Transplantable Melanomas in Hamsters and Gerbils as Models for Human Melanoma. Sensitization in Melanoma Radiotherapy—From Animal Models to Clinical Trials

 , , , and
, , , and
Abstract
:
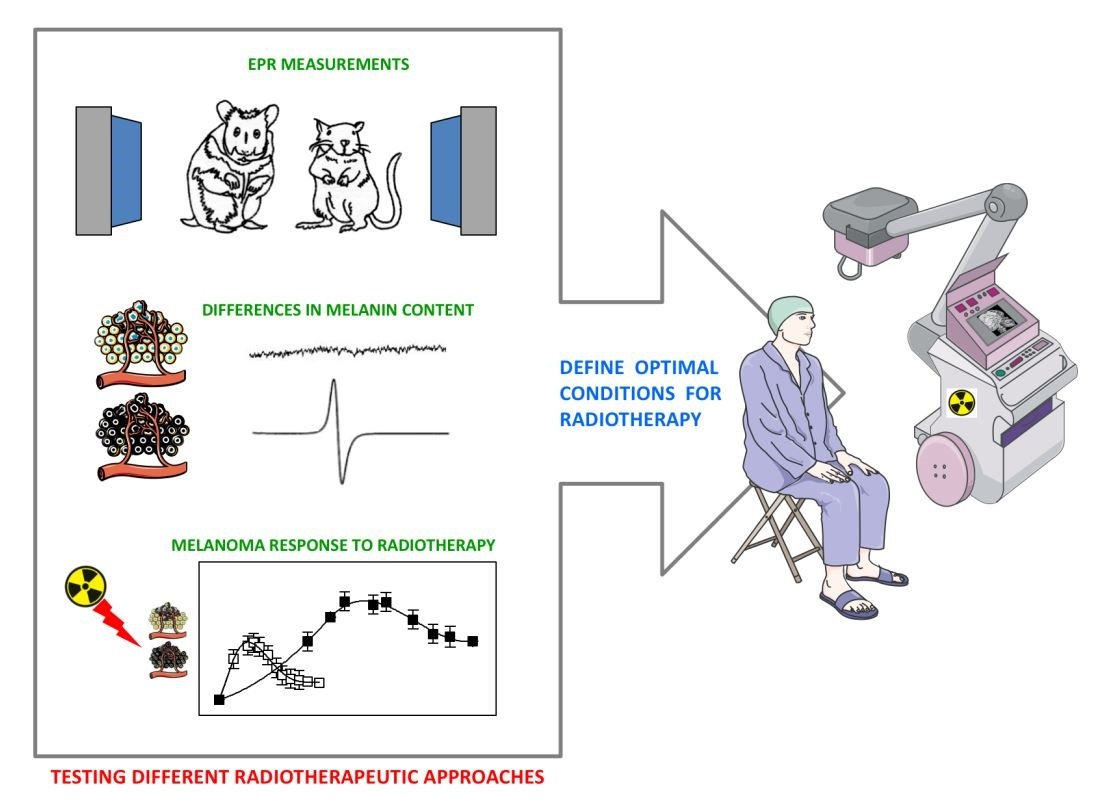
1. Introduction
2. Melanin Functions and Their Radioprotective Role in Melanoma
3. Hamster as a Model of Spontaneously Occurring Melanoma
3.1. Bomirski Hamster Melanoma
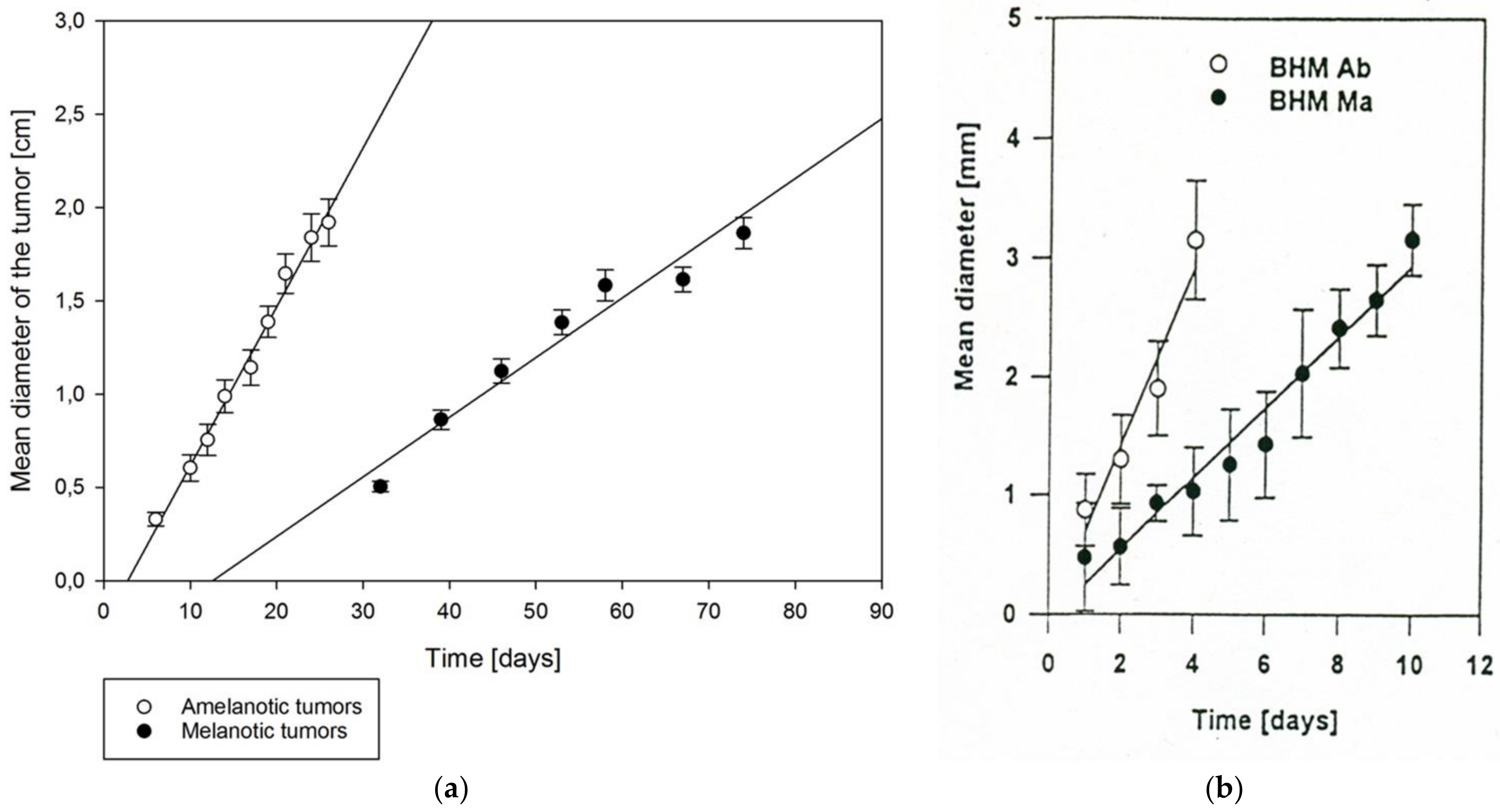
3.2. MI Melanoma and Ab-455
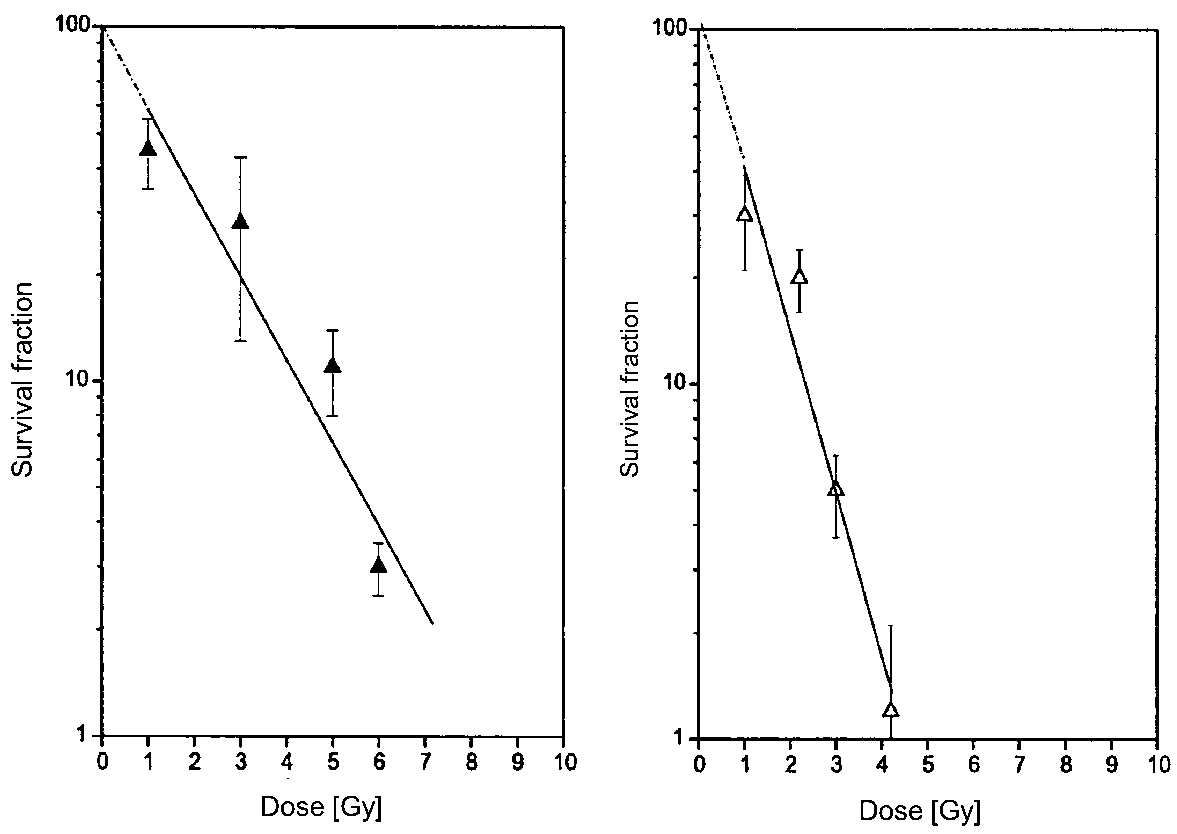
3.3. Radiosensitivity of BHM Growing in the Skin
3.3.1. Effects of Low-LET Radiation
3.3.2. Radio-Chelation Therapy
3.3.3. Effects of Neutrons
3.4. Radiosensitivity of BHM Tumors Transplanted in the Eye
3.4.1. Effects of Low-LET Radiation
3.4.2. Radiotherapy Using Proton Beam Irradiation
3.4.3. Radio-Phototherapy
4. Gerbils as Animal Models for Chemically-Induced Melanomas
4.1. Zeman UJ90 Melanoma
4.2. Irradiation of Zeman UJ90 Melanoma
5. Human Melanoma
5.1. Radiosensitivity of Human Skin Melanoma/Why Is Radiation Not Used in the Treatment of Human Skin Melanoma
5.2. Radiosensitization of Melanoma Cells through Inhibition of Melanoma Pigmentation
5.3. Radiosensitivity of Uveal Melanoma Tumors
5.3.1. Brachytherapy in Clinical Practice
5.3.2. Proton Beam Radiotherapy (PBRT) of Uveal Melanoma
5.3.3. Proteomic Study of Human Skin Melanoma Cells (BLM) Treated with Proton Beam Irradiation
5.3.4. Radio-Phototherapy of Uveal Melanoma
5.3.5. Radio-Chelation Therapy in Clinical Trials
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Urteaga, B.O.; Pack, G.T. On the antiquity of melanoma. Cancer 1966, 19, 607–610. [Google Scholar] [CrossRef]
- Slominski, R.M.; Zmijewski, M.A.; Slominski, A.T. The role of melanin pigment in melanoma. Exp. Dermatol. 2015, 24, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Bomirski, A.; Dominiczak, T.; Nowinska, L. Spontaneous transplantable melanoma in the golden hamster (Mesocricetus auratus). Acta Unio Int. Contra Cancrum 1962, 18, 178–180. [Google Scholar] [PubMed]
- Bomirski, A. Biological Properties of Transplantable Melanomas in the Syrian Hamster during 16 Years of Maintenance by Serial Passages. Habilitation Thesis, Medical School of Gdansk, Gdansk, Poland, 1977. [Google Scholar]
- Bomirski, A.; Słominski, A.; Bigda, J. The natural history of a family of transplantable melanomas in hamsters. Cancer Metastasis Rev. 1988, 7, 95–118. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Paus, R. Bomirski melanomas: A versatile and powerful model for pigment cell and melanoma research. Int. J. Oncol. 1993, 2, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Pajak, S.; Cieszka, K.; Plonka, P.; Lukiewicz, S.; Mihm, M.; Slominski, A. Transplantable melanomas in gerbils (Meriones unguiculatus). I. Origin, morphology and growth rate. Anticancer Res. 1996, 16, 1203–1208. [Google Scholar] [PubMed]
- Plonka, P.M.; Slominski, A.T.; Pajak, S.; Urbanska, K. Transplantable melanomas in gerbils (Meriones unguiculatus). II: Melanogenesis. Exp. Dermatol. 2003, 12, 356–364. [Google Scholar] [CrossRef] [PubMed]
- Billingham, R.E.; Silvers, W.K. The melanocytes of mammals. Q. Rev. Biol. 1960, 35, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Van Den Bossche, K.; Naeyaert, J.-M.; Lambert, J. The Quest for the Mechanism of Melanin Transfer. Traffic 2006, 7, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Lazova, R.; Pawelek, J.M. Why do melanomas get so dark? Exp. Dermatol. 2009, 18, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Lazova, R.; Klump, V.; Pawelek, J. Autophagy in cutaneous malignant melanoma. J. Cutan. Pathol. 2010, 37, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Lembo, S.; Di Caprio, R.; Micillo, R.; Balato, A.; Monfrecola, G.; Panzella, L.; Napolitano, A. Light-independent pro-inflammatory and pro-oxidant effects of purified human hair melanins on keratinocyte cell cultures. Exp. Dermatol. 2017, 26, 592–594. [Google Scholar] [CrossRef] [PubMed]
- Płonka, P.M.; Picardo, M.; Slominski, A.T. Does melanin matter in the dark? Exp. Dermatol. 2017, 26, 595–597. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Paus, R.; Mihm, M.C. Inhibition of melanogenesis as an adjuvant strategy in the treatment of melanotic melanomas: Selective review and hypothesis. Anticancer Res. 1998, 18, 3709–3716. [Google Scholar] [PubMed]
- Brożyna, A.A.; Jóźwicki, W.; Carlson, J.A.; Slominski, A.T. Melanogenesis affects overall and disease-free survival in patients with stage III and IV melanoma. Hum. Pathol. 2013, 44, 2071–2074. [Google Scholar] [CrossRef] [PubMed]
- Englaro, W.; Rezzonico, R.; Durand-Clement, M.; Lallemand, D.; Ortonne, J.-P.; Ballotti, R. Mitogen-activated protein kinase pathway and AP-1 are activated during cAMP-induced melanogenesis in B-16 melanoma cells. J. Biol. Chem. 1995, 270, 24315–24320. [Google Scholar] [CrossRef] [PubMed]
- Ortonne, J.-P.; Ballotti, R. Melanocyte biology and melanogenesis: What’s new? J. Dermatol. Treat. 2000, 11, 15–26. [Google Scholar] [CrossRef]
- Slominski, A.T.; Tobin, D.J.; Shibahara, S.; Wortsman, J. Melanin Pigmentation in Mammalian Skin and Its Hormonal Regulation. Physiol. Rev. 2004, 84, 1155–1228. [Google Scholar] [CrossRef] [PubMed]
- Spagnolo, F.; Queirolo, P. Upcoming strategies for the treatment of metastatic melanoma. Arch. Dermatol. Res. 2012, 304, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Jean, D.; Bar-Eli, M. Regulation of tumor growth and metastasis of human melanoma by the CREB transcription factor family. Mol. Cell. Biochem. 2000, 212, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Buscà, R.; Bertolotto, C.; Ortonne, J.-P.; Ballotti, R. Inhibition of the phosphatidylinositol 3-kinase/p70(S6)-kinase pathway induces B16 melanoma cell differentiation. J. Biol. Chem. 1996, 271, 31824–31830. [Google Scholar] [CrossRef] [PubMed]
- Curtin, J.A.; Busam, K.; Pinkel, D.; Bastian, B.C. Somatic activation of KIT in distinct subtypes of melanoma. J. Clin. Oncol. 2006, 24, 4340–4346. [Google Scholar] [CrossRef]
- Albino, A.P.; Le Strange, R.; Oliff, A.I.; Furth, M.E.; Old, L.J. Transforming ras genes from human melanoma: A manifestation of tumour heterogeneity? Nature 1984, 308, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Padua, R.A.; Barrass, N.; Currie, G.A. A novel transforming gene in a human malignant melanoma cell line. Nature 1984, 311, 671–673. [Google Scholar] [CrossRef] [PubMed]
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Hingorani, S.R.; Jacobetz, M.A.; Robertson, G.P. Suppression of BRAFV599E in Human Melanoma Abrogates Transformation Suppression of BRAF V599E in Human Melanoma Abrogates Transformation 1. Cancer Res. 2003, 63, 5198–5202. [Google Scholar] [PubMed]
- Wellbrock, C.; Ogilvie, L.; Hedley, D.; Karasarides, M.; Martin, J.; Niculescu-Duvaz, D.; Springer, C.J.; Marais, R. V599E B-RAF is an Oncogene in Melanocytes. Cancer Res. 2004, 64, 2338–2342. [Google Scholar] [CrossRef] [PubMed]
- Smalley, K.S.M. Understanding Melanoma Signaling Networks as the Basis for Molecular Targeted Therapy. J. Investig. Dermatol. 2010, 130, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Pawelek, J. Animals under the sun: Effects of ultraviolet radiation on mammalian skin. Clin. Dermatol. 1998, 16, 503–515. [Google Scholar] [CrossRef]
- Wood, J.M.; Jimbow, K.; Boissy, R.E.; Slominski, A.; Plonka, P.M.; Slawinski, J.; Wortsman, J.; Tosk, J. What’s the use of generating melanin? Exp. Dermatol. 1999, 8, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Meredith, P.; Sarna, T. The physical and chemical properties of eumelanin. Pigment Cell Res. 2006, 19, 572–594. [Google Scholar] [CrossRef] [PubMed]
- Meredith, P.; Powell, B.J.; Riesz, J.; Nighswander-Rempel, S.P.; Pederson, M.R.; Moore, E.G. Towards structure–property–function relationships for eumelanin. Soft Matter 2006, 2, 37–44. [Google Scholar] [CrossRef]
- D’Ischia, M.; Napolitano, A.; Pezzella, A.; Meredith, P.; Sarna, T. Chemical and structural diversity in eumelanins: Unexplored bio-optoelectronic materials. Angew. Chem. Int. Ed. 2009, 48, 3914–3921. [Google Scholar] [CrossRef] [PubMed]
- Commoner, B.; Townsend, J.; Pake, G.E. Free radicals in biological materials. Nature 1954, 174, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Blois, M.S.; Zahlan, A.B.; Maling, J.E. Electron Spin Resonance Studies on Melanin. Biophys. J. 1964, 4, 471–490. [Google Scholar] [CrossRef]
- Sarna, T.; Pilas, B.; Land, E.J.; Truscott, T.G. Interaction of radicals from water radiolysis with melanin. BBA Gen. Subj. 1986, 883, 162–167. [Google Scholar] [CrossRef]
- Sarna, T.; Plonka, P.M. Biophysical Studies of Melanin. Biomed. EPR Part A Free Radic. Met. Med. Physiol. 2005, 125–146. [Google Scholar] [CrossRef]
- Sarna, T.; Hyde, J.; Swartz, H. Ion-exchange in melanin: An electron spin resonance study with lanthanide probes. Science 1976, 1132–1134. [Google Scholar] [CrossRef]
- Zecca, L.; Swartz, H. Total and paramagnetic metals in human substantia nigra and its neuromelanin. J. Neural Transm. 1993, 5, 203–213. [Google Scholar] [CrossRef]
- Korytowski, W.; Pilas, B.; Sarna, T.; Kalyanaraman, B. Photoinduced generation of hydrogen peroxide and hydroxyl radicals in melanins. Photochem. Photobiol. 1987, 45, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Borovanský, J.; Elleder, M. Melanosome Degradation: Fact or Fiction. Pigment Cell Res. 2003, 16, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Samokhvalov, A.; Hong, L.; Liu, Y.; Garguilo, J.; Nemanich, R.J.; Edwards, G.S.; Simon, J.D. Oxidation potentials of human eumelanosomes and pheomelanosomes. Photochem. Photobiol. 2005, 81, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.D.; Peles, D.N. The red and the black. Acc. Chem. Res. 2010, 43, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- ElObeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological Properties of Melanin and its Function in Health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Lukiewicz, S.; Pilas, B.; Nowicka, J. Molecular and cellular basis of different radiosensitivity in pigmented and non-pigmented hamster melanoma-cells. In Phenotypic Expression in Pigmented Cells; Seiji, H., Ed.; University of Tokyo Press: Tokyo, Japan, 1981; pp. 647–653. [Google Scholar]
- Lukiewicz, S. Interference with endogenous radioprotectors as a method of radiosensitization. In IAEA’s Modification of Radiosensitivity of Biological Systems; International Atomic Energy Agency: Vienna, Austria, 1976. [Google Scholar]
- Sarna, T.; Dulȩba, A.; Korytowski, W.; Swartz, H. Interaction of melanin with oxygen. Arch. Biochem. Biophys. 1980, 200, 140–148. [Google Scholar] [CrossRef]
- Pathak, M.A.; Riley, F.J.; Fitzpatrick, T.B.; Curwen, W.L. Melanin formation in human skin induced by long-wave ultra-violet and visible light. Nature 1962, 193, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Nakagawa, A.; Muramatsu, T.; Yamashina, Y.; Shirai, T.; Hashimoto, M.W.; Ishigaki, Y.; Ohnishi, T.; Mori, T. Supranuclear melanin caps reduce ultraviolet induced DNA photoproducts in human epidermis. J. Investig. Dermatol. 1998, 110, 806–810. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, A.D.; Howell, R.C.; Jiang, Z.; Bryan, R.A.; Gerfen, G.; Chen, C.C.; Mah, D.; Cahill, S.; Casadevall, A.; Dadachova, E. Physico-chemical evaluation of rationally designed melanins as novel nature-inspired radioprotectors. PLoS ONE 2009, 4, e7229. [Google Scholar] [CrossRef] [PubMed]
- Turick, C.E.; Ekechukwu, A.A.; Milliken, C.E.; Casadevall, A.; Dadachova, E. Gamma radiation interacts with melanin to alter its oxidation-reduction potential and results in electric current production. Bioelectrochemistry 2011, 82, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Rózanowska, M.; Sarna, T.; Land, E.J.; Truscott, T.G. Free radical scavenging properties of melanin interaction of eu- and pheo-melanin models with reducing and oxidising radicals. Free Radic. Biol. Med. 1999, 26, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Land, E.J.; Ramsden, C.A.; Riley, P.A. Quinone Chemistry and Melanogenesis. Methods Enzymol. 2004, 378, 88–109. [Google Scholar] [CrossRef] [PubMed]
- Pey, A.L.; Martinez, A.; Charubala, R.; Maitland, D.J.; Teigen, K.; Calvo, A.; Pfleiderer, W.; Wood, J.M.; Schallreuter, K.U. Specific interaction of the diastereomers 7(R)- and 7(S)-tetrahydrobiopterin with phenylalanine hydroxylase: Implications for understanding primapterinuria and vitiligo. FASEB J. 2006, 20, 2130–2132. [Google Scholar] [CrossRef] [PubMed]
- Gallez, B.; Neveu, M.A.; Danhier, P.; Jordan, B.F. Manipulation of tumor oxygenation and radiosensitivity through modification of cell respiration. A critical review of approaches and imaging biomarkers for therapeutic guidance. Biochim. Biophys. Acta Bioenergy 2017, 1858, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Grossi, G.F.; Durante, M.; Gialanella, G.; Pugliese, M.; Mosse, I. Effects of melanin on high- and low-linear energy transfer (LET) radiation response of human epithelial cells. Radiat. Environ. Biophys. 1998, 37, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Sava, V.; Mosquera, D.; Song, S.; Cardozo-Pelaez, F.; Sánchez-Ramos, J.R. Effects of melanin and manganese on DNA damage and repair in PC12-derived neurons. Free Radic. Biol. Med. 2004, 36, 1144–1154. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gaidamakova, E.K.; Grichenko, O.; Matrosova, V.Y.; Hoeke, V.; Klimenkova, P.; Conze, I.H.; Volpe, R.P.; Tkavc, R.; Gostinčar, C.; et al. Across the tree of life, radiation resistance is governed by antioxidant Mn2+, gauged by paramagnetic resonance. Proc. Natl. Acad. Sci. USA 2017, 114, 201713608. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.R. History of the Capture and Domestication of the Syrian Golden Hamster (Mesocricetus auratus Waterhouse). In The Hamster; Springer: Boston, MA, USA, 1985; pp. 3–20. [Google Scholar]
- McGuire, K.L.; Duncan, W.R.; Tucker, P.W. Syrian hamster DNA shows limited polymorphism at class I-like loci. Immunogenetics 1985, 22, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Silvers, W.K.; Gasser, D.L.; Murphy, M.R. Number of Histocompatibility Loci in Syrian Hamsters. J. Immunol. 1975, 115, 1309–1311. [Google Scholar] [PubMed]
- Valentine, H.; Daugherity, E.K.; Singh, B.; Maurer, K.J. The Experimental Use of Syrian Hamsters. In The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Elsevier: Amsterdam, The Netherlands, 2012; pp. 875–906. ISBN 9780123809209. [Google Scholar]
- Wahl-Jensen, V.; Bollinger, L.; Safronetz, D.; de Kok-Mercado, F.; Scott, D.; Ebihara, H. Use of the Syrian Hamster as a New Model of Ebola Virus Disease and Other Viral Hemorrhagic Fevers. Viruses 2012, 4, 3754–3784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, S. Origin of the Golden Hamster Cricetus auratus as a Laboratory Animal. Nature 1948, 162, 256–257. [Google Scholar] [CrossRef] [PubMed]
- Chorobik, P.; Czaplicki, D.; Ossysek, K.; Bereta, J. Salmonella and cancer: From pathogens to therapeutics. Acta Biochim. Pol. 2013, 60, 285–297. [Google Scholar] [PubMed]
- Slominski, A.; Wortsman, J.; Plonka, P.M.; Schallreuter, K.U.; Paus, R.; Tobin, D.J. Hair follicle pigmentation. J. Investig. Dermatol. 2005, 124, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Pour, P.; Althoff, J.; Salmasi, S.Z.; Stepan, K. Spontaneous Tumors and Common Diseases in Three Types of Hamsters2. JNCI J. Natl. Cancer Inst. 1979, 63, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Kusewitt, D.; Ley, R. Animal Models of Melanoma. Cancer Surv. 1996, 26, 35–70. [Google Scholar] [CrossRef] [PubMed]
- Ghadially, F.N.; Barker, J.F. The histogenesis of experimentally induced melanotic tumours in the Syrian hamster (Cricetus auratus). J. Pathol. Bacteriol. 1960, 79, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Fortner, J.G.; Allen, A.C. Hitherto Unreported Malignant Melanomas in the Syrian Hamster: An Experimental Counterpart of the Human Malignant Melanomas. Cancer Res. 1958, 18, 98–104. [Google Scholar]
- Homburger, F. Chemical carcinogenesis in the Syrian golden hamster. Cancer 1969, 23, 313–338. [Google Scholar] [CrossRef]
- Pawlowski, A.; Lea, P.J. Nevi and melanoma induced by chemical carcinogens in laboratory animals: Similarities and differences with human lesions. J. Cutan. Pathol. 1983, 10, 81–110. [Google Scholar] [CrossRef] [PubMed]
- Bardeesy, N.; Wong, K.K.; DePinho, R.A.; Chin, L. Animal models of melanoma: Recent advances and future prospects. Adv. Cancer Res. 2000, 79, 123–156. [Google Scholar] [CrossRef] [PubMed]
- Greene, H.S.N. A Spontaneous Melanoma in the Hamster with a Propensity for Amelanotic Alteration and Sarcomatous Transformation during Transplantation. Cancer Res. 1958, 18, 422–425. [Google Scholar] [PubMed]
- Burgess, S.E.; Chang, S.; Svitra, P.; Driller, J.; Lizzi, F.L.; Coleman, D.J. Effect of hyperthermia on experimental choroidal melanoma. Br. J. Ophthalmol. 1985, 69, 854–860. [Google Scholar] [CrossRef] [PubMed]
- Franken, K.A.P.; van Delft, J.L.; Dubbelman, T.M.A. R.; de Wolff-Rouendaal, D.; Oosterhuis, J.A.; Star, W.M.; Marijnissen, H.P.A. Hematoporphyrin derivative photoradiation treatment of experimental malignant melanoma in the anterior chamber of the rabbit. Curr. Eye Res. 1985, 4, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Fortner, J.G. Spontaneous tumors, including gastrointestinal neoplasms and malignant melanomas, in the syrian hamster. Cancer 1957, 10, 1153–1156. [Google Scholar] [CrossRef]
- Ott, F. Hamster melanoma M Mel 1 as an ascites tumor. I. Comparison of the various melanoma of the Syrian hamster and the mice. Detailed description of the form of ascites. Arch. Klin. Exp. Dermatol. 1969, 234, 362–383. [Google Scholar] [CrossRef] [PubMed]
- Ott, F. Hamster melanoma M Mel 1 as an ascietes tumor. II. Research. Arch. Klin. Exp. Dermatol. 1969, 234, 384–398. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Costantino, R. L-tyrosine induces tyrosinase expression via a posttranscriptional mechanism. Experientia 1991, 47, 721–724. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Carlson, J.A. Melanoma resistance: A bright future for academicians and a challenge for patient advocates. Mayo Clin. Proc. 2014, 89, 429–433. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Kobzdej, D. Transplantability of Bomirski melanomas in Syrian hamsters. Neoplasma 1983, 30, 51–56. [Google Scholar] [PubMed]
- Scislowski, P.W.D. Biochemical characterization of three hamster melanoma variants. II. Glycolysis and oxygen consumption. Int. J. Biochem. 1984, 16, 327–331. [Google Scholar] [CrossRef]
- Słomiński, A. Rapid melanization of Bomirski amelanotic melanoma cells in cell culture. Biosci. Rep. 1983, 3, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Słomiński, A.; Scisłowski, P.W.; Bomirski, A. Tyrosinase activity in primary cell culture of amelanotic melanoma cells. Biosci. Rep. 1983, 3, 1027–1034. [Google Scholar] [CrossRef] [PubMed]
- Słomiński, A. Some properties of Bomirski Ab amelanotic melanoma cells, which underwent spontaneous melanization in primary cell culture. J. Cancer Res. Clin. Oncol. 1985, 109, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Słomiński, A.; Bomirski, A. Phenotypic changes of Ab hamster melanoma during long-term culture. Anticancer Res. 1985, 5, 403–410. [Google Scholar] [PubMed]
- Bomirski, A.; Słomiński, A. Ultrastructural aspects of melanization of hamster Ab amelanotic melanoma in primary cell culture. Acta Derm.-Venereol. 1986, 66, 520–538. [Google Scholar] [PubMed]
- Słomiński, A.; Bomirski, A.; Scisłowski, P.W.; Zołnierowicz, S. Effects of actinomycin D and cycloheximide on the increase in tyrosinase activity of hamster amelanotic melanoma cells in vitro. Biosci. Rep. 1984, 4, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Daniel, P. L-Dopa binding sites in rodent melanoma cells. Biochim. Biophys. Acta Mol. Basis Dis. 1992, 1139, 324–328. [Google Scholar] [CrossRef]
- Slominski, A.; Kim, T.K.; Brozyna, A.A.; Janjetovic, Z.; Brooks, D.L.P.; Schwab, L.P.; Skobowiat, C.; Jóźwicki, W.; Seagroves, T.N. The role of melanogenesis in regulation of melanoma behavior: Melanogenesis leads to stimulation of HIF-1α expression and HIF-dependent attendant pathways. Arch. Biochem. Biophys. 2014, 563, 79–93. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Pawelek, J. MSH binding in Bomirski amelanotic hamster melanoma cells is stimulated by L-tyrosine. Biosci. Rep. 1987, 7, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Moellmann, G.; Kuklinska, E.; Bomirski, A.; Pawelek, J. Positive regulation of melanin pigmentation by two key substrates of the melanogenic pathway, L-tyrosine and L-dopa. J. Cell Sci. 1988, 89. [Google Scholar]
- Slominski, A. L-tyrosine induces synthesis of melanogenesis related proteins. Life Sci. 1989, 45, 1799–1803. [Google Scholar] [CrossRef]
- Slominski, A.; Jastreboff, P.; Pawelek, J. L-tyrosine stimulates induction of tyrosinase activity by MSH and reduces cooperative interactions between MSH receptors in hamster melanoma cells. Biosci. Rep. 1989, 9, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Moellmann, G.; Kuklinska, E. MSH inhibits growth in a line of amelanotic hamster melanoma cells and induces increases in cyclic AMP levels and tyrosinase activity without inducing melanogenesis. J. Cell Sci. 1989, 92, 551–554. [Google Scholar] [PubMed]
- Slominski, A.; Moellmann, G.; Kuklinska, E. L-Tyrosine, L-DOPA, and Tyrosinase as Positive Regulators of the Subcellular Apparatus of Melanogenesis in Bomirski Ab Amelanotic Melanoma Cells. Pigment Cell Res. 1989, 2, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Howe, J.; Costantino, R.; Slominski, A. On the putative mechanism of induction and regulation of melanogenesis by L-tyrosine. Acta Derm.-Venereol. 1991, 71, 150–152. [Google Scholar] [PubMed]
- Slominski, A.; Costantino, R. Molecular mechanism of tyrosinase regulation by L-DOPA in hamster melanoma cells. Life Sci. 1991, 48, 2075–2079. [Google Scholar] [CrossRef]
- Slominski, A.; Friedrich, T. L-DOPA Inhibits In Vitro Phosphorylation of Melanoma Glycoproteins. Pigment Cell Res. 1992, 5, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A. L-tyrosine-binding proteins on melanoma cells. In Vitro Cell. Dev. Biol. Anim. 1991, 27, 735–738. [Google Scholar] [CrossRef]
- Cichorek, M.; Kozlowska, K.; Bryl, E. The activity of caspases in spontaneous and camptothecin-induced death of melanotic and amelanotic melanoma cell. Cancer Biol. Ther. 2007, 6, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M. Camptothecin-induced death of amelanotic and melanotic melanoma cells in different phases of cell cycle. Neoplasma 2011, 58, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Pajak, S.; Subczynski, W.; Panz, T.; Lukiewicz, S. Rate of oxygen consumption of hamster melanoma cells as a factor influencing their radioresistance. Folia Histochem. Cytochem. 1980, 18, 33–40. [Google Scholar]
- Scisłowski, P.W.; Słomiński, A.; Bomirski, A.; Zydowo, M. Metabolic characterization of three hamster melanoma variants. Neoplasma 1985, 32, 593–598. [Google Scholar] [PubMed]
- Scisłowski, P.W.; Słominski, A. The role of NADP-dependent dehydrogenases in hydroxylation of tyrosine in hamster melanoma. Neoplasma 1983, 30, 239–243. [Google Scholar] [PubMed]
- Li, W.; Slominski, R.; Slominski, A.T. High-resolution magic angle spinning nuclear magnetic resonance analysis of metabolic changes in melanoma cells after induction of melanogenesis. Anal. Biochem. 2009, 386, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Cichorek, M.; Kozłowska, K.; Bryl, E. Mitochondrial transmembrane potential in spontaneous and camptothecin-induced apoptosis of melanotic and amelanotic melanoma cells. Neoplasma 2007, 54, 29–36. [Google Scholar] [PubMed]
- Bomirski, A.; Zawrocka-Wrzolkowa, T.; Pautsch, F. Electron microscopic studies on transplantable melanotic and amelanotic melanomas in hamsters. Arch. Dermatol. Forsch. 1973, 246, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Kozlowska, K.; Bomirski, A. Comparison of the surface glycoprotein components in the isolated cells of hamster melanotic and amelanotic melanomas. Arch. Dermatol. 1976, 256, 197–203. [Google Scholar] [CrossRef]
- Kozłowska, K.; Żurawska-Czupa, B. Heterogeneity of the surface material in isolated cells of transplantable hamster melanomas. Cancer Res. 1983, 43, 1168–1171. [Google Scholar] [PubMed]
- Kozłowska, K.; Nowak, J.; Kwiatkowski, B.; Cichorek, M. ESR study of plasmatic membrane of the transplantable melanoma cells in relation to their biological properties. Exp. Toxicol. Pathol. 1999, 51, 89–92. [Google Scholar] [CrossRef]
- Kozłowska, K.; Witkowski, J.M.; Zarzeczna, M.; Cichorek, M. Diversity of the plasma membrane properties of transplantable hamster melanomas with regard to the expression of P-glycoprotein. Folia Histochem. Cytobiol. 1999, 37, 173–177. [Google Scholar] [PubMed]
- Ren, S.; Slominski, A.; Yu, R.K. Glycosphingolipids in Bomirski Transplantable Melanomas in Hamsters. Cancer Res. 1989, 49, 7051–7056. [Google Scholar] [PubMed]
- Ren, S.; Ariga, T.; Scarsdale, J.N.; Zhang, Y.; Slominski, A.; Livingston, P.O.; Ritter, G.; Kushi, Y.; Yu, R.K. Characterization of a hamster melanoma-associated ganglioside antigen as 7-O-acetylated disialoganglioside GD3. J. Lipid Res. 1993, 34, 1565–1572. [Google Scholar] [PubMed]
- Grossniklaus, H.E.; Dithmar, S.; Albert, D.M. Animal models of uveal melanoma. Melanoma Res. 2000, 10, 195–211. [Google Scholar] [CrossRef]
- Kozłowska, K.; Zurawska-Czupa, B. Comparison of the antigenicity of melanotic and amelanotic melanoma cells after the release of surface glycoproteins by trypsin. Arch. Immunol. 1980, 28, 641–644. [Google Scholar]
- Kozlowska, K.; Zurawska-Czupa, B.; Mierzewski, P.; Kostulak, A. Use of the macrophage migration inhibition test to evaluate antigenic differences in golden hamster transplantable melanomas. Int. J. Cancer 1980, 26, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, K.; Kostulak, A. Comparative analysis of surface antigens of transplantable melanoma in hamsters. I. Studies on serological reactivity of sera with isolated melanocytes by using the. Arch. Immunol. 1984, 32, 59–66. [Google Scholar]
- Kozlowska, K.; Cichorek, M.; Zarzeczna, M.; Wojcik, S. Expression of CD44 on two lines of transplantable melanoma cells–relationship with cytokine secretion and tumor progression. Folia Histochem. Cytobiol. 2004, 42, 29–34. [Google Scholar] [PubMed]
- Wachulska, M.; Kozłowska, K.; Cichorek, M. The DNA ploidy and proliferative activity of transplantable melanoma cells in regard to their secretory function. Neoplasma 2005, 52, 280–286. [Google Scholar] [PubMed]
- Słomiński, A.; ŚcisŁowski, P.W.D.; Bomirski, A. Biochemical characterization of three hamster melanoma variants—I. Tyrosinase activity and melanin content. Int. J. Biochem. 1984, 16, 323–326. [Google Scholar] [CrossRef]
- Slominski, A.; Costantino, R.; Howe, J.; Moellmann, G. Molecular mechanism governing melanogenesis in hamster melanomas: Relative abundance of tyrosinase and catalase-B (gp 75). Anticancer Res. 1991, 11, 257–262. [Google Scholar] [PubMed]
- Woźniak, A.; Drewa, T.; Drewa, O.; Woźniak, B.; Schachtschabel, D.O. Activity of antioxidant enzymes and concentrations of thiobarbituric acid reactive substances (TBARS) in melanotic and amelanotic Bomirski melanoma tissues in the golden hamster (Mesocricetus auratus, Waterhouse). Neoplasma 2002, 49, 401–404. [Google Scholar] [PubMed]
- Bomirski, A.; Wrzolkowa, T.; Arendarczyk, M.; Bomirska, M.; Kuklinska, E.; Slominski, A.; Moellmann, G. Pathology and Ultrastructural Characteristics of a Hypomelanotic Variant of Transplantable Hamster Melanoma With Elevated Tyrosinase Activity. J. Investig. Dermatol. 1987, 89, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Słomiński, A.; Ścisłowski, P.W.D.; Arendarczyk, M. Effect of tissue selection on melanization of MI hamster melanoma. Neoplasma 1984, 31, 551–556. [Google Scholar] [PubMed]
- Pajak, S. Role of Endogenous Radioprotectors in Radioresistance of the Melanotic and Amelanotic form of Bomirski Melanoma. Ph.D. Thesis, Jagiellonian University, Kraków, Poland, 1980. [Google Scholar]
- Gurbiel, R. Oxygen Role in Radiosensitivity of Pigmented Bomirski Hamster Melanoma. Ph.D. Thesis, Jagiellonian University, Kraków, Poland, 1984. (In Polish). [Google Scholar]
- Urbanska, K. Radiosensitization of Pigmented Cells and Animal Melanoma Tumors to Ionizing Radiation. Habilitation Thesis, Jagiellonian University, Kraków, Poland, 2000. (In Polish). [Google Scholar]
- Cao, J.; Jager, M.J. Animal Eye Models for Uveal Melanoma. Ocul. Oncol. Pathol. 2015, 1, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Schuitmaker, J.J.; Vrensen, G.F.J.M.; Van Delft, J.L.; De Wolff-Rouendaal, D.; Dubbelman, T.M.A.R.; De Wolf, A. Morphologic effects of bacteriochlorin a and light in vivo on intraocular melanoma. Investig. Ophthalmol. Vis. Sci. 1991, 32, 2683–2688. [Google Scholar]
- Römer, T.J.; van Delft, J.L.; de Wolff-Rouendaal, D.; Jager, M.J. Hamster Greene melanoma implanted in the anterior chamber of a rabbit eye: A reliable tumor model? Ophthalmic Res. 1992, 24, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.K.; Huh, K.; Gragoudas, E.S.; Young, L.H. Establishment of pigmented choroidal melanomas in a rabbit model. Retina 1994, 14, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Romanowska, B.; Kukiełczak, B.; Bryk, J.; Mirkiewicz-Sieradzka, B.; Heitzmann, J.; Lukiewicz, S. New technique for implanting Bomirski melanoma into the anterior chamber of Syrian hamster eyes. Klin. Oczna 1995, 97, 324–327. [Google Scholar] [PubMed]
- Urbanska, K.; Romanowska-Dixon, B.; Elas, M.; Pajak, S.; Paziewski, E.; Bryk, J.; Kukielczak, B.; Slominski, A.; Zygulska-Mach, H.; Lukiewicz, S. Experimental ruthenium plaque therapy of amelanotic and melanotic melanomas in the hamster eye. Melanoma Res. 2000, 10, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Romanowska-Dixon, B.; Urbanska, K.; Elas, M.; Pajak, S.; Zygulska-Mach, H.; Miodonski, A. Angiomorphology of the pigmented Bomirski melanoma growing in hamster eye. Ann. Anat. 2001, 183, 559–565. [Google Scholar] [CrossRef]
- Romanowska-Dixon, B.; Elas, M.; Swakoń, J.; Sowa, U.; Ptaszkiewicz, M.; Szczygieł, M.; Krzykawska, M.; Olko, P.; Urbańska, K. Metastasis inhibition after proton beam, β- and γ-irradiation of melanoma growing in the hamster eye. Acta Biochim. Pol. 2013, 60, 307–311. [Google Scholar] [PubMed]
- Ogata, T.; Teshima, T.; Kagawa, K.; Hishikawa, Y.; Takahashi, Y.; Kawaguchi, A.; Suzumoto, Y.; Nojima, K.; Furusawa, Y.; Matsuura, N. Particle Irradiation Suppresses Metastatic Potential of Cancer Cells Particle Irradiation Suppresses Metastatic Potential of Cancer Cells. Cancer Res. 2005, 65, 113–120. [Google Scholar] [PubMed]
- Takahashi, Y.; Teshima, T.; Kawaguchi, N.; Hamada, Y.; Mori, S.; Madachi, A.; Ikeda, S.; Mizuno, H.; Ogata, T.; Nojima, K.; et al. Heavy ion irradiation inhibits in vitro angiogenesis even at sublethal dose. Cancer Res. 2003, 63, 4253–4257. [Google Scholar] [PubMed]
- Kukielczak, B.; Cieszka, K.; Matuszak, Z. Experimental photodynamic therapy of Bomirski hamster melanoma using merocyanine 540 and visible light. Curr Top. Biophys. 1995, 19, 66–70. [Google Scholar]
- Kukielczak, B.; Romanowska, B.; Bryk, J. Gamma radiation and MC540 photosensitization of melanoma in the hamster’s eye. Melanoma Res. 1999, 9, 115–124. [Google Scholar] [CrossRef] [PubMed]
- ITIS Integrated Taxonomic Information System. Available online: https://www.itis.gov/ (accessed on 11 October 2017).
- Gulotta, E.F. Meriones unguiculatus. Mamm. Species 1971, 1. [Google Scholar] [CrossRef]
- Agren, G.; Zhou, Q.; Zhong, W. Ecology and social behaviour of Mongolian gerbils, Meriones unguiculatus, at Xilinhot, Inner Mongolia, China. Anim. Behav. 1989, 37, 11–27. [Google Scholar] [CrossRef]
- Susić, V.; Masirević, G. Sleep patterns in the Mongolian gerbil, Meriones unguiculatus. Physiol. Behav. 1986, 37, 257–261. [Google Scholar] [CrossRef]
- Fisher, M.F.; Llewellyn, G.C. The Mongolian Gerbil: Natural History, Care, and Maintenance. Source Am. Biol. Teach. 1978, 40, 557–560. [Google Scholar]
- Yoshimura, H. Behavioral characteristics of scent marking behavior in the Mongolian gerbil (Meriones unguiculatus). Exp. Anim. 1981, 30, 107–112. [Google Scholar] [CrossRef]
- Cheal, M. The gerbil: A unique model for research on aging. Exp. Aging Res. 1986, 12, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Loskota, W.J.; Lomax, P.; Rich, S.T. The Gerbil as a Model for the Study of the Epilepsies. Epilepsia 1974, 15, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Nakaya, T.; Hagiwara, K.; Momiyama, N.; Kagawa, Y.; Taniyama, H.; Ishihara, C.; Sata, T.; Kurata, T.; Ikuta, K. High susceptibility of Mongolian gerbil (Meriones unguiculatus) to Borna disease virus. Vaccine 1999, 17, 480–489. [Google Scholar] [CrossRef]
- Anderson, G.W.; Slone, T.W.; Peters, C.J. Vfrology The gerbil, Meriones unguiculatus, a model for Rift Valley fever viral encephalitis. Arch. Virol. 1988, 102, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, T.; Yasuda, Y.; Nonaka, S. The Genetics of Coat Colors in the Mongolian Gerbil (Meriones unguiculatus). Exp. Anim. 1989, 38, 337–341. [Google Scholar] [CrossRef]
- Gray-Allan, P.; Wong, R. Influence of coat color genes on seizure behavior in Mongolian gerbils. Behav. Genet. 1990, 20, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Benedito, E.; Jiménez-Cervantes, C.; Pérez, D.; Cubillana, J.D.; Solano, F.; Jiménez-Cervantes, J.; Meyer Zum Gottesberge, A.M.; Lozano, J.A.; García-Borrón, J.C. Melanin formation in the inner ear is catalyzed by a new tyrosine hydroxylase kinetically and structurally different from tyrosinase. Biochim. Biophys. Acta Gen. Subj. 1997, 1336, 59–72. [Google Scholar] [CrossRef]
- Handler, A.H.; Magalini, S.I.; Pav, D. Oncogenic Studies on the Mongolian Gerbil. Cancer Res. 1966, 26, 844–847. [Google Scholar]
- Meckley, P.E.; Zwicker, G.M. Naturally-Occurring Neoplasms in the Mongolian Gerbil, Meriones Unguiculatus. Lab. Anim. 1979, 13, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Shumaker, R.C.; Paik, S.K.; Houser, W.D. Tumors in Gerbillinae: A literature review and report of a case. Lab. Anim. Sci. 1974, 24, 688–690. [Google Scholar] [PubMed]
- Naito, M.; Aoyama, H.; Fujioka, Y.; Ito, A. Induction of Gliomas in Mongolian Gerbils (Meriones unguiculatus) Following Neonatal Administration of N-Ethyl-N-nitrosourea2. JNCI J. Natl. Cancer Inst. 1985, 75, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Kleihues, P.; Bücheler, J.; Riede, U.N. Selective Induction of Melanomas in Gerbils (Meriones unguiculatus) Following Postnatal Administration of N-Ethyl-N-nitrosourea2. JNCI J. Natl. Cancer Inst. 1978, 61, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Yukawa, M.; Onodera, T.; Suzuki, K.; Yokomizo, Y.; Suzuki, M.; Mochizuki, K. Impairment of macrophage function in Mongolian gerbils. Vet. Immunol. Immunopathol. 1992. [Google Scholar] [CrossRef]
- Karavodin, L.M.; Ash, L.R. Weak graft-versus-host response in the Mongolian gerbil (Meriones unguiculatus). Lab. Anim. Sci. 1977, 27, 1035–1036. [Google Scholar] [PubMed]
- Karavodin, L.M.; Ash, L.R. Weak mixed lymphocyte culture response in the Mongolian gerbil (Meriones unguiculatus). Lab. Anim. Sci. 1977, 27, 195–203. [Google Scholar] [PubMed]
- Samlowski, W.E.; McGregor, J.R.; Bastian, N.R. Identification of Nitric Oxide-Derived EPR Signals in Human Cancers. In Nitric Oxide in Transplant Rejection and Anti-Tumor Defense; Springer: Boston, MA, USA, 1998; pp. 341–352. [Google Scholar]
- Lukiewicz, S.; Elas, M.; Raczek, J.; Bratasz, A.; Pajak, S.; Cieszka, K. EPR signals detectable in human tumors. In Nitric Oxide in Allograft Rejection and Anti-Tumor Responces; Springer: Boston, MA, USA, 1998; pp. 353–370. [Google Scholar]
- Plonka, P.M.; Plonka, B.K.; Pajak, S.; Lukiewicz, S.J. ESR Studies on the Generation of Nitric Oxide in Rejecting Rat Heart Allografts and In Situ Growing Tumors. Curr. Top. Biophys. 1996, 20, 46–52. [Google Scholar]
- Plonka, P.; Plonka, B.; Cieszka, K.; Raczek, J.; Lukiewicz, S. EPR Analysis of Tumor-Host Interactions. In Nitric Oxide in Transplant Rejection and Anti-Tumor Defense; Springer: Boston, MA, USA, 1998; pp. 313–326. [Google Scholar]
- Lancaster, J.R.; Hibbs, J.B. EPR demonstration of iron-nitrosyl complex formation by cytotoxic activated macrophages. Proc. Natl. Acad. Sci. USA 1990, 87, 1223–1227. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.; Plonka, B.; Drzewinska, M.; Pajak, S. Accelerated Rejection of Cardiac Allografts and Xenografts in Presensitized Rats and Gerbils. In Nitric Oxide in Transplant Rejection and Anti-Tumor Defense; Springer: Boston, MA, USA, 1998; pp. 205–211. [Google Scholar]
- Pustelny, K.; Bielanska, J.; Plonka, P.M.; Rosen, G.M.; Elas, M. In vivo spin trapping of nitric oxide from animal tumors. Nitric Oxide 2007, 16, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.M.; Pierce, G.B. Relationship Between Growth Rate and Differentiation of Melanoma In Vivo. JNCI J. Natl. Cancer Inst. 1964, 32, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Grin, C.M.; Kopf, A.W.; Welkovich, B.; Bart, R.S.; Levenstein, M.J. Accuracy in the Clinical Diagnosis of Malignant Melanoma. Arch. Dermatol. 1990, 126, 763. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Hunt, D.M.; Turbyfill, C. High Resistance of Mongolian Gerbils to Irradiation. Nature 1964, 203, 536–537. [Google Scholar] [CrossRef] [PubMed]
- Nelson, J.M. Survival Time Response of the Mongolian Gerbil after Total Body Irradiation. Radiat. Res. 1971, 48, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, A.P.; Sullivan, M.T.; Lutkenhoff, D.W.; Parkinson, W.C.; Overway, D.J. Radioresistance of Mongolian Gerbils. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1977, 31, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, F.; Nakao, N.; Nikaido, O.; Kondo, S. High Resistance of Cultured Mongolian Gerbil Cells to X-Ray-Induced Killing and Chromosome Aberrations. Radiat. Res. 1992, 131, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Brożyna, A.A.; Jóźwicki, W.; Roszkowski, K.; Filipiak, J.; Slominski, A.T. Melanin content in melanoma metastases affects the outcome of radiotherapy. Oncotarget 2016, 7, 17844–17853. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.J.; Mihm, M.C. Melanoma. N. Engl. J. Med. 2006, 355, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.A.; Ross, J.S.; Slominski, A.; Linette, G.; Mysliborski, J.; Hill, J.; Mihm, M. Molecular diagnostics in melanoma. J. Am. Acad. Dermatol. 2005, 52, 743–775. [Google Scholar] [CrossRef] [PubMed]
- Schadendorf, D.; Fisher, D.E.; Garbe, C.; Gershenwald, J.E.; Grob, J.-J.; Halpern, A.; Herlyn, M.; Marchetti, M.A.; McArthur, G.; Ribas, A.; et al. Melanoma. Nat. Rev. Dis. Prim. 2015, 1, 15003. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.A.; Fisher, D.E. The melanoma revolution: From UV carcinogenesis to a new era in therapeutics. Science 2014, 346, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Schadendorf, D.; Hauschild, A. Melanoma in 2013: Melanoma—The run of success continues. Nat. Rev. Clin. Oncol. 2014, 11, 75–76. [Google Scholar] [CrossRef] [PubMed]
- Shah, D.J.; Dronca, R.S. Latest Advances in Chemotherapeutic, Targeted, and Immune Approaches in the Treatment of Metastatic Melanoma. Mayo Clin. Proc. 2014, 89, 504–519. [Google Scholar] [CrossRef] [PubMed]
- Kwong, L.N.; Davies, M.A. Targeted therapy for melanoma: Rational combinatorial approaches. Oncogene 2014, 33, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, S.; Watson, I.R. Molecular characterisation of cutaneous melanoma: Creating a framework for targeted and immune therapies. Br. J. Cancer 2016, 115, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, V.; Stratigos, A.J. Emerging trends in the epidemiology of melanoma. Br. J. Dermatol. 2014, 170, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.; McKay, M.J. Dispelling the myths surrounding radiotherapy for treatment of cutaneous melanoma. Lancet Oncol. 2006, 7, 575–583. [Google Scholar] [CrossRef]
- Ballo, M.T.; Ang, K.K. Radiotherapy for cutaneous malignant melanoma: Rationale and indications. Oncology 2004, 18, 99–107. [Google Scholar] [PubMed]
- Testori, A.; Rutkowski, P.; Marsden, J.; Bastholt, L.; Chiarion-Sileni, V.; Hauschild, A.; Eggermont, A.M.M. Surgery and radiotherapy in the treatment of cutaneous melanoma. Ann. Oncol. 2009, 20, vi22–vi29. [Google Scholar] [CrossRef] [PubMed]
- Foote, M.; Burmeister, B.; Dwyer, P.; Burmeister, E.; Lambie, D.; Allan, C.; Barbour, A.; Bayley, G.; Pullar, A.; Meakin, J.; et al. An innovative approach for locally advanced stage III cutaneous melanoma. Melanoma Res. 2012, 22, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Mendenhall, W.M.; Shaw, C.; Amdur, R.J.; Kirwan, J.; Morris, C.G.; Werning, J.W. Surgery and adjuvant radiotherapy for cutaneous melanoma considered high-risk for local–regional recurrence. Am. J. Otolaryngol. 2013, 34, 320–322. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Camacho, F.; Andrews, G.; Mackley, H. A retrospective analysis of the role of adjuvant radiotherapy in the treatment of cutaneous melanoma. Cancer Biol. Ther. 2016, 17, 1030–1034. [Google Scholar] [CrossRef] [PubMed]
- Oliver, D.E.; Patel, K.R.; Switchenko, J.; Parker, D.; Lawson, D.H.; Delman, K.A.; Kudchadkar, R.R.; Khan, M.K. Roles of adjuvant and salvage radiotherapy for desmoplastic melanoma. Melanoma Res. 2016, 26, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Song, S.Y.; Kim, W.; Jeong, S.-Y.; Choi, W.; Je, H.U.; Lee, J.-S.; Choi, E.K. Feasibility of Postoperative Radiotherapy Using Conventional Fractionation for Lymph Node Metastasis from Cutaneous Melanoma. Anticancer Res. 2017, 37, 4239–4246. [Google Scholar] [CrossRef] [PubMed]
- Rule, W.G.; Allred, J.B.; Pockaj, B.A.; Markovic, S.N.; DiCaudo, D.J.; Erickson, L.A.; Deming, R.L.; Schild, S.E. Results of NCCTG N0275 (Alliance)—A phase II trial evaluating resection followed by adjuvant radiation therapy for patients with desmoplastic melanoma. Cancer Med. 2016, 5, 1890–1896. [Google Scholar] [CrossRef] [PubMed]
- Guadagnolo, B.A.; Prieto, V.; Weber, R.; Ross, M.I.; Zagars, G.K. The role of adjuvant radiotherapy in the local management of desmoplastic melanoma. Cancer 2014, 120, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Hedblad, M.-A.; Mallbris, L. Grenz ray treatment of lentigo maligna and early lentigo maligna melanoma. J. Am. Acad. Dermatol. 2012, 67, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Farshad, A.; Burg, G.; Panizzon, R.; Dummer, R. A retrospective study of 150 patients with lentigo maligna and lentigo maligna melanoma and the efficacy of radiotherapy using Grenz or soft X-rays. Br. J. Dermatol. 2002, 146, 1042–1046. [Google Scholar] [CrossRef] [PubMed]
- Naganawa, K.; Koto, M.; Takagi, R.; Hasegawa, A.; Ikawa, H.; Shimozato, K.; Kamada, T.; Okamoto, Y. Organizing Committee for the Working Group for Head-and-Neck Cancer. Long-term outcomes after carbon-ion radiotherapy for oral mucosal malignant melanoma. J. Radiat. Res. 2017, 58, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yu, Y.; Wang, H.; Yan, A.; Jiang, X. Evaluation of the prognostic impact of postoperative adjuvant radiotherapy on head and neck mucosal melanoma: A meta-analysis. BMC Cancer 2015, 15, 758. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, K.; Wakatsuki, M.; Kato, S.; Kiyohara, H.; Kamada, T. Working Group for Gynecological Tumors. Clinical trial of carbon ion radiotherapy for gynecological melanoma. J. Radiat. Res. 2014, 55, 343–350. [Google Scholar] [CrossRef]
- McKenna, J.K.; Florell, S.R.; Goldman, G.D.; Bowen, G.M. Lentigo Maligna/Lentigo Maligna Melanoma: Current State of Diagnosis and Treatment. Dermatol. Surg. 2006, 32, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Tio, D.; van der Woude, J.; Prinsen, C.A.C.; Jansma, E.P.; Hoekzema, R.; van Montfrans, C. A systematic review on the role of imiquimod in lentigo maligna and lentigo maligna melanoma: Need for standardization of treatment schedule and outcome measures. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Strom, T.; Caudell, J.J.; Han, D.; Zager, J.S.; Yu, D.; Cruse, C.W.; Marzban, S.S.; Messina, J.L.; Trotti, A.M.; Sondak, V.K.; et al. Radiotherapy influences local control in patients with desmoplastic melanoma. Cancer 2014, 120, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Zbytek, B.; Slominski, R. Inhibitors of melanogenesis increase toxicity of cyclophosphamide and lymphocytes against melanoma cells. Int. J. Cancer 2009, 124, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Kinnaert, E.; Morandini, R.; Simon, S.; Hill, H.Z.; Ghanem, G.; Van Houtte, P. The degree of pigmentation modulates the radiosensitivity of human melanoma cells. Radiat. Res. 2000, 154, 497–502. [Google Scholar] [CrossRef]
- Barranco, S.C.; Romsdahl, M.M.; Humphrey, R.M. The Radiation Response of Human Malignant Melanoma Cells Grown in Vitro. Cancer Res. 1971, 31, 830–833. [Google Scholar] [PubMed]
- Slominski, A.; Ermak, G.; Wortsman, J. Modification of melanogenesis in cultured human melanoma cells. In Vitro Cell. Dev. Biol. Anim. 1999, 35, 564–565. [Google Scholar] [CrossRef] [PubMed]
- Brozyna, A.A.; VanMiddlesworth, L.; Slominski, A.T. Inhibition of melanogenesis as a radiation sensitizer for melanoma therapy. Int. J. Cancer 2008, 123, 1448–1456. [Google Scholar] [CrossRef] [PubMed]
- Janjetovic, Z.; Brozyna, A.; Tuckey, R.; Kim, T.-K.; Nguyen, M.; Jozwicki, W.; Pfeffer, S.; Pfeffer, L.; Slominski, A. High basal NF-kB activity in nonpigmented melanoma cells is associated with an enhanced sensitivity to vitamin D3 derivatives. Br. J. Cancer 2011, 105, 1874–1884. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.; Good-Snitkoff, G. Dopa inhibits induced proliferative activity of murine and human lymphocytes. Anticancer Res. 1992, 12, 753–756. [Google Scholar] [PubMed]
- Shields, C.L.; Kaliki, S.; Furuta, M.; Fulco, E.; Alarcon, C.; Shields, J.A. American joint committee on cancer classification of uveal melanoma (anatomic stage) predicts prognosis in 7731 patients: The 2013 zimmerman lecture. Ophthalmology 2015, 122, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Shields, C.L.; Kaliki, S.; Cohen, M.N.; Shields, P.W.; Furuta, M.; Shields, J.A. Prognosis of uveal melanoma based on race in 8100 patients: The 2015 Doyne Lecture. Eye 2015, 29, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Particle Therapy Co-Operative Group (PTCOG) and Particles Newsletter. Available online: http://www.ptcog.com/ (accessed on 17 August 2017).
- Egger, E.; Zografos, L.; Schalenbourg, A.; Beati, D.; Böhringer, T.; Chamot, L.; Goitein, G. Eye retention after proton beam radiotherapy for uveal melanoma. Int. J. Radiat. Oncol. Biol. Phys. 2003, 55, 867–880. [Google Scholar] [CrossRef]
- Gragoudas, E.S.; Goitein, M.; Verhey, L.; Munzenreider, J.; Suit, H.D.; Koehler, A. Proton beam irradiation. An alternative to enucleation for intraocular melanomas. Ophthalmology 1980, 87, 571–581. [Google Scholar] [CrossRef]
- Allen, A.M.; Pawlicki, T.; Dong, L.; Fourkal, E.; Buyyounouski, M.; Cengel, K.; Plastaras, J.; Bucci, M.K.; Yock, T.I.; Bonilla, L.; et al. An evidence based review of proton beam therapy: The report of ASTRO’s emerging technology committee. Radiother. Oncol. 2012, 103, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Gragoudas, E.S.; Goitein, M.; Koehler, A.M.; Verhey, L.; Tepper, J.; Suit, H.D.; Brockhurst, R.; Constable, I.J. Proton irradiation of small choroidal malignant melanomas. Am. J. Ophthalmol. 1977, 83, 665–673. [Google Scholar] [CrossRef]
- DeLaney, T.F.; Kooy, M.H. Proton and Charged Particle Radiotherapy; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2008; ISBN 978-3-642-00874-0. [Google Scholar]
- Sas-Korczynska, B.; Markiewicz, A.; Romanowska-Dixon, B.; Pluta, E. Preliminary results of proton radiotherapy for choroidal melanoma—The Kraków experience. Wspolczesna Onkol. 2014, 18, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Romanowska-Dixon, B.; Pogrzebielski, A.; Bogdali, A.; Markiewicz, A.; Swakoń, J.; Olko, P.; Jezabek, M.; Sas-Korczyńska, B.; Pluta, E. Proton beam radiotherapy of uveal melanoma—Preliminary results. Klin. Oczna 2012, 114, 173–179. [Google Scholar] [PubMed]
- Verma, V.; Mehta, M. Clinical Outcomes of Proton Radiotherapy for Uveal Melanoma. Clin. Oncol. 2016, 28, e17–e27. [Google Scholar] [CrossRef] [PubMed]
- Romanowska-Dixon, B.; Markiewicz, A.; Sas-Korczynska, B.; Medoń, D.; Walasek, T. Evaluation of intraocular proton beam irradiation complications after choroidal melanoma treatment. Klin. Oczna 2016, 4, 289–292. (In Polish) [Google Scholar]
- Kedracka-Krok, S.; Jankowska, U.; Elas, M.; Sowa, U.; Swakon, J.; Cierniak, A.; Olko, P.; Romanowska-Dixon, B.; Urbanska, K. Proteomic analysis of proton beam irradiated human melanoma cells. PLoS ONE 2014, 9, e84621. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, B.; Sun, B.; Wang, X.; Ban, X.; Sun, T.; Liu, Z.; Zhao, X. A novel function for vimentin: The potential biomarker for predicting melanoma hematogenous metastasis. J. Exp. Clin. Cancer Res. 2010, 29, 109. [Google Scholar] [CrossRef] [PubMed]
- Kubicka-Trzaska, A.; Starzycka, M.; Romanowska-Dixon, B.; Morawski, K. Use of indocyanine green for photodynamic therapy of choroidal melanoma—Preliminary report]. Klin. Oczna 2003, 105, 132–135. [Google Scholar] [PubMed]
- Campagnoli, T.R.; Medina, C.A.; Singh, A.D. Choroidal melanoma initially treated as hemangioma: Diagnostic and therapeutic considerations. Retin. Cases Brief Rep. 2016, 10, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Fabian, I.D.; Stacey, A.W.; Papastefanou, V.; Al Harby, L.; Arora, A.K.; Sagoo, M.S.; Cohen, V.M.L. Primary photodynamic therapy with verteporfin for small pigmented posterior pole choroidal melanoma. Eye 2017, 31, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Austin, E.; Mamalis, A.; Ho, D.; Jagdeo, J. Laser and light-based therapy for cutaneous and soft-tissue metastases of malignant melanoma: A systematic review. Arch. Dermatol. Res. 2017, 309, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Żygulska-Mach, H.; Maciejewski, Z.; Lukiewicz, S.; Iwasiów, B.; Link, E. Clinical Trials on Chemical Radiosensitization of Malignant Melanoma of the Choroid. Ophthalmologica 1979, 178, 194–197. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
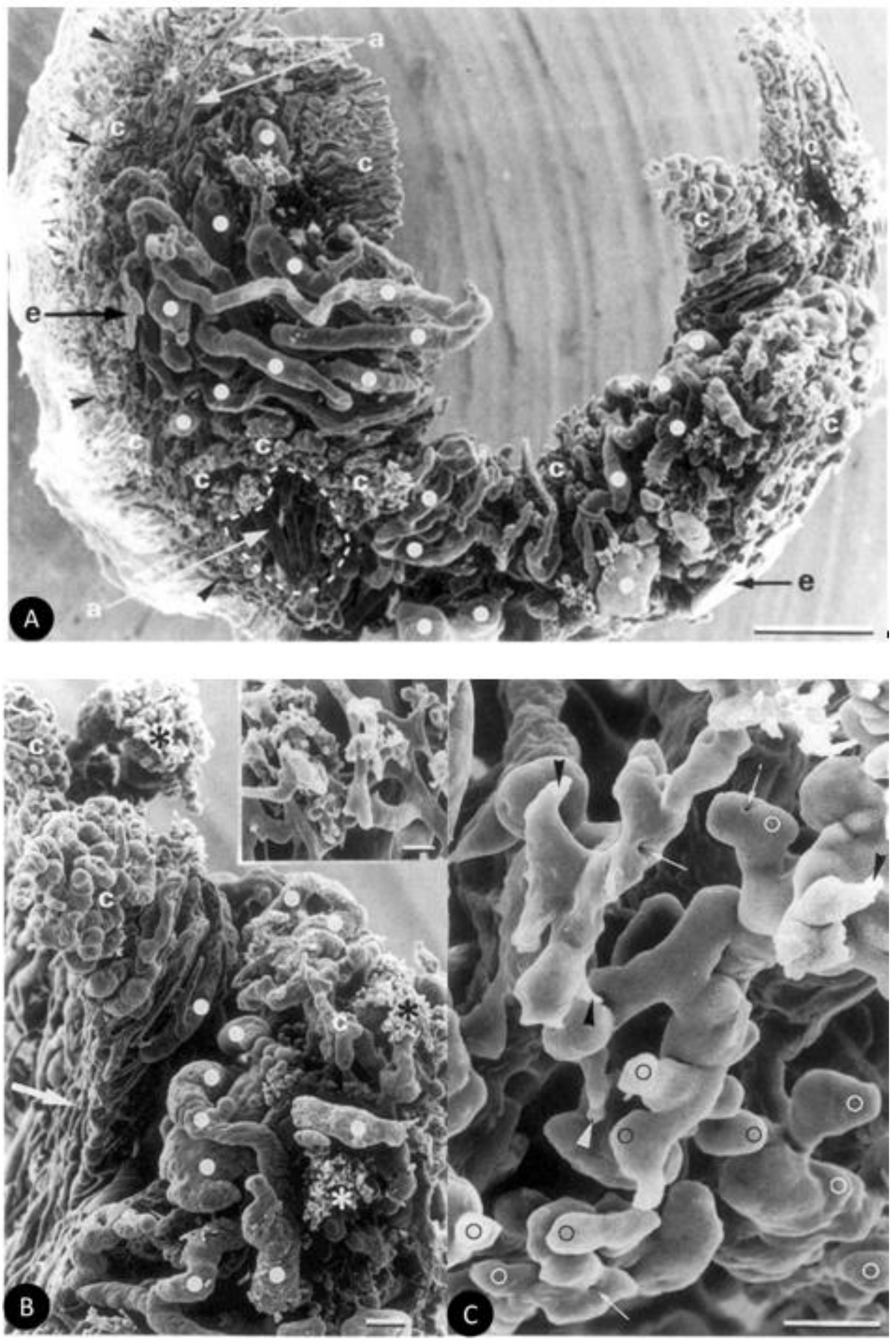{kind=link}
{kind=link}
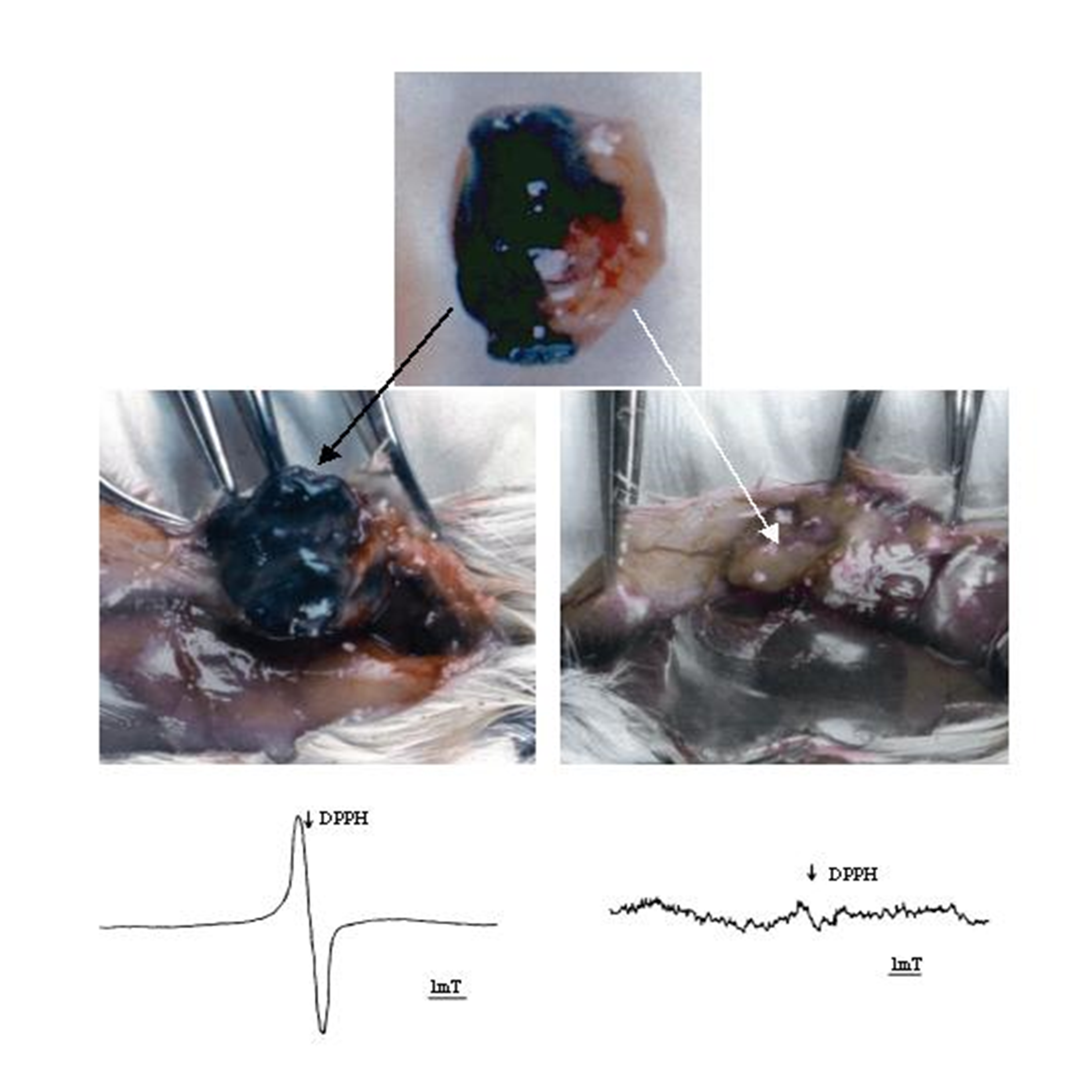{kind=link}
| Melanotic Melanoma Ma | Amelanotic Melanoma Ab | Reference | |
|---|---|---|---|
| Origin | Developed as a spontaneous malignant melanoma of the skin. | Developed as a spontaneous alteration of Ma melanotic melanoma. | [4,5] |
| Year of origin | 1959 | 1963 | [4] |
| Amount of tissue needed for 100% transplantability (mg) | 200 | 50 | [4,5] |
| Transplantation interval (days) | 21 ± 3 | 12 ± 2 | |
| Survival time of implanted hamster (days) | 81 ± 5.8 | 27 ± 1.5 | [5] |
| Most frequent locations of metastases | Lungs, lymph nodes | Kidneys, liver, lymph nodes | [4] |
| Histological and ultrastructural features | [4,110] | ||
| Epitheloidal cells | Polygonal cells | ||
| Melanosomes and premelanosomes | Lack of melanin and melanosomes | ||
| Golgi area is moderately developed | Golgi area is more extensive than in Ma; Products of tyrosinase activity accumulate in the vesicles of the trans-GA | ||
| Moderate amount of ribosomes | Abundant ribosomes | ||
| Mitosis is rare | Mitosis is frequent | ||
| RER and SER are moderately developed | RER and SER are very well developed | ||
| Some mitochondria | Some mitochondria | ||
| Plasma membrane structure: | |||
| 1. Carbohydrates content (nmol/mg of protein) | 1702 | 631 | [111] |
| 2. Heterogeneity | 3 protein fractions 6 glycoprotein fractions | 1 protein fraction 8 glycoprotein fractions | [112] |
| 3. Membrane fluidity and molecular mobility in the plasmatic membrane | Lower degree of order in the phospholipid bilayer; increase in membrane fluidity | [113] | |
| 4. Expression of P glycoprotein (Pgp) | 70% of cells Pgp positive | 10% of cells Pgp positive | [114] |
| 5. Ganglioside content | High level of GM3 Low level of GD3 and 9-O-acetyl-GD3 | Low level of GM3 High level of GD3 and 9-O-acetyl-GD3 | [115,116] |
| 6. Neutral glycolipid content | High level of GL1 | High level of Gb3, Gb4, Gb5 | [117] |
| Antigenicity | Low | Increased in comparison to Ma | [118] |
| Immunogenicity | Low | Increased in comparison to Ma | [119,120] |
| Cytokine secretion | Altered secretion of IL-6, IL-10, TNF-α | [121] | |
| DNA ploidy | 4n | 3n | [5,122] |
| Radiosensitivity | Low | High | [105] |
| Ability for apoptosis | High propensity for spontaneous apoptosis | Low endogenous apoptosis but highly sensitive to camptothecin-induced apoptosis | [103,104] |
| Cell cycle analysis | 30% in S + G2/M phase | 40% in S + G2/M phase | [104] |
| Main biochemical features | |||
| 1. Tyrosinase activity | High tyrosinase activity | Low tyrosinase activity | [123,124] |
| 2. Glycolysis | High aerobic and anaerobic glycolysis | [84] | |
| 3. Antioxidant enzymes | High activity of dismutase/peroxidase | Relatively low activity of dismutases/peroxidase | [125] |
| 4. Mitochondrial transmembrane potential ΔΨ | Relatively low | Relatively high | [109] |
| 5. Oxygen consumption | Relatively high | Relatively low | [106] |
| 6. Enzyme activities | Relatively high activities of citrate synthase, succinate dehydrogenase, malate dehydrogenase higher than in Ab | Relatively high activities of NAD-dependent glycerol-3-phosphate dehydrogenase higher than Ma | [106] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Śniegocka, M.; Podgórska, E.; Płonka, P.M.; Elas, M.; Romanowska-Dixon, B.; Szczygieł, M.; Żmijewski, M.A.; Cichorek, M.; Markiewicz, A.; Brożyna, A.A.; et al. Transplantable Melanomas in Hamsters and Gerbils as Models for Human Melanoma. Sensitization in Melanoma Radiotherapy—From Animal Models to Clinical Trials. Int. J. Mol. Sci. 2018, 19, 1048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041048
Śniegocka M, Podgórska E, Płonka PM, Elas M, Romanowska-Dixon B, Szczygieł M, Żmijewski MA, Cichorek M, Markiewicz A, Brożyna AA, et al. Transplantable Melanomas in Hamsters and Gerbils as Models for Human Melanoma. Sensitization in Melanoma Radiotherapy—From Animal Models to Clinical Trials. International Journal of Molecular Sciences. 2018; 19(4):1048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041048
Chicago/Turabian StyleŚniegocka, Martyna, Ewa Podgórska, Przemysław M. Płonka, Martyna Elas, Bożena Romanowska-Dixon, Małgorzata Szczygieł, Michał A. Żmijewski, Mirosława Cichorek, Anna Markiewicz, Anna A. Brożyna, and et al. 2018. "Transplantable Melanomas in Hamsters and Gerbils as Models for Human Melanoma. Sensitization in Melanoma Radiotherapy—From Animal Models to Clinical Trials" International Journal of Molecular Sciences 19, no. 4: 1048. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041048