondrial signaling during stress a physiological role for alternative oxidase? Physiol. Plant. 2009, 137, 392–406. [Google Scholar] [CrossRef] [PubMed]
- Grabelnych, O.I.; Sumina, O.N.; Funderat, S.P.; Pobezhimova, T.P.; Voinikov, V.K.; Kolesnichenko, A.V. The distribution of electron transport between the main cytochrome and alternative pathways in plant mitochondria during short-term cold stress and cold hardening. J. Therm. Biol. 2004, 29, 165–175. [Google Scholar] [CrossRef]
- Sugie, A.; Naydenov, N.; Mizuno, N.; Nakamura, C.; Takumi, S. Overexpression of wheat alternative oxidase gene Waox1a alters respiration capacity and response to reactive oxygen species under low temperature in transgenic Arabidopsis. Genes Genet. Syst. 2006, 81, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Popov, V.N.; Purvis, A.C.; Skulachev, V.P.; Wagner, A.M. Stress-induced changes in ubiquinone concentration and alternative oxidase in plant mitochondria. Biosci. Rep. 2001, 21, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Heazlewood, J.L.; Day, D.A.; Millar, A.H. Differential Impact of Environmental Stresses on the Pea Mitochondrial Proteome. Mol. Cell. Proteom. 2005, 4, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Rajakulendran, N.; Amirsadeghi, S.; Vanlerberghe, G.C. Impact of mitochondrial alternative oxidase expression on the response of Nicotiana tabacum to cold temperature. Physiol. Plant. 2011, 142, 339–351. [Google Scholar] [CrossRef] [PubMed]
- Fowler, S.; Thomashow, M.F. Arabidopsis Transcriptome Profiling Indicates That Multiple Regulatory Pathways Are Activated during Cold Acclimation in Addition to the CBF Cold Response Pathway. Plant Cell 2002, 14, 1675–1690. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Yang, T.; Du, T.; Huang, Y.; Chen, J.; Yan, J.; He, J.; Guan, R. Mitochondrial genome sequencing helps show the evolutionary mechanism of mitochondrial genome formation in Brassica. BMC Genom. 2011, 12, 497. [Google Scholar] [CrossRef] [PubMed]
- Rikhvanov, E.G.; Gamburg, K.Z.; Varakina, N.N.; Rusaleva, T.M.; Fedoseeva, I.V.; Tauson, E.L.; Stupnikova, I.V.; Stepanov, A.V.; Borovskii, G.B.; Voinikov, V.K. Nuclear-mitochondrial cross-talk during heat shock in Arabidopsis cell culture. Plant J. 2007, 52, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Laisk, A. Kinetics of Photosynthesis and Photorespiration in C3-Plants; Nauka: Moscow, Russia, 1977. [Google Scholar]
- Rurek, M. Diverse accumulation of several dehydrin-like proteins in cauliflower (Brassica oleracea var. botrytis), Arabidopsis thaliana and yellow lupin (Lupinus luteus) mitochondria under cold and heat stress. BMC Plant Biol. 2010, 10, 181. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.-P.; Zhang, Q.-Y.; Tang, Z.-C.; Su, W.-A.; Sun, W.-N. Comparative Proteomic Analysis Provides New Insights into Chilling Stress Responses in Rice. Mol. Cell. Proteom. 2006, 5, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Giegé, P.; Heazlewood, J.L.; Roessner-Tunali, U.; Millar, A.H.; Fernie, A.R.; Leaver, C.J.; Sweetlove, L.J. Enzymes of Glycolysis Are Functionally Associated with the Mitochondrion in Arabidopsis Cells. Plant Cell 2003, 15, 2140–2151. [Google Scholar] [CrossRef] [PubMed]
- Heazlewood, J.L.; Tonti-Filippini, J.S.; Gout, A.M.; Day, D.A.; Whelan, J.; Millar, A.H. Experimental Analysis of the Arabidopsis Mitochondrial Proteome Highlights Signaling and Regulatory Components, Provides Assessment of Targeting Prediction Programs, and Indicates Plant-Specific Mitochondrial Proteins. Plant Cell 2004, 16, 241–256. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.W.A.; Williams, T.C.R.; Morgan, M.; Fernie, A.R.; Ratcliffe, R.G.; Sweetlove, L.J. Glycolytic Enzymes Associate Dynamically with Mitochondria in Response to Respiratory Demand and Support Substrate Channeling. Plant Cell 2007, 19, 3723–3738. [Google Scholar] [CrossRef] [PubMed]
- Goulas, E.; Schubert, M.; Kieselbach, T.; Kleczkowski, L.A.; Gardeström, P.; Schröder, W.; Hurry, V. The chloroplast lumen and stromal proteomes of Arabidopsis thaliana show differential sensitivity to short- and long-term exposure to low temperature. Plant J. 2006, 47, 720–734. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Sweetlove, L.J.; Giegé, P.; Leaver, C.J. Analysis of the Arabidopsis Mitochondrial Proteome. Plant Physiol. 2001, 127, 1711–1727. [Google Scholar] [CrossRef] [PubMed]
- Kruft, V.; Eubel, H.; Jänsch, L.; Werhahn, W.; Braun, H.P. Proteomic Approach to Identify Novel Mitochondrial Proteins in Arabidopsis. Plant Physiol. 2001, 127, 1694–1710. [Google Scholar] [CrossRef] [PubMed]
- Giegé, P.; Sweetlove, L.J.; Leaver, C.J. Identification of mitochondrial protein complexes in Arabidopsis using two-dimensional blue-native polyacrylamide gel electrophoresis. Plant Mol. Biol. Rep. 2003, 21, 133–144. [Google Scholar] [CrossRef]
- Huang, S.; Taylor, N.L.; Narsai, R.; Eubel, H.; Whelan, J.; Millar, A.H. Experimental Analysis of the Rice Mitochondrial Proteome, Its Biogenesis, and Heterogeneity. Plant Physiol. 2009, 149, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Dubinin, J.; Braun, H.-P.; Schmitz, U.; Colditz, F. The mitochondrial proteome of the model legume Medicago truncatula. Biochim. Biophys. Acta 2011, 1814, 1658–1668. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Heazlewood, J.L.; Millar, A.H. The Arabidopsis thaliana 2-D gel mitochondrial proteome: Refining the value of reference maps for assessing protein abundance, contaminants and post-translational modifications. Proteomics 2011, 11, 1720–1733. [Google Scholar] [CrossRef] [PubMed]
- Ito, J.; Heazlewood, J.L.; Millar, A.H. The plant mitochondrial proteome and the challenge of defining the posttranslational modifications responsible for signalling and stress effects on respiratory functions. Physiol. Plant. 2007, 129, 207–224. [Google Scholar] [CrossRef]
- Ito, J.; Taylor, N.L.; Castleden, I.; Weckwerth, W.; Millar, A.H.; Heazlewood, J.L. A survey of the Arabidopsis thaliana mitochondrial phosphoproteome. Proteomics 2009, 9, 4229–4240. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Verrillo, F.; Renzone, G.; Arena, S.; Rocco, M.; Scaloni, A.; Marra, M. Response to biotic and oxidative stress in Arabidopsis thaliana: Analysis of variably phosphorylated proteins. J. Proteom. 2011, 74, 1934–1949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havelund, J.F.; Thelen, J.J.; Møller, I.M. Biochemistry, proteomics, and phosphoproteomics of plant mitochondria from non-photosynthetic cells. Front. Plant Sci. 2013, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Takasaki, H.; Komatsu, S. Comprehensive Phosphoproteome Analysis in Rice and Identification of Phosphoproteins Responsive to Different Hormones/Stresses. J. Proteome Res. 2005, 4, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Solheim, C.; Li, L.; Hatzopoulos, P.; Millar, A.H. Loss of Lon1 in Arabidopsis Changes the Mitochondrial Proteome Leading to Altered Metabolite Profiles and Growth Retardation without an Accumulation of Oxidative Damage. Plant Physiol. 2012, 160, 1187–1203. [Google Scholar] [CrossRef] [PubMed]
- Parkin, I.A.P.; Robinson, S.J. Exploring the Paradoxes of the Brassica Genome Architecture. In Genetics, Genomics and Breeding of Vegetable Brassicas, 1st ed.; Sadowski, J., Kole, C., Eds.; Science Publishers: Enfield, NH, USA; Boca Raton, FL, USA, 2011; pp. 328–348. ISBN 9781578087068. [Google Scholar]
- Faurobert, M.; Mihr, C.; Bertin, N.; Pawlowski, T.; Negroni, L.; Sommerer, N.; Causse, M. Major Proteome Variations Associated with Cherry Tomato Pericarp Development and Ripening. Plant Physiol. 2007, 143, 1327–1346. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Kerim, T.; Rolfe, B.G.; Weinman, J.J. Effect of early cold stress on the maturation of rice anthers. Proteomics 2004, 4, 1873–1882. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Meng, X.; Wang, Q.; Tian, S. Oxidative Damage of Mitochondrial Proteins Contributes to Fruit Senescence: A Redox Proteomics Analysis. J. Proteome Res. 2009, 8, 2449–2462. [Google Scholar] [CrossRef] [PubMed]
- Yun, Z.; Jin, S.; Ding, Y.; Wang, Z.; Gao, H.; Pan, Z.; Xu, J.; Cheng, Y.; Deng, X. Comparative transcriptomics and proteomics analysis of citrus fruit, to improve understanding of the effect of low temperature on maintaining fruit quality during lengthy post-harvest storage. J. Exp. Bot. 2012, 63, 2873–2893. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.; Hjernø, K.; Larsen, M.; Wingsle, G.; Larsen, P.; Fey, S.; Roepstorff, P.; Salomé Pais, M. Proteome Profiling of Populus euphratica Oliv. Upon Heat Stress. Ann. Bot. 2006, 98, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Howell, K.A.; Cheng, K.; Murcha, M.W.; Jenkin, L.E.; Millar, A.H.; Whelan, J. Oxygen Initiation of Respiration and Mitochondrial Biogenesis in Rice. J. Biol. Chem. 2007, 282, 15619–15631. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-G.; Ahsan, N.; Lee, S.-H.; Kang, K.Y.; Bahk, J.D.; Lee, I.-J.; Lee, B.-H. A proteomic approach in analyzing heat-responsive proteins in rice leaves. Proteomics 2007, 7, 3369–3383. [Google Scholar] [CrossRef] [PubMed]
- Millar, A.H.; Whelan, J.; Soole, K.L.; Day, D.A. Organization and regulation of mitochondrial respiration in plants. Annu. Rev. Plant Biol. 2011, 62, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Sarry, J.-E.; Kuhn, L.; Ducruix, C.; Lafaye, A.; Junot, C.; Hugouvieux, V.; Jourdain, A.; Bastien, O.; Fievet, J.B.; Vailhen, D.; et al. The early responses of Arabidopsis thaliana cells to cadmium exposure explored by protein and metabolite profiling analyses. Proteomics 2006, 6, 2180–2198. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Yamamoto, A.; Nakamura, T.; Nouri, M.-Z.; Nanjo, Y.; Nishizawa, K.; Furukawa, K. Comprehensive Analysis of Mitochondria in Roots and Hypocotyls of Soybean under Flooding Stress using Proteomics and Metabolomics Techniques. J. Proteome Res. 2011, 10, 3993–4004. [Google Scholar] [CrossRef] [PubMed]
- Nakaminami, K.; Matsui, A.; Nakagami, H.; Minami, A.; Nomura, Y.; Tanaka, M.; Morosawa, T.; Ishida, J.; Takahashi, S.; Uemura, M.; et al. Analysis of Differential Expression Patterns of mRNA and Protein During Cold-Acclimation and De-Acclimation in Arabidopsis. Mol. Cell. Proteom. 2014, 13, 3602–3611. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Xu, S.L.; Oses-Prieto, J.A.; Putil, S.; Xu, P.; Wang, R.J.; Li, K.H.; Maltby, D.A.; An, L.H.; Burlingame, A.L.; et al. Proteomics Analysis Reveals Post-Translational Mechanisms for Cold-Induced Metabolic Changes in Arabidopsis. Mol. Plant 2011, 4, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Rinalducci, S.; Egidi, M.G.; Karimzadeh, G.; Jazii, F.R.; Zolla, L. Proteomic analysis of a spring wheat cultivar in response to prolonged cold stress. Electrophoresis 2011, 32, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Dumont, E.; Bahrman, N.; Goulas, E.; Valot, B.; Sellier, H.; Hilbert, J.-L.; Vuylsteker, C.; Lejeune-Hénaut, I.; Delbreil, B. A proteomic approach to decipher chilling response from cold acclimation in pea (Pisum sativum L.). Plant Sci. 2011, 180, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.; Sun, H.; Xin, X.; Qin, G.; Liang, Z.; Jing, X. Mitochondrial Damage in the Soybean Seed Axis during Imbibition at Chilling Temperatures. Plant Cell. Physiol. 2009, 50, 1305–1318. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Gao, X.; Li, S.; Shi, M.; Javeed, H.; Jing, X.; Yang, G.; He, G. Proteomic analysis of soybean [Glycine max (L.) Meer.] seeds during imbibition at chilling temperature. Mol. Breed. 2010, 26, 1–17. [Google Scholar] [CrossRef]
- Sánchez-Bel, P.; Egea, I.; Sanchez-Ballesta, M.T.; Martinez-Madrid, C.; Fernandez-Garcia, N.; Romojaro, F.; Olmos, E.; Estrella, E.; Bolarin, M.C.; Flores, F.B. Understanding the mechanisms of chilling injury in bell pepper fruits using the proteomic approach. J. Proteom. 2012, 75, 5463–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Huang, B. Differential proteomic response to heat stress in thermal Agrostis scabra and heat-sensitive Agrostis stolonifera. Physiol. Plant. 2010, 139, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Vítámvás, P.; Prášil, I.T.; Kosová, K.; Planchon, S.; Renaut, J. Analysis of proteome and frost tolerance in chromosome 5A and 5B reciprocal substitution lines between two winter wheats during long-term cold acclimation. Proteomics 2012, 12, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Sweetlove, L.J.; Heazlewood, J.L.; Herald, V.; Holtzapffel, R.; Day, D.A.; Leaver, C.J.; Millar, A.H. The impact of oxidative stress on Arabidopsis mitochondria. Plant J. 2002, 32, 891–904. [Google Scholar] [CrossRef] [PubMed]
- Renaut, J.; Hausman, J.-F.; Bassett, C.; Artlip, T.; Cauchie, H.-M.; Witters, E.; Wisniewski, M. Quantitative proteomic analysis of short photoperiod and low-temperature responses in bark tissues of peach (Prunus persica L. Batsch). Tree Genet. Genomes 2008, 4, 589–600. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Li, J.; Jiang, A.; Cheng, Y.; Zhang, W. Mitochondrial proteome during salt stress-induced programmed cell death in rice. Plant Physiol. Biochem. 2009, 47, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Palmblad, M.; Mills, D.J.; Bindschedler, L.V. Heat-Shock Response in Arabidopsis thaliana Explored by Multiplexed Quantitative Proteomics Using Differential Metabolic Labeling. J. Proteome Res. 2008, 7, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Jiang, L.; Yu, M.; An, X.; Ma, R.; Yu, Z. Proteomic analysis of changes in mitochondrial protein expression during peach fruit ripening and senescence. J. Proteom. 2016, 147, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.J.; Jang, B.; Jung, C.-H.; Ahn, S.J.; Goh, C.-H.; Cho, K.; Han, O.; Kang, H. Functional characterization of a glycine-rich RNA-binding protein 2 in Arabidopsis thaliana under abiotic stress conditions. Plant J. 2007, 50, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Huang, F.; Wang, J.; Ma, X.; Cheng, Y.; Liu, J. A proteomic analysis of cold stress responses in rice seedlings. Proteomics 2005, 5, 3162–3172. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Lin, X.-H.; Zhou, Y.-C.; Chen, X.-L.; Liu, X.; Lu, X.-X. Proteome analysis of maize seeds: The effect of artificial ageing. Physiol. Plant. 2011, 143, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, Q.; Kong, L.; Xia, F.; Yan, H.; Zhu, Y.; Mao, P. Proteomic and Physiological Analysis of the Response of Oat (Avena sativa) Seeds to Heat Stress under Different Moisture Conditions. Front. Plant Sci. 2016, 7, 896. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Rudhe, C.; Hulett, J.M.; Lithgow, T.; Glaser, E.; Day, D.A.; Millar, A.H.; Whelan, J. Environmental stresses inhibit and stimulate different protein import pathways in plant mitochondria. FEBS Lett. 2003, 547, 125–130. [Google Scholar] [CrossRef]
- Funck, D.; Eckard, S.; Müller, G. Non-redundant functions of two proline dehydrogenase isoforms in Arabidopsis. BMC Plant Biol. 2010, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Lu, Q.; Verma, D.P. Reciprocal regulation of delta 1-pyrroline-5-carboxylate synthetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants. Mol. Gen. Genet. 1996, 253, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Honig, A.; Stein, H.; Suzuki, N.; Mittler, R.; Zilberstein, A. Unraveling Δ1-Pyrroline-5-Carboxylate-Proline Cycle in Plants by Uncoupled Expression of Proline Oxidation Enzymes. J. Biol. Chem. 2009, 284, 26482–26492. [Google Scholar] [CrossRef] [PubMed]
- Fuller, M.P.; Metwali, E.M.R.; Eed, M.H.; Jellings, A.J. Evaluation of Abiotic Stress Resistance in Mutated Populations of Cauliflower (Brassica oleracea var. botrytis). Plant Cell Tissue Organ Cult. 2006, 86, 239. [Google Scholar] [CrossRef]
- Hadi, F.; Gilpin, M.; Fuller, M.P. Identification and expression analysis of CBF/DREB1 and COR15 genes in mutants of Brassica oleracea var. botrytis with enhanced proline production and frost resistance. Plant Physiol. Biochem. 2011, 49, 1323–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, W.-T.; Lin, B.; Zhang, M.; Hua, X.-J. Proline Accumulation is Inhibitory to Arabidopsis Seedlings during Heat Stress. Plant Physiol. 2011, 156, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Lee, U.; Wie, C.; Escobar, M.; Williams, B.; Hong, S.-W.; Vierling, E. Genetic Analysis Reveals Domain Interactions of Arabidopsis Hsp100/Clpb and Cooperation with the Small Heat Shock Protein Chaperone System. Plant Cell 2005, 17, 559–571. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.L.; Day, D.A.; Millar, A.H. Environmental Stress Causes Oxidative Damage to Plant Mitochondria Leading to Inhibition of Glycine Decarboxylase. J. Biol. Chem. 2002, 277, 42663–42668. [Google Scholar] [CrossRef] [PubMed]
- Herman, E.M.; Rotter, K.; Premakumar, R.; Elwinger, G.; Bae, H.; Ehler-King, L.; Chen, S.; Livingston, D.P. Additional freeze hardiness in wheat acquired by exposure to −3 °C is associated with extensive physiological, morphological, and molecular changes. J. Exp. Bot. 2006, 57, 3601–3618. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, F.; Peng, S.; Ma, W.; Korpelainen, H.; Li, C. Comparative physiological, ultrastructural and proteomic analyses reveal sexual differences in the responses of Populus cathayana under drought stress. Proteomics 2010, 10, 2661–2677. [Google Scholar] [CrossRef] [PubMed]
- Imin, N.; Kerim, T.; Weinman, J.J.; Rolfe, B.G. Low Temperature Treatment at the Young Microspore Stage Induces Protein Changes in Rice Anthers. Mol. Cell. Proteom. 2006, 5, 274–292. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-W.; Rakwal, R.; Agrawal, G.K.; Jung, Y.-H.; Shibato, J.; Jwa, N.-S.; Iwahashi, Y.; Iwahashi, H.; Kim, D.H.; Shim, I.-S.; et al. A hydroponic rice seedling culture model system for investigating proteome of salt stress in rice leaf. Electrophoresis 2005, 26, 4521–4539. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Shan, X.H.; Wu, Y.; Su, S.Z.; Li, S.P.; Liu, H.K.; Han, J.Y.; Xue, C.M.; Yuan, Y.P. iTRAQ-based quantitative proteomic analysis reveals new metabolic pathways responding to chilling stress in maize seedlings. J. Proteom. 2016, 146, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Byun, Y.-J.; Kim, H.-J.; Lee, D.-H. LongSAGE analysis of the early response to cold stress in Arabidopsis leaf. Planta 2009, 229, 1181–1200. [Google Scholar] [CrossRef] [PubMed]
- Naydenov, N.G.; Khanam, S.; Siniauskaya, M.; Nakamura, C. Profiling of mitochondrial transcriptome in germinating wheat embryos and seedlings subjected to cold, salinity and osmotic stresses. Genes Genet. Syst. 2010, 85, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.P.; Millar, A.H.; Taylor, N.L. Wheat Mitochondrial Proteomes Provide New Links between Antioxidant Defense and Plant Salinity Tolerance. J. Proteome Res. 2010, 9, 6595–6604. [Google Scholar] [CrossRef] [PubMed]
- Heidarvand, L.; Maali-Amiri, R. Physio-biochemical and proteome analysis of chickpea in early phases of cold stress. J. Plant Physiol. 2013, 170, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Holtzapffel, R.C.; Finnegan, P.M.; Millar, A.H.; Badger, M.R.; Day, D.A. Mitochondrial protein expression in tomato fruit during on-vine ripening and cold storage. Funct. Plant Biol. 2002, 29, 827–834. [Google Scholar] [CrossRef]
- Bocian, A.; Kosmala, A.; Rapacz, M.; Jurczyk, B.; Marczak, L.; Zwierzykowski, Z. Differences in leaf proteome response to cold acclimation between Lolium perenne plants with distinct levels of frost tolerance. J. Plant Physiol. 2011, 168, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Lin, K.-H.; Huang, H.-C.; Lin, C.-Y. Cloning, expression and physiological analysis of broccoli catalase gene and Chinese cabbage ascorbate peroxidase gene under heat stress. Plant Cell Rep. 2010, 29, 575–593. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M. Relationships between growth and gas exchange characteristics in some salt-tolerant amphidiploid Brassica species in relation to their diploid parents. Environ. Exp. Bot. 2001, 45, 155–163. [Google Scholar] [CrossRef]
- Baryla, A.; Carrier, P.; Franck, F.; Coulomb, C.; Sahut, C.; Havaux, M. Leaf chlorosis in oilseed rape plants (Brassica napus) grown on cadmium-polluted soil: Causes and consequences for photosynthesis and growth. Planta 2001, 212, 696–709. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Khan, N.A.; Tuteja, N. Differential cadmium stress tolerance in five indian mustard (Brassica juncea L.) cultivars: An evaluation of the role of antioxidant machinery. Plant Signal. Behav. 2011, 6, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Maheshwari, P.; Wani, A.S.; Irfan, M.; Alyemeni, M.N.; Ahmad, A. Comparative effect of 28 homobrassinolide and salicylic acid in the amelioration of NaCl stress in Brassica juncea L. Plant Physiol. Biochem. 2012, 53, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zheng, Q.; Liu, M.; Long, X.; Liu, Z.; Shen, Q.; Guo, S. Difference in Sodium Spatial Distribution in the Shoot of Two Canola Cultivars under Saline Stress. Plant Cell. Physiol. 2012, 53, 1083–1092. [Google Scholar] [CrossRef] [PubMed]
- Bunce, J.A. Acclimation of photosynthesis to temperature in Arabidopsis thaliana and Brassica oleracea. Photosynthetica 2008, 46, 517–524. [Google Scholar] [CrossRef]
- Hu, W.H.; Shi, K.; Song, X.S.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q. Different effects of chilling on respiration in leaves and roots of cucumber (Cucumis sativus). Plant Physiol. Biochem. 2006, 44, 837–843. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Xiao, Y.A.; Zeng, J.J.; Hu, X.H. Photosynthesis, respiration and antioxidant enzymes in pepper leaves under drought and heat stresses. Biol. Plant. 2010, 54, 761–765. [Google Scholar] [CrossRef]
- Dahal, K.; Kane, K.; Gadapati, W.; Webb, E.; Savitch, L.V.; Singh, J.; Sharma, P.; Sarhan, F.; Longstaffe, F.J.; Grodzinski, B.; et al. The effects of phenotypic plasticity on photosynthetic performance in winter rye, winter wheat and Brassica napus. Physiol. Plant. 2012, 144, 169–188. [Google Scholar] [CrossRef] [PubMed]
- Savitch, L.V.; Barker-Astrom, J.; Ivanov, A.G.; Hurry, V.; Oquist, G.; Huner, N.P.; Gardeström, P. Cold acclimation of Arabidopsis thaliana results in incomplete recovery of photosynthetic capacity, associated with an increased reduction of the chloroplast stroma. Planta 2001, 214, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Copolovici, L.; Kännaste, A.; Pazouki, L.; Niinemets, U. Emissions of green leaf volatiles and terpenoids from Solanum lycopersicum are quantitatively related to the severity of cold and heat shock treatments. J. Plant Physiol. 2012, 169, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jiang, H.; Peng, S.; Korpelainen, H.; Li, C. Sex-related differences in morphological, physiological, and ultrastructural responses of Populus cathayana to chilling. J. Exp. Bot. 2011, 62, 675–686. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.-B.; Ma, L.; Xi, H.-F.; Duan, W.; Li, S.-H.; Loescher, W.; Wang, J.-F.; Wang, L.-J. Photosynthetic Responses to Heat Treatments at Different Temperatures and Following Recovery in Grapevine (Vitis amurensis L.) Leaves. PLoS ONE 2011, 6, e23033. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; wei Meng, Q.; Brestic, M.; Olsovska, K.; Yang, X. Photosynthesis is improved by exogenous calcium in heat-stressed tobacco plants. J. Plant Physiol. 2011, 168, 2063–2071. [Google Scholar] [CrossRef] [PubMed]
- Efeoğlu, B.; Terzioğlu, S. Photosynthetic responses of two wheat varieties to high temperature. EurAsian J. BioSci. 2009, 3, 97–106. [Google Scholar] [CrossRef]
- Yin, Y.; Li, S.; Liao, W.; Lu, Q.; Wen, X.; Lu, C. Photosystem II photochemistry, photoinhibition, and the xanthophyll cycle in heat-stressed rice leaves. J. Plant Physiol. 2010, 167, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Hüve, K.; Bichele, I.; Rasulov, B.; Niinemets, U. When it is too hot for photosynthesis: Heat-induced instability of photosynthesis in relation to respiratory burst, cell permeability changes and H2O2 formation. Plant Cell Environ. 2011, 34, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial Malate Dehydrogenase Lowers Leaf Respiration and Alters Photorespiration and Plant Growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef] [PubMed]
- Boutry, M.; Faber, A.-M.; Charbonnier, M.; Briquet, M. Microanalysis of plant mitochondrial protein synthesis products. Plant Mol. Biol. 1984, 3, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Staszak, A.M.; Pawłowski, T.A. Proteomic Analysis of Embryogenesis and the Acquisition of Seed Dormancy in Norway Maple (Acer platanoides L.). Int. J. Mol. Sci. 2014, 15, 10868–10891. [Google Scholar] [CrossRef] [PubMed]
- Pawłowski, T.A.; Staszak, A.M. Analysis of the embryo proteome of sycamore (Acer pseudoplatanus L.) seeds reveals a distinct class of proteins regulating dormancy release. J. Plant Physiol. 2016, 195, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Ramagli, L.S.; Rodriguez, L.V. Quantitation of microgram amounts of protein in two-dimensional polyacrylamide gel electrophoresis sample buffer. Electrophoresis 1985, 6, 559–563. [Google Scholar] [CrossRef]
- Heukeshoven, J.; Dernick, R. Silver staining of proteins. In Electrophoresis Forum, 1st ed.; Radola, B.J., Ed.; Technische Universität München: Munich, Germany, 1986. [Google Scholar]
- Neuhoff, V.; Arold, N.; Taube, D.; Ehrhardt, W. Improved staining of proteins in polyacrylamide gels including isoelectric focusing gels with clear background at nanogram sensitivity using Coomassie Brilliant Blue G-250 and R-250. Electrophoresis 1988, 9, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Durek, P.; Schmidt, R.; Heazlewood, J.L.; Jones, A.; MacLean, D.; Nagel, A.; Kersten, B.; Schulze, W.X. PhosPhAt: The Arabidopsis thaliana phosphorylation site database. An update. Nucleic Acids Res. 2010, 38, D828–D834. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Gammeltoft, S.; Brunak, S. Sequence and structure-based prediction of eukaryotic protein phosphorylation sites. J. Mol. Biol. 1999, 294, 1351–1362. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Thelen, J.J.; Dunker, A.K.; Xu, D. Musite, a Tool for Global Prediction of General and Kinase-specific Phosphorylation Sites. Mol. Cell. Proteom. 2010, 9, 2586–2600. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.-P.; Qiu, J.-D.; Sun, X.-Y.; Suo, S.-B.; Huang, S.-Y.; Liang, R.-P. PMeS: Prediction of Methylation Sites Based on Enhanced Feature Encoding Scheme. PLoS ONE 2012, 7, e38772. [Google Scholar] [CrossRef] [PubMed]
- Zybailov, B.; Sun, Q.; van Wijk, K.J. Workflow for Large Scale Detection and Validation of Peptide Modifications by RPLC-LTQ-Orbitrap: Application to the Arabidopsis thaliana Leaf Proteome and an Online Modified Peptide Library. Anal. Chem. 2009, 81, 8015–8024. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Rudella, A.; Ramirez Rodriguez, V.; Zybailov, B.; Olinares, P.D.B.; van Wijk, K.J. Subunits of the Plastid ClpPR Protease Complex Have Differential Contributions to Embryogenesis, Plastid Biogenesis, and Plant Development in Arabidopsis. Plant Cell 2009, 21, 1669–1692. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Gooley, A.A.; Herbert, B.R.; Molloy, M.P.; Binz, P.A.; Ou, K.; Sanchez, J.C.; Bairoch, A.; Williams, K.L.; et al. High-throughput mass spectrometric discovery of protein post-translational modifications. J. Mol. Biol. 1999, 289, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Lamattina, L.; Gonzalez, D.; Gualberto, J.; Grienenberger, J.M. Higher plant mitochondria encode an homologue of the nuclear-encoded 30-kDa subunit of bovine mitochondrial complex I. Eur. J. Biochem. 1993, 217, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Rayapuram, N.; Hagenmuller, J.; Grienenberger, J.M.; Bonnard, G.; Giegé, P. The Three Mitochondrial Encoded Ccmf Proteins Form a Complex That Interacts with CCMH and c-Type Apocytochromes in Arabidopsis. J. Biol. Chem. 2008, 283, 25200–25208. [Google Scholar] [CrossRef] [PubMed]
- Rayapuram, N.; Hagenmuller, J.; Grienenberger, J.-M.; Giegé, P.; Bonnard, G. AtCCMA Interacts with AtCcmB to Form a Novel Mitochondrial ABC Transporter Involved in Cytochrome c Maturation in Arabidopsis. J. Biol. Chem. 2007, 282, 21015–21023. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. a | Mean % Volume b | Assignment; Species; FunCat c | Protein Record Version | UniProt Accession No. | AGI Identifier | Nominal | Observed | Mascot Score; emPAI d | Coverage (%) | Uniq. Peps; Tot. Peps e | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K | C; H | CA; HA | M | pI | M | pI | ||||||||
| 1 | 0.53 ± 0.02 | 1.06 ± 0.08 (+2.00) | 0.68 ± 0.19 [+1.28] | Mitochondrial heat shock protein 70-1; Arabidopsis; PrF | CAB37531.1 | Q9SZJ3 | At4g37910 | 71.4 | 5.31 | 79 | 5.35 | 4732; 3.61 | 33 | 53; 129 |
| 2 | 0.11 ± 0.01 | 0.21 ± 0.06 [+1.91] | 0.22 ± 0.03 (+2.00) | Pyruvate dehydrogenase E1 beta subunit; Arabidopsis; CM | NP_199898.1 | Q38799 | At5g50850 | 39.4 | 5.67 | 39 | 5.16 | 5484; 2.94 | 29 | 73; 146 |
| 3 | 0.18 ± 0.01 | 0.40 ± 0.09 (+2.22) | 0.21 ± 0.01 (+1.17) | 3-phosphoglycerate dehydrogenase-like protein; Arabidopsis; AM | NP_195146.1 | O49485 | At4g34200 | 63.6 | 6.16 | 72 | 5.43 | 7707; 1.48 | 31 | 112; 266 |
| 4 | 0.11 ± 0.01 | 0.15 ± 0.01 (+1.36) | 0.21 ± 0.03 (+1.91) | Phosphoglycerate kinase 1; Arabidopsis; CM | NP_187884.1 | Q9LD57 | At3g12780 | 50.1 | 5.91 | 43 | 5.20 | 630; 1.44 | 28 | 9; 26 |
| 5 | 0.15 ± 0.01 | 0.19 ± 0.04 (+1.27) | 0.29 ± 0.03 (+1.93) | Malate dehydrogenase (NAD), mitochondrial; Arabidopsis: CM | NP_564625.1 | Q9ZP06 | At1g53240 | 36.0 | 8.54 | 37 | 5.58 | 409; 0.98 | 22 | 10; 18 |
| 6 | 0.19 ± 0.02 | 0.37 ± 0.05 (+1.95) | 0.16 ± 0.03 (−1.18) | Heat shock protein 81-2 (HSP 90 related); Arabidopsis; PrF | NP_200414.1 | P55737 | At5g56030 | 80.2 | 4.95 | 84 | 5.20 | 277; 0.43 | 14 | 0; 12 |
| 7 | 0.16 ± 0.05 | 0.39 ± 0.09 (+2.44) | 0.13 ± 0.07 (−1.23) | Heat shock protein 90; Arabidopsis; PrF | BAF00175.1 | Q0WRS4 | At3g07770 | 90.8 | 5.26 | 89 | 5.06 | 1996; 1.11 | 19 | 15; 75 |
| 8 | 0.17 ± 0.02 | 0.64 ± 0.19 (+3.76) | 0.48 ± 0.07 (+2.82) | 3-phosphoglycerate dehydrogenase-like protein; Arabidopsis; AM | NP_195146.1 | O49485 | At4g34200 | 63.6 | 6.16 | 73 | 5.56 | 6778; 1.74 | 30 | 88; 237 |
| 9 | 0.25 ± 0.03 | 0.72 ± 0.31 (+2.88) | 0.72 ± 0.16 (+2.88) | 3-phosphoglycerate dehydrogenase-like protein; Arabidopsis; AM | NP_195146.1 | O49485 | At4g34200 | 63.6 | 6.16 | 74 | 5.49 | 7997; 1.36 | 27 | 111; 301 |
| 10 | 0.05 ± 0.03 | 0.06 ± 0.01 (+1.20) | 0.19 ± 0.04 (+3.80) | Phosphoglycerate kinase 1; Arabidopsis; CM | NP_187884.1 | Q9LD57 | At3g12780 | 50.2 | 5.91 | 81 | 5.50 | 1376; 2.57 | 42 | 22; 53 |
| 11 | 0.14 ± 0.08 | 0.12 ± 0.11 (−1.17) | 1.29 ± 0.25 (+9.21) | Putative succinyl-CoA ligase (GDP-forming) beta subunit, mitochondrial; Arabidopsis; CM | NP_179632.1 | O82662 | At2g20420 | 45.6 | 6.30 | 41 | 5.15 | 265; 0.88 | 17 | 11; 11 |
| 12 | 0.56 ± 0.15 | 0.32 ± 0.17 (−1.75) | 0.11 ± 0.02 (−5.09) | Putative mitochondrial processing peptidase; Arabidopsis; PrF | BAE98412.1 | Q42290 | At3g02090 | 51.5 | 5.71 | 80 | 5.79 | 833; 1.38 | 26 | 17; 42 |
| 13 | 0.36 ± 0.14 | 0.47 ± 0.09 (+1.30) | 0.19 ± 0.04 (−1.89) | ATPase subunit 1; Brassica napus; RC | YP_717155.1 | Q6YSN4 | AtMg01190 | 55.4 | 6.01 | 58 | 6.07 | 3195; 1.12 | 23 | 76; 168 |
| 14 | 0.35 ± 0.14 | 0.22 ± 0.04 (−1.59) | 0.08 ± 0.03 (−4.37) | Δ-1-pyrroline-5-carboxylate dehydrogenase precursor; Arabidopsis; AM | AAK73756.1 | Q8VZC3 | At5g62530 | 62.2 | 6.26 | 70 | 6.24 | 784; 0.67 | 18 | 3; 41 |
| 15 | 0.54 ± 0.06 | 0.33 ± 0.06 (−1.64) | 0.09 ± 0.01 (−6.00) | Δ-1-pyrroline-5-carboxylate dehydrogenase precursor; Arabidopsis; AM | AAK73756.1 | Q8VZC3 | At5g62530 | 62.2 | 6.26 | 69 | 6.33 | 1622; 0.95 | 21 | 0; 78 |
| 16 | 0.60 ± 0.14 | 0.77 ± 0.11 (+1.28) | 0.31 ± 0.05 (−1.93) | Mitochondrial elongation factor Tu; Arabidopsis; PrS | CAA61511.1 | Q9ZT91 | At4g02930 | 51.6 | 5.53 | 42 | 6.00 | 3123; 2.45 | 37 | 24; 100 |
| 17 | 0.31 ± 0.08 | 0.42 ± 0.07 (+1.35) | 0.18 ± 0.04 (−1.72) | Mitochondrial elongation factor Tu; Arabidopsis; PrS | CAA61511.1 | Q9ZT91 | At4g02930 | 51.6 | 5.53 | 42 | 6.19 | 2502; 2.05 | 33 | 35; 77 |
| 18 | 0.58 ± 0.15 | 0.52 ± 0.07 (−1.11) | 0.14 ± 0.02 (−4.14) | Isocitrate dehydrogenase-like protein; Arabidopsis; CM | CAB87626.1 | Q9LYK1 | At5g14590 | 52.3 | 7.11 | 45 | 6.37 | 697; 0.96 | 23 | 14; 23 |
| 19 | 0.26 ± 0.06 | 0.17 ± 0.03 (−1.53) | 0.11 ± 0.01 (−2.36) | Citrate synthase (SI); Arabidopsis; CM | NP_850415.1 | P20115 | At2g44350 | 53.1 | 6.41 | 48 | 6.57 | 999; 0.62 | 18 | 34; 40 |
| 20 | 0.40 ± 0.09 | 0.23 ± 0.07 (−1.74) | 0.15 ± 0.01 (−2.67) | Citrate synthase (SI); Arabidopsis: CM | NP_850415.1 | P20115 | At2g44350 | 53.1 | 6.41 | 47 | 6.83 | 2126; 0.83 | 21 | 51; 84 |
| 21 | 0.08 ± 0.06 | 0.16 ± 0.08 (+2.00) | 0.61 ± 0.08 (+7.62) | 10 kDa chaperonin; Arabidopsis; PrF | NP_563961.1 | P34893 | At1g14980 | 10.8 | 6.74 | 19 | 9.38 | 107; 1.30 | 31 | 8; 8 |
| 22 | LAf | 0.80 ± 0.20 | 0.53 ± 0.01 | ATP synthase, d chain, mitochondrial; Arabidopsis; RC | NP_190798.1 | Q9FT52 | At3g52300 | 19.6 | 5.09 | 20 | 4.59 | 979; 1.60 | 25 | 18; 39 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rurek, M.; Czołpińska, M.; Pawłowski, T.A.; Krzesiński, W.; Spiżewski, T. Cold and Heat Stress Diversely Alter Both Cauliflower Respiration and Distinct Mitochondrial Proteins Including OXPHOS Components and Matrix Enzymes. Int. J. Mol. Sci. 2018, 19, 877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030877
Rurek M, Czołpińska M, Pawłowski TA, Krzesiński W, Spiżewski T. Cold and Heat Stress Diversely Alter Both Cauliflower Respiration and Distinct Mitochondrial Proteins Including OXPHOS Components and Matrix Enzymes. International Journal of Molecular Sciences. 2018; 19(3):877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030877
Chicago/Turabian StyleRurek, Michał, Magdalena Czołpińska, Tomasz Andrzej Pawłowski, Włodzimierz Krzesiński, and Tomasz Spiżewski. 2018. "Cold and Heat Stress Diversely Alter Both Cauliflower Respiration and Distinct Mitochondrial Proteins Including OXPHOS Components and Matrix Enzymes" International Journal of Molecular Sciences 19, no. 3: 877. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19030877