Comparative Analysis between the In Vivo Biodistribution and Therapeutic Efficacy of Adipose-Derived Mesenchymal Stromal Cells Administered Intraperitoneally in Experimental Colitis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
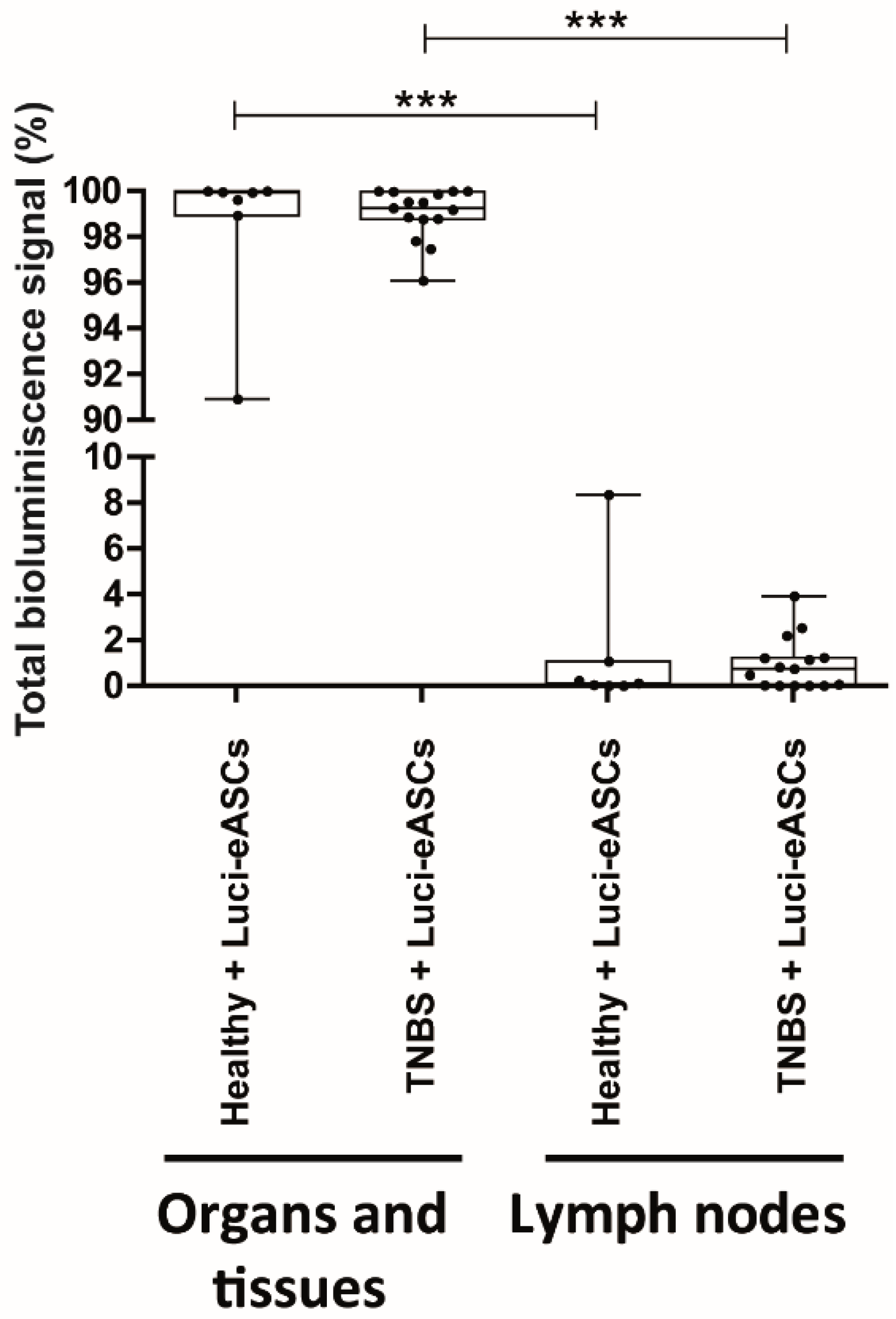
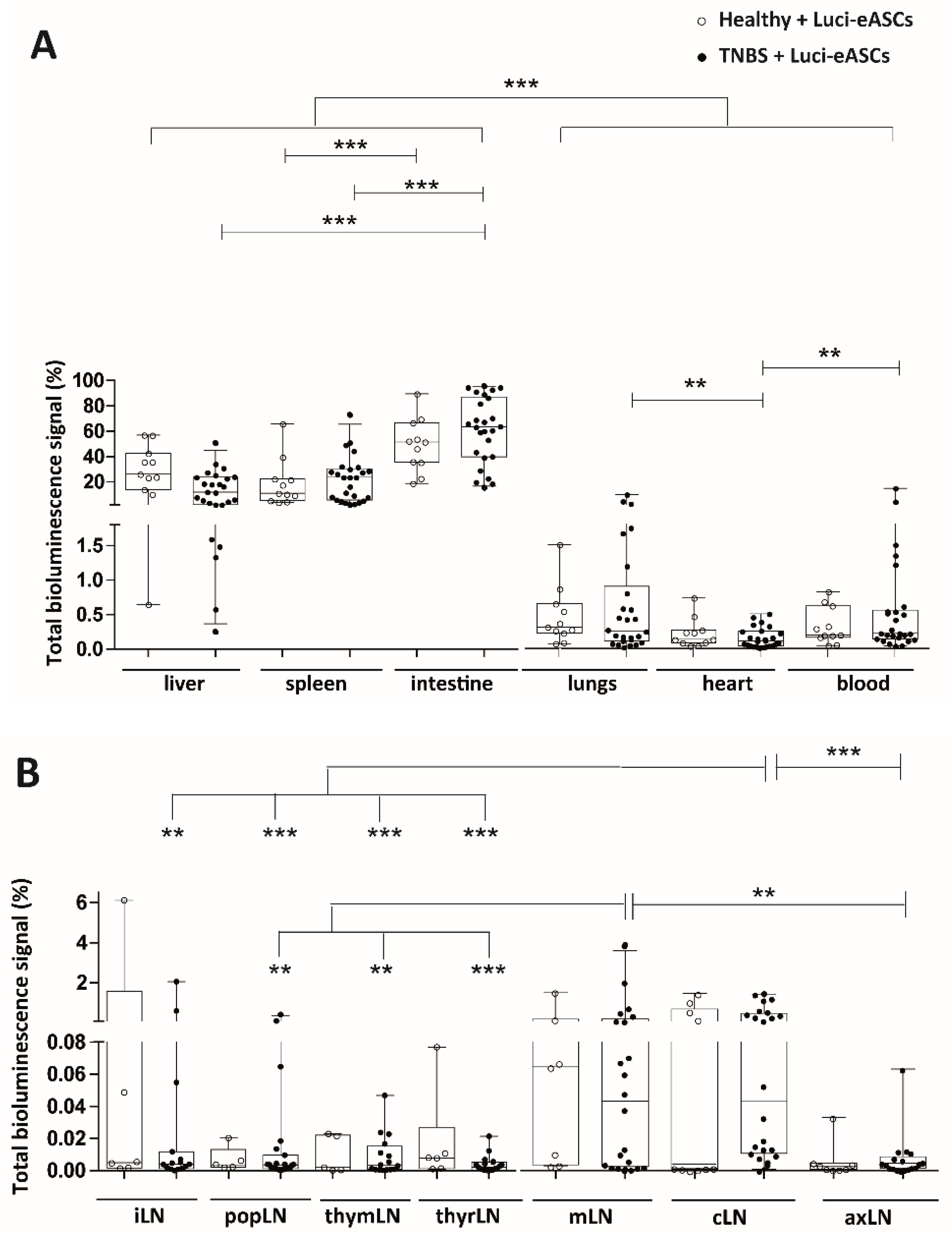
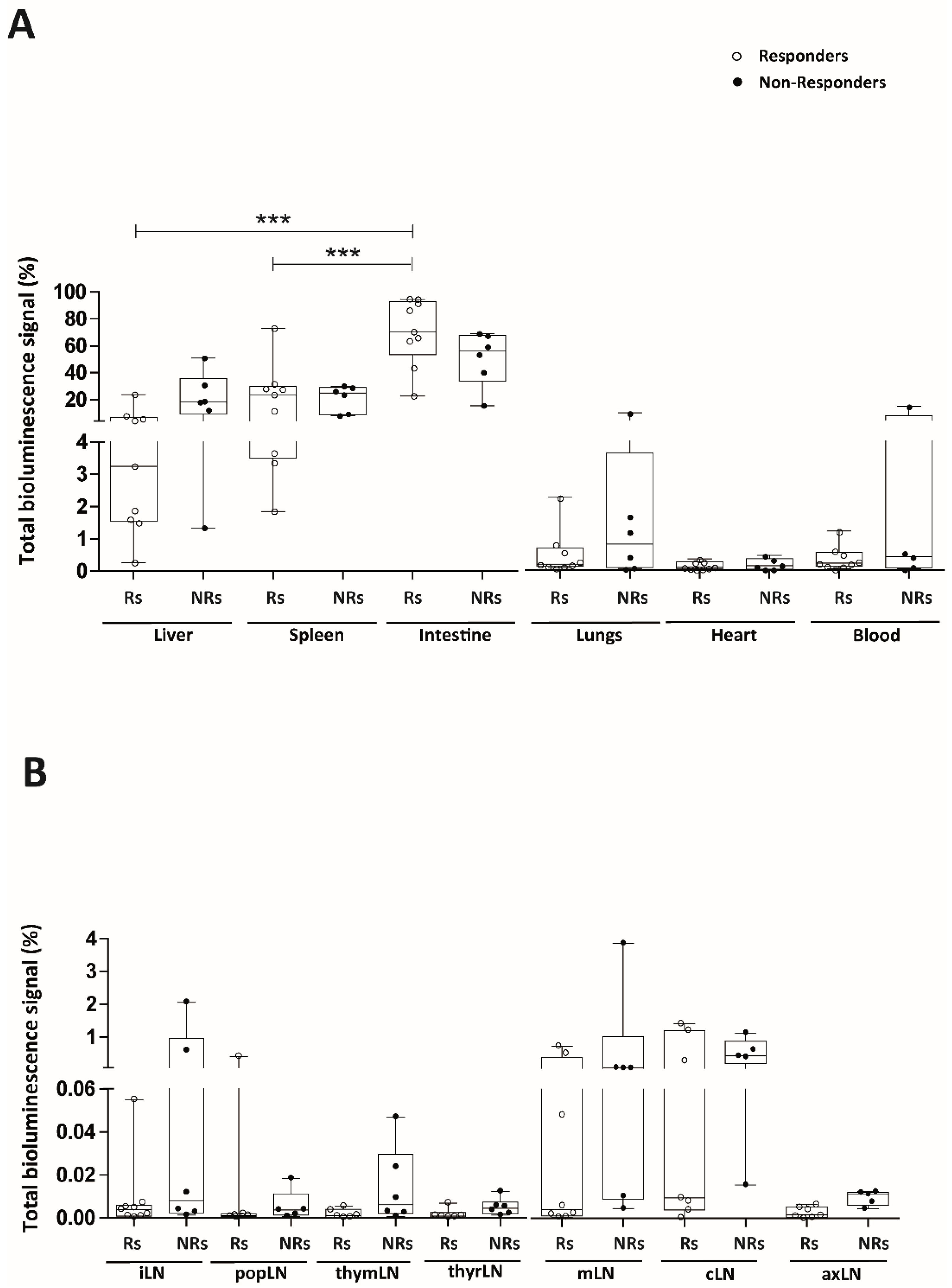
2.1. Intraperitoneally Administered Luci-eASCs Accumulate Preferentially in the Main Organs and Tissues Independently of the Inflammatory Status of the Mice
2.2. TNBS-Colitic Mice Treated with Luci-eASCs Showed an Increase in the Bioluminescence Signal in the Intestine
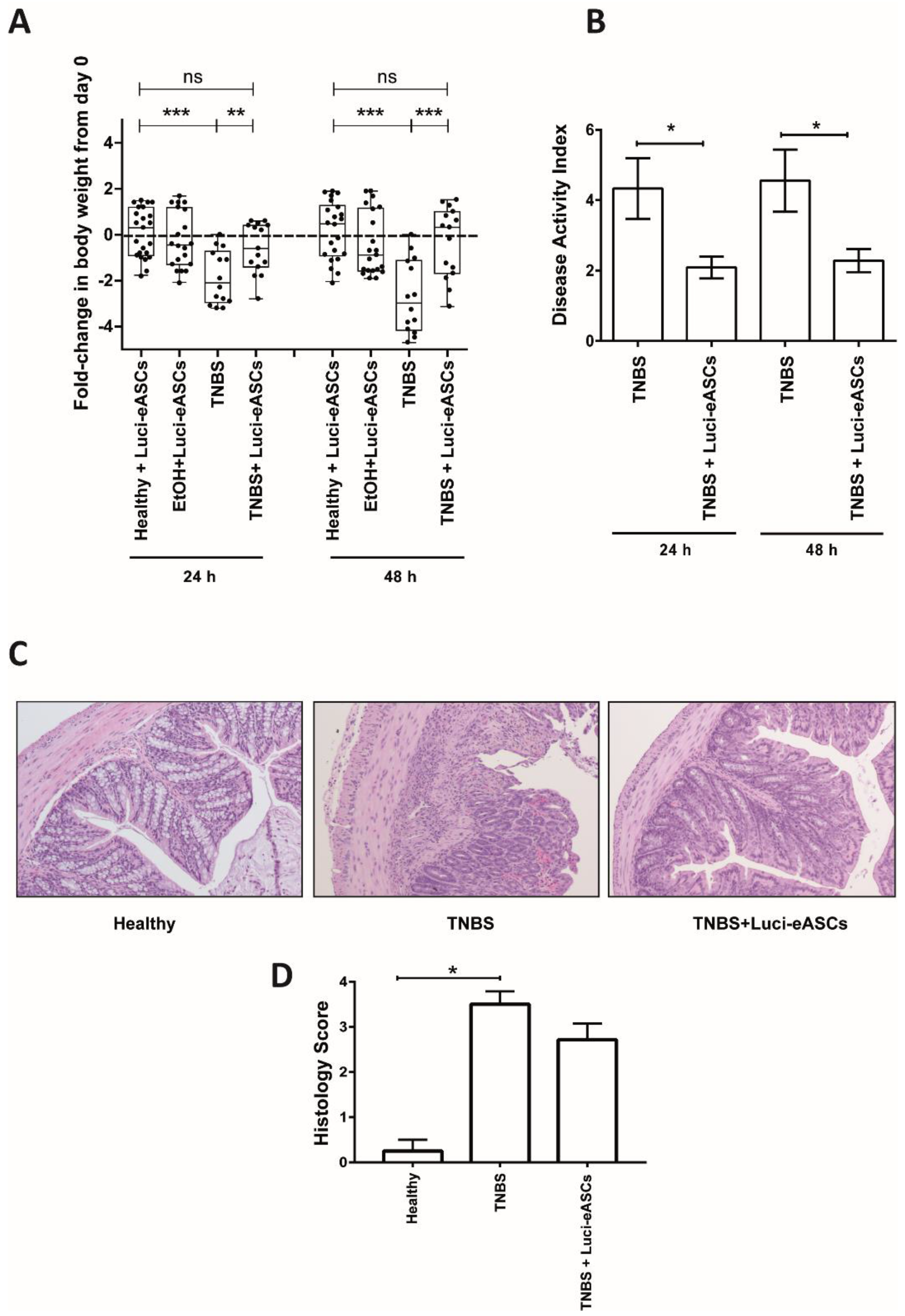
2.3. Luci-eASCs Administered IP Protect Against TNBS-Induced Colitis
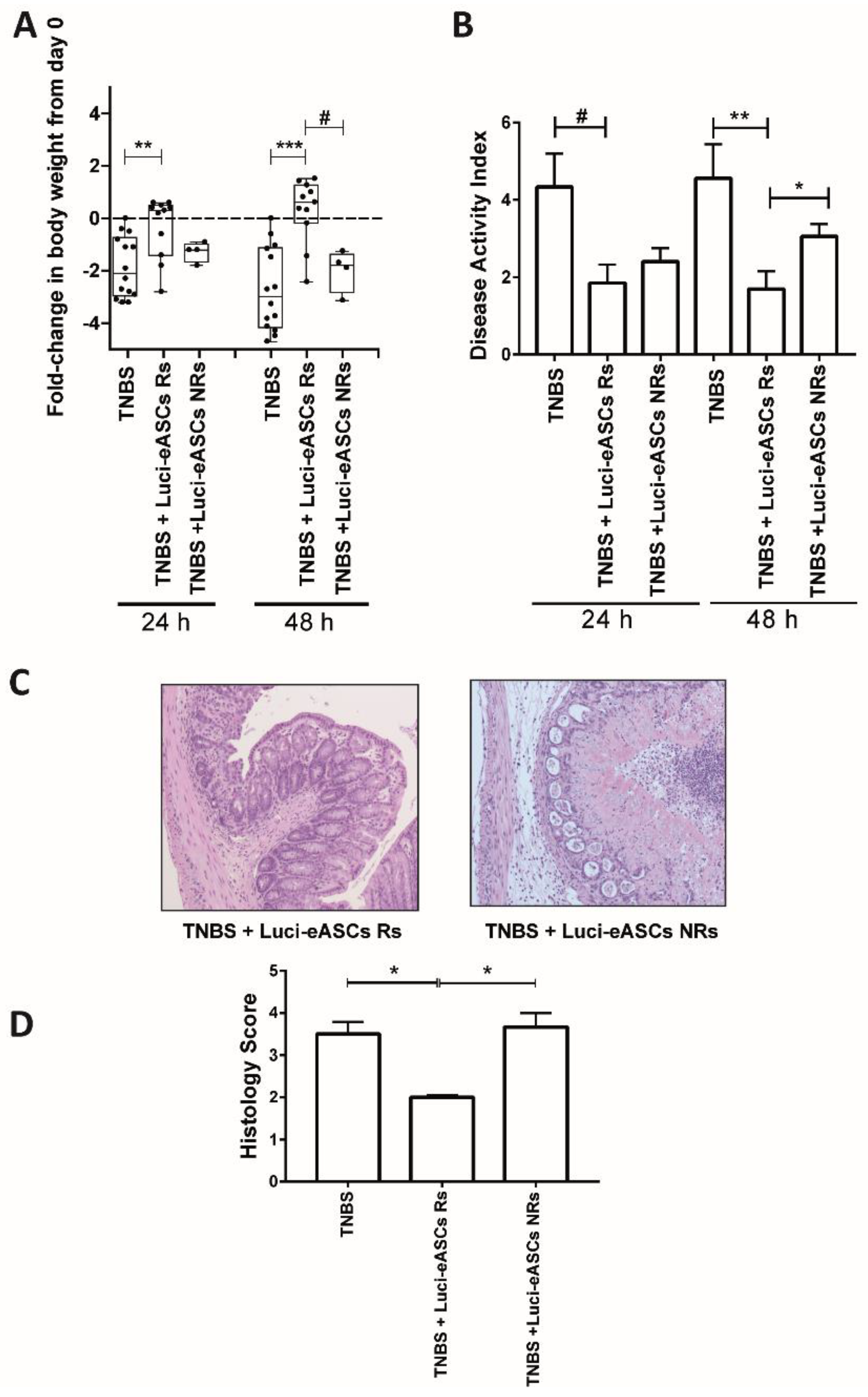
2.4. Comparative Analysis in the Biodistribution and the Therapeutic Effects of the Luci-eASCs
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Generation of Human Expanded Adipose-Derived Stem Cells
4.3. Generation of Luciferase+ Transduced eASCs (Luci-eASCs)
4.4. Immunophenotyping of Luci-eASCs
4.5. Immunosuppression Assay of Luci-eASCs
4.6. Induction and Evaluation of Colitis after Treatment with Luci-eASCs
4.7. Histological Analysis
4.8. In Vivo Optical Bioluminescence Imaging
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- English, K.; French, A.; Wood, K.J. Mesenchymal stromal cells: Facilitators of successful transplantation? Cell Stem Cell 2010, 7, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Bunnell, B.A.; Frazier, T.; Rowan, B.; Shah, F.; Thomas-Porch, C.; Wu, X. Adipose-derived stromal/stem cells: A primer. Organogenesis 2013, 9, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yu, F.; Sun, Y.; Jiang, B.; Zhang, W.; Yang, J.; Xu, G.T.; Liang, A.; Liu, S. Characteristics and Potential Applications of Human Dental Tissue-Derived Mesenchymal Stem Cells. Stem Cells 2014, 33, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Luz-Crawford, P.; Torres, M.J.; Noel, D.; Fernandez, A.; Toupet, K.; Alcayaga-Miranda, F.; Tejedor, G.; Jorgensen, C.; Illanes, S.E.; Figueroa, F.E.; et al. The immunosuppressive signature of menstrual blood mesenchymal stem cells entails opposite effects on experimental arthritis and graft versus host diseases. Stem Cells 2016, 34, 456–469. [Google Scholar] [CrossRef] [PubMed]
- Su, W.R.; Zhang, Q.Z.; Shi, S.H.; Nguyen, A.L.; Le, A.D. Human gingiva-derived mesenchymal stromal cells attenuate contact hypersensitivity via prostaglandin E2-dependent mechanisms. Stem Cells 2011, 29, 1849–1860. [Google Scholar] [CrossRef] [PubMed]
- Sessarego, N.; Parodi, A.; Podesta, M.; Benvenuto, F.; Mogni, M.; Raviolo, V.; Lituania, M.; Kunkl, A.; Ferlazzo, G.; Bricarelli, F.D.; et al. Multipotent mesenchymal stromal cells from amniotic fluid: Solid perspectives for clinical application. Haematologica 2008, 93, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Alvaro-Gracia, J.M.; Jover, J.A.; Garcia-Vicuna, R.; Carreno, L.; Alonso, A.; Marsal, S.; Blanco, F.; Martinez-Taboada, V.M.; Taylor, P.; Martin-Martin, C.; et al. Intravenous administration of expanded allogeneic adipose-derived mesenchymal stem cells in refractory rheumatoid arthritis (Cx611): Results of a multicentre, dose escalation, randomised, single-blind, placebo-controlled phase Ib/IIa clinical trial. Ann. Rheum. Dis. 2017, 76, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santalla, M.; Menta, R.; Mancheño-Corvo, P.; Lopez-Belmonte, J.; DelaRosa, O.; Bueren, J.A.; Dalemans, W.; Lombardo, E.; Garin, M.I. Adipose-derived mesenchymal stromal cells modulate experimental autoimmune arthritis by inducing an early regulatory innate cell signature. Immun. Inflamm. Dis. 2016, 4, 213–244. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santalla, M.; Mancheño-Corvo, P.; Menta, R.; Lopez-Belmonte, J.; DelaRosa, O.; Bueren, J.A.; Dalemans, W.; Lombardo, E.; Garin, M.I. Human adipose-derived mesenchymal stem cells modulate experimental autoimmune arthritis by modifying early adaptive T cell responses. Stem Cells 2015, 33, 3493–3503. [Google Scholar] [CrossRef] [PubMed]
- Nagaishi, K.; Arimura, Y.; Fujimiya, M. Stem cell therapy for inflammatory bowel disease. J. Gastroenterol. 2015, 50, 280–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Wang, L.; Cong, X.; Liu, G.; Zhou, J.; Bai, B.; Li, Y.; Bai, W.; Li, M.; Ji, H.; et al. Human umbilical cord mesenchymal stem cell therapy for patients with active rheumatoid arthritis: Safety and efficacy. Stem Cells Dev. 2013, 22, 3192–3202. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, H.; Liang, J.; Li, X.; Feng, X.; Wang, H.; Hua, B.; Liu, B.; Lu, L.; Gilkeson, G.S.; et al. Allogeneic mesenchymal stem cell transplantation in severe and refractory systemic lupus erythematosus: 4 years of experience. Cell Transplant. 2013, 22, 2267–2277. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, G.I.; Augello, A.; De Bari, C. Role of mesenchymal stem cells in reestablishing immunologic tolerance in autoimmune rheumatic diseases. Arthritis Rheumatol. 2011, 63, 2547–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.; Zhang, H.; Hua, B.; Wang, H.; Lu, L.; Shi, S.; Hou, Y.; Zeng, X.; Gilkeson, G.S.; Sun, L. Allogenic mesenchymal stem cells transplantation in refractory systemic lupus erythematosus: A pilot clinical study. Ann. Rheum. Dis. 2010, 69, 1423–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constantin, G.; Marconi, S.; Rossi, B.; Angiari, S.; Calderan, L.; Anghileri, E.; Gini, B.; Bach, S.D.; Martinello, M.; Bifari, F.; et al. Adipose-derived mesenchymal stem cells ameliorate chronic experimental autoimmune encephalomyelitis. Stem Cells 2009, 27, 2624–2635. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Soto-Gutierrez, A.; Parekkadan, B.; Kitagawa, Y.; Tompkins, R.G.; Kobayashi, N.; Yarmush, M.L. Mesenchymal stem cells: Mechanisms of immunomodulation and homing. Cell. Transplant. 2010, 19, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Santalla, M.; Mancheño-Corvo, P.; Escolano, A.; Menta, R.; Delarosa, O.; Abad, J.L.; Büscher, D.; Redondo, J.M.; Bueren, J.A.; Dalemans, W.; et al. Biodistribution and efficacy of human adipose-derived mesenchymal stem cells following intranodal administration in experimental colitis. Front. Immunol. 2017, 8, 638. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.J.; Prigent, J.; Dollet, P.E.; Ravau, J.; Larbanoix, L.; Van Simaeys, G.; Bol, A.; Gregoire, V.; Goldman, S.; Deblandre, G.; et al. Long-Term In Vivo Monitoring of Adult-Derived Human Liver Stem/Progenitor Cells by Bioluminescence Imaging, Positron Emission Tomography, and Contrast-Enhanced Computed Tomography. Stem Cells Dev. 2017, 26, 986–1002. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liang, C.; Hu, H.; Zhou, L.; Xu, B.; Wang, X.; Han, Y.; Nie, Y.; Jia, S.; Liang, J.; et al. Intraperitoneal injection (IP), Intravenous injection (IV) or anal injection (AI)? Best way for mesenchymal stem cells transplantation for colitis. Sci. Rep. 2016, 6, 30696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toupet, K.; Maumus, M.; Luz-Crawford, P.; Lombardo, E.; Lopez-Belmonte, J.; van Lent, P.; Garin, M.I.; van den Berg, W.; Dalemans, W.; Jorgensen, C.; et al. Survival and biodistribution of xenogenic adipose mesenchymal stem cells is not affected by the degree of inflammation in arthritis. PLoS ONE 2015, 10, e0114962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.W.; Lee, H.W.; Shin, S.; Park, J.H.; Yun, T.W.; Youn, H.Y.; Kim, S.J. Comparative efficacies of long-term serial transplantation of syngeneic, allogeneic, xenogeneic, or CTLA4Ig overproducing xenogeneic adipose tissue-derived mesenchymal stem cells on murine systemic lupus erythematosus. Cell Transplant. 2015, 25, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.S.; Golding, J.P.; Chapon, C.; Jones, W.A.; Bhakoo, K.K. Homing of stem cells to sites of inflammatory brain injury after intracerebral and intravenous administration: A longitudinal imaging study. Stem Cell Res. Ther. 2010, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Hamidian, J.S.; Estrada, C.; Li, Y.; Cheng, E.; Davies, J.E. Human Umbilical Cord Perivascular Cells and Human Bone Marrow Mesenchymal Stromal Cells Transplanted Intramuscularly Respond to a Distant Source of Inflammation. Stem Cells Dev. 2018, 27, 415–429. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, F.E.; Sobh, M.A.; Abouelkheir, M.; Khater, Y.; Mahmoud, K.; Saad, M.A.; Sobh, M.A. Study of the Effect of Route of Administration of Mesenchymal Stem Cells on Cisplatin-Induced Acute Kidney Injury in Sprague Dawley Rats. Int. J. Stem Cells 2016, 9, 79–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munir, H.; McGettrick, H.M. Mesenchymal Stem Cell Therapy for Autoimmune Disease: Risks and Rewards. Stem Cells Dev. 2015, 24, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Eggenhofer, E.; Benseler, V.; Kroemer, A.; Popp, F.C.; Geissler, E.K.; Schlitt, H.J.; Baan, C.C.; Dahlke, M.H.; Hoogduijn, M.J. Mesenchymal stem cells are short-lived and do not migrate beyond the lungs after intravenous infusion. Front. Immunol. 2012, 26, 297. [Google Scholar] [CrossRef] [PubMed]
- Castelo-Branco, M.T.; Soares, I.D.; Lopes, D.V.; Buongusto, F.; Martinusso, C.A.; do Rosario, A., Jr; Souza, S.A.; Gutfilen, B.; Fonseca, L.M.; Elia, C.; et al. Intraperitoneal but not intravenous cryopreserved mesenchymal stromal cells home to the inflamed colon and ameliorate experimental colitis. PLoS ONE 2012, 7, e33360. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Dennis, J.E.; Muzic, R.F.; Lundberg, M.; Caplan, A.I. The dynamic in vivo distribution of bone marrow-derived mesenchymal stem cells after infusion. Cells Tissues Org. 2001, 169, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Ko, J.H.; Jeong, H.J.; Ko, A.Y.; Kim, M.K.; Wee, W.R.; Yoon, S.O.; Oh, J.Y. Mesenchymal Stem/Stromal Cells Protect against Autoimmunity via CCL2-Dependent Recruitment of Myeloid-Derived Suppressor Cells. J. Immunol. 2015, 194, 3634–3645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assis, A.C.; Carvalho, J.L.; Jacoby, B.A.; Ferreira, R.L.; Castanheira, P.; Diniz, S.O.; Cardoso, V.N.; Goes, A.M.; Ferreira, A.J. Time-dependent migration of systemically delivered bone marrow mesenchymal stem cells to the infarcted heart. Cell Transplant. 2010, 19, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Eggenhofer, E.; Luk, F.; Dahlke, M.H.; Hoogduijn, M.J. The life and fate of mesenchymal stem cells. Front. Immunol. 2014, 19, 148. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.A.; Gonzalez-Rey, E.; Rico, L.; Buscher, D.; Delgado, M. Adipose-derived mesenchymal stem cells alleviate experimental colitis by inhibiting inflammatory and autoimmune responses. Gastroenterology 2009, 136, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Mancheno-Corvo, P.; Lopez-Santalla, M.; Menta, R.; DelaRosa, O.; Mulero, F.; Del Rio, B.; Ramirez, C.; Buscher, D.; Bueren, J.A.; Lopez-Belmonte, J.; et al. Intralymphatic Administration of Adipose Mesenchymal Stem Cells Reduces the Severity of Collagen-Induced Experimental Arthritis. Front. Immunol. 2017, 8, 462. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rey, E.; Anderson, P.; Gonzalez, M.A.; Rico, L.; Buscher, D.; Delgado, M. Human adult stem cells derived from adipose tissue protect against experimental colitis and sepsis. Gut 2009, 58, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Bramhall, M.; Florez-Vargas, O.; Stevens, R.; Brass, A.; Cruickshank, S. Quality of methods reporting in animal models of colitis. Inflamm. Bowel Dis. 2015, 21, 1248–1259. [Google Scholar] [CrossRef] [PubMed]
- Goncalves Fda, C.; Schneider, N.; Pinto, F.O.; Meyer, F.S.; Visioli, F.; Pfaffenseller, B.; Lopez, P.L.; Passos, E.P.; Cirne-Lima, E.O.; Meurer, L.; et al. Intravenous vs intraperitoneal mesenchymal stem cells administration: What is the best route for treating experimental colitis? World J. Gastroenterol. 2014, 20, 18228–18239. [Google Scholar] [CrossRef] [PubMed]
- Bazhanov, N.; Ylostalo, J.H.; Bartosh, T.J.; Tiblow, A.; Mohammadipoor, A.; Foskett, A.; Prockop, D.J. Intraperitoneally infused human mesenchymal stem cells form aggregates with mouse immune cells and attach to peritoneal organs. Stem Cell Res. Ther. 2016, 7, 27. [Google Scholar] [CrossRef] [PubMed]
- Parekkadan, B.; Tilles, A.W.; Yarmush, M.L. Bone marrow-derived mesenchymal stem cells ameliorate autoimmune enteropathy independently of regulatory T. cells. Stem Cells 2008, 26, 1913–1919. [Google Scholar] [CrossRef] [PubMed]
- Maria, A.T.; Toupet, K.; Maumus, M.; Fonteneau, G.; Le Quellec, A.; Jorgensen, C.; Guilpain, P.; Noel, D. Human adipose mesenchymal stem cells as potent anti-fibrosis therapy for systemic sclerosis. J. Autoimmun. 2016, 70, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez-Fernandez, M.; Rodriguez-Frutos, B.; Ramos-Cejudo, J.; Otero-Ortega, L.; Fuentes, B.; Vallejo-Cremades, M.T.; Sanz-Cuesta, B.E.; Diez-Tejedor, E. Comparison between xenogeneic and allogeneic adipose mesenchymal stem cells in the treatment of acute cerebral infarct: Proof of concept in rats. J. Transl. Med. 2015, 13, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cargnoni, A.; Gibelli, L.; Tosini, A.; Signoroni, P.B.; Nassuato, C.; Arienti, D.; Lombardi, G.; Albertini, A.; Wengler, G.S.; Parolini, O. Transplantation of allogeneic and xenogeneic placenta-derived cells reduces bleomycin-induced lung fibrosis. Cell Transplant. 2009, 18, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Anderson, P.; Souza-Moreira, L.; Morell, M.; Caro, M.; O’Valle, F; Gonzalez-Rey, E.; Delgado, M. Adipose-derived mesenchymal stromal cells induce immunomodulatory macrophages which protect from experimental colitis and sepsis. Gut 2013, 62, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Dong, C.; Chen, X.; Fang, Z.; Xu, J.; Liu, M.; Zhang, X.; Gu, D.S.; Wang, D.; Du, W.; et al. Human umbilical cord mesenchymal stem cells ameliorate mice trinitrobenzene sulfonic acid (TNBS)-induced colitis. Cell Transplant. 2011, 20, 1395–1408. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Shin, T.H.; Lee, B.C.; Yu, K.R.; Seo, Y.; Lee, S.; Seo, M.S.; Hong, I.S.; Choi, S.W.; Seo, K.W.; et al. Human umbilical cord blood mesenchymal stem cells reduce colitis in mice by activating NOD2 signaling to COX2. Gastroenterology 2013, 145, 1392–1403, e1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Martin-Arranz, E.; Martin-Arranz, M.; Robredo, T.; Mancheño-Corvo, P.; Menta, R.; Alves, F.; Suarez-de-Parga, F.; Mora-Sanz, P.; DelaRosa, O.; Büscher, D.; et al. Endoscopic submucosal injection of adipose-derived mesenchymal stem cells ameliorates TNBS-induced colitis in rats and prevents stenosis. Stem Res. Ther. 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menta, R.; Mancheno-Corvo, P.; Del Rio, B.; Ramirez, C.; DelaRosa, O.; Dalemans, W.; Lombardo, E. Tryptophan concentration is the main mediator of the capacity of adipose mesenchymal stromal cells to inhibit T-lymphocyte proliferation in vitro. Cytotherapy 2014, 16, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- DelaRosa, O.; Lombardo, E.; Beraza, A.; Mancheno-Corvo, P.; Ramirez, C.; Menta, R.; Rico, L.; Camarillo, E.; Garcia, L.; Abad, J.L.; et al. Requirement of IFN-gamma-mediated indoleamine 2,3-dioxygenase expression in the modulation of lymphocyte proliferation by human adipose-derived stem cells. Tissue Eng. Part. A 2009, 15, 2795–2806. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopez-Santalla, M.; Mancheño-Corvo, P.; Escolano, A.; Menta, R.; Delarosa, O.; Redondo, J.M.; Bueren, J.A.; Dalemans, W.; Lombardo, E.; Garin, M.I. Comparative Analysis between the In Vivo Biodistribution and Therapeutic Efficacy of Adipose-Derived Mesenchymal Stromal Cells Administered Intraperitoneally in Experimental Colitis. Int. J. Mol. Sci. 2018, 19, 1853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071853
Lopez-Santalla M, Mancheño-Corvo P, Escolano A, Menta R, Delarosa O, Redondo JM, Bueren JA, Dalemans W, Lombardo E, Garin MI. Comparative Analysis between the In Vivo Biodistribution and Therapeutic Efficacy of Adipose-Derived Mesenchymal Stromal Cells Administered Intraperitoneally in Experimental Colitis. International Journal of Molecular Sciences. 2018; 19(7):1853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071853
Chicago/Turabian StyleLopez-Santalla, Mercedes, Pablo Mancheño-Corvo, Amelia Escolano, Ramon Menta, Olga Delarosa, Juan M. Redondo, Juan A. Bueren, Wilfried Dalemans, Eleuterio Lombardo, and Marina I. Garin. 2018. "Comparative Analysis between the In Vivo Biodistribution and Therapeutic Efficacy of Adipose-Derived Mesenchymal Stromal Cells Administered Intraperitoneally in Experimental Colitis" International Journal of Molecular Sciences 19, no. 7: 1853. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071853