Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.)

College of Horticulture, Northwest A&F University, Yangling 712100, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(4), 1038; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041038
Submission received: 28 January 2018
/
Revised: 26 March 2018
/
Accepted: 27 March 2018
/
Published: 30 March 2018
(This article belongs to the Special Issue Plant Defense Genes Against Biotic Stresses)
Abstract
:RNA silencing is an evolutionarily conserved mechanism that regulates variety of cellular processes in plants. Argonaute protein (AGO), Dicer-like protein (DCL) and RNA-dependent RNA polymerase (RDR) are critical components of RNA silencing. These efficient and indispensable components of the RNAi pathway have not been identified and characterized in pepper. In this study, we identified 12 CaAGO, 4 CaDCL and 6 CaRDR genes in pepper and compared them with those of Arabidopsis, tobacco, potato and tomato. Detailed phylogenetic analyses revealed that each CaAGO, CaDCL and CaRDR protein family were classified into four clades. The tissue specific expression and respond to abiotic or biotic stress were studied. The real-time quantitative polymerase chain reaction (PCR) results demonstrated that CaAGO2, CaAGO10b, CaDCL2 and CaDCL4 were upregulated with cucumber mosaic virus (CMV), potato virus Y (PVY) and tobacco mosaic virus (TMV) infections, whereas they showed difference expression patterns in response to abiotic stress. In addition, we found that many of the candidate genes were induced by phytohormones and H2O2 treatment. Our results provide useful information for further elucidation of gene silencing pathways and RNAi-mediated host immunity in pepper.
1. Introduction
Plants have evolved some control mechanisms that efficiently prevent pathogen invasion to protect themselves from pathogen attack throughout their life-cycles [1]. RNA silencing is one such mechanism, which is highly conserved in most eukaryotes and controls sequence specific regulation of gene expression. Dicer-like (DCL), Argonaute (AGO), and RNA-dependent RNA Polymerase (RDR) proteins are the key components of RNA silencing machinery [1,2,3]. DCL proteins belong to the RNase III family of endoribonucleases that contain DExD, Helicase-C, DUF283, PAZ, RNase III and dsRNA-binding domains [4,5]. DCLs process double-stranded RNAs (dsRNAs) into 21–24 nucleotide small RNA duplexes [4]. Previous studies indicated that AGO proteins contained the PAZ domain and PIWI domain. The PAZ domain can bend small RNA into a specific binding pocket, whereas the PIWI domain can cleave target mRNA similar to RNase H [6,7]. RDR proteins, containing a RNA-dependent RNA polymerase (RdRP) domain, catalyze the dsRNA formation from single-stranded RNAs (ssRNAs) [8].
In recent years, studies of the AGO, DCL and RDR gene families in Arabidopsis, rice, tomato and maize have advanced our understanding of RNA silencing [9,10,11]. There are 10 AtAGOs, four AtDCLs and six AtRDRs in Arabidopsis thaliana [11]. In rice, eight OsDCLs, 19 OsAGOs and five OsRDRs genes were identified, in which OsAGO2 showed specific upregulation in response to cold, salt and dehydration stress [10]. Likewise, genes for seven SlDCLs, 15 SlAGOs and six SlRDRs were identified in tomato. The expression models of tandem gene duplications among SlDCL2s indicate that the DCL2 family plays an important role in the evolution of tomato [9]. Similarly, a total of seven, five and eight CsAGOs, CsDCLs, and CsRDR genes, respectively, have been identified in cucumber. All CsAGOs, especially CsAGO1c, CsAGO1d, and CsAGO7, were profusely upregulated in leaves and tendrils compared to that in other organs, whereas all CsDCL genes showed a higher up regulation in tendrils, with almost no expression of CsDCL1, CsDCL4a, or CsDCL4b in other organs. In addition, CsRDR1a, CsRDR2, CsRDR3, and CsRDR6 were relatively upregulated in tendrils, but almost all CsRDRs are downregulated in other organs [12]. Genome of the allopolyploid species of Brassica napus possessed eight BnDCLs, 27 BnAGOs, and 16 BnRDRs [13,14]. In grapevines, a total of four VvDCLs, 13 VvAGOs, and five VvRDRs were identified. It was worth mentioning that one gene, VvAGO10a, was only expressed in the stem, suggesting that VvAGO10a might function in the regulation of siRNAs in the grapevine stem [15].Thus, these key components of RNA silencing machinery of various plant species exhibited considerable variation and likely contributed to a diverse set of functions in different species of plants.
Pepper is one of the most important vegetable crops in the world. However, its productivity is severely affected by viral disease [16,17]. In previous study, we cloned CaRDR1 from pepper, which was induced by salicylic acid (SA) and tobacco mosaic virus (TMV). CaRDR1 played a positive role in pepper TMV resistance by regulating antioxidant enzymes’ activities and the expression of RNA silencing-related genes [18]. In this study, the expression pattern of pepper AGO, DCL and RDR gene families were examined in response to biotic/abiotic stress. These results provide useful information for further elucidation of RNA silencing pathways and RNAi-mediated host immunity in pepper.
2. Results
In this study, expression levels of RNA silencing related genes were investigated in response to biotic and abiotic stress conditions. In addition, effects of these treatments were evaluated by detecting the expression of stress-related genes [19,20,21]. CaPR1 was induced by cucumber mosaic virus (CMV), potato virus Y (PVY) and TMV infections. The expression level of CaDEF1 upregulated after abscisic acid (ABA), H2O2, MeJA, SA, NaCl and PEG treatments, and CaEREBP-C1 was induced by cold treatment (Figure S1). The results indicated that the stresses worked on the plants.
2.1. Identification and Structural Analysis of CaAGO, CaDCL and CaRDR Genes
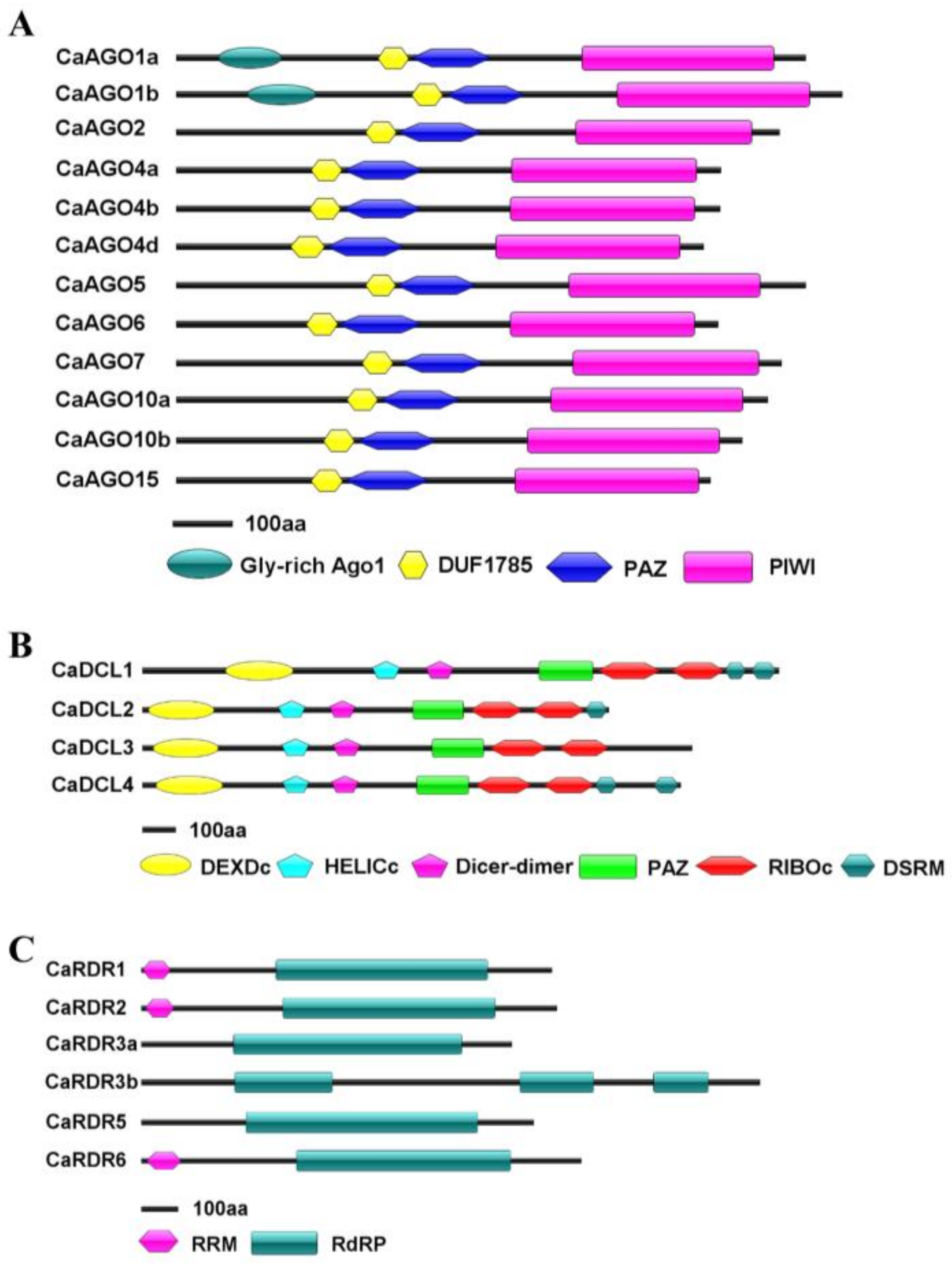
To identify potential CaAGO, CaDCL and CaRDR genes in the pepper genome, we obtained the Hidden Markov Model (HMM) profiles of the conserved PIWI, DCL (RNase III) and RdRP, and then used BLAST-p to search a draft pepper genome sequence on the genome database (http://peppersequence.genomics.cn/page/species/index.jsp and Table 1). Subsequently, the structural integrity of conserved domains was evaluated, and redundant sequences were eliminated. Twelve CaAGOs, four CaDCLs and six CaRDRs were identified in pepper. The identified AGOs showed coding potentials of −100 kDa proteins. Early studies showed that AGO proteins typically have a PAZ domain and a PIWI domain [6,7]. CaAGOs shared a DUF1785 domain, a PAZ domain and a C-terminus PIWI domain, which were highly consistent with known plant AGO proteins by SMART analysis (Figure 1A). In addition, a Gly-rich AGO1 domain was found in front of the DUF1785 domain in CaAGO1a/b proteins. The pepper genome encoded four hypothetical CaDCLs, which contained the conserved DEXDc, HELICc, Dicer-dimer, PAZ, RIBOc and DSRM domains of DCL proteins in plants (Figure 1B). In addition, CaDCL3 lacked C-terminal DSRM regions (Figure 1B). The four DCLs showed coding potentials of 158–214 kDa proteins. Six hypothetical CaRDRs in pepper shared a common motif corresponding to the catalytic β′ subunit of RdRP [22]. They showed coding potentials of 114–135 kDa proteins. In contrast, homologous CaRDR3b was the largest protein in the RDR family, most likely to encode 1682-amino acid polypeptides. Besides the conserved RdRP domain, there was an RRM (RNA recognition motif) domain that existed in the N-terminus of CaRDR1, CaRDR2 and CaRDR6 (Figure 1C). These analyses demonstrated that DCL, RDR, and AGO proteins, along with their correct domains, are well conserved in pepper.
2.2. Phylogenetic Analysis of CaAGO, CaDCL and CaRDR Genes
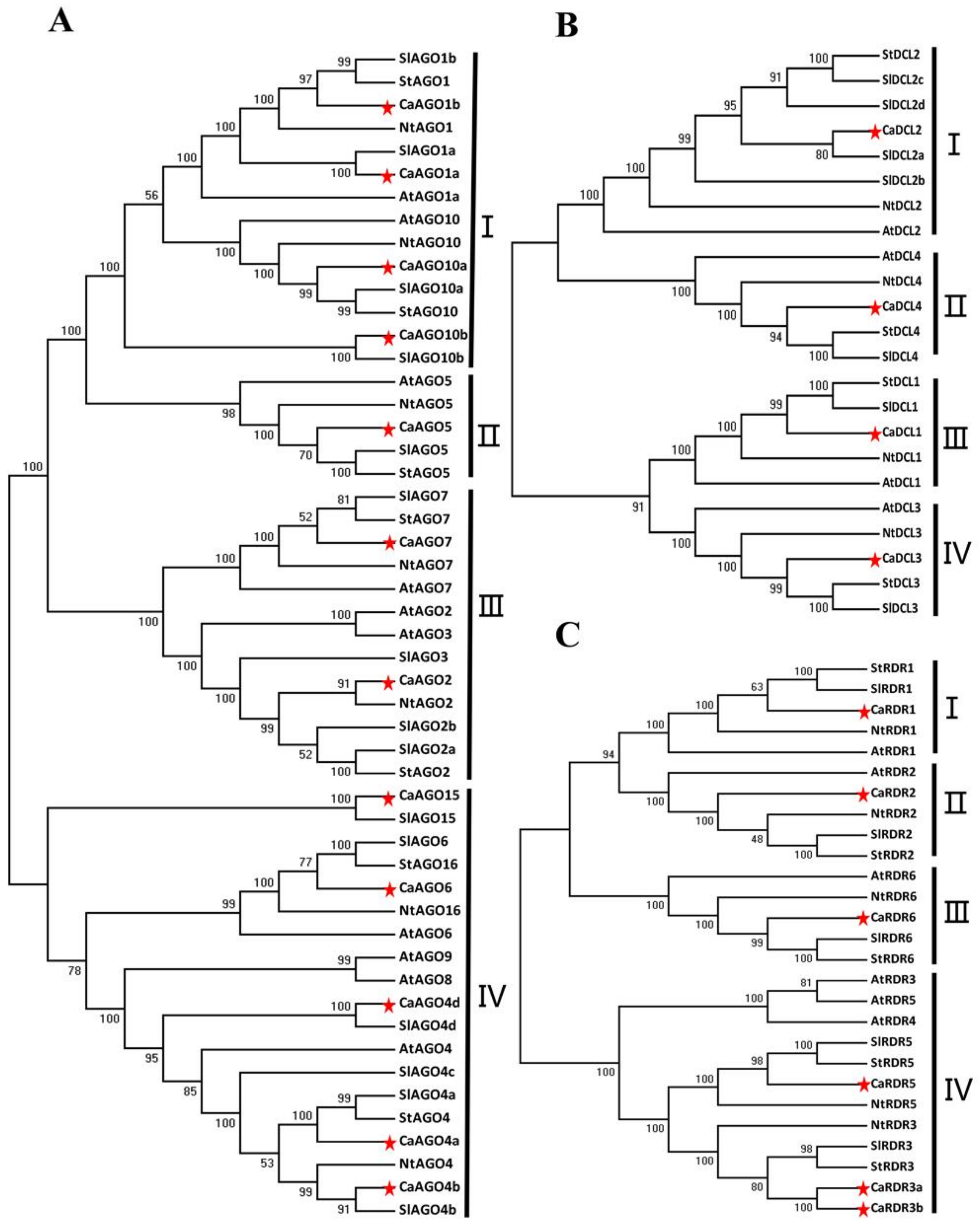
The AGO, DCL and RDR proteins of Arabidopsis, tomato and tobacco were used to study the phylogenetic relationships and functional diversities of CaAGOs, CaRDRs and CaDCLs in pepper. The 12 CaAGOs were separated into four distinct groups in the phylogenetic tree (Figure 2A). The groups were named according to their identity to tomato AGO proteins. Among all clades, group I was with four CaAGOs proteins (CaAGO1a, CaAGO1b, CaAGO10a, CaAGO10b). Group II and group III contained CaAGO5 and two CaAGOs (CaAGO2 and CaAGO7), respectively. There were five CaAGOs in group VI, which were CaAGO15, CaAGO6, CaAGO4a, CaAGO4b and CaAGO4d. The CaDCLs showed high sequence conservation compared with tomato. The four CaDCLs could be classified into four distinct clades (Figure 2B). Each clade contained one member that was closely allied with SlDCL orthologs at a high similarity. These results indicated that high conservation of DCL family in dicots. The phylogenetic tree derived from CaRDRs sequences was divided into four clades (Figure 2C). Among the four groups, group I contained one member, CaRDR1, as shown in Figure 2C. Groups II and III also contained one member, CaRDR2 and CaRDR6, respectively. There were three CaRDRs in group VI, which were CaRDR3a, CaRDR3b and CaRDR5.
2.3. Expression Pattern of CaAGOs, CaDCLs and CaRDRs in Various Organs
In order to determine the expression pattern of candidate genes in different organs of pepper, real-time quantitative polymerase chain reaction (qRT-PCR) was performed to analyze the transcript level of CaAGOs, CaDCLs and CaRDRs. The various organs of pepper: roots, stems, leaves, flowers, and fruits were investigated. The results showed that most of the CaAGO genes were expressed in all five organs except for CaAGO4d and CaAGO15. This was probably due to no expression or an extremely low expression of these genes in these organs. The spatial expression data that normalized with CaUbi3 were compared with data for roots. Compared to their expression in root, 10 of the CaAGO genes exhibited higher transcript level (fold > 2) in flower, especially CaAGO1b, CaAGO5 and CaAGO10b (Figure 3, Figure S2). CaAGO2 showed high expression in fruit; CaAGO4a showed high expression in leaves; CaAGO6, CaAGO10a and CaAGO10b showed high expression in stems compared to their expression in roots. Similarly, the expression results showed that all CaDCLs and CaRDRs were also expressed in various organs (Figure 3). All of the CaDCLs exhibited a higher expression level in flowers as compared to roots (Figure 3). CaRDR1 and CaRDR5 were highly expressed in stems, while the other CaRDRs exhibited a higher level of expression in flowers as compared to their expression in root tissues (Figure 3).
2.4. Biotic Stress Induces Expression of CaAGO, CaDCL and CaRDR Genes
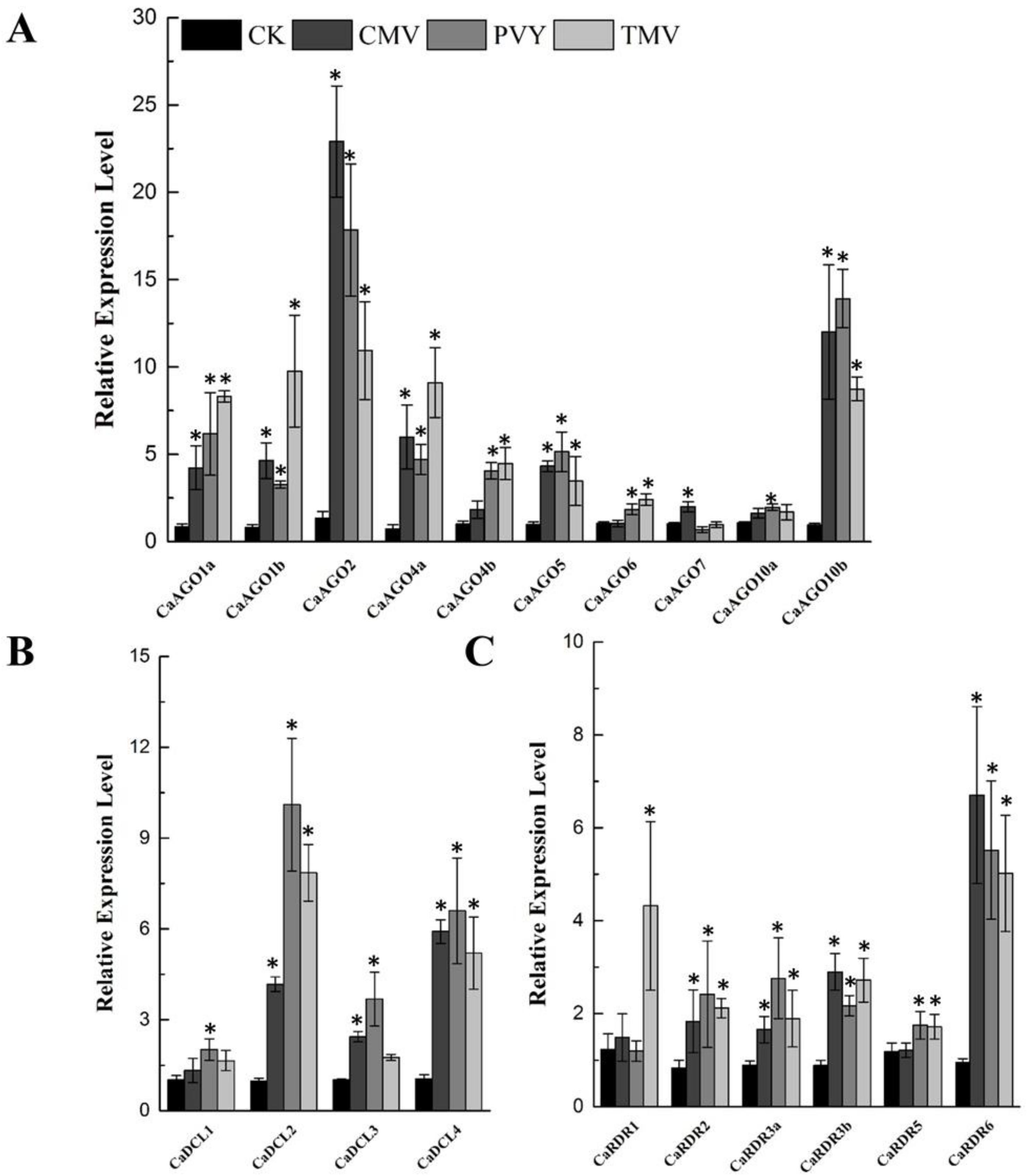
To unravel the functions of CaAGOs, CaDCLs and CaRDRs in response to biotic stressors, we inoculated pepper leaves with TMV, CMV and PVY, and measured the expression of CaAGOs, CaDCLs and CaRDRs. At 7 day-post inoculation (dpi) with the viruses, the expression of CaAGO genes was differentially expressed in pepper leaves (Figure 4A). The transcripts of CaAGO2 and CaAGO10b were significantly induced by CMV inoculation, accounting for 20-fold and 10-fold increases, respectively (Figure 4A), the expression of these genes were also upregulated by PVY inoculation (>10 folds) (Figure 4A). Similarly, an upregulation in the expression of CaAGO1a/1b, CaAGO2, CaAGO4a and CaAGO10b was observed at 7 dpi with TMV (Figure 4A). In this study, the transcripts of CaDCL2 and CaDCL4 responded to all viruses (Figure 4B); however, a relatively higher expression of CaDCL3 was observed when challenged with PVY. The transcripts of CaRDR6 were significantly induced upon virus inoculation, and even more CaRDR1 was induced by TMV (Figure 4C). The results suggest that these genes commonly participated in virus-induced resistance pathways.
2.5. Abiotic Stress Induces CaAGO, CaDCL and CaRDR Expression
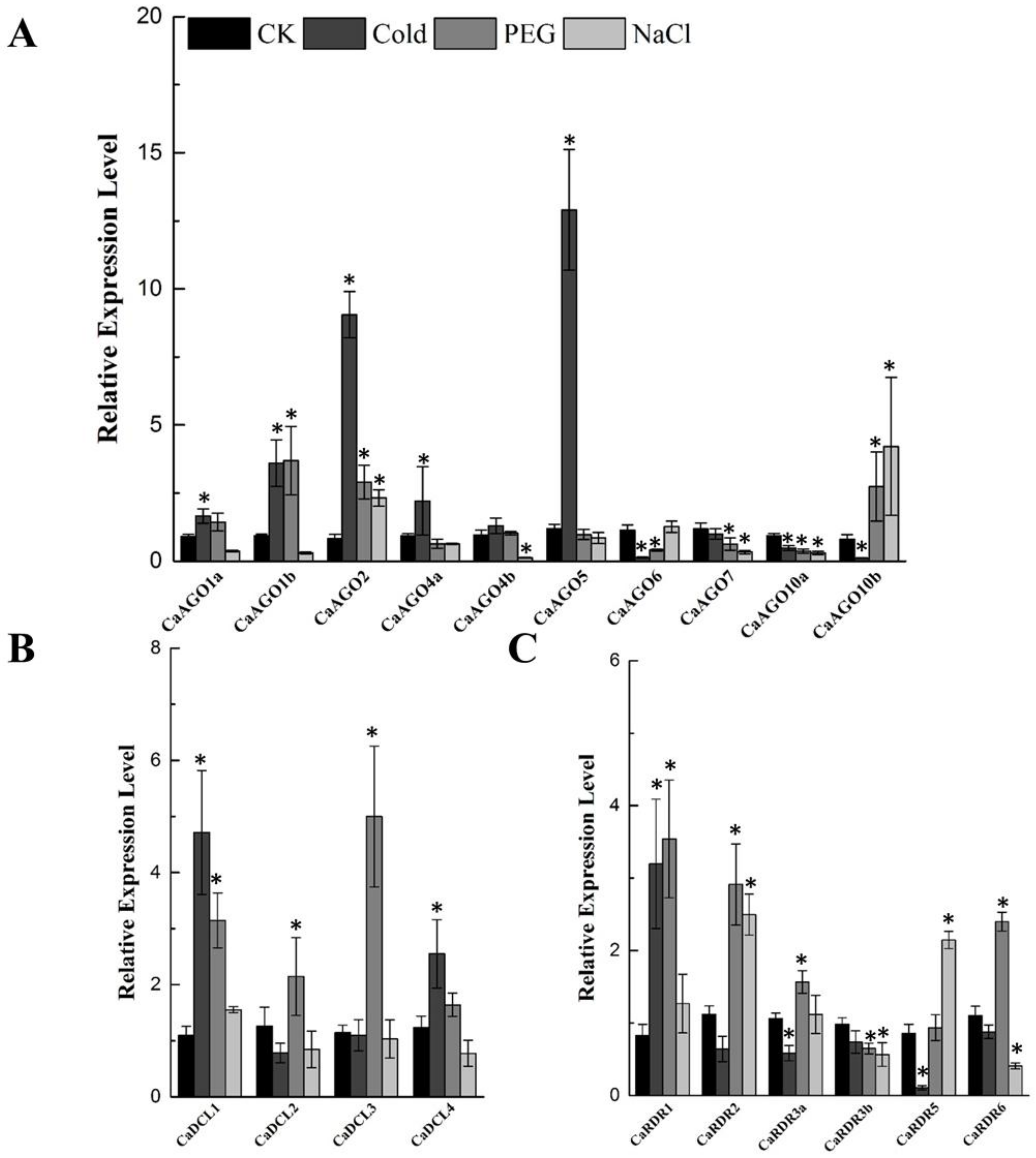
Evidence from prior research shows that RNA silencing plays a critical role in plant tolerance to abiotic stress [9]. Therefore, the expression patterns of CaAGOs, CaDCLs and CaRDRs were measured at 24 h-posttreatment with cold, drought and salinity in pepper. Although cold treatment positively induced CaAGO1b, CaAGO2 and CaAGO5 expression, it suppressed the expression of CaAGO6, CaAGO10a and CaAGO10b in pepper (Figure 5A). Drought and salinity increased the transcripts of CaAGO2 and CaAGO10b mildly. Interestingly, CaAGO10a was downregulated under all abiotic stresses. CaDCL1 and CaDCL4 were upregulated by cold treatment (Figure 5B). Drought stress induced transcripts of CaDCL1 and CaDCL3, especially CaDCL3, by 5-fold (Figure 5B). Among CaRDRs, CaRDR1 expressed in response to cold and drought treatment (Figure 5C). Drought treatment also increased the transcripts of CaRDR2 and CaRDR6, whereas CaRDR2 and CaRDR5 were upregulated by salinity. The expression of CaRDR3b was not induced by abiotic treatment (Figure 5C).
2.6. CaAGOs, CaDCLs and CaRDRs Are Responsive to Phytohormones and H2O2
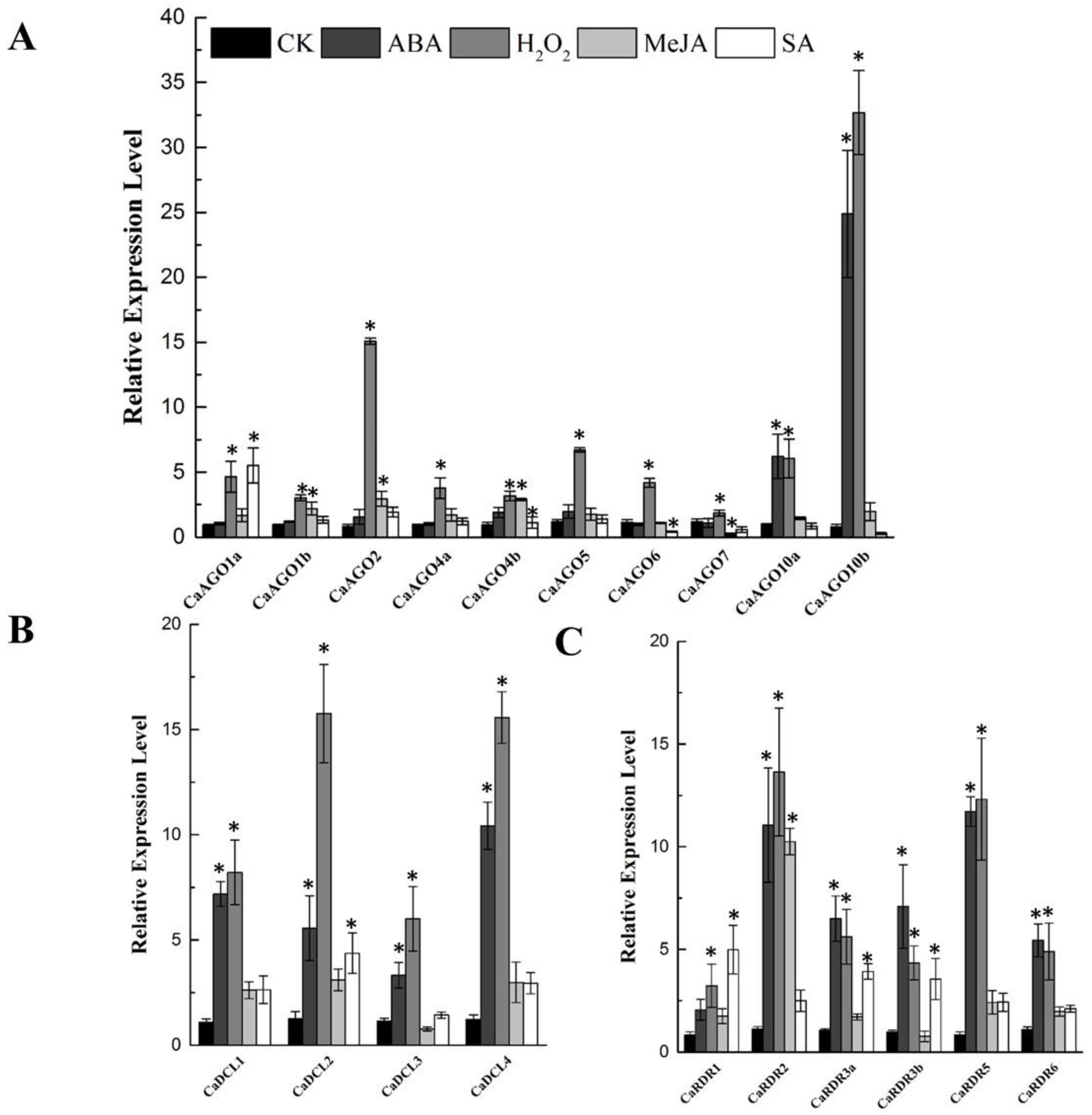
Phytohormones and H2O2 function as signals in mediating plant response to abiotic and biotic stress. The expression of CaAGOs, CaDCLs and CaRDRs was assessed after ABA, H2O2, methyl jasmonate (MeJA) and SA treatment. Gene expression analysis at 24 h after phytohormones and H2O2 treatment showed that CaAGO10a and CaAGO10b were significantly induced by ABA, especially CaAGO10b (20 folds). Similarly, H2O2 increased the transcripts of CaAGO1a, CaAGO2, CaAGO5, CaAGO6, CaAGO10a and CaAGO10b (Figure 6A), in which the expression of AGO10b was upregulated by 32-fold with H2O2 treatment (Figure 6A). The expression of CaAGOs were hardly affected by MeJA (Figure 6A). CaAGO1a expression was upregulated by SA (Figure 6A). The transcripts of CaDCLs were significantly induced by ABA and H2O2. Similarly, MeJA increased the expression of CaDCLs, expect for CaDCL3, whereas SA induced the expression of CaDCL2 by 4-fold (Figure 6B). The transcripts of CaRDR2 and CaRDR5 were significantly induced by both ABA and H2O2. Likewise, MeJA significantly induced the transcript of CaRDR2 (Figure 6C) and SA induced the expression of CaRDR1 and CaRDR3a/b in pepper (Figure 6C).
3. Discussion
In plants, RNA silencing plays an important role in sequence specific regulation of gene expression via posttranscriptional regulation and chromatin modification during abiotic stress, viral defense and plant development. Therefore, it is indispensable to explore the temporal and spatial expression patterns of the core elements of the RNA silencing machinery. In addition, availability of the pepper genome sequence has enabled genome wide gene expression analysis in pepper [23]. In the present investigation, 12 CaAGOs, four CaDCLs and six CaRDRs genes were identified in the pepper genome and a phylogenetic analysis for each gene family was carried out. Finally, the expression patterns of CaAGOs, CaDCLs and CaRDRs under biotic or abiotic stress and treatment with phytochromes were analyzed in pepper. Our results unveiled important roles of CaAGOs, CaDCLs and CaRDRs that provide new insights into gene silencing pathways and RNAi-mediated host immunity in pepper.
3.1. Argonaute (AGO) Proteins in Pepper
Argonautes are the highly basic RNA binding proteins characterized by the presence of PAZ and PIWI domains [24]. Genes for 12 CaAGOs were identified in the pepper genome in this study (Table 1). CaAGO4d and CaAGO15 were barely detected by real-time qPCR. Other CaAGO genes exhibited diverse expression patterns in different organs (Figure 3). CaAGO1a, CaAGO1b, CaAGO10a and CaAGO10b were grouped into the same cluster (Figure 2). In Arabidopsis, AtAGO1 was expressed generally in leaves, roots, flowers and siliques, but mutant ago1 showed a dwarf and sterile phenotype [10,25,26]. Notably, AGO10 acted as a locker of miR165/miR166 in shoot apical meristem (SAM) development, while miR165/miR166 cooperated with AGO1 to suppress SAM maintenance [27,28]. In the present study, we found an increased expression of CaAGO1a/CaAGO1b in flowers compared to that in other organs, whereas CaAGO10a/CaAGO10b showed high expression in stems. These results were in agreement with a previous report, in which BnAGO1a was profusely expressed in flowers of Brassica napus [14]. AGO1 stabilized miR168 posttranscriptional and the transcripts of AGO1 were regulated by miR168, which played an important role in plant development [29]. AGO1 and AGO10 interacted with miR172 and miR165/166 regulate the SAM and floral meristems development through targeting APETALA2 (AP2) and type III homeodomain-leucine zipper (HD-Zip) genes, respectively [28,30,31,32]. Cells in the SAM whether differentiated or not were regulated by HD-Zip transcription factors [33]. AP2 played a key role in the specification of reproductive and perianth organ identities in flower development [34]. The AS1 and AS2 genes played important roles in leaf development [35,36]. AGO1 interacted with AS1 and AS2 for plant development, and AGO1 was required for repressing class I KNOX genes in the developing leaves [37]. AGO1a and AGO2 were co-expressed with MADS15 involved in the flowering process and flower development in rice [38]. An miR168 binding site was found in CaAGO1a/b (Figure S3A,B). It suggested that AGO1/10 might participate in the plant development, especially flower development via miR168 with regulating the development related genes’ expression.
AGO2-like proteins play a crucial role in battle against viral infections guided by siRNAs generated from double stranded virus RNAs that are synthesized by RdRP using viral RNA as templates [39]. Likewise, AGO2 functioned in defense against various viruses, including TCV, Potato virus X, CMV, and Tomato bushy stunt virus [39,40,41,42]. In Arabidopsis, PVX failed to infect wild-type but not ago2 mutants, suggesting that AGO2 is required to suppress PVX infection [43]. While AtAGO2 was highly induced by Pseudomonas syringae pv. tomato (Pst) in wild-type, ago2 mutants displayed an enhanced susceptibility to Pst [44]. Similarly, in Nicotiana benthamiana, NbAGO2 contributed to anti-viral defense, and suppression of the NbAGO2 expression enhanced susceptibility to TBSV [45]. Both ago1 and ago2 mutants were hypersensitive to viral infection in plant [46,47,48,49]. In plants, AGO2 mRNA is targeted by the miR403 [50,51]. Transcript levels of AGO1 but not AGO2 were repressed after SMV infection, which were regulated by the upregulation of miR168a and miR403a [52]. RNA-mediated defense contains multiple layers in the interactions between plants and viruses. AGO1 represented a first layer and AGO2 acted as the second layer when AGO1 was overcome by viral suppressors of silencing, and the second layer was also activated when the first layer was suppressed because AGO2 was repressed by AGO1 via miR403 [39]. CaAGO2 contained an miR403 binding site (Figure S3C). Our real-time PCR data showed that CaAGO2 was significantly induced by biotic stress (Figure 4A). CaAGO2 might be participated in pepper virus defense via miR403. AGO1 mainly acted in miRNA and siRNA pathways for post-transcriptional gene silencing (PTGS) [53]. AGO1 functions to ensure targeting and efficient clearance of viral RNAs [54]. The expression of CaAGO1a/b and CaAGO10b increased by CMV, PVY and TMV infection. It implied that CaAGO1 might have played a positive role in pepper through other regulatory mechanisms. In line with previous reports, our findings suggested that the AGO family genes might function collaboratively in RNA silencing-mediated viral defense in plants. Importantly, CaAGOs were also induced by aboitic stress (Figure 5). Here, cold treatment increased the transcripts of CaAGO2 and CaAGO5, whereas drought and salinity induced an upregulation in CaAGO10b expression. In sunflowers, miR403 played critical roles in responses to stress [55]. CaAGO2 might be regulated the response of abiotic stresses via miR403. In rice, OsAGO2 also shows similar upregulation in response to cold, salt and dehydration stress [10]. Furthermore, CaAGO10a/10b expressions were induced by ABA (Figure 6). ABA is a universal hormone in plants, and it was a core component in multiple plant signaling pathways to mediate several responses, including gene regulation, stomatal closure and plant growth modulation [56,57,58]. Our results suggested that CaAGO10b might play an important role in the response of pepper plants to osmotic stress by regulating ABA responsive genes. However, CaAGO10b shows a strong induction in the presence of ABA rather than NaCl. It implied that ABA might participate in multiple plant physiological mechanism via CaAGO10b, and this hypothesis needs to be confirmed in further study.
3.2. Dicer-Like (DCL) Proteins in Pepper
Dicer enzymes work to process dsRNA into small RNA of diverse size. DCL1 and DCL4 are well known to trigger post-transcriptional gene silencing (PTGS), DCL2 generated 22-nt siRNAs, which share functional overlap in antiviral defense with DCL4-generated 21-nt siRNAs, and DCL3-produced 24-nt heterochromatic siRNA (hc-siRNA) mediated DNA methylation, gene silencing and chromatin modification [59,60,61,62]. In this study, four DCL genes clustered into four subgroups were found in pepper (Figure 2). CaDCL1 and CaDCL3 exhibited a higher expression level in flowers than other organs. In Arabidopsis, DCL1 and DCL3 promote flowering, whereas double mutants of dcl1 and dcl3 exhibited a delay in flowering [63]. It implied that CaDCL1 and CaDCL3 might also be involved in pepper flower development. Moreover, the transcripts of CaDCL2 and CaDCL4 were significantly induced upon virus inoculation (Figure 4B). In tomato, TYLCV infection upregulates expression of SlDCL1, 2a, 2c, 2d and 3, which supports our current results [9]. The expression of CaDCLs was induced under different abotic stress and hormonal treatment (Figure 5 and Figure 6). These findings were in agreement with a previous study on tomato, which showed an increased expression of SlDCL1, 2a, 2bc and 2d in response to various stress [9].
3.3. RNA-Dependent RNA Polymerase (RDR) Proteins in Pepper
RDRs participate in dsRNAs synthesis to initiate a new round of RNA silencing [8,64,65]. Six RDRs had been identified in Arabidopsis [8]. Similarly, we found six CaRDR genes in pepper, which was expressed in all five tissues. Several lines of evidence suggested that plant RDR1 was involved in antiviral defense [66,67]. In Arabidopsis, AtRDR6 acted in many RNA silencing pathways, leading to defense against viruses, such as TMV and CMV [65,68,69,70]. NgRDR6 could be induced by ABA, GA, MeJA, CMV, but not by PVY, TMV, H2O2 and SA in Nicotiana glutinosa [71]. In the present study, CaRDR6 was induced by CMV, PVY, TMV, ABA and H2O2, indicating an important role of RDR6 in different signal pathways in a range of plant species (Figure 4C). AtRDR2 played a crucial role in the biogenesis of hc-siRNAs that induced a DNA methylation pathway with the participation of AGO4 and DCL3 [72,73]. In tomato, TYLCV infection enhanced the expression of SlRDR2 and SlDCL3 [9]. In Solanum chilense, the tomato yellow leaf curl virus resistance loci Ty-1 and Ty-3 were homologous to Arabidopsis RDR3, 4, and 5 [74], suggesting that RDR3, 4, and 5 might be involved in plant viral defense by their active participations in generating siRNAs. In the current study, transcripts of CaRDR3a and CaRDR3b were significantly induced upon virus inoculation. Furthermore, expression of CaRDR2 was enhanced by virus infection and plant hormones (Figure 4C and Figure 6C).
4. Materials and Methods
4.1. Identification of Putative Pepper DCL, AGO, and RDR Genes
Protein sequences of tomato DCLs, AGOs, and RDRs were downloaded from NCBI (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/) and SGN (https://solgenomics.net/). Protein sequence was analyzed for domain structure using Pfam (http://pfam.xfam.org/). DCLs, AGOs, and RDRs in pepper were identified by using Hidden Markov Model (HMM) profiles and BLAST-P to search the Pepper Genome database (http://peppersequence.genomics.cn/page/species/index.jsp). All identified genes in this study were named after the homologies sequence in the same gene family of tomato. The conserved domains of the gene sequences were searched using the Simple Modular Architecture Research Tool (SMART, http://smart.embl-heidelberg.de/). The molecular weight (MW) of CaDCL, CaAGO, and CaRDR proteins were predicted using ExPASy ComputepI/Mwtool (http://au.expasy.org/tools/pi_tool.html). The gene information, including accession number, chromosome location, coding sequence (CDS) length and encoded protein length were downloaded from the pepper genome database.
4.2. Phylogenetic Analysis
4.3. Plant and Treatment
Pepper (Capsicum annuum L.) cultivars P79 was used in the present study. Pepper seedlings were grown in a plant growth chamber under a 16 h/8 h light/dark period at 25 °C/20 °C. Tissue samples were collected from roots, leaves, stems, flowers and fruit.
In addition, 2 mM SA, 100 μM MeJA, 100 μM ABA and 10 mM H2O2 were used to spray the leaves of 8-week-old pepper seedlings [77,78]. The corresponding solvent was used to deal with control plants. Leaves were collected at 24 h after treatment. Samples were frozen in liquid nitrogen and stored at −80 °C.
For abiotic stress, such as drought, salinity and cold, 8-week-old seedlings of pepper were treated with 20% (w/v) polyethylene glycol (PEG), 200 mM sodium chloride (NaCl) and placed at 4 °C, respectively. Leaves were collected at 24 h after treatment. Samples were frozen in liquid nitrogen and stored at −80 °C.
Two to three lower leaves of 8-week-old pepper seedlings were inoculated with TMV, PVY and CMV (0.01 M phosphate buffer, pH 7.0) mechanically. Leaves were collected at 7 d after treatment. Samples were frozen in liquid nitrogen and stored at −80 °C.
4.4. Quantitative Real-Time PCR (qRT-PCR)
An Omega plant RNA kit (Omega Bio-tek, Guangzhou, China) was used for RNA extracted. qRT-PCR performed using iQ5 Real-Time PCR Detection System (Bio-Rad Corp., Hercules, CA, USA) with SYBR® Premix Ex Taq (TaKaRa, Beijing, China). Pepper ubiquitin-conjugating protein (CaUbi3) was used as an internal reference gene [54]. Three biological replicates were performed for qRT-PCR assay. Gene relative expression levels were determined using the 2−ΔΔCt method [79]. Primers for qRT-PCR were listed in Table S2.
4.5. Statistical Analysis
SPSS software was used for statistical analysis. The treatments were compared with the control using Tukey’s test at p < 0.05.
5. Conclusions
In this study, a total of 12 CaAGO, four CaDCL and six CaRDR genes were identified in pepper plants. We discussed the structures and conserved domains of these genes and performed a detailed phylogenetic analysis that classified CaAGO, CaDCL and CaRDR gene families into four clades. Tissue specific expression analysis revealed that CaAGO, CaDCL and CaRDR genes showed multiple patterns of expression in different organs. We also analyzed expression of CaAGOs, CaDCLs and CaRDRs in response to abiotic and biotic stressors. CMV, PVY and TMV infections upregulated CaAGO2, CaAGO10b, CaDCL2, CaDCL4 and CaRDR6 expression, whereas cold, drought and salinity treatments induced various CaAGOs, CaDCLs and CaRDRs expression in pepper. Additionally, a potential involvement of phytohormones in regulating many of the candidate genes was speculated. RNA silencing components showed distinct role in stress responses of pepper. The results play a basis role in further functional characterization of these genes.
Supplementary Materials
Supplementary materials can be found at https://0-www-mdpi-com.brum.beds.ac.uk/1422-0067/19/4/1038/s1.
Acknowledgments
This work was supported by the National Key Technology R&D Program of China (2013BAD01B04-14).
Author Contributions
Yan Liang and Lei Qin conceived and designed the experiments; Lei Qin and Ning Mo performed the experiments; Lei Qin and Tayeb Muhammad analyzed the data; Lei Qin wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AGO | Argonaute Protein |
| ABA | Abscisic Acid |
| CMV | Cucumber Mosaic Virus |
| DCL | Dicer-Like Protein |
| MeJA | Methyl Jasmonate |
| MW | Molecular Weight |
| NaCl | Sodium Chloride |
| PEG | Polyethylene Glycol |
| PVY | Potato Virus Y |
| RDR | RNA-Dependent RNA Polymerase protein |
| RISCs | RNA-induced Silencing Complexes |
| SA | Salicylic Acid |
| ssRNAs | Single-Stranded RNAs |
| TMV | Tobacco Mosaic Virus |
References
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.J.; Carrington, J.C. Specialization and evolution of endogenous small RNA pathways. Nat. Rev. Genet. 2007, 8, 884–896. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Post-transcriptional small RNA pathways in plants: Mechanisms and regulations. Genes Dev. 2006, 20, 759–771. [Google Scholar] [CrossRef] [PubMed]
- Carmell, M.A.; Hannon, G.J. RNase III enzymes and the initiation of gene silencing. Nat. Struct. Mol. Biol. 2004, 11, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Margis, R.; Fusaro, A.F.; Smith, N.A.; Curtin, S.J.; Watson, J.M.; Finnegan, E.J.; Waterhouse, P.M. The evolution and diversification of dicers in plants. FEBS Lett. 2006, 580, 2442–2450. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Doudna, J.A. A three-dimensional view of the molecular machinery of RNA interference. Nature 2009, 457, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Simon, B.; Kirkpatrick, J.P.; Eckhardt, S.; Reuter, M.; Rocha, E.A.; Andradenavarro, M.A.; Sehr, P.; Pillai, R.S.; Carlomagno, T. Recognition of 2′-o-methylated 3′-end of piRNA by the PAZ domain of a PIWI protein. Structure 2011, 19, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M.; Krczal, G. Nomenclature and functions of RNA-directed rna polymerases. Trends Plant Sci. 2006, 11, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Yang, G.S.; Chen, W.T.; Mao, Z.C.; Kang, H.X.; Chen, G.H.; Yang, Y.H.; Xie, B.Y. Genome-wide identification of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analyses in response to viral infection and abiotic stresses in Solanum lycopersicum. Gene 2012, 501, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, M.; Arora, R.; Lama, T.; Nijhawan, A.; Khurana, J.P.; Tyagi, A.K.; Kapoor, S. Genome-wide identification, organization and phylogenetic analysis of Dicer-like, Argonaute and RNA-dependent RNA polymerase gene families and their expression analysis during reproductive development and stress in rice. BMC Genom. 2008, 9, 451. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Plant Argonautes. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Gan, D.; Liang, D.; Wu, J.; Zhan, M.; Yang, F.; Xu, W.; Zhu, S.; Shi, J. Genome-wide identification of the Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in cucumber (Cucumis sativus L.). J. Plant Growth Regul. 2016, 35, 135–150. [Google Scholar] [CrossRef]
- Cao, J.Y.; Xu, Y.P.; Li, W.; Li, S.S.; Rahman, H.; Cai, X.Z. Genome-wide identification of Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in Brassica species and functional analyses of their Arabidopsis homologs in resistance to Sclerotinia sclerotiorum. Front. Plant Sci. 2016, 7, 1614. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zheng, W.; Zhong, Z.; Chen, X.; Wang, A.; Wang, Z. Genome-wide analysis of RNA-interference pathway in Brassica napus, and the expression profile of BnAGOs in response to Sclerotinia sclerotiorum infection. Eur. J. Plant Pathol. 2016, 146, 1–15. [Google Scholar] [CrossRef]
- Zhao, H.; Zhao, K.; Wang, J.; Chen, X.; Chen, Z.; Cai, R.; Xiang, Y. Comprehensive analysis of Dicer-like, Argonaute, and RNA-dependent RNA polymerase gene families in grapevine (vitis vinifera). J. Plant Growth Regul. 2015, 34, 108–121. [Google Scholar] [CrossRef]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (lib.) de bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Erental, A.; Dickman, M.B.; Yarden, O. Sclerotial development in sclerotinia sclerotiorum: Awakening molecular analysis of a “dormant” structure. Fungal Biol. Rev. 2008, 22, 6–16. [Google Scholar] [CrossRef]
- Qin, L.; Mo, N.; Zhang, Y.; Muhammad, T.; Zhao, G.; Zhang, Y.; Liang, Y. CaRDR1, an RNA-dependent RNA polymerase plays a positive role in pepper resistance against TMV. Front. Plant Sci. 2017, 8, 1068. [Google Scholar] [CrossRef] [PubMed]
- Do, H.M.; Lee, S.C.; Jung, H.W.; Sohn, K.H.; Hwang, B.K. Differential expression and in situ localization of a pepper defensin (CADEF1) gene in response to pathogen infection, abiotic elicitors and environmental stresses in Capsicum annuum. Plant Sci. 2004, 166, 1297–1305. [Google Scholar]
- Yu, B.K.; Lee, J.H.; Shin, S.J.; Hwang, E.W.; Kwon, H.B. Molecular characterization of cold stress-related transcription factors, CAEREBP-C1, -C2, -C3, and CaWRKY1a from Capsicum annuum L. J. Plant Biol. 2013, 56, 106–114. [Google Scholar] [CrossRef]
- Lcvan, L.; Eavan, S. The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant Pathol. 1999, 55, 85–97. [Google Scholar]
- Iyer, L.M.; Koonin, E.V.; Aravind, L. Evolutionary connection between the catalytic subunits of DNA-dependent RNA polymerases and eukaryotic RNA-dependent RNA polymerases and the origin of RNA polymerases. BMC Struct. Biol. 2003, 3, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.S.; Roe, S.M.; Barford, D. Structural insights into mRNA recognition from a PIWI domain-siRNA guide complex. Nature 2005, 434, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Fagard, M.; Boutet, S.; Morel, J.B.; Bellini, C.; Vaucheret, H. AGO1, QDE-2, and RDE-1 are related proteins required for post-transcriptional gene silencing in plants, quelling in fungi, and RNA interference in animals. Proc. Natl. Acad. Sci. USA 2000, 97, 11650–11654. [Google Scholar] [CrossRef] [PubMed]
- Bohmert, K.; Camus, I.; Bellini, C.; Bouchez, D.; Caboche, M.; Benning, C. AGO1 defines a novel locus of Arabidopsis controlling leaf development. EMBO J. 1998, 17, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Hu, F.; Wang, R.; Zhou, X.; Sze, S.H.; Liou, L.W.; Barefoot, A.; Dickman, M.; Zhang, X. Arabidopsis Argonaute10 specifically sequesters miR166/165 to regulate shoot apical meristem development. Cell 2011, 145, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Liu, X.; Yan, J.; Wang, W.; Yumul, R.E.; Yun, J.K.; Dinh, T.T.; Liu, J.; Cui, X.; Zheng, B. Argonaute10 and Argonaute1 regulate the termination of floral stem cells through two micrornas in Arabidopsis. PLoS Genet. 2011, 7, e1001358. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Mallory, A.C.; Bartel, D.P. AGO1 homeostasis entails coexpression of miR168 and AGO1 and preferential stabilization of miR168 by AGO1. Mol. Cell 2006, 22, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ji, L.; Le, B.H.; Zhai, J.; Chen, J.; Luscher, E.; Gao, L.; Liu, C.; Cao, X.; Mo, B. Argonaute10 promotes the degradation of miR165/6 through the SDN1 and SDN2 exonucleases in Arabidopsis. PLoS Biol. 2017, 15, e2001272. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.B.; Provost, P.; Ekwall, K.; Hobman, T.C. Ago1 and Dcr1, two core components of the RNA interference pathway, functionally diverge from Rdp1 in regulating cell cycle events in Schizosaccharomyces pombe. Mol. Biol. Cell 2004, 15, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Fernándeznohales, P.; Domenech, M.J.; Martínez de Alba, A.E.; Micol, J.L.; Ponce, M.R.; Madueño, F. AGO1 controls arabidopsis inflorescence architecture possibly by regulating TFL1 expression. Ann. Bot. 2014, 114, 1471–1481. [Google Scholar]
- Barton, M.K. Twenty years on: The inner workings of the shoot apical meristem, a developmental dynamo. Dev. Biol. 2010, 341, 95–113. [Google Scholar] [CrossRef] [PubMed]
- Irish, V.F. The flowering of Arabidopsis flower development. Plant J. Cell Mol. Biol. 2010, 61, 1014–1028. [Google Scholar]
- Byrne, M.E.; Barley, R.; Curtis, M.; Arroyo, J.M.; Dunham, M.; Hudson, A.; Martienssen, R.A. Asymmetric leaves1 mediates leaf patterning and stem cell functionin Arabidopsis. Nature 2000, 408, 967–971. [Google Scholar] [CrossRef] [PubMed]
- Semiarti, E.; Ueno, Y.; Tsukaya, H.; Iwakawa, H.; Machida, C.; Machida, Y. The Asymmetric leaves2 gene of Arabidopsis thaliana regulates formation of a symmetric lamina, establishment of venation and repression of meristem-related homeobox genes in leaves. Development 2001, 128, 1771–1783. [Google Scholar] [PubMed]
- Li, Y.; Huang, W.; Hua, W.; Cai, R.; Xu, Y.; Hai, H. Characterizations of a hypomorphic Argonaute1 mutant reveal novel ago1 functions in arabidopsis lateral organ development. Plant Mol. Biol. 2006, 61, 63–78. [Google Scholar]
- Yang, Y.; Jun, Z.; Yi-Dan, O.; Jialing, Y. The integrative expression and co-expression analysis of the ago gene family in rice. Gene 2013, 528, 221–235. [Google Scholar] [CrossRef] [PubMed]
- Harvey, J.J.; Lewsey, M.G.; Patel, K.; Westwood, J.; Heimstädt, S.; Carr, J.P.; Baulcombe, D.C. An antiviral defense role of AGO2 in plants. PLoS ONE 2011, 6, e14639. [Google Scholar] [CrossRef] [PubMed]
- Jaubert, M.; Bhattacharjee, S.; Mello, A.F.S.; Perry, K.L.; Moffett, P. Argonaute2 mediates RNA-silencing antiviral defenses against Potato virus X in Arabidopsis. Plant Physiol. 2011, 156, 1556–1564. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, H.B.; Alvarado, V.Y.; Vegaarreguin, J.C.; Ciomperlik, J.; Odokonyero, D.; Brosseau, C.; Jaubert, M.; Zamora, A.; Moffett, P. Identification of an Argonaute for antiviral RNA silencing in Nicotiana Benthamiana. Plant Physiol. 2011, 156, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, X.; Singh, J.; Li, D.; Qu, F. Temperature-dependent survival of turnip crinkle virus-infected Arabidopsis plants relies on an RNA silencing-based defense that requires DCL2, AGO2, and HEN1. J. Virol. 2012, 86, 6847–6854. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, C.; Moffett, P. Functional and genetic analysis identify a role for Arabidopsis Argonaute5 in antiviral RNA silencing. Plant Cell 2015, 27, 1742–1754. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, H.; Gao, S.; Wang, W.C.; Katiyaragarwal, S.; Huang, H.D.; Raikhel, N.; Jin, H. Arabidopsis Argonaute 2 regulates innate immunity via miRNA393∗-mediated silencing of a Golgi-localized SNARE gene, MEMB12. Mol. Cell 2011, 42, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Odokonyero, D.; Mendoza, M.R.; Alvarado, V.Y.; Zhang, J.; Wang, X.; Scholthof, H.B. Transgenic down-regulation of ARGONAUTE2 expression in Nicotiana benthamiana interferes with several layers of antiviral defenses. Virology 2015, 486, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ludman, M.; Burgyán, J.; Fátyol, K. Crispr/Cas9 mediated inactivation of argonaute 2 reveals its differential involvement in antiviral responses. Sci. Rep. 2017, 7, 1010. [Google Scholar] [CrossRef] [PubMed]
- Diermann, N.; Matoušek, J.; Junge, M.; Riesner, D.; Steger, G. Characterization of plant miRNAs and small RNAs derived from Potato Spindle Tuber Viroid (PSTVD) in infected tomato. Biol. Chem. 2010, 391, 1379–1390. [Google Scholar] [CrossRef] [PubMed]
- Bazzini, A.A.; Manacorda, C.A.; Tohge, T.; Conti, G.; Rodriguez, M.C.; Nunesnesi, A.; Villanueva, S.; Fernie, A.R.; Carrari, F.; Asurmendi, S. Metabolic and miRNA profiling of TMV infected plants reveals biphasic temporal changes. PLoS ONE 2011, 6, e28466. [Google Scholar] [CrossRef] [PubMed]
- Morel, J.B.; Godon, C.; Mourrain, P.; Beclin, C.; Boutet, S.; Feuerbach, F.; Proux, F.; Vaucheret, H. Fertile hypomorphic argonaute (ago1) mutants impaired in post-transcriptional gene silencing and virus resistance. Plant Cell 2002, 14, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xian, Z.; Huang, W.; Li, Z. Evidence for the biological function of miR403 in tomato development. Sci. Horticult. 2015, 197, 619–626. [Google Scholar] [CrossRef]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Bao, D.; Ganbaatar, O.; Cui, X.; Yu, R.; Bao, W.; Falk, B.W.; Wuriyanghan, H. Downregulation of genes coding for core RNAi components and disease resistance proteins via corresponding microRNAs might be correlated with successful SMV infection in soybean. Mol. Plant Pathol. 2018, 19, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Liu, J.; Takatoshi, K.; Jongchan, W.; Tolulope, O.; Markus, H.; Thomas, T.; Nam-Hai, C.; Wang, X.J. Deep sequencing of small RNAs specifically associated with Arabidopsis AGO1 and AGO4 uncovers new AGO functions. Plant J. 2011, 67, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Ye, X.; Morris, T.J. Arabidopsis DRB4, AGO1, AGO7, and RDR6 participate in a DCL4-initiated antiviral RNA silencing pathway negatively regulated by DCL1. Proc. Natl. Acad. Sci. USA 2008, 105, 14732–14737. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, K.R.; Mirlohi, S.; Khalaji, F.; Fakhari, Z.; Shiran, B.; Fallahi, H.; Rafiei, F.; Budak, H.; Ebrahimie, E. Differential expression of seven conserved microRNAs in response to abiotic stress and their regulatory network in Helianthus annuus. Front. Plant Sci. 2015, 6, 741. [Google Scholar]
- Hirayama, T.; Shinozaki, K. Perception and transduction of abscisic acid signals: Keys to the function of the versatile plant hormone ABA. Trends Plant Sci. 2007, 12, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Melotto, M.; Underwood, W.; Koczan, J.; Nomura, K.; He, S.Y. Plant stomata function in innate immunity against bacterial invasion. Cell 2006, 126, 969–980. [Google Scholar] [CrossRef] [PubMed]
- Zelicourt, A.D.; Colcombet, J.; Hirt, H. The role of MAPK modules and ABA during abiotic stress signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Johansen, L.K.; Gustafson, A.M.; Kasschau, K.D.; Lellis, A.D.; Zilberman, D.; Jacobsen, S.E.; Carrington, J.C. Genetic and functional diversification of small RNA pathways in plants. PLoS Biol. 2004, 2, E104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Allen, E.; Wilken, A.; Carrington, J.C. Dicer-like 4 functions in trans-acting small interfering RNA biogenesis and vegetative phase change in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2005, 102, 12984–12989. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Denli, A.M.; Hannon, G.J. Biochemical specialization within Arabidopsis RNA silencing pathways. Mol. Cell 2005, 19, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Watanabe, Y. Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions. Proc. Natl. Acad. Sci. USA 2004, 101, 12753–12758. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, R.J.; Hong, L.; Fitzpatrick, K.E.; Amasino, R.M. Dicer-like 1 and Dicer-like 3 redundantly act to promote flowering via repression of flowering locus c in Arabidopsis thaliana. Genetics 2007, 176, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O. Origin, biogenesis, and activity of plant microRNAs. Cell 2009, 136, 669–687. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Bao, F.S.; Xie, Z. Small RNA deep sequencing reveals role for Arabidopsis thaliana RNA-dependent RNA polymerases in viral siRNA biogenesis. PLoS ONE 2009, 4, e4971. [Google Scholar] [CrossRef]
- Liao, Y.W.; Liu, Y.R.; Liang, J.Y.; Wang, W.P.; Zhou, J.; Xia, X.J.; Zhou, Y.H.; Yu, J.Q.; Shi, K. The relationship between the plant-encoded RNA-dependent RNA polymerase 1 and alternative oxidase in tomato basal defense against tobacco mosaic virus. Planta 2015, 241, 641–650. [Google Scholar] [CrossRef]
- Yu, D.; Fan, B.; Macfarlane, S.A.; Chen, Z. Analysis of the involvement of an inducible Arabidopsis RNA-dependent RNA polymerase in antiviral defense. Mol. Plant-Microbe Interact. 2003, 16, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Curaba, J.; Chen, X. Biochemical activities of Arabidopsis RNA-dependent RNA polymerase 6. J. Biol. Chem. 2008, 283, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Vaistij, F.E.; Jones, L. Compromised virus-induced gene silencing in RDR6-deficient plants. Plant Physiol. 2009, 149, 1399–1407. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.B.; Wu, Q.F.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.M.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Wang, M.; Gao, Z.; Zhu, C.; Guo, X. Isolation of a novel RNA-dependent RNA polymerase 6 from Nicotiana glutinosa, NgRDR6, and analysis of its response to biotic and abiotic stresses. Mol. Biol. Rep. 2011, 38, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Kasschau, K.D.; Fahlgren, N.; Chapman, E.J.; Sullivan, C.M.; Cumbie, J.S.; Givan, S.A.; Carrington, J.C. Genome-wide profiling and analysis of Arabidopsis siRNAs. PLoS Biol. 2007, 5, e57. [Google Scholar] [CrossRef] [PubMed]
- Raja, P.; Sanville, B.C.; Buchmann, R.C.; Bisaro, D.M. Viral genome methylation as an epigenetic defense against geminiviruses. J. Virol. 2008, 82, 8997–9007. [Google Scholar] [CrossRef] [PubMed]
- Verlaan, M.G.; Hutton, S.F.; Ibrahem, R.M.; Kormelink, R.; Visser, R.G.; Scott, J.W.; Edwards, J.D.; Bai, Y. The tomato yellow leaf curl virus resistance genes Ty-1 and Ty-3 are allelic and code for DFDGD-class RNA-dependent RNA polymerases. PLoS Genet. 2013, 9, e1003399. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega 5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Hunter, L.J.R.; Westwood, J.H.; Heath, G.; Macaulay, K.; Smith, A.G.; MacFarlane, S.A.; Palukaitis, P.; Carr, J.P. Regulation of RNA-dependent RNA polymerase 1 and isochorismate synthase gene expression in Arabidopsis. PLoS ONE 2013, 8, e66530. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Yang, S.; Yan, Y.; Xiao, Z.; Cheng, J.; Wu, J.; Qiu, A.; Yan, L.; Mou, S.; Guan, D. CaWRKY6 transcriptionally activates CaWRKY40, regulates ralstonia solanacearum resistance, and confers high-temperature and high-humidity tolerance in pepper. J. Exp. Bot. 2015, 66, 3163–3174. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−△△Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Structural analysis of CaAGOs (A), CaRDRs (B) and CaDCLs (C) in pepper. Domains are indicated as boxes in different colors.
Figure 1.
Structural analysis of CaAGOs (A), CaRDRs (B) and CaDCLs (C) in pepper. Domains are indicated as boxes in different colors.

Figure 2.
Phylogenetic analysis of putative Argonaute protein (AGO), RNA-dependent RNA polymerase (RDR) and Dicer-like protein (DCL) proteins of pepper. Unrooted neighbor-joining trees constructed from multiple alignments of total (A) AGO, (B) DCL and (C) RDR protein sequences of pepper, tomato, Arabidopsis, tobacco and potato. Bootstrap support values from 1000 replications are indicated above the branches. Each gene family is divided into different clades as shown in the figure. Sequences of tomato, Arabidopsis, tobacco and potato were downloaded from the NCBI database. The red star indicated the proteins in pepper.
Figure 2.
Phylogenetic analysis of putative Argonaute protein (AGO), RNA-dependent RNA polymerase (RDR) and Dicer-like protein (DCL) proteins of pepper. Unrooted neighbor-joining trees constructed from multiple alignments of total (A) AGO, (B) DCL and (C) RDR protein sequences of pepper, tomato, Arabidopsis, tobacco and potato. Bootstrap support values from 1000 replications are indicated above the branches. Each gene family is divided into different clades as shown in the figure. Sequences of tomato, Arabidopsis, tobacco and potato were downloaded from the NCBI database. The red star indicated the proteins in pepper.

Figure 3.
Heatmap showing the expression pattern of CaAGO, CaDCL and CaRDR genes in various organs. Relative expression levels of CaAGO, CaDCL and CaRDR genes in pepper were determined by real-time quantitative polymerase chain reaction (qRT-PCR) at corresponding organs, including roots, leaves, flowers, stems and fruit. The CaUbi3 was used as the reference gene. The color scale for each value is shown on the down pane.
Figure 3.
Heatmap showing the expression pattern of CaAGO, CaDCL and CaRDR genes in various organs. Relative expression levels of CaAGO, CaDCL and CaRDR genes in pepper were determined by real-time quantitative polymerase chain reaction (qRT-PCR) at corresponding organs, including roots, leaves, flowers, stems and fruit. The CaUbi3 was used as the reference gene. The color scale for each value is shown on the down pane.

Figure 4.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression in response to viral infections. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.
Figure 4.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression in response to viral infections. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.

Figure 5.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression under abiotic stress. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.
Figure 5.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression under abiotic stress. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.

Figure 6.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression under phytohormone and H2O2 treatment. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.
Figure 6.
qRT-PCR analyses of CaAGOs (A), CaDCLs (B) and CaRDRs (C) expression under phytohormone and H2O2 treatment. The pepper CaUbi3 was used as the reference gene, and three biological replicates were performed for these experiments. Error bars indicate the standard errors. Asterisks indicate the significant differences (p < 0.05) between control and treatment.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of CaDCL, CaAGO, and CaRDR genes in pepper.
| Gene Name | Accession Number | Chr | Range | CDS (bp) | Protein (aa) | Mw (kD) |
|---|---|---|---|---|---|---|
| CaAGO1a | Capana06g000835 | 6 | 13,219,325–13,229,295 | 3156 | 1051 | 116.40 |
| CaAGO1b | Capana03g001538 | 3 | 28,791,304–28,798,662 | 3339 | 1112 | 122.52 |
| CaAGO2 | Capana02g001299 | 2 | 119,455,171–119,460,059 | 3027 | 1008 | 112.42 |
| CaAGO4a | Capana01g001805 | 1 | 70,676,331–70,686,308 | 2733 | 910 | 102.02 |
| CaAGO4b | Capana06g000702 | 6 | 11,153,712–11,162,578 | 2730 | 909 | 101.33 |
| CaAGO4d | Capana08g001169 | 8 | 125,940,932–125,950,280 | 2646 | 881 | 98.44 |
| CaAGO5 | Capana06g000572 | 6 | 8,647,138–8,654,022 | 3048 | 1015 | 111.73 |
| CaAGO6 | Capana07g001363 | 7 | 175,512,846–175,528,611 | 2718 | 905 | 101.76 |
| CaAGO7 | Capana01g002131 | 1 | 114,240,252–114,244,871 | 3036 | 1011 | 115.15 |
| CaAGO10a | Capana03g004637 | 3 | 261,275,238–261,283,423 | 2967 | 988 | 110.83 |
| CaAGO10b | Capana09g000331 | 9 | 10,489,518–10,495,890 | 2844 | 947 | 107.02 |
| CaAGO15 | Capana03g001292 | 3 | 22,639,388–22,645,377 | 2682 | 893 | 100.94 |
| CaDCL1 | Capana10g000732 | 10 | 43,673,571–43,714,716 | 5736 | 1911 | 214.29 |
| CaDCL2 | Capana12g002509 | 12 | 220,158,256–220,168,547 | 4206 | 1401 | 158.07 |
| CaDCL3 | Capana08g000619 | 8 | 95,835,542–95,908,094 | 4956 | 1651 | 186.11 |
| CaDCL4 | Capana07g000265 | 7 | 11,520,463–11,570,768 | 4854 | 1617 | 182.23 |
| CaRDR1 | Capana11g001709 | 11 | 191,597,130–191,608,269 | 3351 | 1116 | 127.40 |
| CaRDR2 | Capana03g000988 | 3 | 16,378,521–16,387,609 | 3393 | 1130 | 128.28 |
| CaRDR3a | Capana07g000168 | 7 | 8,346,974–8,399,430 | 3024 | 1007 | 114.57 |
| CaRDR3b | Capana08g000377 | 8 | 51,083,159–51,128,885 | 5049 | 1682 | 191.94 |
| CaRDR5 | Capana09g000243 | 9 | 7,491,673–7,505,214 | 3204 | 1067 | 122.95 |
| CaRDR6 | Capana05g000179 | 5 | 2,848,056–2,858,044 | 3591 | 1196 | 135.98 |
CDS: Coding sequence; MW: Molecular weight.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Qin, L.; Mo, N.; Muhammad, T.; Liang, Y. Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.). Int. J. Mol. Sci. 2018, 19, 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041038
AMA Style
Qin L, Mo N, Muhammad T, Liang Y. Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.). International Journal of Molecular Sciences. 2018; 19(4):1038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041038
Chicago/Turabian StyleQin, Lei, Ning Mo, Tayeb Muhammad, and Yan Liang. 2018. "Genome-Wide Analysis of DCL, AGO, and RDR Gene Families in Pepper (Capsicum Annuum L.)" International Journal of Molecular Sciences 19, no. 4: 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041038
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.