A Biocontrol Strain of Bacillus subtilis WXCDD105 Used to Control Tomato Botrytis cinerea and Cladosporium fulvum Cooke and Promote the Growth of Seedlings


Abstract
:
1. Introduction
2. Results
2.1. The Separation, Purification, and Screening of Antagonist Strains
2.2. Identification of Strain WXCDD105
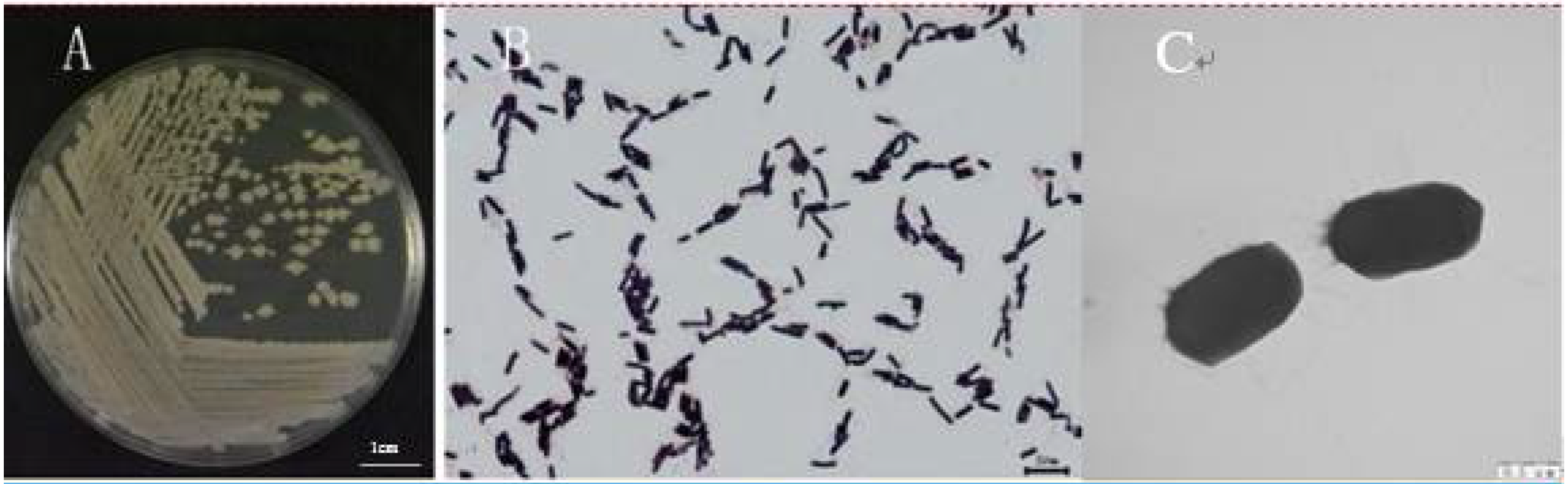
2.2.1. Strain WXCDD105 Morphology Detection
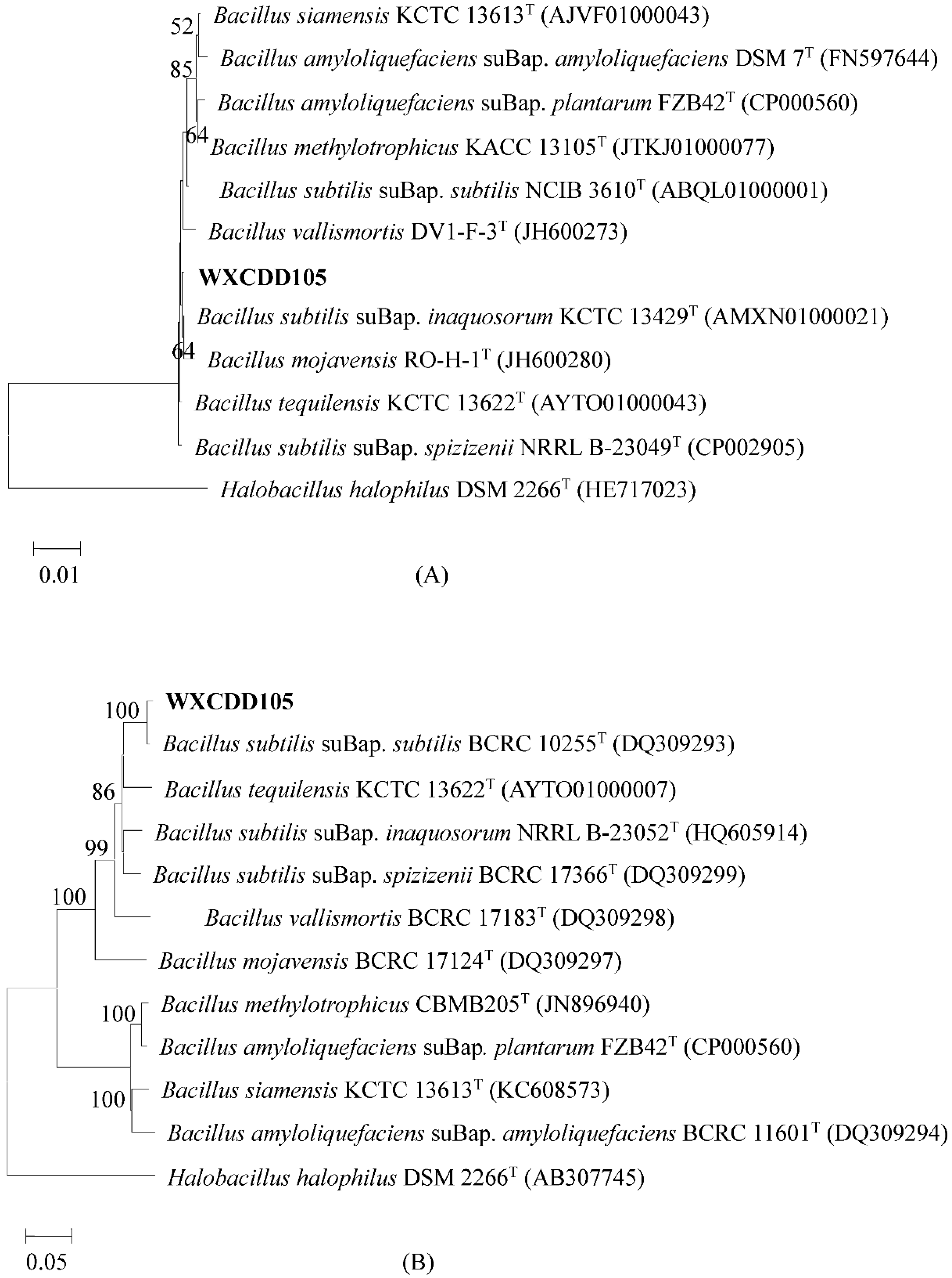
2.2.2. Molecular Identification of Bacterial Strain WXCDD105
2.3. Effect of Strain WXCDD105 Aseptic Filtrate on the Growth of Mycelia of Botrytis cinerea and Fuliva fulva
2.4. The Biocontrol Effect of Strain WXCDD105 on Tomato Gray Mold and Tomato Leaf Mold
2.5. Determination of Bacteriostasis Spectra of Strain WXCDD105
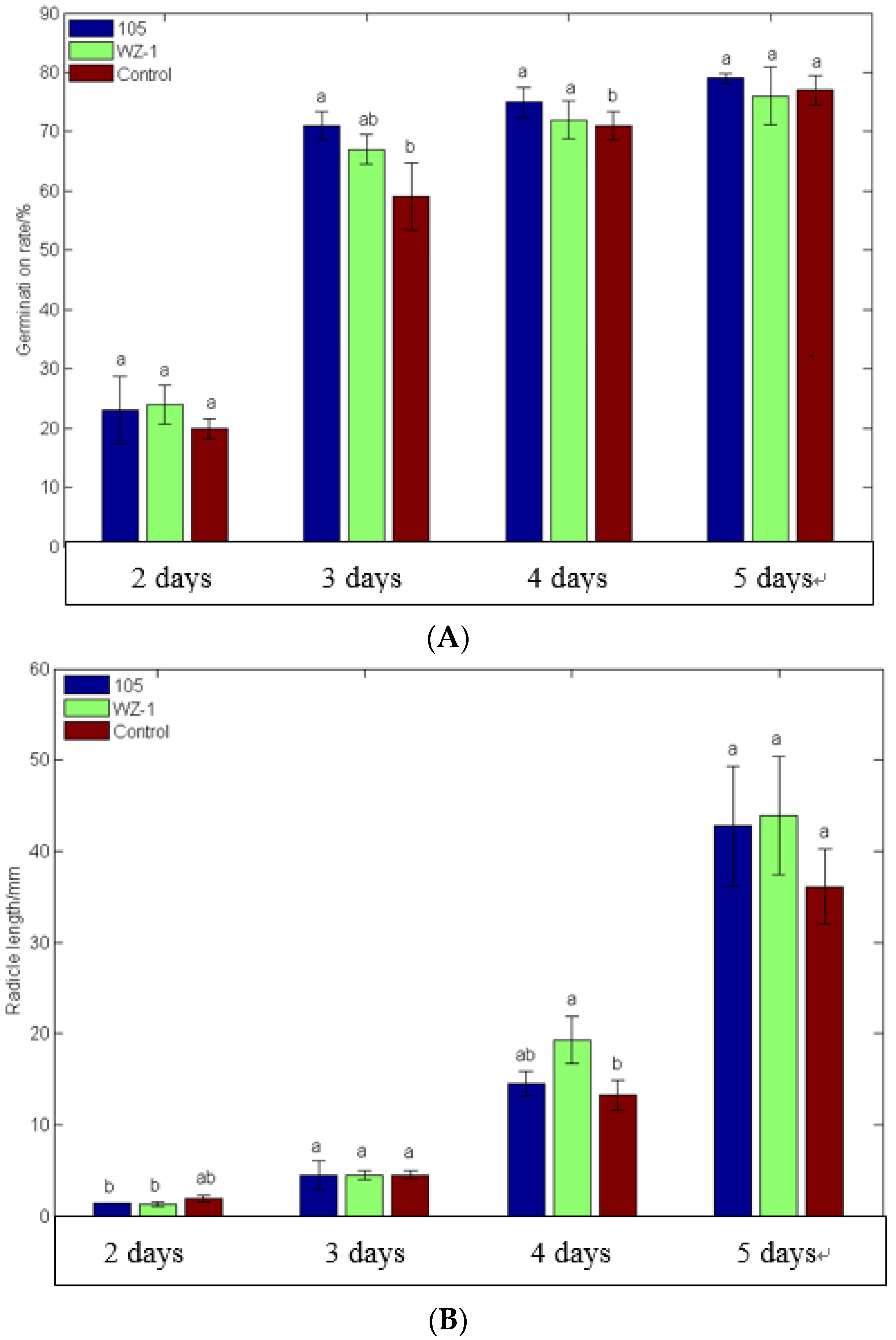
2.6. Growth-Promoting Effects of Strain WXCDD105
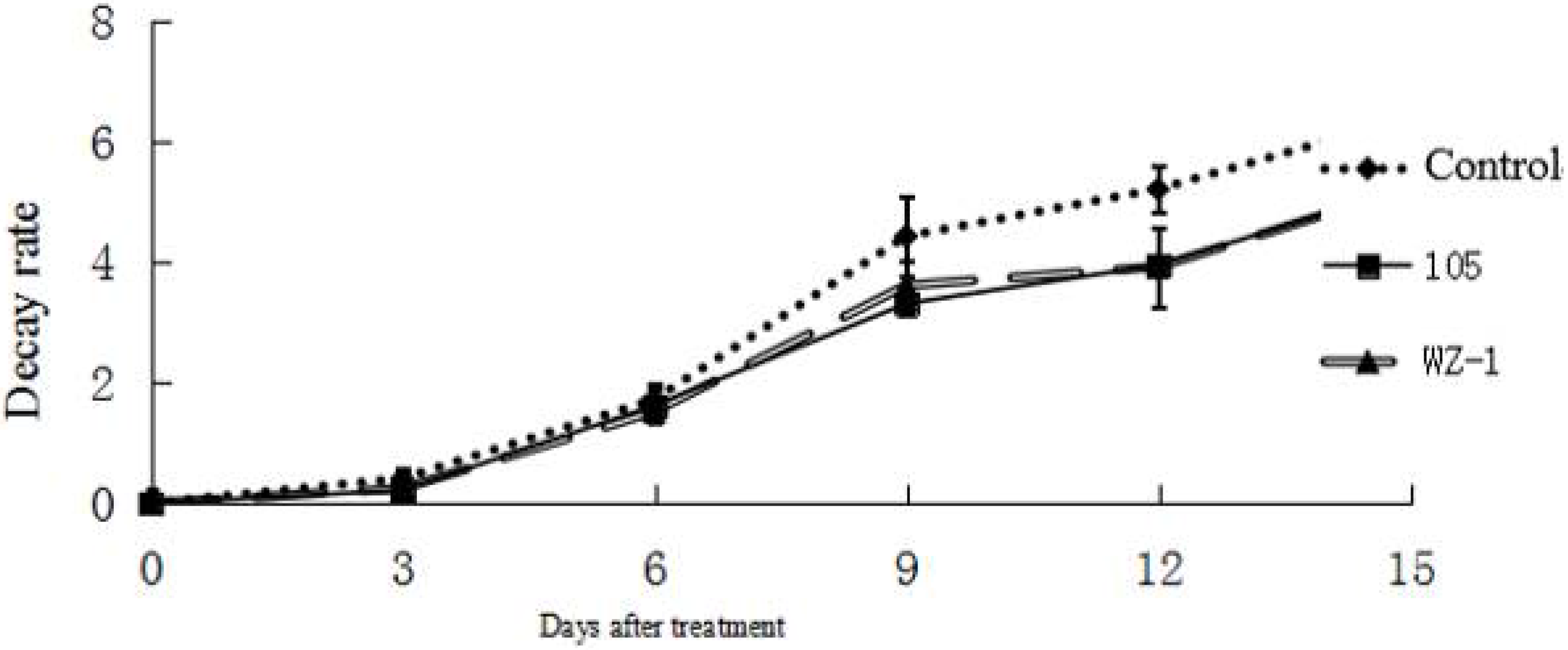
2.7. The Effect of Strain WXCDD105 on the Physiological Quality of Tomato Fruit
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Isolation and Screening of Antagonistic Bacteria
4.3. Morphological and Biochemical Characteristics of Strain WXCDD105
4.4. The Inhibiting Effect of the Biocontrol Bacterium WXCDD105 Fermentation Filtrate on the Growth of Tomato-Grown Mold and Leaf Mold
4.5. Antifungal Activity Assay Using Strain WXCDD105 in the Greenhouse
4.6. Determination of Biocontrol Bacterial Antibacterial Spectrum
4.7. Effects of Biocontrol Bacteria on the Growth of Tomato Seeds and Seedlings
4.8. The Effects of Bacteria Strain WXCDD105 on the Physiological Quality of Tomato Fruits
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| WD1 | Botrytis cinerea |
| WD2 | Cladosporium fulvum Cooke |
| LB | Luria–Bertani |
| PDA | Potato dextrose agar |
| PDB | Potato dextrose broth |
| DI | Disease index |
| BXYL | Beta-XYLOSIDASE |
| LysA | L-Lysine-ARYLAMIDASE |
| AspA | Aspartic-ARYLAMIDASE |
| LeuA | Leucine-ARYLAMIDASE |
| PheA | Phenylalanine ARYLAMIDASE |
| ProA | Proline ARYLAMIDASE |
| BGAL | BETA-GALACTOSIDASE |
| PyrA | Pyrrolydonyl ARYLAMIDASE |
| AGAL | ALPHA-GALACTOSIDASE |
| AlaA | Alanine ARYLAMIDASE |
| TyrA | Tyrosine ARYLAMIDASE |
| BNAG | BETA-N-ACETYL-GLUCOSAMINIDASE |
| APPA | Ala-Phe-Pro-ARYLAMIDASE |
| CDEX | Alanine-phenylalanine-proline ARYLAMIDASE |
| dGAL | D-GALACTOSE |
| GLYG | Glycogen |
| INO | Inositol |
| MdG | Methyl glucoside acidification |
| ELLM | ELLMAN |
| MdX | Methyl-d-xyloside |
| AMAN | Alpha mannosidase |
| MTE | Germination three sugar |
| GlyA | Glycine ARYLAMIDASE |
| dMAN | D-MANNITOL |
| dMNE | D-MANNOSE |
| dMLZ | D-MELEZITOSE |
| NAG | N-ACETYL-GLUCOSAMINE |
| PLE | PALATINOSE |
| IRHA | L-rhamnose monohydrate |
| BGLU | BETA-GLUCOSIDASE |
| INU | Inulin |
| dGLU | D-GLUCOSE |
| dRIB | Ribose |
| PSCNa | Decomposing ammonia assimilation |
| OLD | Old sugar |
| ESC | ESCULIN hydrolyse |
| TTZ | Polymyxin B resistance |
| POLYB_R | Polymyxin B tolerance |
References
- Al-Saleh, M.A. Pathogenic variability among five bacterial isolates of Xanthomonas campestris pv. vesicatoria, causing spot disease on tomato and their response to salicylic acid. J. Saudi Soc. Agric. Sci. 2011, 10, 47–51. [Google Scholar] [CrossRef]
- Babadoost, M. Leaf mold (Fulvia fulva), a serious threat to high tunnel tomato production in illinois. Acta Hortic. 2011, 914, 93–96. [Google Scholar] [CrossRef]
- Williamson, B.; Tudzynski, B.P.; Kan, J.A.L.V. Botrytis cinerea: The cause of grey mould disease. Mol. Plant Pathol. 2007, 8, 561–580. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.J. National Academy of Agricultural Science, RDA, Suwon, Republic of Korea, Study on the Control of Leaf Mold, Powdery Mildew and Gray Mold for Organic Tomato Cultivation. Korean J. Organ. Agric. 2012, 20, 655–668. [Google Scholar] [CrossRef]
- Wahab, S. Biotechnological approaches in the management of plant pests, diseases and weeds for sustainable agriculture. J. Biopestic. 2009, 2, 115–134. [Google Scholar]
- Correa, O.S.; Soria, M.A. Potential of Bacilli for Biocontrol and Its Exploitation in Sustainable Agriculture. Microbiol. Monogr. 2010, 18, 197–209. [Google Scholar]
- Medeiros, F.H.V.D.; Martins, S.J.; Zucchi, T.D.; Melo, I.S.D.; Batista, L.R.; Machado, J.D.C. Controle biológico de fungos de armazenamento produtores de micotoxinas. Ciênc Agrotec 2012, 36, 483–497. [Google Scholar] [CrossRef]
- Smith, S.M. Biological control with Trichogramma: Advances, successes, and potential of their use. Annu. Rev. Entomol. 1996, 41, 375–406. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Feng, H.; Wang, L.; Li, Z.; Shi, Y.; Zhao, L.; Feng, Z.; Zhu, H. Potential of Endophytic Fungi Isolated from Cotton Roots for Biological Control against Verticillium Wilt Disease. PLoS ONE 2017, 12, e0170557. [Google Scholar] [CrossRef] [PubMed]
- Heimpel, G.E.; Ragsdale, D.W.; Venette, R.; Hopper, K.R.; O’Neil, R.J.; Rutledge, C.E.; Wu, Z.S. Prospects for importation biological control of the soybean aphid: Anticipating potential costs and benefits. Ann. Entomol. Soc. Am. 2004, 97, 249–258. [Google Scholar] [CrossRef]
- Raspor, P.; Mikličmilek, D.; Avbelj, M.; Čadež, N. Biocontrol of grey mould disease on grape caused by Botrytis cinerea with autochthonous wine yeasts. Food Technol. Biotechnol. 2010, 48, 336–343. [Google Scholar]
- Elad, Y.; Kohl, J.; Fokkema, N.J. Control of infection and sporulation of Botrytis cinerea in bean and tomato by saprophytic bacteria and fungi. Eur. J. Plant Pathol. 1994, 100, 315–336. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Sonenshein, A.L.; Hoch, J.A.; Losick, R. Bacillus Subtilis and Its Closest Relatives: From Genes to Cells; American Society of Microbiology: Washington, DC, USA, 2002; pp. 263–264. [Google Scholar]
- Dubnau, D.; Losick, R. Bistability in bacteria. Mol. Microbiol. 2010, 61, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Yavuztürk, B.G.; Imer, D.Y.; Park, P.K.; Koyuncu, I. Evaluation of a novel anti-biofouling microorganism (Bacillus sp. T5) for control of membrane biofouling and its effect on bacterial community structure in membrane bioreactors. Water Sci. Technol. 2018, 77, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Davey, M.E.; O’Toole, G.A. Microbial Biofilms: From Ecology to Molecular Genetics. Microbiol. Mol. Biol. Rev. 2000, 64, 847–867. [Google Scholar] [CrossRef] [PubMed]
- Idris, E.S.E.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-Dependent Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth Promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant-Microbe Interact. 2007, 20, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Kakar, K.U.; Duan, Y.P.; Nawaz, Z.; Sun, G.; Almoneafy, A.A.; Hassan, M.A.; Elshakh, A.; Li, B.; Xie, G.L. A novel rhizobacterium Bk7 for biological control of brown sheath rot of rice caused by Pseudomonas fuscovaginae and its mode of action. Eur. J. Plant Pathol. 2014, 138, 819–834. [Google Scholar] [CrossRef]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Kakar, K.U.; Nawaz, Z.; Cui, Z.; Almoneafy, A.A.; Zhu, B.; Xie, G.L. Characterizing the mode of action of Brevibacillus laterosporus B4 for control of bacterial brown strip of rice caused by A. avenae subsp. avenae RS-1. World J. Microbiol. Biotechnol. 2014, 30, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Höfte, M.; Bakker, P.A.H.M. Competition for Iron and Induced Systemic Resistance by Siderophores of Plant Growth Promoting Rhizobacteria; Springer: Berlin/Heidelberg, Germany, 2007; pp. 121–133. [Google Scholar]
- Rashad, Y.M.; Al-Askar, A.A.; Ghoneem, K.M.; Saber, W.I.A.; Hafez, E.E. Chitinolytic Streptomyces griseorubens E44G enhances the biocontrol efficacy against Fusarium wilt disease of tomato. Phytoparasitica 2017, 45, 227–237. [Google Scholar] [CrossRef]
- Abdalla, O.A.; Bibi, S.; Zhang, S. Application of plant growth-promoting rhizobacteria to control Papaya ringspot virus and Tomato chlorotic spot virus. Arch. Phytopathol. Plant Protect. 2017, 54, 584–597. [Google Scholar] [CrossRef]
- Anton, D.; Bender, I.; Kaart, T.; Roasto, M.; Heinonen, M.; Luik, A.; Püssa, T. Changes in Polyphenols Contents and Antioxidant Capacities of Organically and Conventionally Cultivated Tomato (Solanum lycopersicum L.) Fruits during Ripening. Int. J. Anal. Chem. 2017, 2017, 2367453. [Google Scholar] [CrossRef] [PubMed]
- Boukaew, S.; Prasertsan, P.; Troulet, C.; Bardin, M. Biological control of tomato gray mold caused by Botrytis cinerea by using Streptomyces spp. Biocontrol 2017, 62, 793–803. [Google Scholar] [CrossRef]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani Damping-Off of Tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996, 62, 4081–4085. [Google Scholar] [PubMed]
- Shali, A.; Rigi, G.; Pornour, M.; Ahmadian, G. Expression and Secretion of Cyan Fluorescent Protein (CFP) in B. subtilis using the Chitinase Promoter from Bacillus pumilus SG2. Iran. Biomed. J. 2017, 21, 240. [Google Scholar] [CrossRef] [PubMed]
- Al-Masri, M.I. Biological Control of Gray Mold Disease (Botrytis cinerea) on Tomato and Bean Plants by Using Local Isolates of Trichoderma harzianum. Dirasat Agric. Sci. 2005, 32, 145–156. [Google Scholar]
- Higgins, D.; Thompson, J.; Gibson, T.W. Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406. [Google Scholar] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Boshart, M.; Weber, F.; Jahn, G.; Dorsch-Häsler, K.; Fleckenstein, B.; Schaffner, W. A very strong enhancer is located upstream of an immediate early gene of human cytomegalovirus. Cell 1985, 41, 521. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Botrytis cinereal WD1 | Cladosporium fulvum Cooke WD2 | ||
|---|---|---|---|---|
| Inhibition Zone/mm | Inhibition Rate/% | Inhibition Zone/mm | Inhibition Rate/% | |
| 105 | 4.99 ± 0.56 a | 71.57 a | 5.01 ± 0.75 a | 71.64 a |
| 51 | 4.00 ± 0.63 a | 69.13 a | 4.00 ± 0.51 a | 69.13 a |
| 98 | 0.93 ± 0.12 c | 61.56 c | 0.92 ± 0.14 c | 61.53 c |
| 15 | 1.61 ± 0.41 b | 63.23 b | 1.62 ± 0.32 b | 63.26 b |
| 109 | 1.00 ± 0.11 a | 61.74 a | 1.00 ± 0.11 a | 61.74 a |
| The Bacterial Strain WXCDD105 | |||||||
|---|---|---|---|---|---|---|---|
| Identification Index | Result | Identification Index | Result | Identification Index | Result | Identification Index | Result |
| BXYL | + | LysA | − | AspA | − | LeuA | + |
| PheA | − | ProA | − | BGAL | + | PyrA | + |
| AGAL | + | AlaA | − | TyrA | − | BNAG | − |
| APPA | + | CDEX | − | dGAL | − | GLYG | − |
| INO | + | MdG | + | ELLM | − | MdX | − |
| AMAN | − | MTE | + | GlyA | + | dMAN | + |
| dMNE | + | dMLZ | − | NAG | − | PLE | + |
| lRHA | − | BGLU | + | BMAN | − | PHC | − |
| PVATE | + | AGLU | − | dTAG | − | dTRE | + |
| INU | + | dGLU | + | dRIB | + | PSCNa | − |
| NaCl 6.5% | + | KAN | − | OLD | − | ESC | + |
| TTZ | + | POLYB_R | − | ||||
| Treatment | Botrytis cinerea WD1 | Cladosporium fulvum Cooke WD2 | ||
|---|---|---|---|---|
| Mycelium Dry Weight/g | Inhibition Rate/% | Mycelium Dry Weight/g | Inhibition Rate/% | |
| WXCDD105 | 0.024 ± 0.009 | 95.28 | 0.029 ± 0.006 | 94.44 |
| Control | 0.508 ± 0.014 | - | 0.522 ± 0.025 | - |
| Treatment | Tomato Gray Mold Disease | Tomato Leaf Mold Disease | ||
|---|---|---|---|---|
| Disease Index | Control Efficiency/% | Disease Index | Control Efficiency/% | |
| WXCDD105 | 10.73 ± 2. 41 b | 74.70 | 12.09 ± 2.89 b | 72.07 |
| 40% Pyrimethanil | 13.67 ± 2.63 b | 67.77 | - | - |
| 10% Polyoxin | - | - | 12.95 ± 2.74 b | 70.09 |
| Control | 42.41 ± 1.58 a | - | 43.29 ± 1.53 a | - |
| Pathogenic Fungi | Colony Diameter (mm) | Inhibition Rate/% | |
|---|---|---|---|
| Control | Treatment | ||
| Corynespora cassiicola | 88.13 ± 0.78 | 11.49 ± 0.25 | 94.47 ab |
| Botrytis cinerea | 88.56 ± 0.99 | 9.13 ± 0.42 | 97.37 a |
| Sclerotiniasclerotiorum | 88.51 ± 0.55 | 16.79 ± 0.25 | 87.99 c |
| Setosphaeria turcica | 88.36 ± 2.70 | 13.24 ± 1.00 | 92.33 bc |
| Fusarium oxysporum f. sp. cucumerinum | 88.83 ± 0.38 | 16.17 ± 1.72 | 88.79 c |
| Fusarium oxysporum f. sp. niveum | 88.33 ± 3.85 | 25.34 ± 5.68 | 77.45 d |
| Septoria lycopersici Speg. | 88.15 ± 0.45 | 14.86 ± 0.49 | 90.31 bc |
| Fusarium oxysporium Schelcht | 88.11 ± 1.57 | 23.98 ± 3.18 | 79.07 d |
| Fusarium oxysporum f. sp. lycopersici | 88.89 ± 2.15 | 16.04 ± 0.02 | 88.96 c |
| Fusarium oxysporum f. sp. melonis | 88.50 ± 1.57 | 22.41 ± 0.83 | 81.08 d |
| Processing Method | Plant Height/cm | Stem Diameter/mm | Main Root Length/cm | Whole Plant Fresh Weight/g |
|---|---|---|---|---|
| 10 times dilution | 15.30 ± 0.72 b | 3.21 ± 0.04 b | 13.13 ± 1.78 b | 3.52 ± 0.91 b |
| 100 times dilution | 17.30 ± 1.31 a | 3.90 ± 0.14 a | 17.90 ± 1.84 a | 5.64 ± 0.14 a |
| Control | 10.40 ± 0.85 c | 2.75 ± 0.09 c | 8.43 ± 1.81 c | 2.30 ± 0.84 b |
| Treatment | Rotting Rate/% | Firmness/kg·cm−2 | Soluble Solids Content/% | Titratable Acid Content/% |
|---|---|---|---|---|
| Control | 56.67 ± 1.21 a | 0.57 ± 0.06 b | 6.24 ± 0.21 a | 0.32 ± 0.13 a |
| WZ-1 | 38.34 ± 1.13 b | 0.71 ± 0.02 a | 6.25 ± 0.13 a | 0.33 ± 0.04 a |
| WXCDD105 | 28.15 ± 1.02 c | 0.71 ± 0.03 a | 6.27 ± 0.22 a | 0.35 ± 0.06 a |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Shi, Y.; Wang, D.; Yao, Z.; Wang, Y.; Liu, J.; Zhang, S.; Wang, A. A Biocontrol Strain of Bacillus subtilis WXCDD105 Used to Control Tomato Botrytis cinerea and Cladosporium fulvum Cooke and Promote the Growth of Seedlings. Int. J. Mol. Sci. 2018, 19, 1371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051371
Wang H, Shi Y, Wang D, Yao Z, Wang Y, Liu J, Zhang S, Wang A. A Biocontrol Strain of Bacillus subtilis WXCDD105 Used to Control Tomato Botrytis cinerea and Cladosporium fulvum Cooke and Promote the Growth of Seedlings. International Journal of Molecular Sciences. 2018; 19(5):1371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051371
Chicago/Turabian StyleWang, Hui, Yuying Shi, Doudou Wang, Zhongtong Yao, Yimei Wang, Jiayin Liu, Shumei Zhang, and Aoxue Wang. 2018. "A Biocontrol Strain of Bacillus subtilis WXCDD105 Used to Control Tomato Botrytis cinerea and Cladosporium fulvum Cooke and Promote the Growth of Seedlings" International Journal of Molecular Sciences 19, no. 5: 1371. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051371