LncRNA ITGB2-AS1 Could Promote the Migration and Invasion of Breast Cancer Cells through Up-Regulating ITGB2

Abstract
:1. Introduction
2. Results
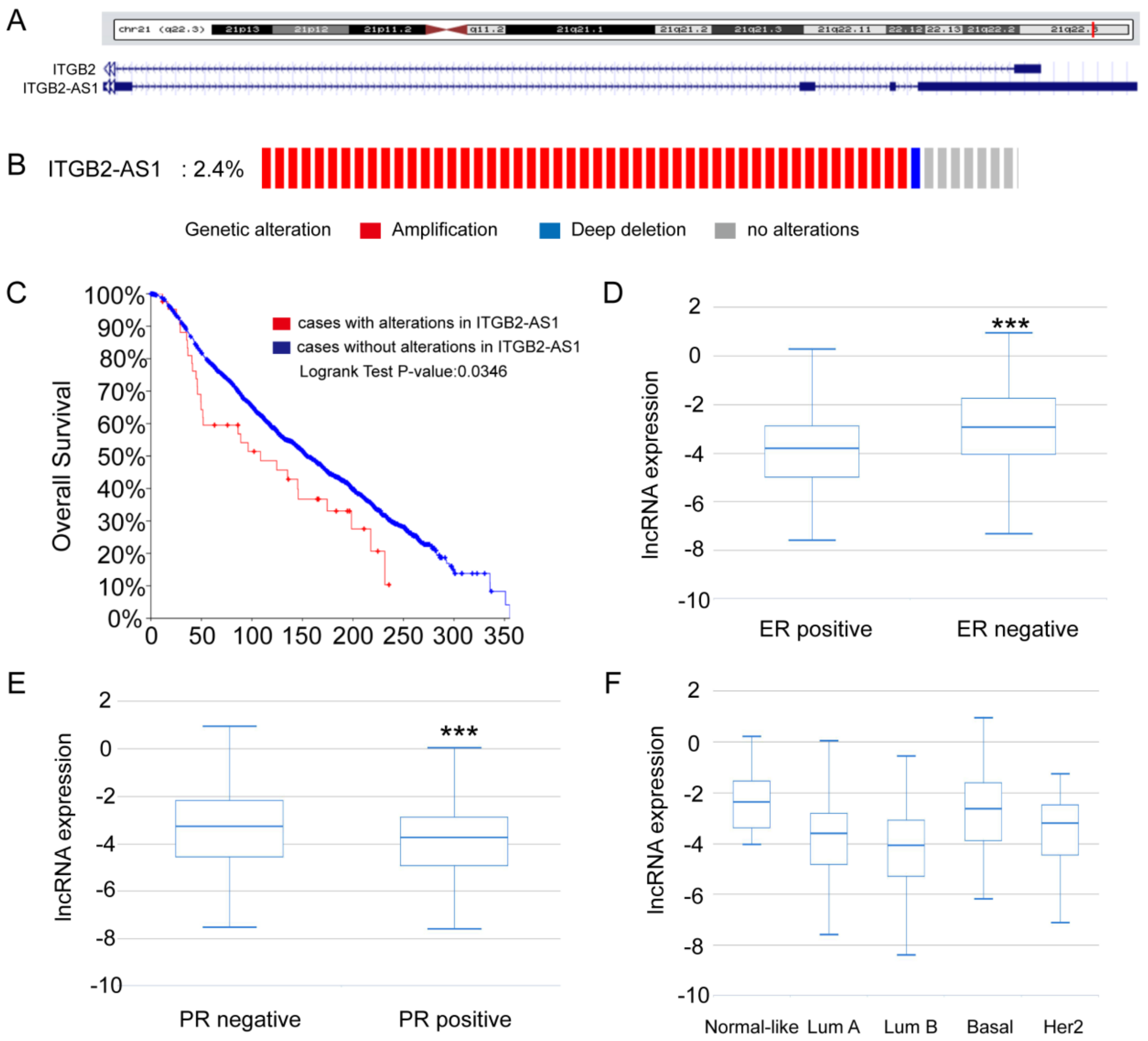
2.1. The Expression Analysis of ITGB2-AS1 on Online Database
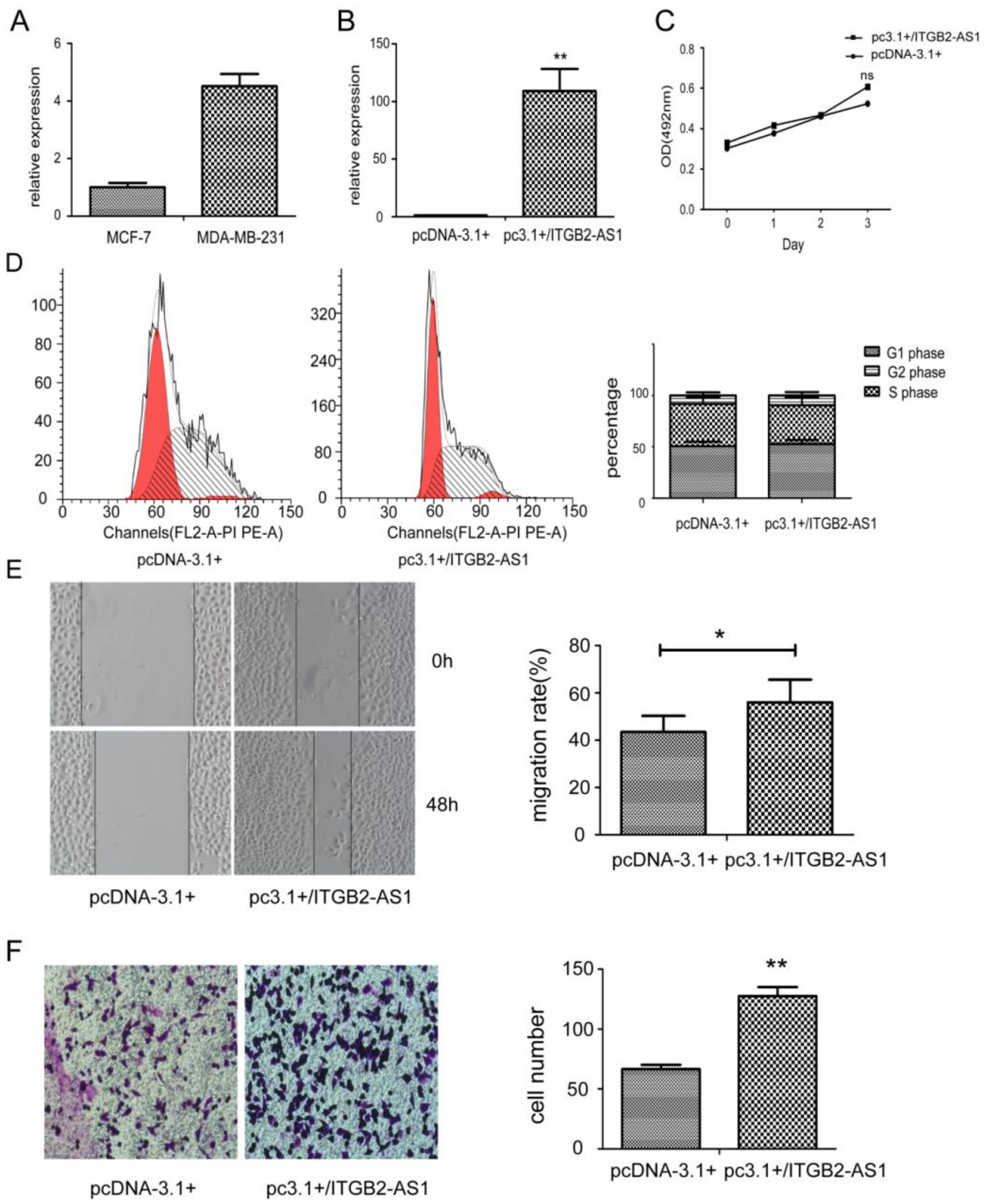
2.2. Overexpression of ITGB2-AS1 Could Promote the Migration and Invasion of MCF-7 Cell
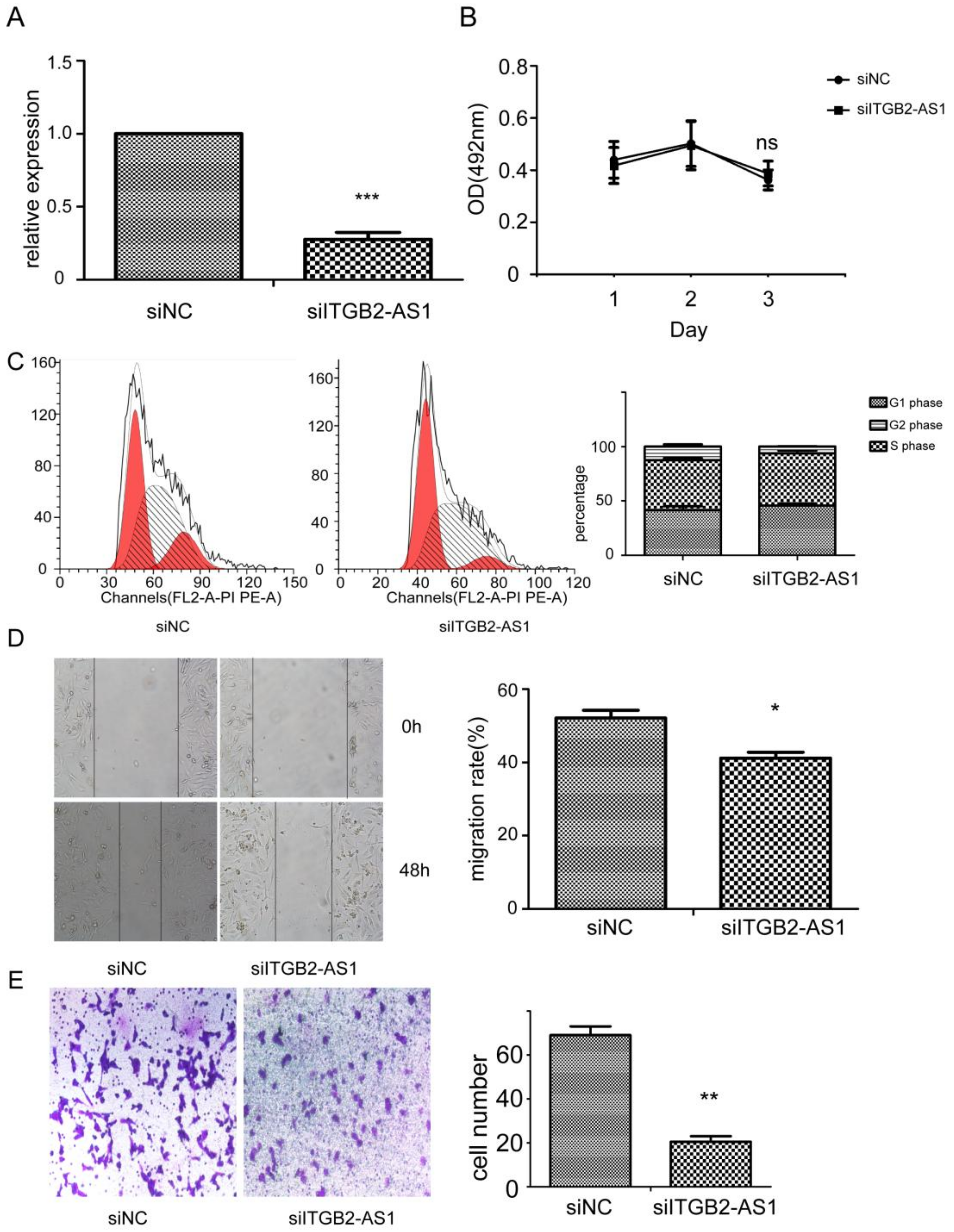
2.3. Knock-Down of ITGB2-AS1 Exerted Inhibitory Effect on Breast Cancer
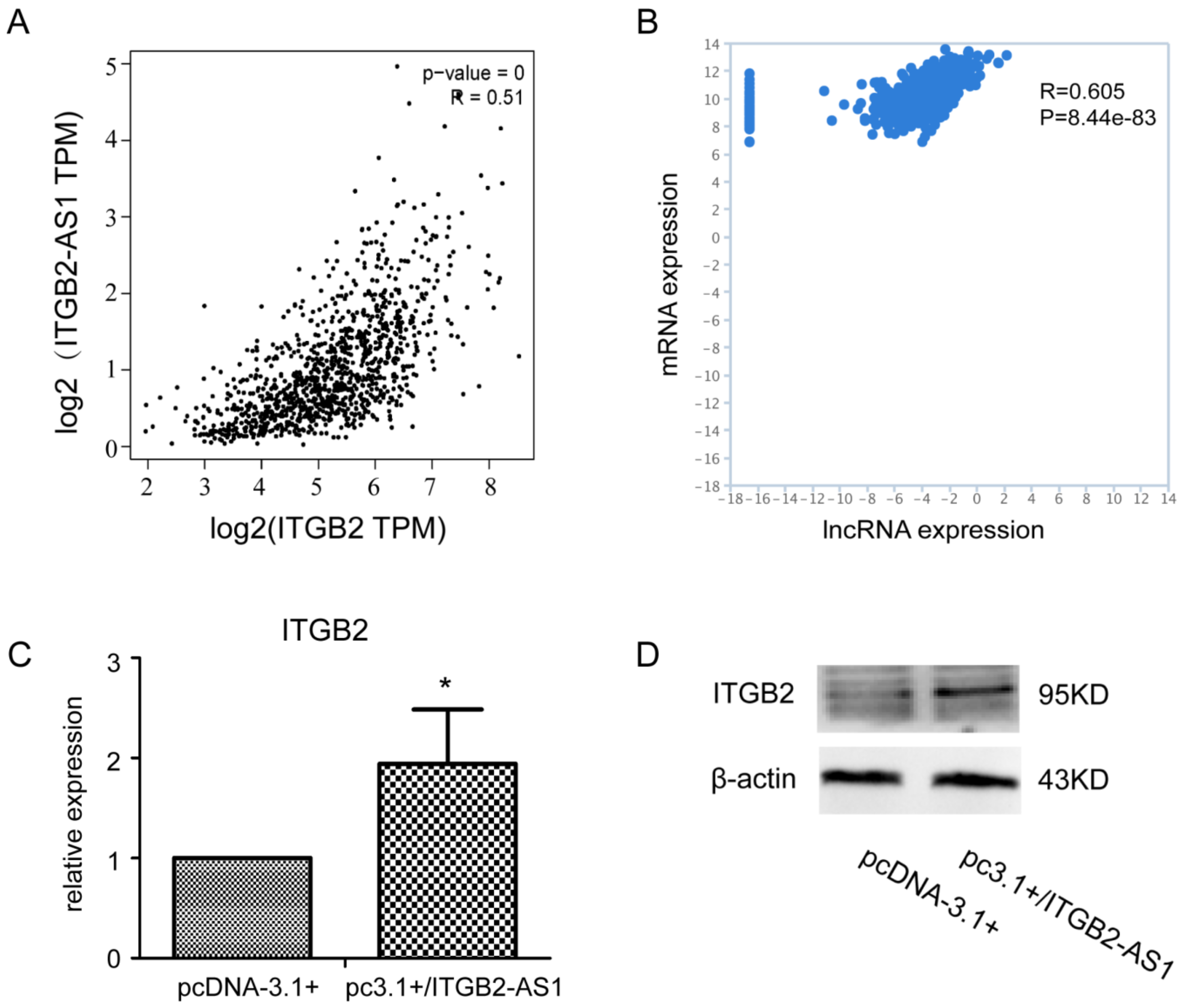
2.4. There Is a Positive Correlation between the Expression of ITGB2-AS1 and ITGB2
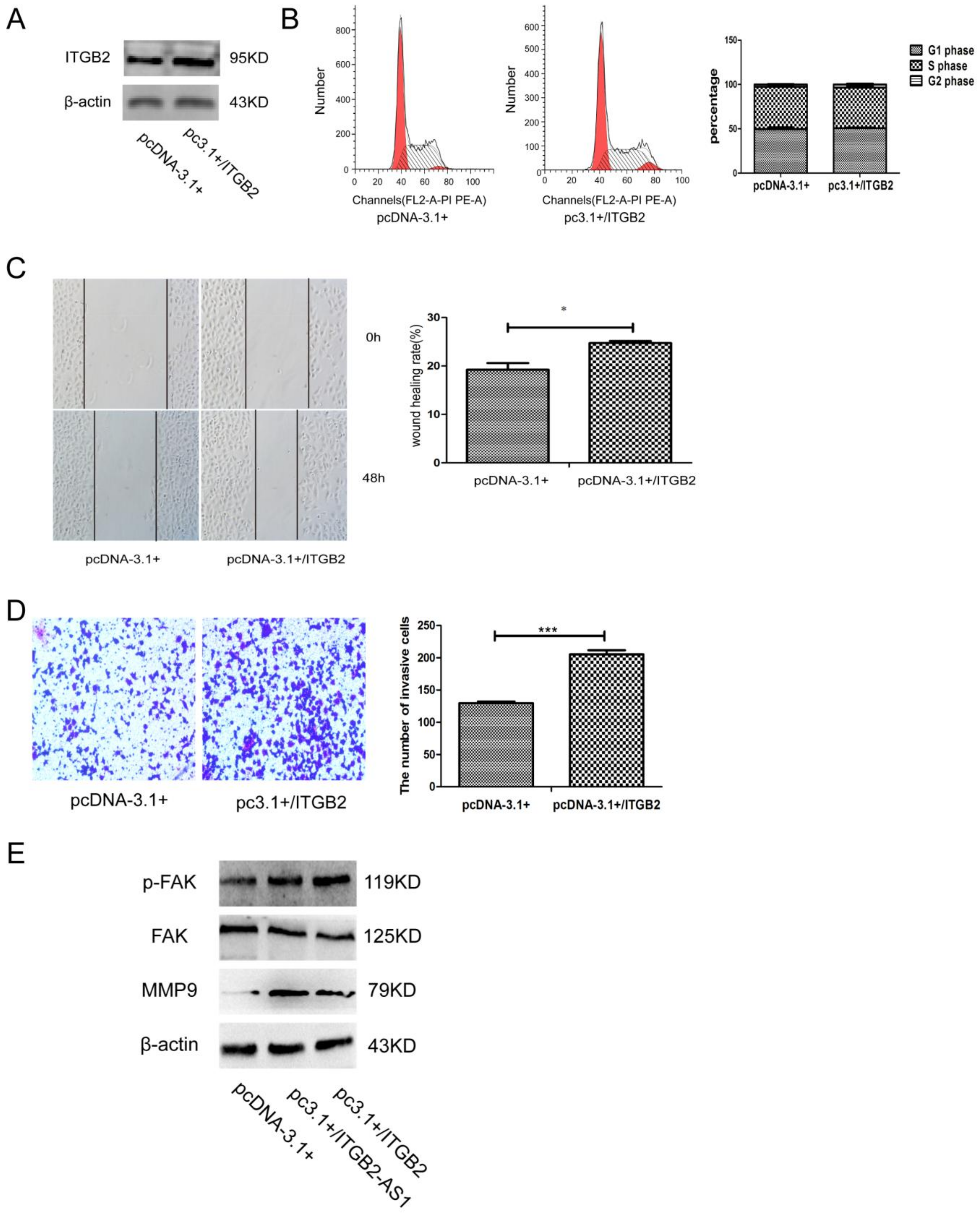
2.5. Overexpression of ITGB2 also Could Facilitate the Migration and Invasion of MCF-7 Cell
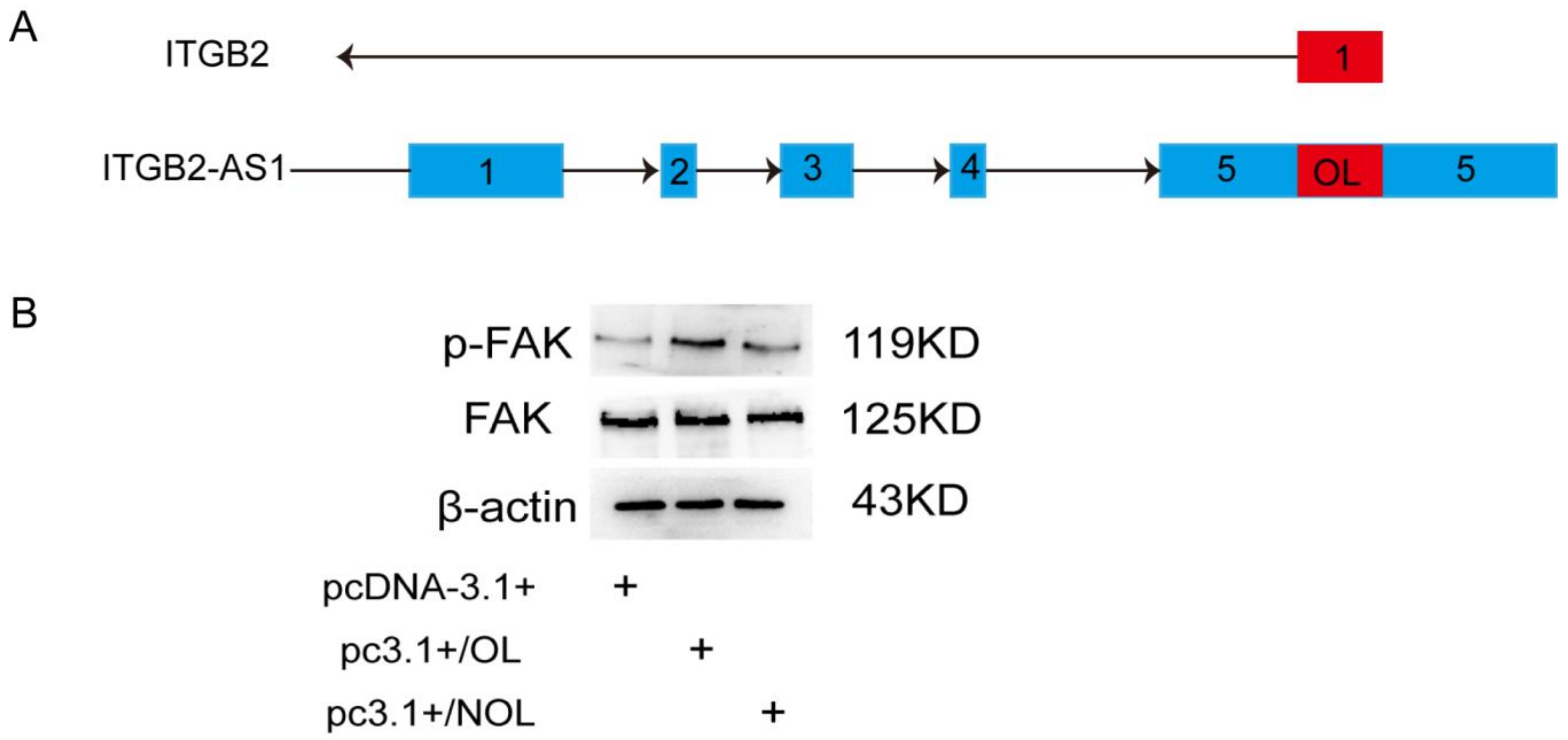
2.6. The OL Fragment Plays a Critical Role in the Promotion Effect of ITGB2-AS1 on MCF-7 Cell
3. Discussion
4. Materials and Methods
4.1. Cell Line and Cell Culture
4.2. Online Database
4.3. RNA Isolation
4.4. Plasmid Transfection
4.5. siRNA Transfection
4.6. Western Blotting
4.7. MTT Assay
4.8. Flow Cytometry
4.9. Wound Healing Assay
4.10. Transwell Chamber Assay
4.11. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- GBD2015 Mortality and Causes of Death Collaborators. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980–2015: A systematicanalysis for the global burden of disease study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar]
- Rossi, S.; Schinzari, G.; Basso, M.; Strippoli, A.; Dadduzio, V.; D’Argento, E.; Cassano, A.; Barone, C. Maintenance hormonal and chemotherapy treatment in metastatic breast cancer: A systematic review. Future Oncol. 2016, 12, 1299–1307. [Google Scholar] [CrossRef] [PubMed]
- Fitzmaurice, C.; Allen, C.; Barber, R.M.; Barregard, L.; Bhutta, Z.A.; Brenner, H.; Dicker, D.J.; Chimed-Orchir, O.; Dandona, R.; Dandona, L.; et al. Global, regional, and national cancer incidence, mortality, years of life lost, years lived with disability, and disability-adjusted life-years for 32 cancer groups, 1990 to 2015: A systematic analysis for the global burden of disease study. JAMA Oncol. 2017, 3, 524–548. [Google Scholar] [PubMed]
- Fan, L.; Zheng, Y.; Yu, K.D.; Liu, G.Y.; Wu, J.; Lu, J.S.; Shen, K.W.; Shen, Z.Z.; Shao, Z.M. Breast cancer in a transitional society over 18 years: Trends and present status in Shanghai, China. Breast Cancer Res. Treat. 2009, 117, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Sun, S.; Yuan, J.P.; Wang, Y.H.; Cao, T.Z.; Zheng, H.M.; Jiang, X.Q.; Gong, Y.P.; Tu, Y.; Yao, F.; et al. Characteristics of breast cancer in Central China, literature review and comparison with USA. Breast 2016, 30, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Strasser-Weippl, K.; Li, J.-J.; Louis, J.S.; Finkelstein, D.M.; Yu, K.; Chen, W.; Shao, Z.; Goss, P.E. Breast cancer in China. Lancet Oncol. 2014, 15, e279–e289. [Google Scholar] [CrossRef]
- Quail, D.F.; Olson, O.C.; Bhardwaj, P.; Walsh, L.A.; Akkari, L.; Quick, M.L.; Chen, I.C.; Wendel, N.; Ben-Chetrit, N.; Walker, J.; et al. Obesity alters the lung myeloid cell landscape to enhance breast cancer metastasis through IL5 and GM-CSF. Nat. Cell Biol. 2017, 19, 974–987. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, N.M.; Zhou, X.K.; Gucalp, A.; Morris, P.G.; Howe, L.R.; Giri, D.D.; Morrow, M.; Wang, H.; Pollak, M.; Jones, L.W.; et al. Systemic correlates of white adipose tissue inflammation in early-stage breast cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 2283–2289. [Google Scholar] [CrossRef] [PubMed]
- Pusztai, L.; Karn, T.; Safonov, A.; Abu-Khalaf, M.M.; Bianchini, G. New strategies in breast cancer: immunotherapy. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2016, 22, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Kang, Q.; Sun, M.H.; Cheng, H.; Peng, Y.; Montag, A.G.; Deyrup, A.T.; Jiang, W.; Luu, H.H.; Luo, J.; Szatkowski, J.P.; et al. Characterization of the distinct orthotopic bone-forming activity of 14 BMPs using recombinant adenovirus-mediated gene delivery. Gene Ther. 2004, 11, 1312–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Weng, Y.; Zhang, M.; Li, Y.; Fan, M.; Guo, Y.; Sun, Y.; Li, W.; Shi, Q. BMP9 inhibits the growth and migration of lung adenocarcinoma A549 cells in a bone marrow stromal cells derived microenvironment through the MAPK/ERK and NF-κB pathways. Oncol. Rep. 2016, 36, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.X.; Wang, D.X.; Wu, Q.X.; Ren, C.M.; Li, Y.; Chen, Q.Z.; Zeng, Y.H.; Shao, Y.; Yang, J.Q.; Bai, Y.; et al. BMP9/p38 MAPK is essential for the antiproliferative effect of resveratrol on human colon cancer. Oncol. Rep. 2016, 35, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, J.H.; Huang, J.G. Bone morphogenetic protein 9 facilitates osteocarcinoma cell apoptosis and inhibits in vivo tumor growth. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Garcíaálvaro, M.; Addante, A.; Roncero, C.; Fernández, M.; Fabregat, I.; Sánchez, A.; Herrera, B. BMP9-induced survival effect in liver tumor cells requires p38MAPK activation. Int. J. Mol. Sci. 2015, 16, 20431–20448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Feng, H.; Ren, W.; Sun, X.; Luo, J.; Tang, M.; Zhou, L.; Weng, Y.; He, T.C.; Zhang, Y. BMP9 inhibits the proliferation and invasiveness of breast cancer cells MDA-MB-231. J. Cancer Res. Clin. Oncol. 2011, 137, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Liu, Y.; Wan, S.; Fei, C.; Wang, W.; Chen, Y.; Zhang, Z.; Wang, T.; Wang, J.; Zhou, L.; et al. BMP9 inhibits proliferation and metastasis of HER2-positive SK-BR-3 breast cancer cells through ERK1/2 and PI3K/AKT pathways. PLoS ONE 2014, 9, e96816. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Sun, L.; Liu, Q.; Gong, C.; Yao, Y.; Lv, X.; Lin, L.; Yao, H.; Su, F.; Li, D.; et al. A cytoplasmic NF-κB interacting long noncoding RNA blocks IκB phosphorylation and suppresses breast cancer metastasis. Cancer Cell 2015, 27, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Niknafs, Y.S.; Han, S.; Teng, M.; Speers, C.; Zhang, C.; Wilder-Romans, K.; Iyer, M.K.; Pitchiaya, S.; Malik, R.; Hosono, Y.; et al. The lncRNA landscape of breast cancer reveals a role for DSCAM-AS1 in breast cancer progression. Nat. Commun. 2016, 7, 12791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogill, S.B.; Wang, L. Co-expression network analysis of human lncRNAs and cancer genes. Cancer Inform. 2014, 13 (Suppl. 5), 49–59. [Google Scholar] [CrossRef] [PubMed]
- cBioportal for Cancer Genomics. Available online: http://www.cbioportal.org/ (accessed on 1 June 2018).
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanric. Available online: http://ibl.mdanderson.org/tanric/_design/basic/index.html (accessed on 1 June 2018).
- Li, J.; Han, L.; Roebuck, P.; Diao, L.; Liu, L.; Yuan, Y.; Weinstein, J.N.; Liang, H. TANRIC: An interactive open platform to explore the function of lncRNAs in cancer. Cancer Res. 2015, 75, 3728–3737. [Google Scholar] [CrossRef] [PubMed]
- GEPIA (Gene Expression Profiling Interactive Analysis). Available online: http://gepia.cancer-pku.cn/ (accessed on 1 June 2018).
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2018, 13, 4429–4434. [Google Scholar] [CrossRef] [PubMed]
- Vennin, C.; Spruyt, N.; Dahmani, F.; Julien, S.; Bertucci, F.; Finetti, P.; Chassat, T.; Bourette, R.P.; Le Bourhis, X.; Adriaenssens, E. H19 non coding RNA-derived miR-675 enhances tumorigenesis and metastasis of breast cancer cells by downregulating c-Cbl and Cbl-b. Oncotarget 2015, 6, 29209–29223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Li, Y.; Li, Y.; Ren, K.; Li, X.; Han, X.; Wang, J. Long non-coding RNA H19 promotes the proliferation and invasion of breast cancer through upregulating DNMT1 expression by sponging miR-152. J. Biochem. Mol. Toxicol. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Zhou, N.; Watabe, K.; Lu, Z.; Wu, F.; Xu, M.; Mo, Y.Y. Long non-coding RNA UCA1 promotes breast tumor growth by suppression of p27 (Kip1). Cell Death Dis. 2014, 5, e1008. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Key Laboratory of Diagnostic Medicine Designated by the Chinese Ministry of Education, Chongqing Medical University, Chongqing 400000, China. Measured by Quantitative PCR. Unpublished work, 2018.
- Tufarelli, C.; Stanley, J.A.S.; Garrick, D.; Sharpe, J.A.; Ayyub, H.; Wood, W.G.; Higgs, D.R. Transcription of antisense RNA leading to gene silencing and methylation as a novel cause of human genetic disease. Nat. Genet. 2003, 34, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Gius, D.; Onyango, P.; Muldoon-Jacobs, K.; Karp, J.; Feinberg, A.P.; Cui, H. Epigenetic silencing of tumour suppressor gene p15 by its antisense RNA. Nature 2008, 451, 202–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, R.R.; Mondal, T.; Mohammad, F.; Enroth, S.; Redrup, L.; Komorowski, J.; Nagano, T.; Mancini-Dinardo, D.; Kanduri, C. Kcnq1ot1 antisense noncoding RNA mediates lineage-specific transcriptional silencing through chromatin-level regulation. Mol. Cell 2008, 32, 232–246. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Product Length (bp) |
|---|---|---|
| itgb2-as1 | Forward: AGGAGATGGAACGAGGAAA Reverse: TTAGTGGTCTGCGAAGGTG | 231 |
| itgb2 | Forward: TTCGGGTCCTTCGTGGACA Reverse: ACTGGTTGGAGTTGTTGGTCA | 148 |
| β-actin | Forward: CCACGAAACTACCTTCAACTCC Reverse: GTGATCTCCTTCTGCATCCTGT | 132 |
| siRNA | Sense Strand | Anti-Sense Strand |
|---|---|---|
| siNC | 5′-UUCUCCGAACGUGUCACGUTT-3′ | 5′-ACGUGACACGUUCGGAGAATT-3′ |
| siITGB2-AS1 | 5′-GCAGACCACUAAACCUCAUTT-3′ | 5′-AUGAGGUUUAGUGGUCUGCTT-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Gou, L.; Xia, J.; Wan, Q.; Jiang, Y.; Sun, S.; Tang, M.; He, T.; Zhang, Y. LncRNA ITGB2-AS1 Could Promote the Migration and Invasion of Breast Cancer Cells through Up-Regulating ITGB2. Int. J. Mol. Sci. 2018, 19, 1866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071866
Liu M, Gou L, Xia J, Wan Q, Jiang Y, Sun S, Tang M, He T, Zhang Y. LncRNA ITGB2-AS1 Could Promote the Migration and Invasion of Breast Cancer Cells through Up-Regulating ITGB2. International Journal of Molecular Sciences. 2018; 19(7):1866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071866
Chicago/Turabian StyleLiu, Mengyao, Liyao Gou, Jing Xia, Qun Wan, Yayun Jiang, Shilei Sun, Min Tang, Tongchuan He, and Yan Zhang. 2018. "LncRNA ITGB2-AS1 Could Promote the Migration and Invasion of Breast Cancer Cells through Up-Regulating ITGB2" International Journal of Molecular Sciences 19, no. 7: 1866. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071866