Postnatal Development and Distribution of Sympathetic Innervation in Mouse Skeletal Muscle

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
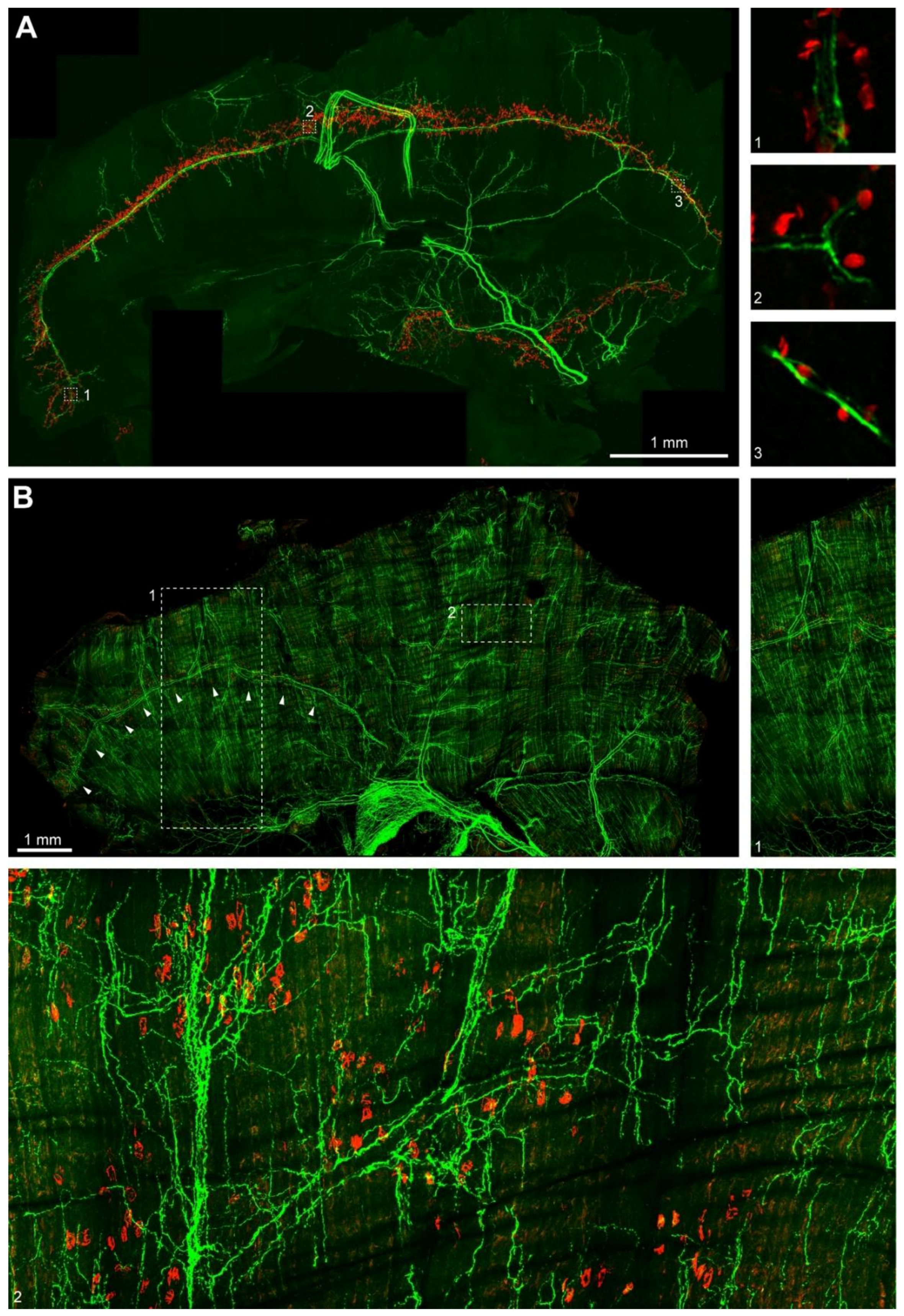
2.1. Sympathetic Innervation is Widely Distributed in Hindleg Muscles
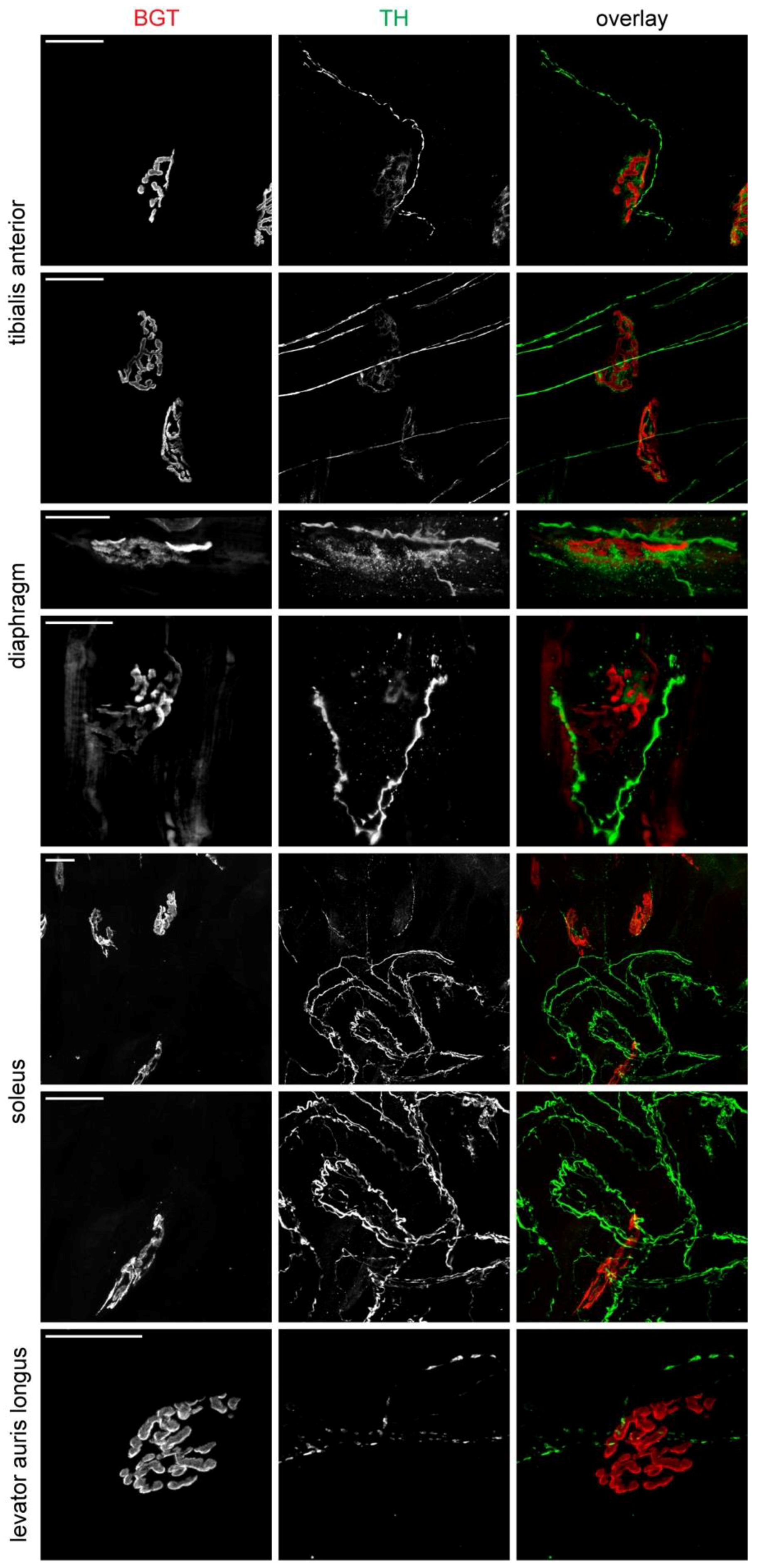
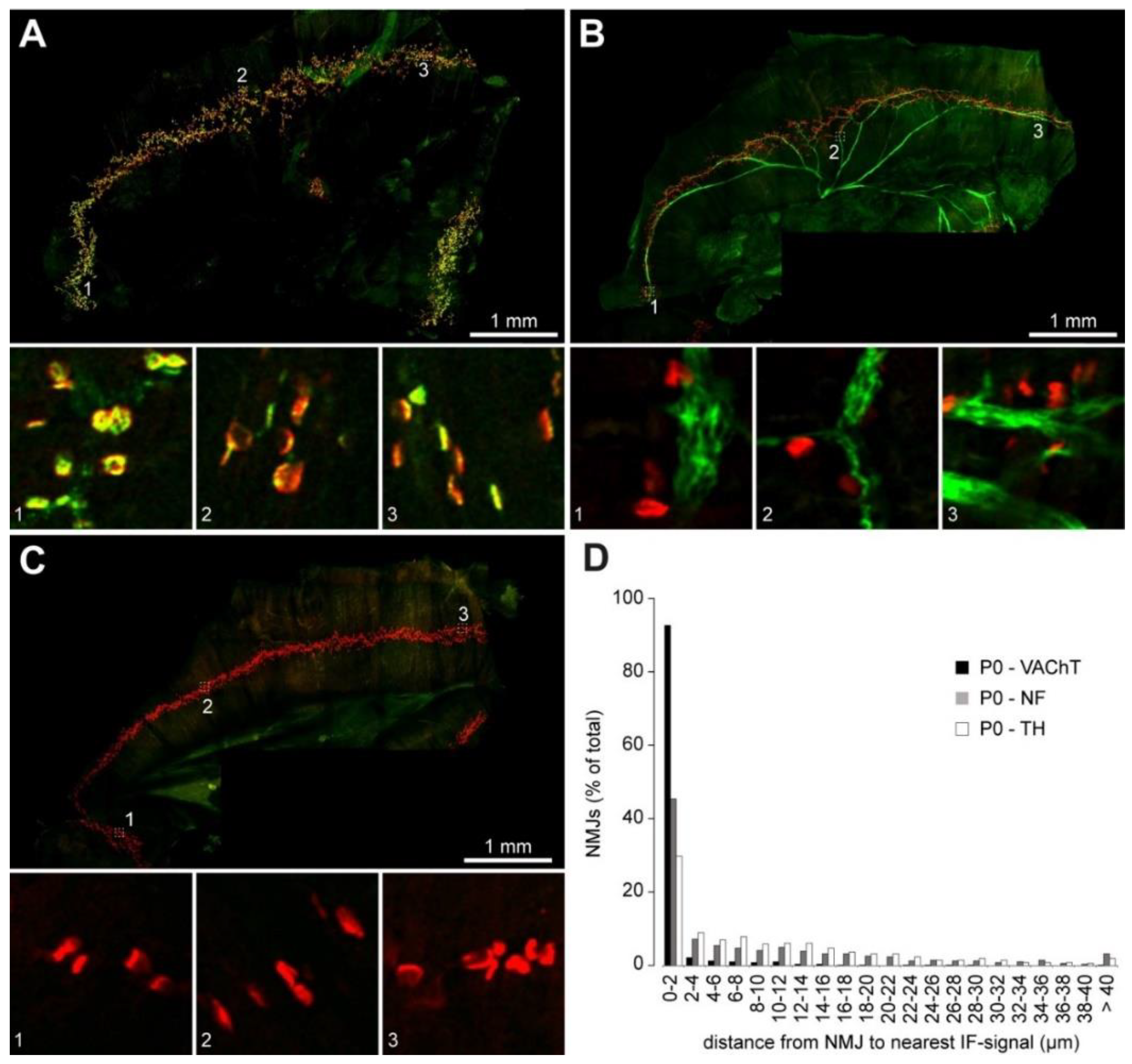
2.2. Sympathetic Innervation Contacts NMJs in Different Muscles
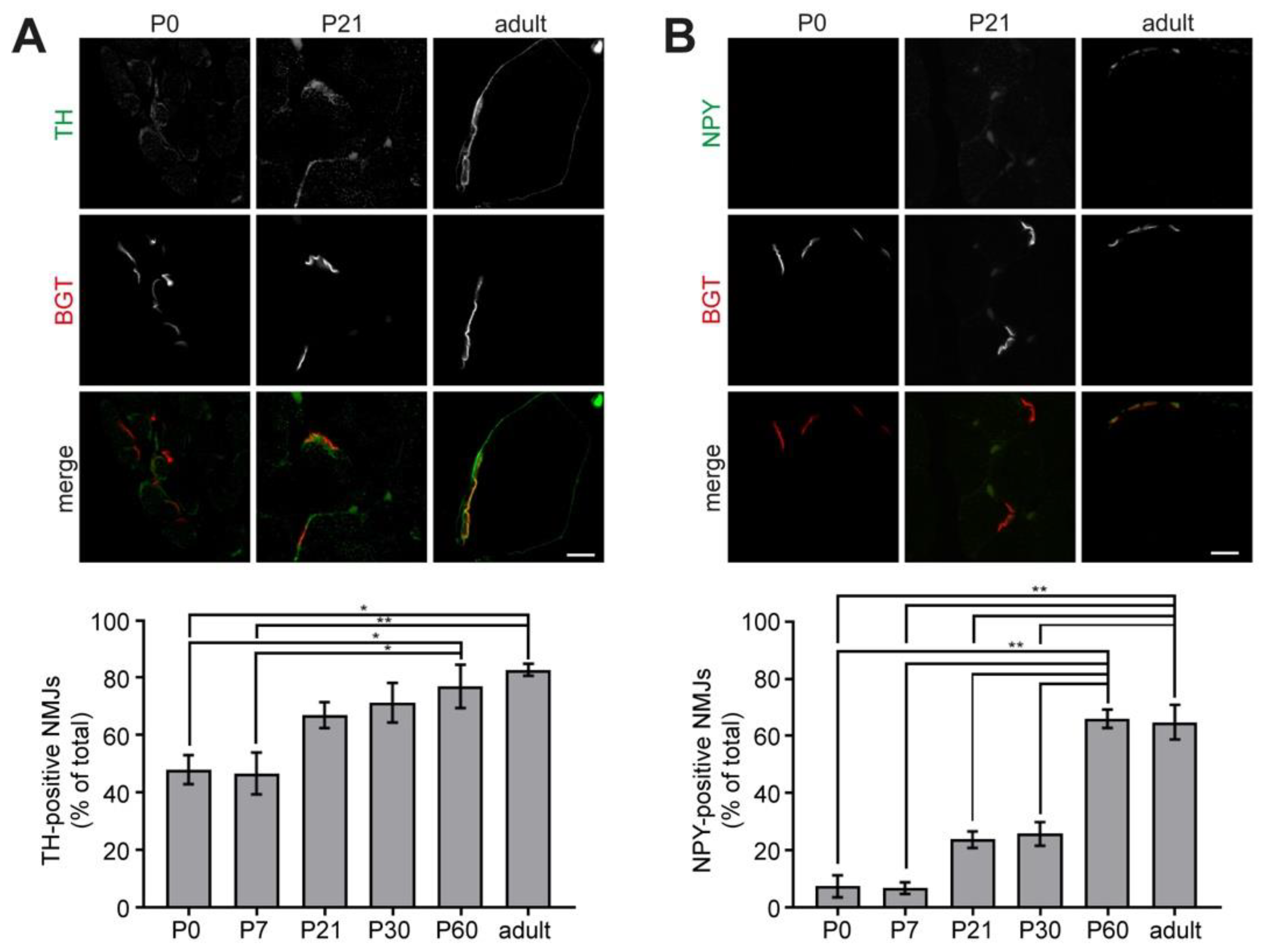
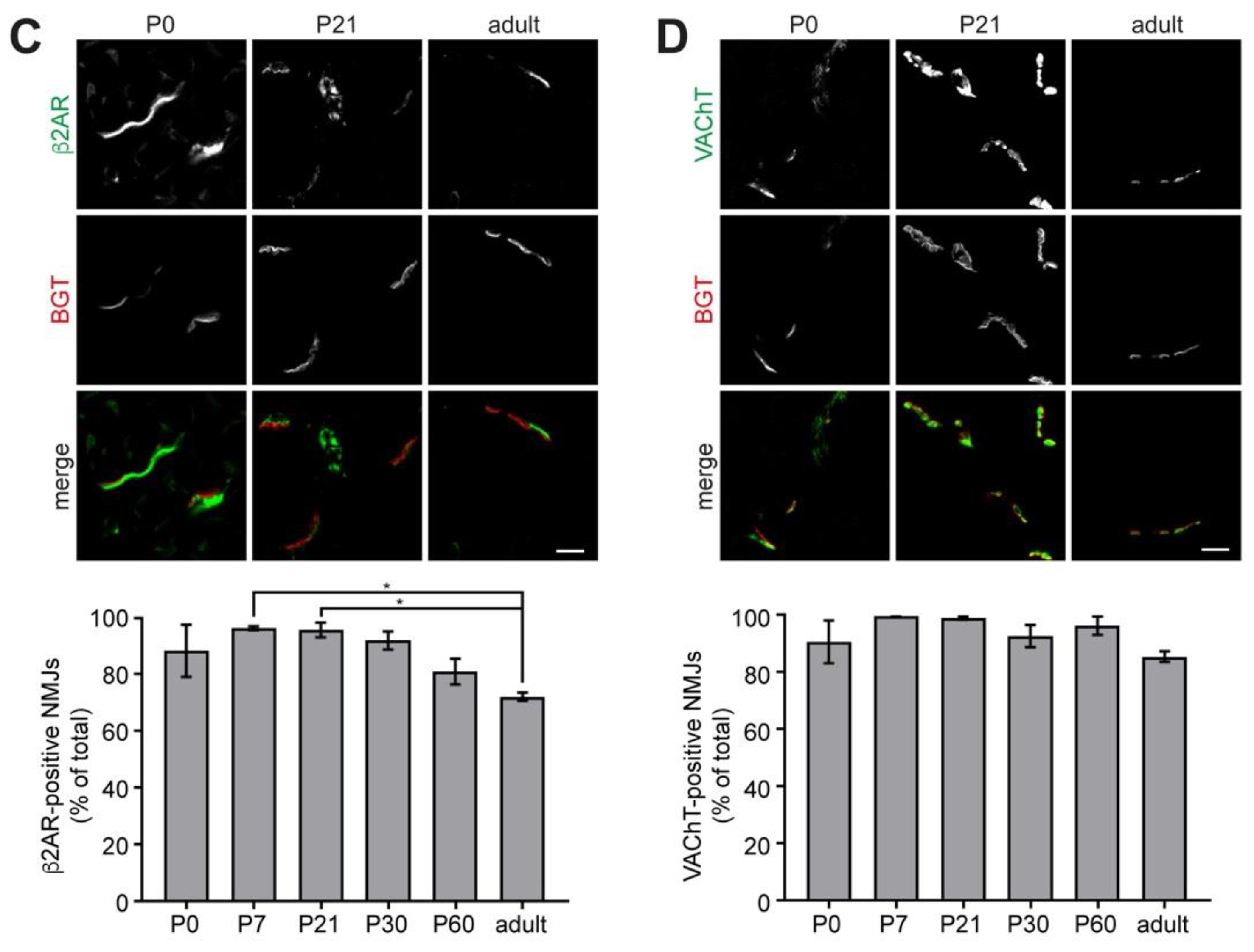
2.3. Sympathetic Innervation of NMJs Increases during Postnatal Development
2.4. Sympathetic Innervation in Mouse Diaphragm Aligns with the Synapse Band
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Preparation
4.2. Western Blot
4.3. Immunofluorescence
4.4. Microscopy
4.5. Image Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AChR | nicotinic acetylcholine receptor |
| BGT | α-bungarotoxin |
| BnP | blocking and permeabilization solution |
| β2AR | β2-adrenergic receptor |
| CMS | congenital myasthenic syndrome |
| GAPDH | glycerol aldehyde-3-phosphate-dehydrogenase |
| NMJ | neuromuscular junction |
| NPY | neuropeptide Y |
| PTwH | 0.2% Tween in PBS with 10 µg/mL heparin |
| TH | tyrosine hydroxylase |
| VAChT | vesicular acetylcholine transporter |
References
- Nicole, S.; Azuma, Y.; Bauché, S.; Eymard, B.; Lochmüller, H.; Slater, C. Congenital Myasthenic Syndromes or Inherited Disorders of Neuromuscular Transmission: Recent Discoveries and Open Questions. J. Neuromuscul. Dis. 2017, 4, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Beeson, D.; Palace, J. Therapeutic strategies for congenital myasthenic syndromes. Ann. N. Y. Acad. Sci. 2018, 1412, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, R.; Khan, M.M.; Lustrino, D.; Labeit, S.; Kettelhut, Í.C.; Navegantes, L.C.C. Alterations of cAMP-dependent signaling in dystrophic skeletal muscle. Front. Physiol. 2013, 4, 290. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Lustrino, D.; Silveira, W.A.; Wild, F.; Straka, T.; Issop, Y.; O’Connor, E.; Cox, D.; Reischl, M.; Marquardt, T.; et al. Sympathetic innervation controls homeostasis of neuromuscular junctions in health and disease. Proc. Natl. Acad. Sci. 2016, 113, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Boeke, J. Die motorische Endplatte bei den höheren Vertebraten, ihre Entwickelung, Form und Zusammenhang mit der Muskelfaser. Anat Anz 1909, 35, 240–256. [Google Scholar]
- Boeke, J. Ueber eine aus marklosen Fasern hervorgehende zweite Art von hypolemmalen Nervenendplatten bei den quergestreiften Muskelfasern der Vertebraten. Anat Anz 1909, 35, 481–484. [Google Scholar]
- Boeke, J. Die doppelte (motorische und sympathische) efferente Innervation der quergestreiften Muskelfasern. Anat Anz 1913, 44, 343–356. [Google Scholar]
- Hinsey, J.C. The Innervation of Skeletal Muscle. Physiol Rev 1934, 14, 514–585. [Google Scholar] [CrossRef]
- Barker, D.; Saito, M. Autonomic innervation of receptors and muscle fibres in cat skeletal muscle. Proc. R. Soc. Lond. B Biol. Sci. 1981, 212, 317–332. [Google Scholar] [CrossRef]
- Radovanovic, D.; Peikert, K.; Lindström, M.; Domellöf, F.P. Sympathetic innervation of human muscle spindles. J. Anat. 2015, 226, 542–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wörl, J.; Neuhuber, W.L. Enteric co-innervation of motor endplates in the esophagus: state of the art ten years after. Histochem. Cell Biol. 2005, 123, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Hempfling, C.; Seibold, R.; Shiina, T.; Heimler, W.; Neuhuber, W.L.; Wörl, J. Enteric co-innervation of esophageal striated muscle fibers: a phylogenetic study. Auton. Neurosci. 2009, 151, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Chan-Palay, V.; Engel, A.G.; Wu, J.Y.; Palay, S.L. Coexistence in human and primate neuromuscular junctions of enzymes synthesizing acetylcholine, catecholamine, taurine, and gamma-aminobutyric acid. Proc. Natl. Acad. Sci. USA 1982, 79, 7027–7030. [Google Scholar] [CrossRef] [PubMed]
- Chan-Palay, V.; Engel, A.G.; Palay, S.L.; Wu, J.Y. Synthesizing enzymes for four neuroactive substances in motor neurons and neuromuscular junctions: light and electron microscopic immunocytochemistry. Proc. Natl. Acad. Sci. USA 1982, 79, 6717–6721. [Google Scholar] [CrossRef] [PubMed]
- Silveira, W.A.; Gonçalves, D.A.; Graça, F.A.; Andrade-Lopes, A.L.; Bergantin, L.B.; Zanon, N.M.; Godinho, R.O.; Kettelhut, I.C.; Navegantes, L.C.C. Activating cAMP/PKA signaling in skeletal muscle suppresses the ubiquitin-proteasome-dependent proteolysis: implications for sympathetic regulation. J. Appl. Physiol. 2014, 117, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolf, R.; Deschenes, M.R.; Sandri, M. Neuromuscular junction degeneration in muscle wasting. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xiong, W.-C.; Mei, L. Neuromuscular Junction Formation, Aging, and Disorders. Annu. Rev. Physiol. 2018, 80, 159–188. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.; Araujo, I.; Araujo, A. Autonomic nervous system dysfunction in motor neuron diseases. J. Rare Dis. Res. Treat. 2018, 3, 1–5. [Google Scholar] [CrossRef]
- Sanes, J.R.; Lichtman, J.W. Development of the vertebrate neuromuscular junction. Annu. Rev. Neurosci. 1999, 22, 389–442. [Google Scholar] [CrossRef] [PubMed]
- Gilles, J.-F.; Dos Santos, M.; Boudier, T.; Bolte, S.; Heck, N. DiAna, an ImageJ tool for object-based 3D co-localization and distance analysis. Methods 2017, 115, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Gibbins, I.L.; Jobling, P.; Morris, J.L. Functional organization of peripheral vasomotor pathways. Acta Physiol. Scand. 2003, 177, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Hodges, G.J.; Jackson, D.N.; Mattar, L.; Johnson, J.M.; Shoemaker, J.K. Neuropeptide Y and neurovascular control in skeletal muscle and skin. Am. J. Physiol. Integr. Comp. Physiol. 2009, 297, R546–R555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sculptoreanu, A.; Scheuer, T.; Catterall, W.A. Voltage-dependent potentiation of L-type Ca2+ channels due to phosphorylation by cAMP-dependent protein kinase. Nature 1993, 364, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.P.; Dulhunty, A.F. Beta-adrenergic potentiation of E-C coupling increases force in rat skeletal muscle. Muscle Nerve 1993, 16, 1317–1325. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, R.; Magalhães, P.J.; Pozzan, T. Direct in vivo monitoring of sarcoplasmic reticulum Ca2+ and cytosolic cAMP dynamics in mouse skeletal muscle. J. Cell Biol. 2006, 173, 187–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peyronnard, J.M.; Charron, L.F.; Lavoie, J.; Messier, J.P. Motor, sympathetic and sensory innervation of rat skeletal muscles. Brain Res. 1986, 373, 288–302. [Google Scholar] [CrossRef]
- Tintignac, L.A.; Brenner, H.-R.; Rüegg, M.A. Mechanisms Regulating Neuromuscular Junction Development and Function and Causes of Muscle Wasting. Physiol. Rev. 2015, 95, 809–852. [Google Scholar] [CrossRef] [PubMed]
- Gilbey, M.P.; Spyer, K.M. Essential organization of the sympathetic nervous system. Best Pract. Res. Clin. Endoc. Metab. 1993, 7, 259–278. [Google Scholar] [CrossRef]
- Stuelsatz, P.; Keire, P.; Almuly, R.; Yablonka-Reuveni, Z. A Contemporary Atlas of the Mouse Diaphragm. J. Histochem. Cytochem. 2012, 60, 638–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, D.; Segal, S.S. Neurovascular Proximity in the Diaphragm Muscle of Adult Mice. Microcirculation 2012, 19, 306–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz, P.M.R.; Palace, J.; Beeson, D. Congenital myasthenic syndromes and the neuromuscular junction. Curr. Opin. Neurol. 2014, 27, 566–575. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.; Toniolo, L.; Nagaraj, N.; Ciciliot, S.; Vindigni, V.; Schiaffino, S.; Reggiani, C.; Mann, M. Single Muscle Fiber Proteomics Reveals Fiber-Type-Specific Features of Human Muscle Aging. Cell Rep. 2017, 19, 2396–2409. [Google Scholar] [CrossRef] [PubMed]
- Heuslein, J.L.; Murrell, K.P.; Leiphart, R.J.; Llewellyn, R.A.; Meisner, J.K.; Price, R.J. Vascular growth responses to chronic arterial occlusion are unaffected by myeloid specific focal adhesion kinase (FAK) deletion. Sci. Rep. 2016, 6, 27029. [Google Scholar] [CrossRef] [PubMed]
- Renier, N.; Wu, Z.; Simon, D.J.; Yang, J.; Ariel, P.; Tessier-Lavigne, M. iDISCO: A Simple, Rapid Method to Immunolabel Large Tissue Samples for Volume Imaging. Cell 2014, 159, 896–910. [Google Scholar] [CrossRef] [PubMed]
- Röder, I.V.; Strack, S.; Reischl, M.; Dahley, O.; Khan, M.M.; Kassel, O.; Zaccolo, M.; Rudolf, R. Participation of myosin Va and Pka type I in the regeneration of neuromuscular junctions. PLoS ONE 2012, 7, e40860. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Straka, T.; Vita, V.; Prokshi, K.; Hörner, S.J.; Khan, M.M.; Pirazzini, M.; Williams, M.P.I.; Hafner, M.; Zaglia, T.; Rudolf, R. Postnatal Development and Distribution of Sympathetic Innervation in Mouse Skeletal Muscle. Int. J. Mol. Sci. 2018, 19, 1935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071935
Straka T, Vita V, Prokshi K, Hörner SJ, Khan MM, Pirazzini M, Williams MPI, Hafner M, Zaglia T, Rudolf R. Postnatal Development and Distribution of Sympathetic Innervation in Mouse Skeletal Muscle. International Journal of Molecular Sciences. 2018; 19(7):1935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071935
Chicago/Turabian StyleStraka, Tatjana, Veronica Vita, Kaltrina Prokshi, Sarah Janice Hörner, Muzamil Majid Khan, Marco Pirazzini, Marion Patrick Ivey Williams, Mathias Hafner, Tania Zaglia, and Rüdiger Rudolf. 2018. "Postnatal Development and Distribution of Sympathetic Innervation in Mouse Skeletal Muscle" International Journal of Molecular Sciences 19, no. 7: 1935. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071935