Overexpression of BraLTP2, a Lipid Transfer Protein of Brassica napus, Results in Increased Trichome Density and Altered Concentration of Secondary Metabolites

Abstract
:1. Introduction
2. Results
2.1. Gene and Protein Sequence Characterization
2.2. Spatiotemporal Expression of BraLTP2
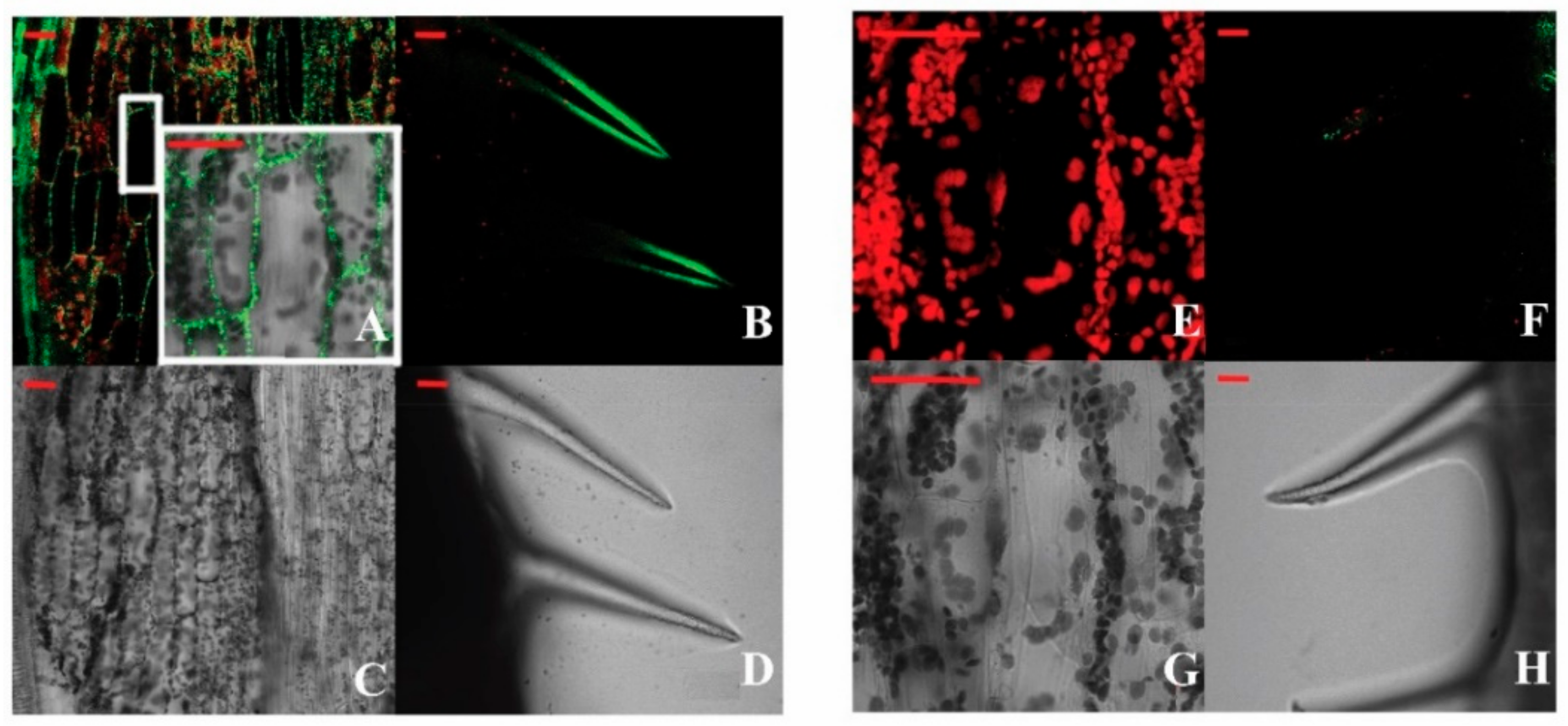
2.3. Localization of BraLTP2::GFP (Green Fluorescent Protein) Fusion Protein
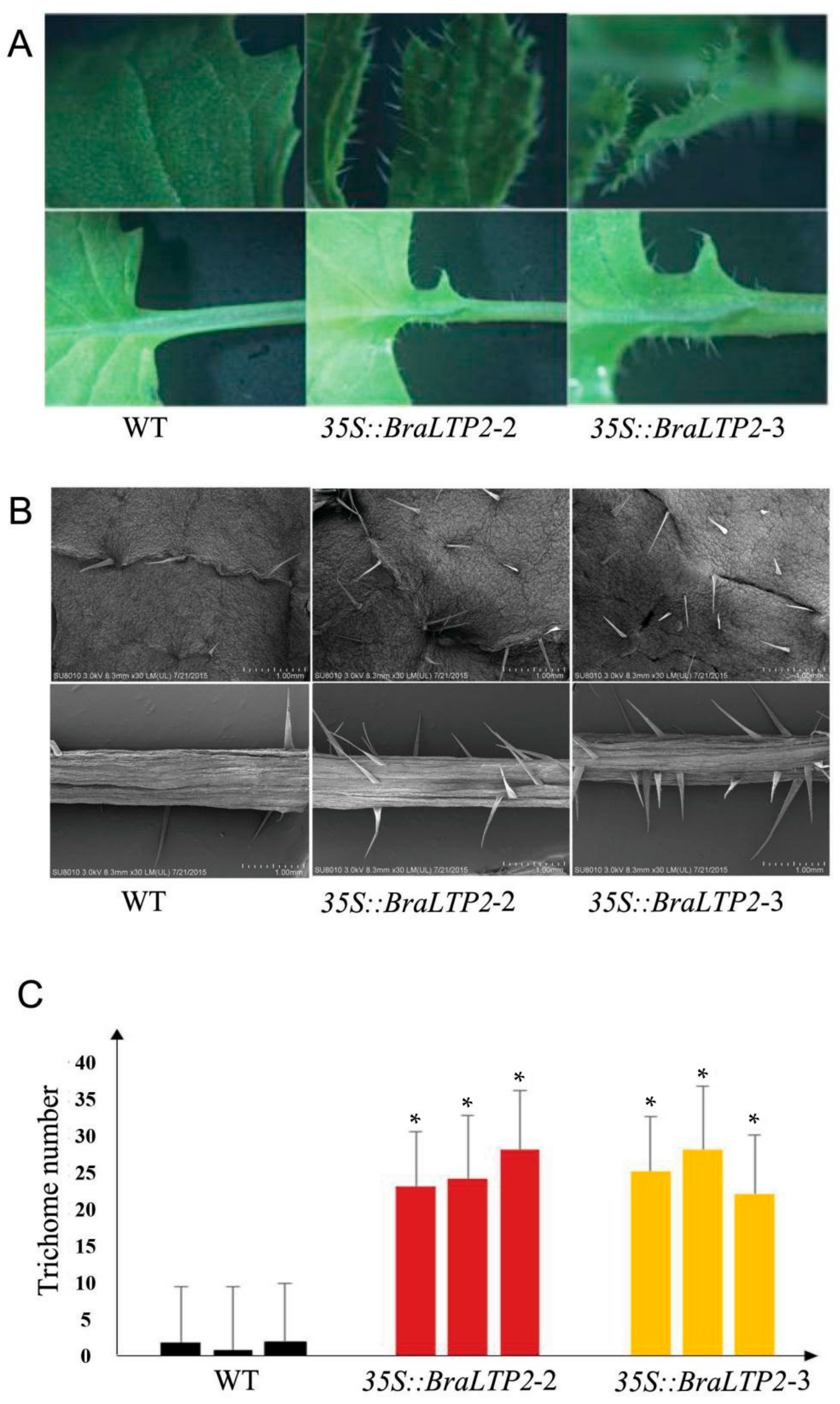
2.4. Overexpressing BraLTP2 Increases Trichome Number
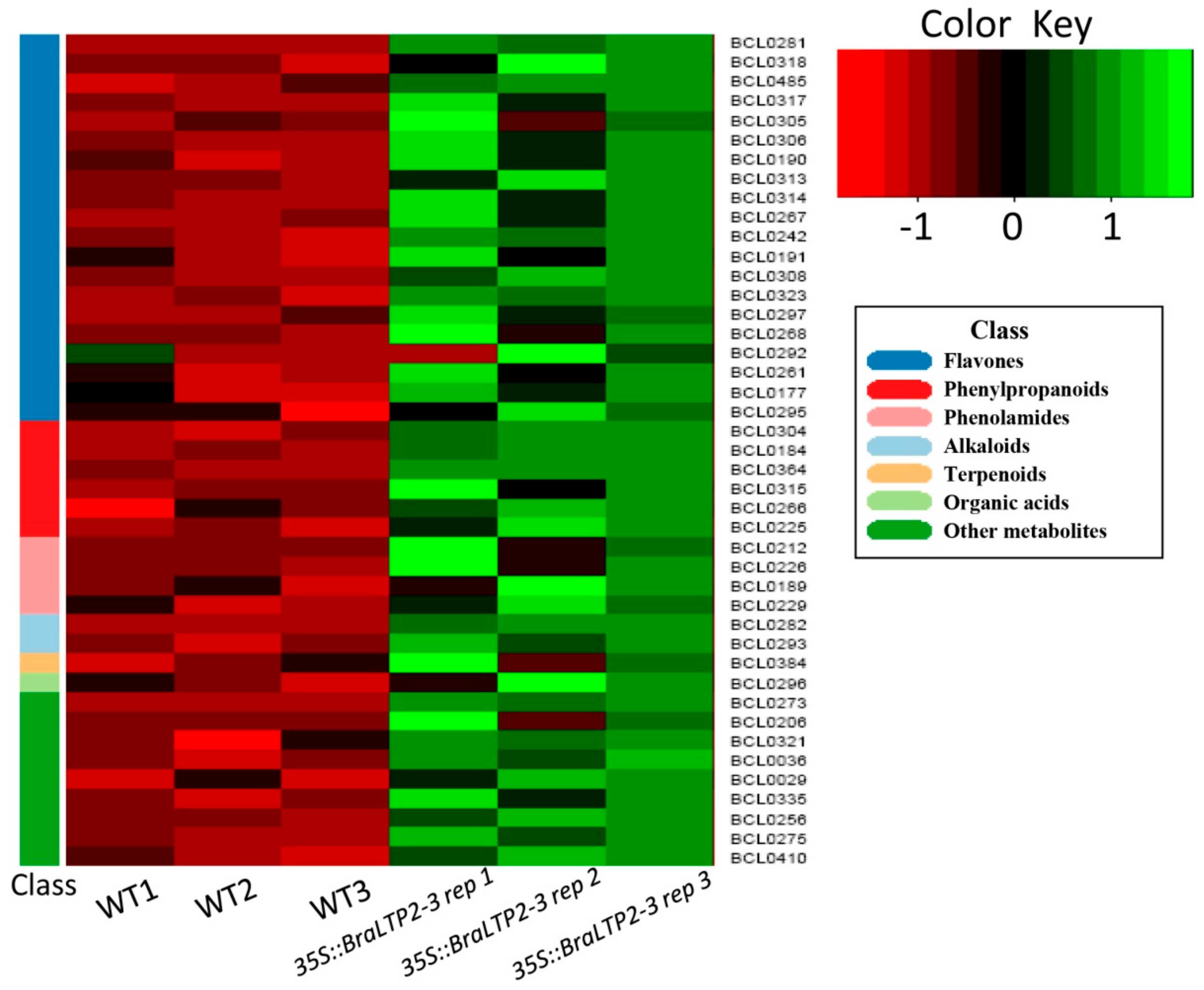
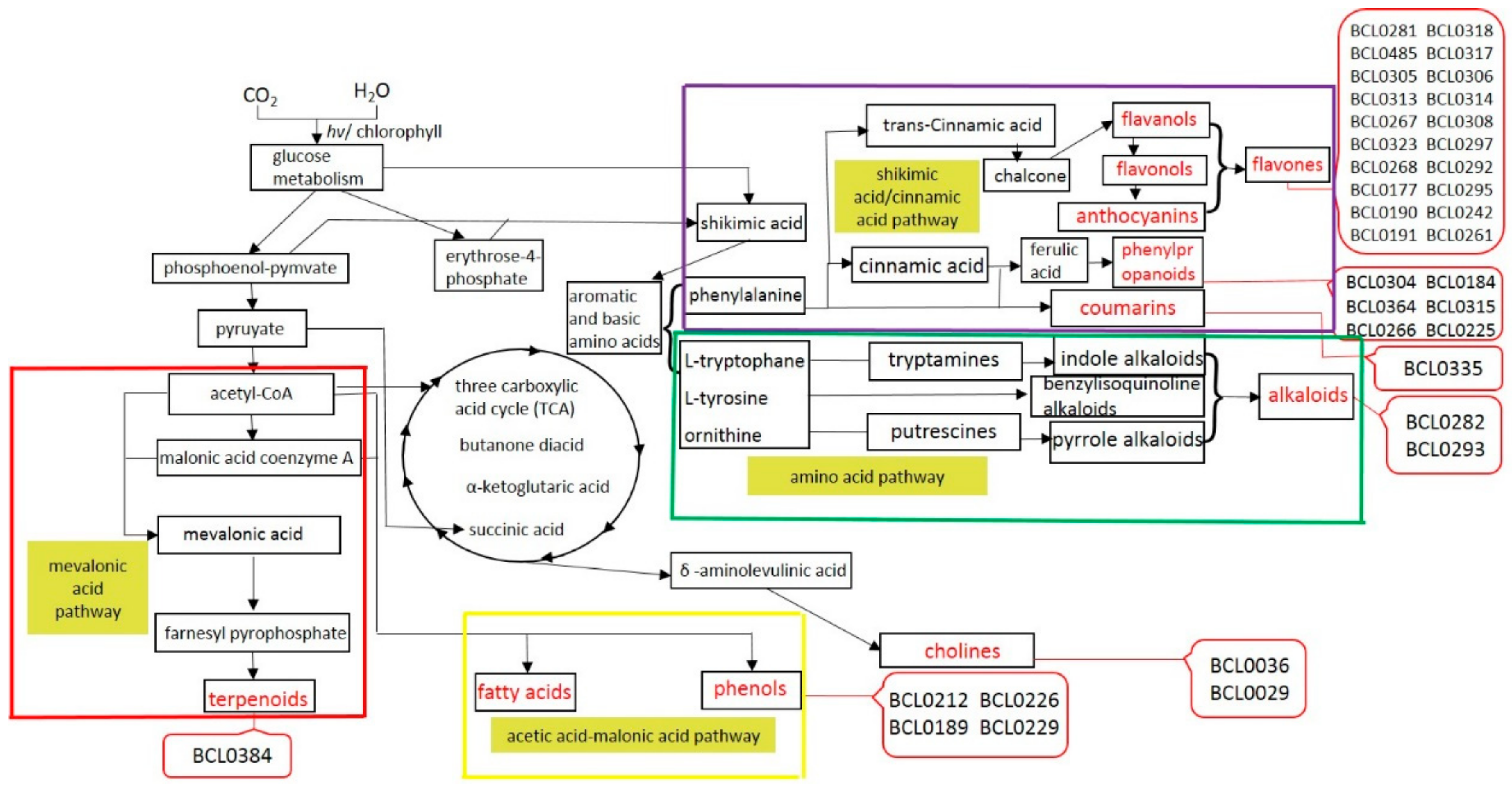
2.5. Overexpressing BraLTP2 Affects Secondary Metabolites in B. napus
2.6. Activities of Antioxidant Enzymes
3. Discussion
3.1. Functional Characterization of BraLTP2
3.2. BraLTP2 Is Expressed in Leaf Epidermal Trichomes.
3.3. Overexpressing BraLTP2 Leads to an Increase of Trichome Number
3.4. Overexpressing BraLTP2 Alters Different Secondary Metabolites Produced by Different Biosynthesis Pathways.
3.5. Overexpressing BraLTP2 Demonstrates Enhanced Activities of the Main Antioxidant Enzymes
4. Materials and Methods
4.1. Plant Material
4.2. Gene, Protein, and Promoter Sequence Analysis
4.3. Vector Construction and Genetic Transformation
4.3.1. Gene Cloning and Vector Construction
4.3.2. Genetic Transformation
4.4. Real-Time PCR Analysis
4.5. Histochemical Analysis of GUS Expression
4.6. Microscopic Observation of the BraLTP2::GFP Fusion Protein
4.7. Trichome Observation
4.8. Analysis of Secondary Metabolites from Transgenic Plants by Liquid Chromatography Tandem Mass Spectrometry (LC-MS)
4.9. Determination of Anti-Oxidant Enzymes Activities
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgements
Conflicts of Interest
References
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar]
- Mathur, J.; Chua, N.H. Microtubule stabilization leads to growth reorientation in Arabidopsis trichomes. Plant Cell 2000, 12, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, J.A.; Agrawal, A.A. The Role of Plant Trichomes and Caterpillar Group Size on Growth and Defence of the Pipevine Swallowtail Battus philenor. J. Anim. Ecol. 2001, 70, 997–1005. [Google Scholar] [CrossRef]
- Voirin, B.; Bayet, C.; Colson, M. Demonstration that flavone aglycones accumulate in the peltate glands of Mentha x piperita leaves. Phytochemistry 1993, 34, 85–87. [Google Scholar] [CrossRef]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as antioxidants in plants: Location and functional significance. Plant Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Gang, D.R.; Wang, J.; Dudareva, N.; Nam, K.H.; Simon, J.E.; Lewinsohn, E.; Pichersky, E. An investigation of the storage and biosynthesis of phenylpropenes in sweet basil. Plant Physiol. 2001, 125, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Hallahan, D.L.; Callow, J.A.; Gray, J.C. Plant Trichomes. In Advances in Botanical Research; Academic Press: London, UK, 2000; Volume 31, pp. 1–35. [Google Scholar]
- Valletta, A.; Santamaria, A. R.; Canini, A.; Canuti, L.; Pasqua, G. Trichomes in Camptotheca acuminata Decaisne (Nyssaceae): Morphology, distribution, structure, and secretion. Plant Biosyst. 2013, 147, 548–556. [Google Scholar] [CrossRef]
- Shepherd, R.W.; Bass, W.T.; Houtz, R.L.; Wagner, G.J. Phylloplanins of tobacco are defensive proteins deployed on aerial surfaces by short glandular trichomes. Plant Cell 2005, 17, 1851–1861. [Google Scholar] [CrossRef] [PubMed]
- Mellon, J.E.; Zelaya, C.A.; Dowd, M.K.; Beltz, S.B.; Klich, M.A. Inhibitory effects of gossypol, gossypolone, and apogossypolone on a collection of economically important filamentous fungi. J. Agric. Food Chem. 2012, 60, 2740–2745. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E.; Duke, S.O. Trichomes and Root Hairs: Natural Pesticide Factories. Pestic. Outlook 2003, 14, 175–178. [Google Scholar] [CrossRef]
- Impei, S.; Gismondi, A.; Canini, A. Metabolic and biological profile of autochthonous Vitis vinifera L. ecotypes. Food Funct. 2015, 6, 1526–1538. [Google Scholar] [CrossRef] [PubMed]
- Gismond, A.; di Marco, G.; Canuti, L.; Canini, A. Antiradical activity of phenolic metabolites extracted from grapes of white and red Vitis vinifera L. cultivars. Vitis 2017, 56, 19–26. [Google Scholar]
- Karabourniotis, G.; Kotsabassidis, D.; Manetas, Y. Trichome density and its protective potential against ultraviolet-B radiation damage during leaf development. Can. J. Bot. 1995, 73, 376–383. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Kofidis, G.; Fasseas, C.; Liakoura, V.; Drossopoulos, I. Polyphenol deposition in leaf hairs of Olea europaea (Oleaceae) and Quercus ilex (Fagaceae). Am. J. Bot. 1998, 85, 1007. [Google Scholar] [CrossRef] [PubMed]
- Lange, B.M.; Wildung, M.R.; Stauber, E.J.; Sanchez, C.; Pouchnik, D.; Croteau, R. Probing essential oil biosynthesis and secretion by functional evaluation of expressed sequence tags from mint glandular trichomes. Proc. Natl. Acad. Sci. USA 2000, 97, 2934–2939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gang, D.R.; Beuerle, T.; Werck-Reichhart, D.; Pichersky, E. Differential production of meta hydroxylated phenylpropanoids in sweet basil peltate glandular trichomes and leaves is controlled by the activities of specific acyltransferases and hydroxylases. Plant Physiol. 2002, 130, 1536–1544. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Paiva, N.L.; May, G.D.; Dixon, R.A. Transcriptome analysis of alfalfa glandular trichomes. Planta 2005, 221, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Bertea, C.M.; Voster, A.; Verstappen, F.W.; Maffei, M.; Beekwilder, J.; Bouwmeester, H.J. Isoprenoid biosynthesis in Artemisia annua: Cloning and heterologous expression of a germacrene A synthase from a glandular trichome cDNA library. Arch. Biochem. Biophys. 2006, 448, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Tian, L.; Aziz, N.; Broun, P.; Dai, X.; He, J.; King, A.; Zhao, P.X.; Dixon, R.A. Terpene biosynthesis in glandular trichomes of hop. Plant Physiol. 2008, 148, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Chatzopoulou, F.M.; Makris, A.M.; Argiriou, A.; Degenhardt, J.; Kanellis, A.K. EST analysis and annotation of transcripts derived from a trichome-specific cDNA library from Salvia fruticosa. Plant Cell Rep. 2010, 29, 523–534. [Google Scholar] [CrossRef] [PubMed]
- Harada, E.; Kim, J.A.; Meyer, A.J.; Hell, R.; Clemens, S.; Choi, Y.E. Expression profiling of tobacco leaf trichomes identifies genes for biotic and abiotic stresses. Plant Cell Physiol. 2010, 51, 1627–1637. [Google Scholar] [CrossRef] [PubMed]
- Schilmiller, A.L.; Miner, D.P.; Larson, M.; McDowell, E.; Gang, D.R.; Wilkerson, C.; Last, R.L. Studies of a biochemical factory: Tomato trichome deep expressed sequence tag sequencing and proteomics. Plant Physiol. 2010, 153, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Debono, A.; Yeats, T.H.; Rose, J.K.; Bird, D.; Jetter, R.; Kunst, L.; Samuels, L. Arabidopsis LTPG is a glycosylphosphatidylinositol-anchored lipid transfer protein required for export of lipids to the plant surface. Plant Cell 2009, 21, 1230–1238. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.B.; Go, Y.S.; Bae, H.J.; Park, J.H.; Cho, S.H.; Cho, H.J.; Lee, D.S.; Park, O.K.; Hwang, I.; Suh, M.C. Disruption of glycosylphosphatidylinositol-anchored lipid transfer protein gene altered cuticular lipid composition, increased plastoglobules, and enhanced susceptibility to infection by the fungal pathogen Alternaria brassicicola. Plant Physiol. 2009, 150, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.E.; Lim, S.; Kim, H.J.; Han, J.Y.; Lee, M.H.; Yang, Y.; Kim, J.A.; Kim, Y.S. Tobacco NtLTP1, a glandular-specific lipid transfer protein, is required for lipid secretion from glandular trichomes. Plant J. 2012, 70, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Thoma, S.; Kaneko, Y.; Somerville, C. A non-specific lipid transfer protein from Arabidopsis is a cell wall protein. Plant J. 1993, 3, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kader, J.C. Lipid-Transfer Proteins in Plants. Annu. Rev. Plant Physiol. Mol. Biol. 1996, 47, 627–654. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Chantret, N.; Gautier, M.F. Genome-wide analysis of the rice and Arabidopsis non-specific lipid transfer protein (nsLtp) gene families and identification of wheat nsLtp genes by EST data mining. BMC Genomics 2008, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Kader, J.C. Proteins and the intracellular exchange of lipids. I. Stimulation of phospholipid exchange between mitochondria and microsomal fractions by proteins isolated from potato tuber. Biochim. Biophys. Acta 1975, 380, 31–44. [Google Scholar] [CrossRef]
- Wang, L.; Yu, S.; Tong, C.; Zhao, Y.; Liu, Y.; Song, C.; Zhang, Y.; Zhang, X.; Wang, Y.; Hua, W.; et al. Genome sequencing of the high oil crop sesame provides insight into oil biosynthesis. Genome Biol. 2014, 15, R39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, G.; Xu, K.; Chen, B.; Yan, G.; Li, F.; Qiao, J.; Zhang, T.; Wu, X. Genome-wide survey and expression analysis of the putative non-specific lipid transfer proteins in Brassica rapa L. PLoS ONE 2014, 9, e84556. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Zhong, X. Non-specific lipid transfer proteins in maize. BMC Plant Biol. 2014, 14, 281. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.; Kim, D.; Bu, S.; Kim, J.; Lee, S.; Kim, J.; Johnson, J.; Seo, Y. Isolation and characterization of lipid transfer protein (LTP) genes from a wheat-rye translocation line. Plant Cell Rep. 2002, 20, 961–966. [Google Scholar]
- Jang, C.S.; Lee, H.J.; Chang, S.J.; Seo, Y.W. Expression and promoter analysis of the TaLTP1 gene induced by drought and salt stress in wheat (Triticum aestivum L.). Plant Sci. 2004, 167, 995–1001. [Google Scholar] [CrossRef]
- Jung, H.W.; Kim, K.D.; Hwang, B.K. Identification of pathogen-responsive regions in the promoter of a pepper lipid transfer protein gene (CALTPI) and the enhanced resistance of the CALTPI transgenic Arabidopsis against pathogen and environmental stresses. Planta 2005, 221, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Saupe, S.; Klinkenberg, J.; Riedel, M. The nonspecific lipid transfer protein AtLtpI-4 is involved in suberin formation of Arabidopsis thaliana crown galls. Plant Physiol. 2016, 172, 1911–1927. [Google Scholar] [CrossRef] [PubMed]
- Sarowar, S.; Kim, Y.J.; Kim, K.D.; Hwang, B.K.; Ok, S.H.; Shin, J.S. Overexpression of lipid transfer protein (LTP) genes enhances resistance to plant pathogens and LTP functions in long-distance systemic signaling in tobacco. Plant Cell Rep. 2009, 28, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Gou, J.; Sun, Y.; Yuan, L.; Tang, Q.; Yang, X.; Pei, Y.; Luo, K. Enhanced resistance to fungal pathogens in transgenic Populus tomentosa Carr. by overexpression of an nsLTP-like antimicrobial protein gene from motherwort (Leonurus japonicus). Tree Physiol. 2010, 30, 1599–1605. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.; Kieslich, C.A.; Morikis, D.; Kim, S.C.; Lord, E.M. A gain-of-function mutation of Arabidopsis lipid transfer protein 5 disturbs pollen tube tip growth and fertilization. Plant Cell 2009, 21, 3902–3914. [Google Scholar] [CrossRef] [PubMed]
- Chae, K.; Gonong, B.J.; Kim, S.C.; Kieslich, C.A.; Morikis, D.; Balasubramanian, S.; Lord, E.M. A multifaceted study of stigma/style cysteine-rich adhesin (SCA)-like Arabidopsis lipid transfer proteins (LTPs) suggests diversified roles for these LTPs in plant growth and reproduction. J. Exp. Bot. 2010, 61, 4277–4290. [Google Scholar] [CrossRef] [PubMed]
- Ambrose, C.; DeBono, A.; Wasteneys, G. Cell geometry guides the dynamic targeting of apoplastic GPI-linked lipid transfer protein to cell wall elements and cell borders in Arabidopsis thaliana. PLoS ONE 2013, 8, e81215. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.; Kim, Y.S.; Park, S.; Back, K. Senescence-induced serotonin biosynthesis and its role in delaying senescence in rice leaves. Plant Physiol. 2009, 150, 1380–1393. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, A.; Giridhar, P.; Ravishankar, G.A. Phytoserotonin: A review. Plant Signal Behav. 2011, 6, 800–809. [Google Scholar] [PubMed]
- Dong, X.; Chen, W.; Wang, W.; Zhang, H.; Liu, X.; Luo, J. Comprehensive profiling and natural variation of flavonoids in rice. J. Integr. Plant Biol. 2014, 56, 876–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagaharu, U. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. Jpn. J. Bot. 1935, 7, 389–452. [Google Scholar]
- Guo, L.; Yang, H.; Zhang, X.; Yang, S. Lipid transfer protein 3 as a target of MYB96 mediates freezing and drought stress in Arabidopsis. J. Exp. Bot. 2013, 64, 1755–1767. [Google Scholar] [CrossRef] [PubMed]
- Telfer, A.; Bollman, K.M.; Poethig, R.S. Phase change and the regulation of trichome distribution in Arabidopsis thaliana. Development 1997, 124, 645–654. [Google Scholar] [PubMed]
- Szymanski, D.B.; Jilk, R.A.; Pollock, S.M.; Marks, M.D. Control of GL2 expression in Arabidopsis leaves and trichomes. Development 1998, 125, 1161–1171. [Google Scholar] [PubMed]
- Hung, C.Y.; Lin, Y.; Zhang, M.; Pollock, S.; Marks, M.D.; Schiefelbein, J. A common position-dependent mechanism controls cell-type patterning and GLABRA2 regulation in the root and hypocotyl epidermis of Arabidopsis. Plant Physiol. 1998, 117, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Cai, W.J.; Wang, S.; Shan, C.M.; Wang, L.J.; Chen, X.Y. Temporal control of trichome distribution by microRNA156-targeted SPL genes in Arabidopsis thaliana. Plant Cell 2010, 22, 2322–2335. [Google Scholar] [CrossRef] [PubMed]
- Kader, J.C. Lipid-transfer proteins: A puzzling family of plant proteins. Trends Plant Sci. 1997, 2, 66–70. [Google Scholar] [CrossRef]
- Djordjevic, M.A.; Oakes, M.; Li, D.X.; Hwang, C.H.; Hocart, C.H.; Gresshoff, P.M. The glycine max xylem sap and apoplast proteome. J. Proteome Res. 2007, 6, 3771–3779. [Google Scholar] [CrossRef] [PubMed]
- Kusumawati, L.; Imin, N.; Djordjevic, M.A. Characterization of the secretome of suspension cultures of Medicago species reveals proteins important for defense and development. J. Proteome Res. 2008, 7, 4508–4520. [Google Scholar] [CrossRef] [PubMed]
- Pyee, J.; Yu, H.; Kolattukudy, P.E. Identification of a lipid transfer protein as the major protein in the surface wax of broccoli (Brassica oleracea) leaves. Arch. Biochem. Biophys. 1994, 311, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Mundy, J.; Rogers, J.C. Selective expression of a probable amylase/protease inhibitor in barley aleurone cells: Comparison to the barley amylase/subtilisin inhibitor. Planta 1986, 169, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Dani, V.; Simon, W.J.; Duranti, M.; Croy, R.R. Changes in the tobacco leaf apoplast proteome in response to salt stress. Proteomics 2005, 5, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Hülskamp, M.; Misra, S.; Jurgens, G. Genetic dissection of trichome cell development in Arabidopsis. Cell 1994, 76, 555–566. [Google Scholar] [CrossRef]
- Larkin, J.C.; Oppenheimer, D.G.; Lloyd, A.M.; Paparozzi, E.T.; Marks, M.D. Roles of the GLABROUS1 and TRANSPARENT TESTA GLABRA Genes in Arabidopsis Trichome Development. Plant Cell 1994, 6, 1065–1076. [Google Scholar] [CrossRef] [PubMed]
- Sohal, A.K.; Pallas, J.A.; Jenkins, G.I. The promoter of a Brassica napus lipid transfer protein gene is active in a range of tissues and stimulated by light and viral infection in transgenic Arabidopsis. Plant Mol. Biol. 1999, 41, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Kunst, L.; Samuels, A.L. Biosynthesis and secretion of plant cuticular wax. Prog. Lipid Res. 2003, 42, 51–80. [Google Scholar] [CrossRef]
- Liu, F.; Xiong, X.; Wu, L.; Fu, D.; Hayward, A.; Zeng, X.; Cao, Y.; Wu, Y.; Li, Y.; Wu, G. BraLTP1, a lipid transfer protein gene involved in epicuticular wax deposition, cell proliferation and flower development in Brassica napus. PLoS ONE 2014, 9, e110272. [Google Scholar] [CrossRef] [PubMed]
- Kirik, V.; Lee, M.M.; Wester, K.; Herrmann, U.; Zheng, Z.; Oppenheimer, D.; Schiefelbein, J.; Hülskamp, M. Functional diversification of MYB23 and GL1 genes in trichome morphogenesis and initiation. Development 2005, 132, 1477–1485. [Google Scholar] [CrossRef] [PubMed]
- Plett, J.M.; Wilkins, O.; Campbell, M.M.; Ralph, S.G.; Regan, S. Endogenous overexpression of Populus MYB186 increases trichome density, improves insect pest resistance, and impacts plant growth. Plant J. 2010, 64, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Gigolashvili, T.; Berger, B.; Mock, H.P.; Muller, C.; Weisshaar, B.; Flugge, U.I. The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. Plant J. 2007, 50, 886–901. [Google Scholar] [CrossRef]
- Jakoby, M.J.; Falkenhan, D.; Mader, M.T.; Brininstool, G.; Wischnitzki, E.; Platz, N.; Hudson, A.; Hülskamp, M.; Larkin, J.; Schnittger, A. Transcriptional profiling of mature Arabidopsis trichomes reveals that NOECK encodes the MIXTA-like transcriptional regulator MYB106. Plant Physiol. 2008, 148, 1583. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.M.; Walbot, V.; Davis, R.W. Arabidopsis and Nicotiana anthocyanin production activated by maize regulators R and C1. Science 1992, 258, 1773–1775. [Google Scholar] [CrossRef] [PubMed]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Oxford University Press: New York, NY, USA, 2014; Volume 6, pp. 1065–1068. [Google Scholar]
- Wen, W.; Li, D.; Li, X.; Gao, Y.; Li, W.; Li, H.; Liu, J.; Liu, H.; Chen, W.; Luo, J. Metabolome-based genome-wide association study of maize kernel leads to novel biochemical insights. Nat. Commun. 2014, 5, 3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, M.; Gao, Y. Evolutionarily distinct BAHD N-acyltransferases are responsible for natural variation of aromatic amine conjugates in rice. Plant Cell 2016, 28, 1533–1550. [Google Scholar] [CrossRef] [PubMed]
- Yannai, S. Dictionary of Food Compounds with CD-ROM: Additives, Flavors, and Ingredients; CRC Press: Boca Raton, FL, USA, 2013; Volume 63, p. 123. [Google Scholar]
- Lorenzen, M.; Racicot, V.; Strack, D.; Chapple, C. Sinapic acid ester metabolism in wild type and a sinapoylglucose-accumulating mutant of Arabidopsis. Plant Physiol. 1996, 112, 1625–1630. [Google Scholar] [CrossRef] [PubMed]
- Stobiecki, M. Application of mass spectrometry for identification and structural studies of flavonoid glycosides. Phytochemistry 2000, 54, 237–256. [Google Scholar] [CrossRef]
- Lepiniec, L.; Debeaujon, I.; Routaboul, J.M.; Baudry, A.; Pourcel, L.; Nesi, N.; Caboche, M. Genetics and biochemistry of seed flavonoids. Annu. Rev. Plant Biol. 2006, 57, 405–430. [Google Scholar] [CrossRef] [PubMed]
- Golldack, D.; Luking, I.; Yang, O. Plant tolerance to drought and salinity: Stress regulating transcription factors and their functional significance in the cellular transcriptional network. Plant Cell Rep. 2011, 30, 1383–1391. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.S.; Chern, M.; Bartley, L.E.; Han, M.; Jung, K.H.; Lee, I.; Walia, H.; Richter, T.; Xu, X.; Cao, P.; et al. Towards establishment of a rice stress response interactome. PLoS Genet. 2011, 7, e1002020. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.G.; Swinny, E.E.; Winefield, C.; Markham, K.R. Flavonoids and UV photoprotection in Arabidopsis mutants. Z. Naturforsch. C. 2001, 56, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Sheahan, J.J. Sinapate Esters Provide greater UV-B attenuation than flavonoids in Arabidopsis thaliana (Brassicaceae). Am. J. Bot. 1996, 83, 679–686. [Google Scholar] [CrossRef]
- N’Guessan, J.D.; Bidié, A.P.; Lenta, B.N.; Weniger, B.; André, P.; Guédéguina, F. In vitro assays for bioactivity-guided isolation of antisalmonella and antioxidant compounds in Thonningia sanguinea flowers. Afr. J. Biotechnol. 2007, 6, 1685–1689. [Google Scholar]
- Hawas, U.W. Antioxidant activity of brocchlin carboxylic acid and its methyl ester from Chrozophora brocchiana. Nat. Prod. Res. 2007, 21, 632–640. [Google Scholar] [CrossRef] [PubMed]
- Herbette, S.; Menn, A.L.; Rousselle, P.; Ameglio, T.; Faltin, Z.; Branlard, G.; Eshdat, Y.; Julien, J.L.; Drevet, J.R.; Roeckel-Drevet, P. Modification of photosynthetic regulation in tomato overexpressing glutathione peroxidase. Biochim. Biophys. Acta 2005, 1724, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Ascorbate and glutathione: The heart of the redox hub. Plant Physiol. 2011, 155, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, J.; Zhang, X.; Xu, Q.; Zhou, S.; Xu, S. Effect of Salt Priming on Germination and Seedling Establishment of Rape Seed under Different Water Conditions. Seed 2004, 23, 6–8. [Google Scholar]
- Lin, R.; Wang, X.; Luo, Y.; Du, W.; Guo, H.; Yin, D. Effects of soil cadmium on growth, oxidative stress and antioxidant system in wheat seedlings (Triticum aestivum L.). Chemosphere 2007, 69, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Shang, S.; Feng, X.; Zhang, G.; Wu, F. Modulation of exogenous selenium in cadmium-induced changes in antioxidative metabolism, cadmium uptake, and photosynthetic performance in the 2 tobacco genotypes differing in cadmium tolerance. Environ. Toxicol. Chem. 2015, 34, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Hong, C.; Chen, X.; Xu, Y.; Liu, Y.; Jiang, D.; Zheng, B. Different growth and physiological responses to cadmium of the three miscanthus species. PLoS ONE 2016, 11, e0153475. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef] [PubMed]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: Beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [PubMed]
- Yang, C.; Li, H.; Zhang, J.; Luo, Z.; Gong, P.; Zhang, C.; Li, J.; Wang, T.; Zhang, Y.; Lu, Y.; et al. A regulatory gene induces trichome formation and embryo lethality in tomato. Proc. Natl. Acad. Sci. USA 2011, 108, 11836–11841. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel integrated method for large-scale detection, identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X.; Zhang, H.; Dong, H.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Dodge, A.D. The mode of action of the bipyridylium herbicides, paraquat and diquat. Endeavour 1971, 30, 130–135. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component Name | Organism | Position in (+) Strand | Sequence | Function | |
|---|---|---|---|---|---|
| Start | End | ||||
| ABRE | Arabidopsis thaliana | –1184 –254 –166 | –1179 –250 –160 | CACGTGACGTGCACGCGG | cis–acting element involved in the abscisic acid responsiveness |
| ARE | Zea mays | –661 –546 | –655 –541 | TAACGTATGGTTT | cis–acting regulatory element essential for the anaerobic induction |
| P-Box | Petroselinum crispum | –975 | –970 | TTGACC | Gibberellin-responsive element |
| CGTCA-motif | Hordeum vulgare | –213 | –207 | CCTTTTG | cis–acting regulatory element involved in the Methyl jasmonate-responsiveness |
| DRE | Arabidopsis thaliana | –1141 | –1134 | TGGCCGAC | Regulatory element involved in cold- and dehydration-responsiveness |
| G-Box | Antirrhinum majus | –778 | –773 | CACGTT | cis–acting regulatory element involved in light responsiveness |
| MBS | Arabidopsis thaliana | –1090 | –1085 | CAACTG | MYB binding site involved in drought-inducibility |
| MYB | Arabidopsis thaliana | –109 –67 | –104 –62 | CTGTTA | MYB binding sites |
| Class | Number of Compounds Detected | Number of Compounds with Significant Quantitative Differences | Number of Compounds with Higher Concentration in 35::BraLTP2 | Number of Compounds with Lower Concentration in 35::BraLTP2 |
|---|---|---|---|---|
| Flavonoids | 55 | 21 | 20 | 1 |
| Phenylpropanoids | 32 | 10 | 6 | 4 |
| Phenolamides | 19 | 13 | 4 | 9 |
| Alkaloids | 24 | 8 | 2 | 6 |
| Terpenoids | 10 | 2 | 1 | 1 |
| Organic acids | 34 | 2 | 1 | 1 |
| Other metabolites | 97 | 17 | 9 | 8 |
| Total | 271 | 73 | 43 | 30 |
| No. a | Flavones | WT b | BraLTP2b | Fold Change | VIP |
|---|---|---|---|---|---|
| BCL0281 | Kaempferide | 6.07 × 102 | 3.15 × 104 | 51.87 | 3.35 |
| BCL0318 | Prunin | 1.27 × 103 | 7.03 × 103 | 5.56 | 2.11 |
| BCL0485 | Kaempferol | 1.03 × 104 | 5.59 × 104 | 5.43 | 2.22 |
| BCL0317 | Chrysoeriol O-hexoside | 2.02 × 103 | 1.00 × 104 | 4.95 | 2.10 |
| BCL0305 | Kaempferol 3-O-glucoside (Astragalin) | 2.00 × 104 | 9.20 × 104 | 4.60 | 1.81 |
| BCL0306 | Luteolin 5-O-hexoside | 4.14 × 104 | 1.70 × 105 | 4.10 | 1.98 |
| BCL0313 | Naringenin 7-O-glucoside | 5.65 × 103 | 2.08 × 104 | 3.68 | 1.87 |
| BCL0314 | Apigenin 7-O-glucoside (Cosmosiin) | 2.66 × 103 | 9.35 × 103 | 3.51 | 1.84 |
| BCL0267 | Luteolin O-hexosyl-O-hexoside | 2.37 × 104 | 7.42 × 104 | 3.13 | 1.75 |
| BCL0308 | Apigenin 7-O-glucoside | 5.35 × 103 | 1.54 × 104 | 2.87 | 1.72 |
| BCL0323 | Naringenin O-malonylhexoside | 1.77 × 104 | 5.03 × 104 | 2.84 | 1.72 |
| BCL0297 | 3’,4’,5’-Dihydrotricetin O-hexosyl-O-hexoside | 3.97 × 103 | 1.04 × 104 | 2.62 | 1.59 |
| BCL0268 | Quercetin O-hexoside | 1.01 × 104 | 2.59 × 104 | 2.57 | 1.49 |
| BCL0292 | Kaempferol 3-O-β -rutinoside (Nicotiflorin) | 5.36 × 103 | 1.36 × 104 | 2.54 | 1.20 |
| BCL0177 | Luteolin-3’,7-di-O-glucoside | 5.71 × 106 | 1.36 × 107 | 2.38 | 1.51 |
| BCL0295 | 3,4,2’,4’,6’-Pentamethoxychalcone | 1.31 × 104 | 2.62 × 104 | 2.00 | 1.27 |
| BCL0190 | Cyanidin 3-O-glucoside (Anthocyanins) | 1.54 × 106 | 6.24 × 106 | 4.06 | 1.96 |
| BCL0242 | Cyanidin (Anthocyanins) | 4.87 × 103 | 1.46 × 104 | 3.00 | 1.76 |
| BCL0191 | Delphinidin O-hexoside (Anthocyanins) | 5.52 × 104 | 1.65 × 105 | 2.98 | 1.69 |
| BCL0261 | Rosinidin O-hexoside (Anthocyanins) | 3.40 × 103 | 8.49 × 103 | 2.50 | 1.52 |
| No. | Phenylpropanoids | WT | BraLTP2 | Fold Change | VIP |
|---|---|---|---|---|---|
| BCL0304 | Ferulic acid | 2.60 × 103 | 1.49 × 104 | 5.7 | 2.24 |
| BCL0184 | Brevifolincarboxylic acid | 1.61 × 104 | 7.94 × 104 | 4.93 | 2.13 |
| BCL0364 | Geranyl acetate | 1.86 × 104 | 7.48 × 104 | 4.03 | 2.00 |
| BCL0315 | trans-Cinnamaldehyde | 4.57 × 103 | 1.11 × 104 | 2.44 | 1.50 |
| BCL0266 | Syringic acid | 2.00 × 103 | 4.89 × 103 | 2.44 | 1.52 |
| BCL0225 | Coumarin | 9.77 × 106 | 3.22 × 106 | 2.27 | 1.48 |
| No. | Phenolamides | WT | BraLTP2 | Fold Change | VIP |
|---|---|---|---|---|---|
| BCL0212 | N-Feruloyl agmatine | 4.51 × 103 | 1.78 × 105 | 39.54 | 3.13 |
| BCL0226 | N-Sinapoyl agmatine | 2.71 × 103 | 1.92 × 104 | 7.08 | 2.26 |
| BCL0189 | N-Coumaroyl agmatine | 2.02 × 103 | 8.02 × 103 | 3.97 | 1.82 |
| BCL0229 | N-Feruloyl spermidine | 8.84 × 104 | 1.79 × 105 | 2.03 | 1.34 |
| No. | Alkaloids, Terpenoids, and Organic acids | WT | BraLTP2 | Fold Change | VIP | |
|---|---|---|---|---|---|---|
| BCL0282 | Alkaloids (Tryptamines) | N-Nexosyl-p-coumaroyl serotonin | 1.56 × 103 | 1.65 × 105 | 105.77 | 3.69 |
| BCL0293 | 5-Methoxy-N,N-dimethyltryptamine | 5.36 × 104 | 1.85 × 105 | 3.45 | 1.85 | |
| BCL0384 | Terpenoids | Phytocassane C | 6.71 × 103 | 1.60 × 104 | 2.39 | 1.32 |
| BCL0296 | Organic acids | m-Anisic-acid | 7.44 × 103 | 1.86 × 104 | 2.83 | 1.57 |
| No. | Other Metabolites | WT | BraLTP2 | Fold Change | VIP | |
|---|---|---|---|---|---|---|
| BCL0273 | Indoles and its derivatives | IAA-Asp-N-Glc | 7.41 × 102 | 4.23 × 104 | 57.09 | 3.38 |
| BCL0321 | 1-Methoxyindole-3-carbaldehyde | 7.44 × 103 | 1.86 × 104 | 2.50 | 1.56 | |
| BCL0036 | Cholines | O-Phosphocholine | 9.49 × 103 | 3.36 × 104 | 3.54 | 1.88 |
| BCL0029 | sn-Glycero-3-phosphocho | 8.99 × 105 | 2.17 × 106 | 2.41 | 1.52 | |
| BCL0335 | Coumarins and its derivatives | 4-Methylumbelliferone | 1.49 × 104 | 3.51 × 104 | 2.35 | 1.26 |
| BCL0206 | Others | N-α-Benzenolarginine ethylester | 9.01 × 102 | 1.15 × 104 | 12.79 | 2.50 |
| BCL0256 | Bergamottin | 6.23 × 105 | 3.16 × 106 | 5.08 | 2.14 | |
| BCL0275 | 1-(3,4-Dichlorophenyl)-3-methylurea | 1.89 × 105 | 8.77 × 105 | 4.63 | 2.08 | |
| BCL0410 | 2-Amino-9-methyl-4-octadecene-1,3,8-triol | 4.80 × 103 | 1.12 × 104 | 2.33 | 1.49 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, N.; Liu, F.; Wang, P.; Yan, X.; Gao, H.; Zeng, X.; Wu, G. Overexpression of BraLTP2, a Lipid Transfer Protein of Brassica napus, Results in Increased Trichome Density and Altered Concentration of Secondary Metabolites. Int. J. Mol. Sci. 2018, 19, 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061733
Tian N, Liu F, Wang P, Yan X, Gao H, Zeng X, Wu G. Overexpression of BraLTP2, a Lipid Transfer Protein of Brassica napus, Results in Increased Trichome Density and Altered Concentration of Secondary Metabolites. International Journal of Molecular Sciences. 2018; 19(6):1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061733
Chicago/Turabian StyleTian, Nini, Fang Liu, Pandi Wang, Xiaohong Yan, Hongfei Gao, Xinhua Zeng, and Gang Wu. 2018. "Overexpression of BraLTP2, a Lipid Transfer Protein of Brassica napus, Results in Increased Trichome Density and Altered Concentration of Secondary Metabolites" International Journal of Molecular Sciences 19, no. 6: 1733. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061733