Respiratory Bordetella bronchiseptica Carriage is Associated with Broad Phenotypic Alterations of Peripheral CD4+CD25+ T Cells and Differentially Affects Immune Responses to Secondary Non-Infectious and Infectious Stimuli in Mice


Abstract
:1. Introduction
2. Results
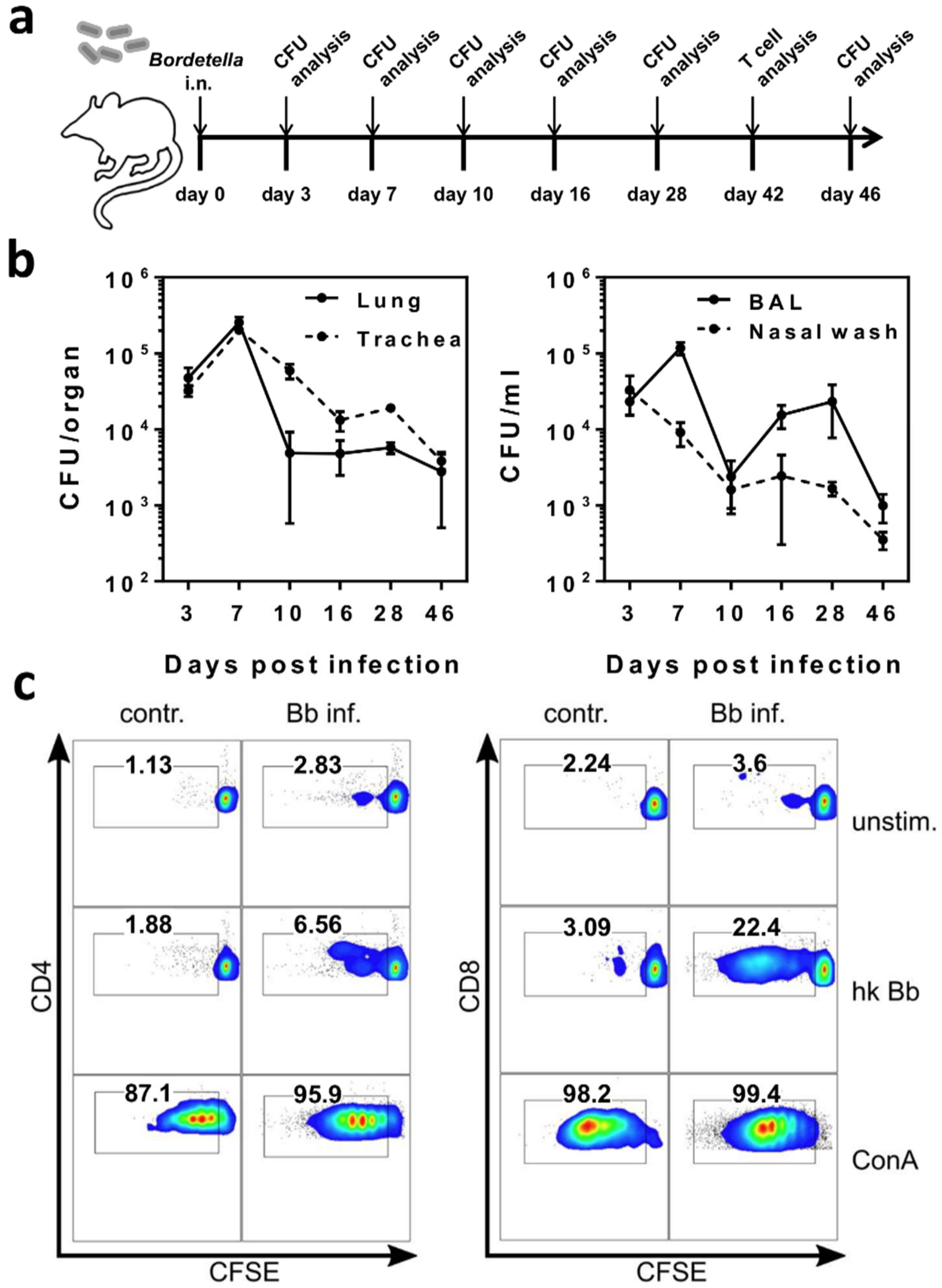
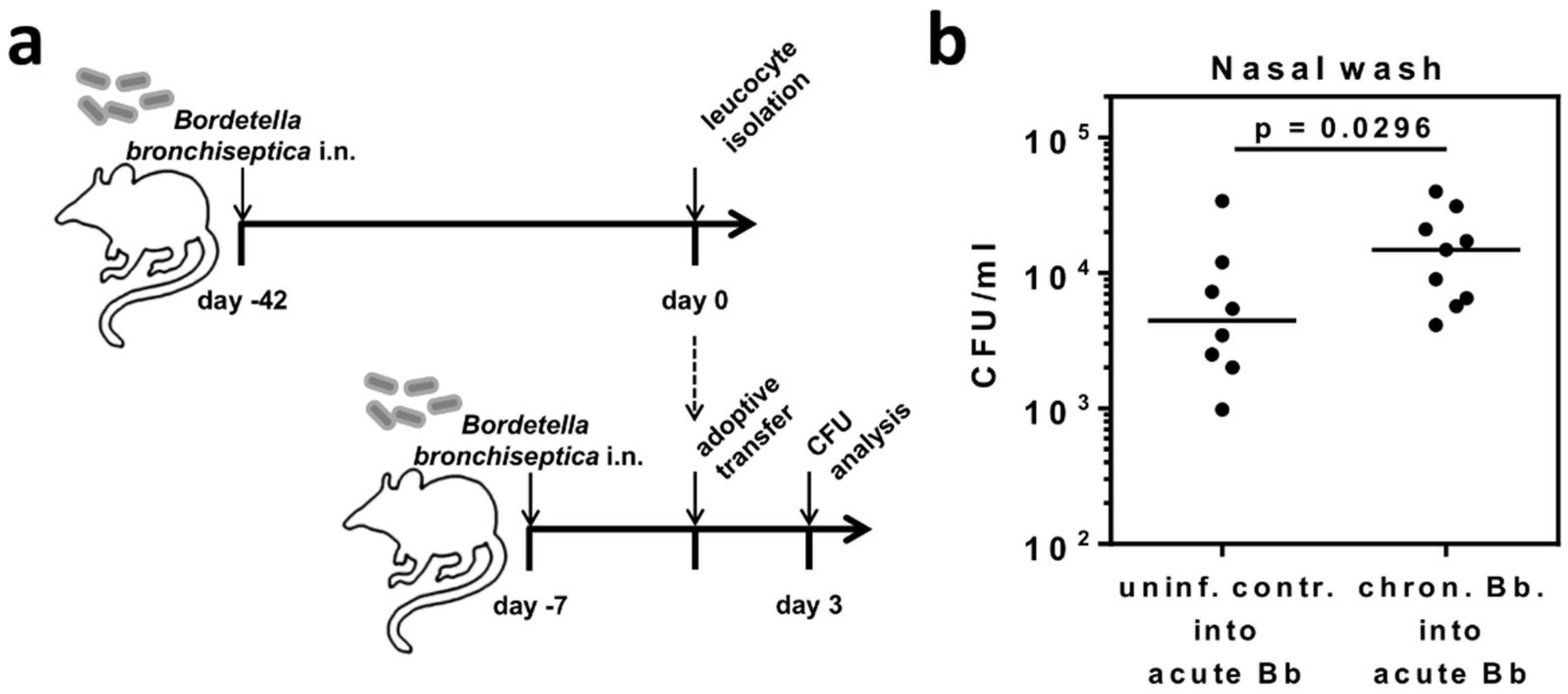
2.1. B. bronchiseptica Establishes Respiratory Tract Carriage Despite the Presence of Pathogen-Specific T Cells in Mice
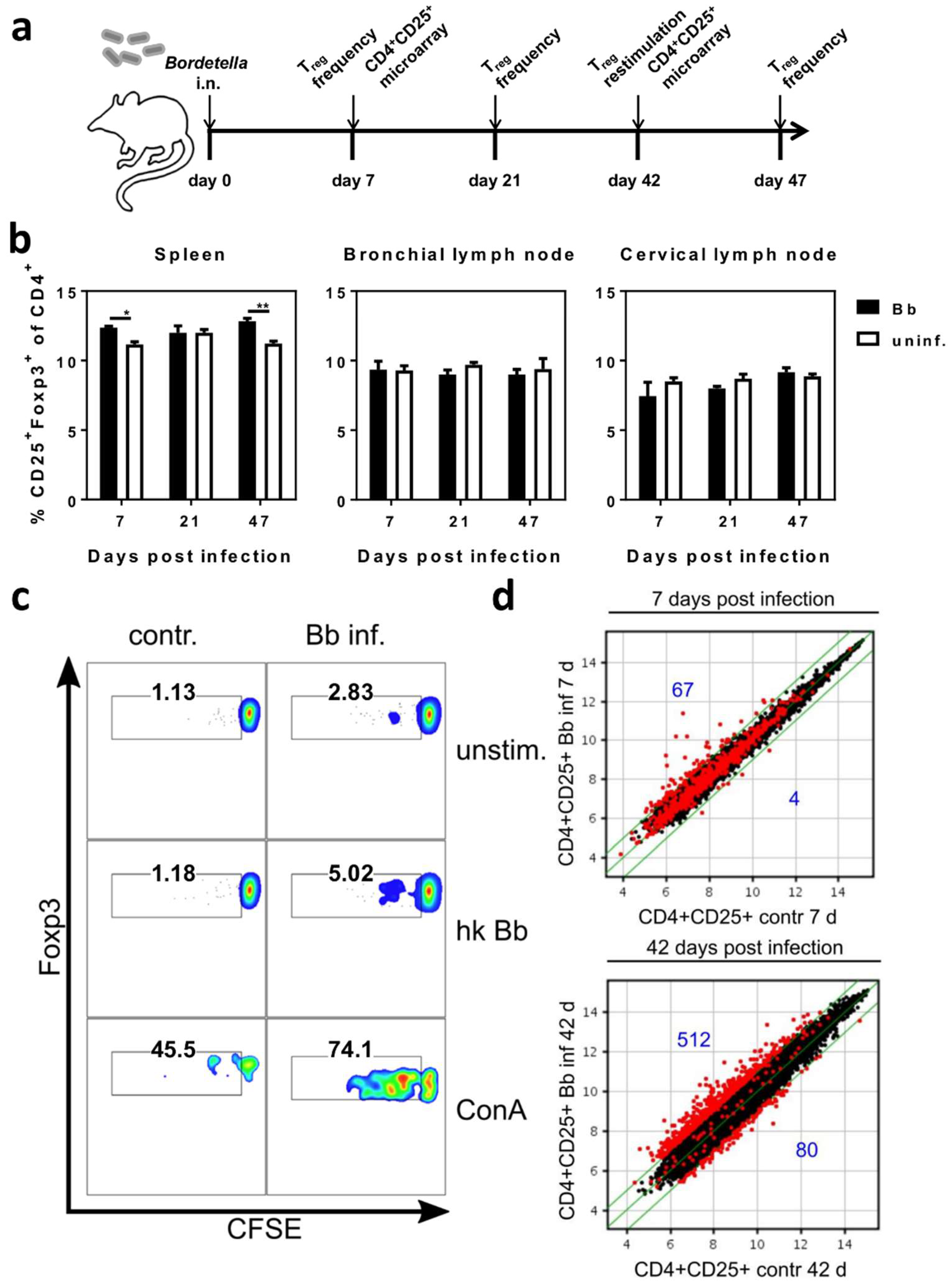
2.2. Respiratory B. bronchiseptica Carriage Is Associated with Broad Phenotypic Alterations of Peripheral CD4+CD25+ T Cells and the Induction of Pathogen-Specific Regulatory T Cells
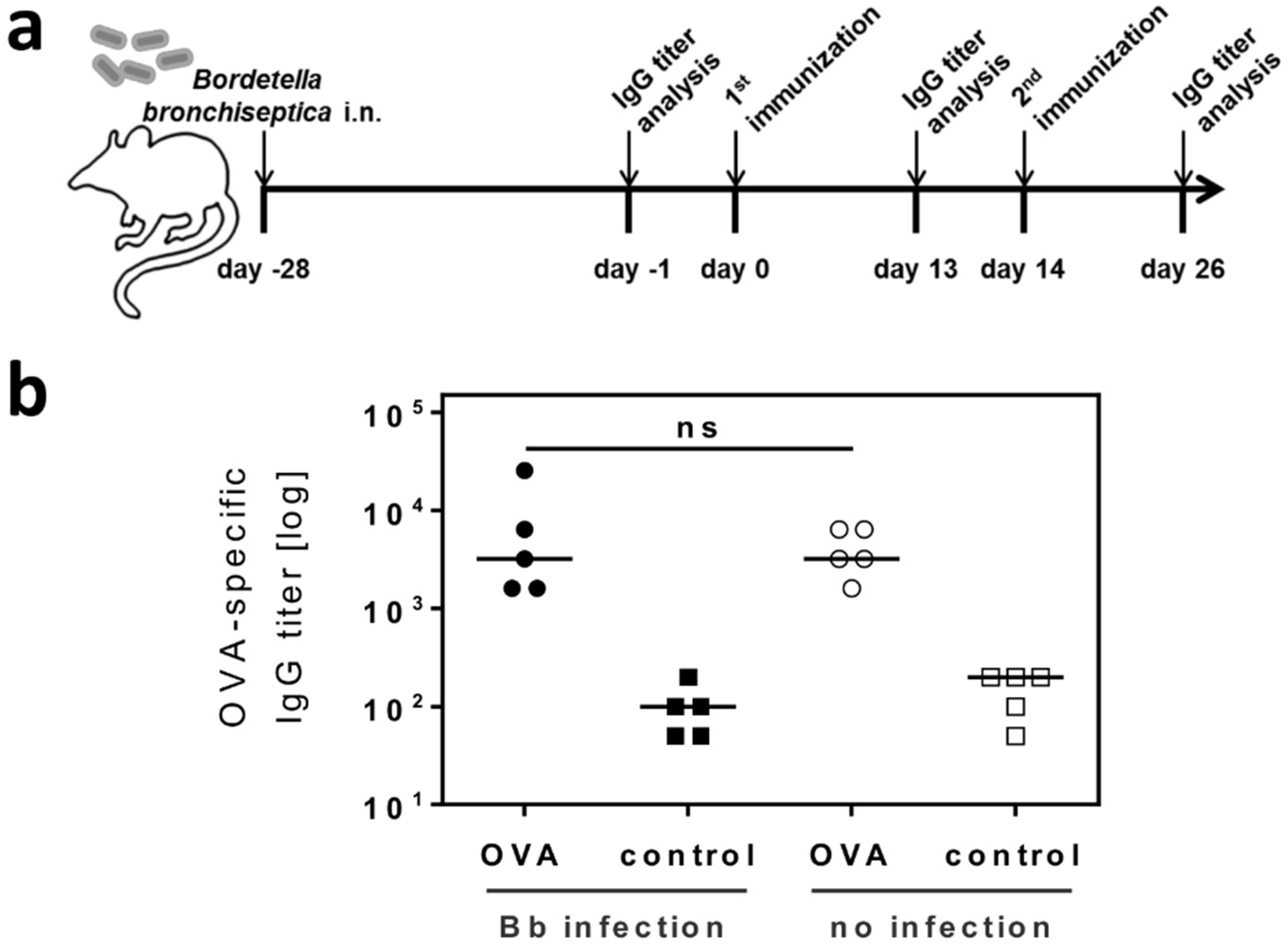
2.3. Respiratory B. bronchiseptica Carriage Does Not Alter the Humoral Immune Response to Vaccination
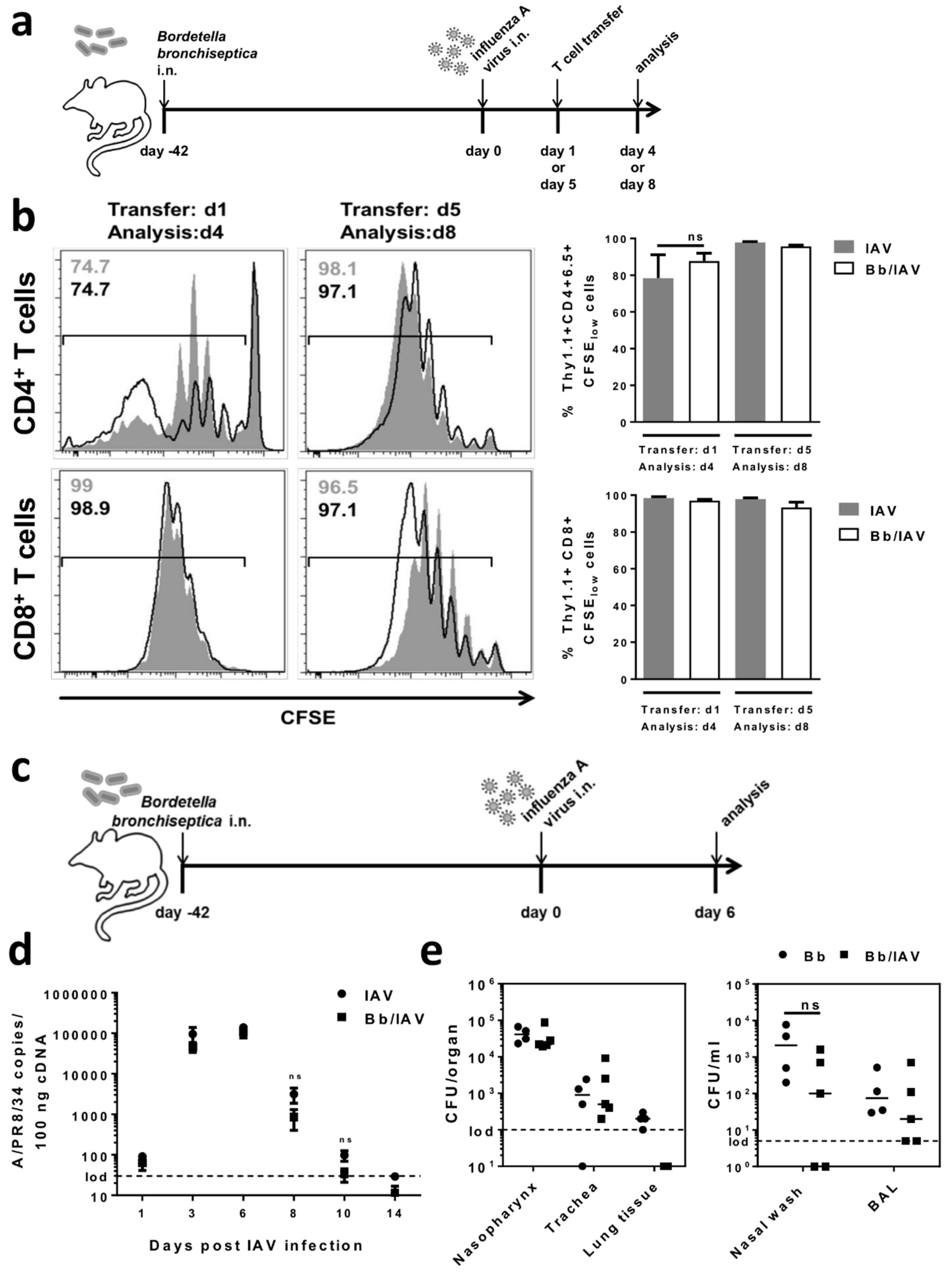
2.4. Respiratory B. bronchiseptica Carriage Does Not Affect the Local Antiviral Immune Response Following Secondary Infection with Influenza A Virus
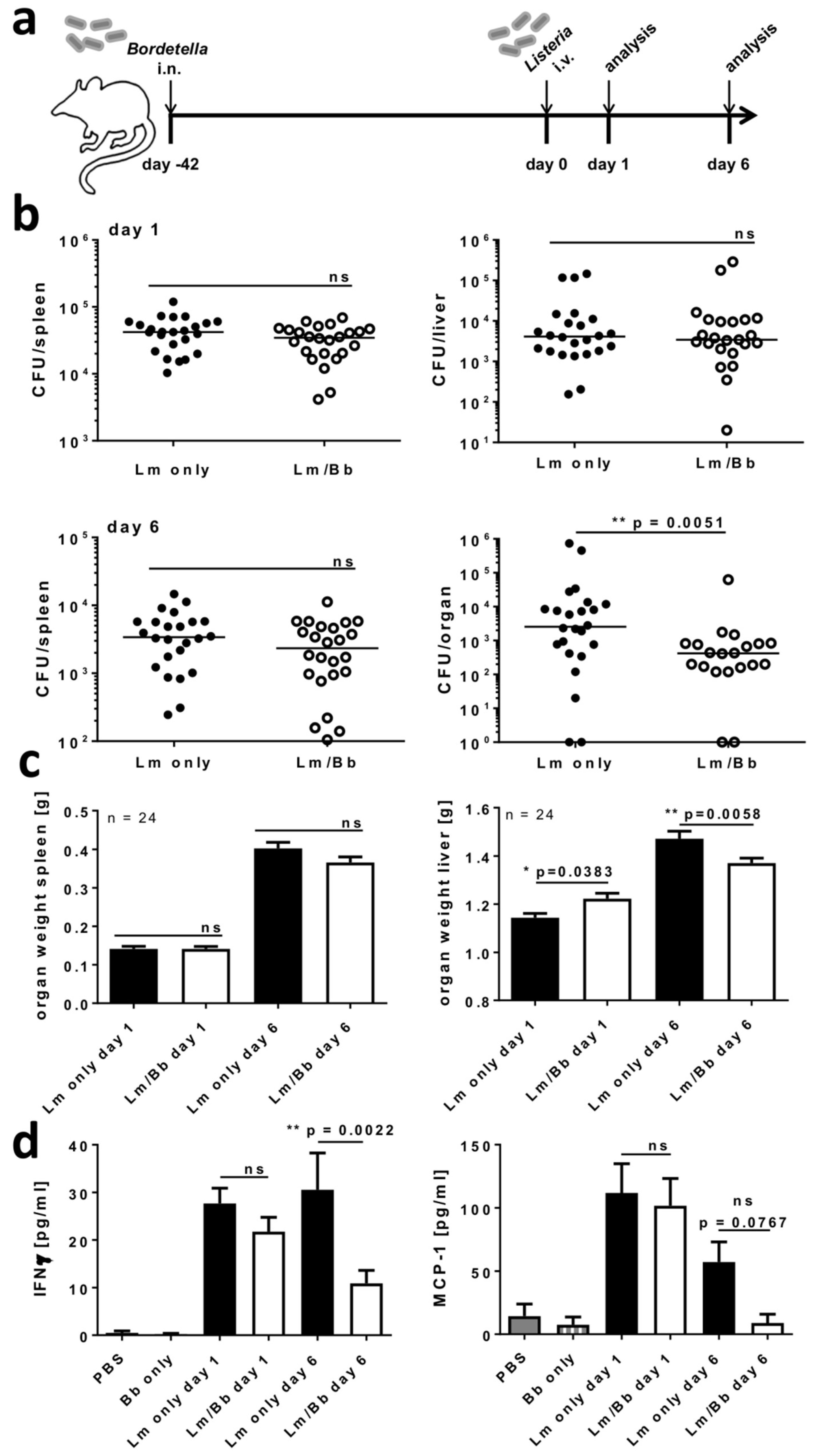
2.5. Respiratory B. bronchiseptica Carriage Alters Immunity to Systemic L. monocytogenes Infection
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Bacterial and Viral Pathogens
4.3. Bacterial and Viral Infections
4.4. Quantification of Pathogen Loads Following Infection
4.5. Restimulation of Splenocytes with Heat-Killed B. bronchiseptica
4.6. Flow Cytometry
4.7. Adoptive Transfer of Leukocytes Isolated from B. bronchiseptica Carriers
4.8. Microarray Analyses of Sorted CD4+CD25+ T Cells
4.9. Immunization with the Model Antigen OVA
4.10. Detection of OVA-Specific Serum IgG
4.11. Adoptive Transfer of IAV-Specific CD4+ and CD8+ T Cells
4.12. Quantification of IFN-γ and MCP-1 in Serum
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| DOAJ | Directory of open access journals |
References
- Dickson, R.P.; Erb-Downward, J.R.; Martinez, F.J.; Huffnagle, G.B. The microbiome and the respiratory tract. Annu. Rev. Physiol. 2016, 78, 481–504. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.G.; Segal, L.N. Lung microbiota and its impact on the mucosal immune phenotype. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Kolls, J.K. T cell-mediated host immune defenses in the lung. Annu. Rev. Immunol. 2013, 31, 605–633. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Croft, M. Control of regulatory T cells and airway tolerance by lung macrophages and dendritic cells. Ann. Am. Thorac. Soc. 2014, 11 (Suppl. 5), S306–S313. [Google Scholar] [CrossRef] [PubMed]
- Hasenberg, M.; Stegemann-Koniszewski, S.; Gunzer, M. Cellular immune reactions in the lung. Immunol. Rev. 2013, 251, 189–214. [Google Scholar] [CrossRef] [PubMed]
- Weitnauer, M.; Mijosek, V.; Dalpke, A.H. Control of local immunity by airway epithelial cells. Mucosal Immunol. 2016, 9, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Garbi, N.; Lambrecht, B.N. Location, function, and ontogeny of pulmonary macrophages during the steady state. Pflugers Arch. 2017, 469, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Kolahian, S.; Oz, H.H.; Zhou, B.; Griessinger, C.M.; Rieber, N.; Hartl, D. The emerging role of myeloid-derived suppressor cells in lung diseases. Eur. Respir. J. 2016, 47, 967–977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garib, F.Y.; Rizopulu, A.P. T-regulatory cells as part of strategy of immune evasion by pathogens. Biochemistry (Mosc.) 2015, 80, 957–971. [Google Scholar] [CrossRef] [PubMed]
- Cyktor, J.C.; Turner, J. Interleukin-10 and immunity against prokaryotic and eukaryotic intracellular pathogens. Infect. Immun. 2011, 79, 2964–2973. [Google Scholar] [CrossRef] [PubMed]
- Levast, B.; Li, Z.; Madrenas, J. The role of IL-10 in microbiome-associated immune modulation and disease tolerance. Cytokine 2015, 75, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, H.; Hermansen, M.N.; Buchvald, F.; Loland, L.; Halkjaer, L.B.; Bonnelykke, K.; Brasholt, M.; Heltberg, A.; Vissing, N.H.; Thorsen, S.V.; et al. Childhood asthma after bacterial colonization of the airway in neonates. N. Engl. J. Med. 2007, 357, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.L.; Wesselingh, S.; Rogers, G.B. Host-microbiome interactions in acute and chronic respiratory infections. Cell. Microbiol. 2016, 18, 652–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Linstow, M.L.; Schonning, K.; Hoegh, A.M.; Sevelsted, A.; Vissing, N.H.; Bisgaard, H. Neonatal airway colonization is associated with troublesome lung symptoms in infants. Am. J. Respir. Crit. Care Med. 2013, 188, 1041–1042. [Google Scholar] [CrossRef] [PubMed]
- McAleer, J.P.; Kolls, J.K. Contributions of the intestinal microbiome in lung immunity. Eur. J. Immunol. 2018, 48, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, D.R.; Welsh, D.A.; Shellito, J.E. Regulation of lung immunity and host defense by the intestinal microbiota. Front. Microbiol. 2015, 6, 1085. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Chiang, C.H.; Chuang, C.H.; Liu, S.L.; Jheng, Y.H.; Ryu, J.H. Spillover of cytokines and reactive oxygen species in ventilator-induced lung injury associated with inflammation and apoptosis in distal organs. Respir. Care 2014, 59, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Rahman Fink, N.; Chawes, B.L.; Thorsen, J.; Stokholm, J.; Krogfelt, K.; Schjorring, S.; Kragh, M.; Bonnelykke, K.; Brix, S.; et al. Neonates colonized with pathogenic bacteria in the airways have a low-grade systemic inflammation. Allergy 2018, 6, 1085. [Google Scholar] [CrossRef] [PubMed]
- Teichert, T.; Vossoughi, M.; Vierkotter, A.; Sugiri, D.; Schikowski, T.; Hoffmann, B.; Schulte, T.; Roden, M.; Raulf-Heimsoth, M.; Luckhaus, C.; et al. Investigating the spill-over hypothesis: Analysis of the association between local inflammatory markers in sputum and systemic inflammatory mediators in plasma. Environ. Res. 2014, 134, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Yevsa, T.; Ebensen, T.; Fuchs, B.; Zygmunt, B.; Libanova, R.; Gross, R.; Schulze, K.; Guzman, C.A. Development and characterization of attenuated metabolic mutants of Bordetella bronchiseptica for applications in vaccinology. Environ. Microbiol. 2013, 15, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Melvin, J.A.; Scheller, E.V.; Miller, J.F.; Cotter, P.A. Bordetella pertussis pathogenesis: Current and future challenges. Nat. Rev. Microbiol. 2014, 12, 274–288. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.A.; Jones, A.M. Phosphorelay control of virulence gene expression in Bordetella. Trends Microbiol. 2003, 11, 367–373. [Google Scholar] [CrossRef]
- Trainor, E.A.; Nicholson, T.L.; Merkel, T.J. Bordetella pertussis transmission. Pathog. Dis. 2015, 73, ftv068. [Google Scholar] [CrossRef] [PubMed]
- Hewlett, E.L.; Burns, D.L.; Cotter, P.A.; Harvill, E.T.; Merkel, T.J.; Quinn, C.P.; Stibitz, E.S. Pertussis pathogenesis—What we know and what we don’t know. J. Infect. Dis. 2014, 209, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Banemann, A.; Gross, R. Phase variation affects long-term survival of Bordetella bronchiseptica in professional phagocytes. Infect. Immun. 1997, 65, 3469–3473. [Google Scholar] [PubMed]
- Forde, C.B.; Parton, R.; Coote, J.G. Bioluminescence as a reporter of intracellular survival of Bordetella bronchiseptica in murine phagocytes. Infect. Immun. 1998, 66, 3198–3207. [Google Scholar] [PubMed]
- Buboltz, A.M.; Nicholson, T.L.; Weyrich, L.S.; Harvill, E.T. Role of the type III secretion system in a hypervirulent lineage of Bordetella bronchiseptica. Infect. Immun. 2009, 77, 3969–3977. [Google Scholar] [CrossRef] [PubMed]
- Mann, P.; Goebel, E.; Barbarich, J.; Pilione, M.; Kennett, M.; Harvill, E. Use of a genetically defined double mutant strain of Bordetella bronchiseptica lacking adenylate cyclase and type III secretion as a live vaccine. Infect. Immun. 2007, 75, 3665–3772. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, L.S.; Rolin, O.Y.; Muse, S.J.; Park, J.; Spidale, N.; Kennett, M.J.; Hester, S.E.; Chen, C.; Dudley, E.G.; Harvill, E.T. A type VI secretion system encoding locus is required for Bordetella bronchiseptica immunomodulation and persistence in vivo. PLoS ONE 2012, 7, e45892. [Google Scholar] [CrossRef] [PubMed]
- Reissinger, A.; Skinner, J.A.; Yuk, M.H. Downregulation of mitogen-activated protein kinases by the Bordetella bronchiseptica Type III secretion system leads to attenuated nonclassical macrophage activation. Infect. Immun. 2005, 73, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, N.A.; Skinner, J.A.; Yuk, M.H. Bordetella bronchiseptica modulates macrophage phenotype leading to the inhibition of CD4+ T cell proliferation and the initiation of a Th17 immune response. J. Immunol. 2006, 177, 7131–7138. [Google Scholar] [CrossRef] [PubMed]
- Guzman, C.A.; Rohde, M.; Bock, M.; Timmis, K.N. Invasion and intracellular survival of Bordetella bronchiseptica in mouse dendritic cells. Infect. Immun. 1994, 62, 5528–5537. [Google Scholar] [PubMed]
- Gueirard, P.; Ave, P.; Balazuc, A.M.; Thiberge, S.; Huerre, M.; Milon, G.; Guiso, N. Bordetella bronchiseptica persists in the nasal cavities of mice and triggers early delivery of dendritic cells in the lymph nodes draining the lower and upper respiratory Tract. Infect. Immun. 2003, 71, 4137–4143. [Google Scholar] [CrossRef] [PubMed]
- Nagamatsu, K.; Kuwae, A.; Konaka, T.; Nagai, S.; Yoshida, S.; Eguchi, M.; Watanabe, M.; Mimuro, H.; Koyasu, S.; Abe, A. Bordetella evades the host immune system by inducing IL-10 through a type III effector, BopN. J. Exp. Med. 2009, 206, 3073–3088. [Google Scholar] [CrossRef] [PubMed]
- Pilione, M.R.; Harvill, E.T. The Bordetella bronchiseptica type III secretion system inhibits gamma interferon production that is required for efficient antibody-mediated bacterial clearance. Infect. Immun. 2006, 74, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Skinner, J.A.; Pilione, M.R.; Shen, H.; Harvill, E.T.; Yuk, M.H. Bordetella type III secretion modulates dendritic cell migration resulting in immunosuppression and bacterial persistence. J. Immunol. 2005, 175, 4647–4652. [Google Scholar] [CrossRef] [PubMed]
- Bruder, D.; Probst-Kepper, M.; Westendorf, A.M.; Geffers, R.; Beissert, S.; Loser, K.; von Boehmer, H.; Buer, J.; Hansen, W. Neuropilin-1: A surface marker of regulatory T cells. Eur. J. Immunol. 2004, 34, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Hansen, W.; Loser, K.; Westendorf, A.M.; Bruder, D.; Pfoertner, S.; Siewert, C.; Huehn, J.; Beissert, S.; Buer, J. G protein-coupled receptor 83 overexpression in naive CD4+CD25− T cells leads to the induction of Foxp3+ regulatory T cells in vivo. J. Immunol. 2006, 177, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Oida, T.; Hirota, K.; Nakamura, K.; Nomura, T.; Uchiyama, T.; Sakaguchi, S. Foxp3-dependent and -independent molecules specific for CD25+CD4+ natural regulatory T cells revealed by DNA microarray analysis. Int. Immunol. 2006, 18, 1197–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 control over Foxp3+ regulatory T cell function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.; Huehn, J.; de la Rosa, M.; Maszyna, F.; Kretschmer, U.; Krenn, V.; Brunner, M.; Scheffold, A.; Hamann, A. Expression of the integrin alpha Ebeta 7 identifies unique subsets of CD25+ as well as CD25− regulatory T cells. Proc. Natl. Acad. Sci. USA 2002, 99, 13031–13036. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Yu, H.; Dang, E.V.; Barbi, J.; Pan, X.; Grosso, J.F.; Jinasena, D.; Sharma, S.M.; McCadden, E.M.; Getnet, D.; et al. Eos mediates Foxp3-dependent gene silencing in CD4+ regulatory T cells. Science 2009, 325, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Hori, S.; Nomura, T.; Sakaguchi, S. Control of regulatory T cell development by the transcription factor Foxp3. Science 2003, 299, 1057–1061. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G. Microbiota influences vaccine and mucosal adjuvant efficacy. Immune Netw. 2017, 17, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Lynn, D.J.; Pulendran, B. The potential of the microbiota to influence vaccine responses. J. Leukoc. Biol. 2018, 103, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.N.; Himes, J.E.; Martinez, D.R.; Permar, S.R. The impact of the gut microbiota on humoral immunity to pathogens and vaccination in early infancy. PLoS Pathog. 2016, 12, e1005997. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. The influence of probiotics on vaccine responses—A systematic review. Vaccine 2018, 36, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Braciale, T.J.; Sun, J.; Kim, T.S. Regulating the adaptive immune response to respiratory virus infection. Nat. Rev. Immunol. 2012, 12, 295–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGill, J.; Heusel, J.W.; Legge, K.L. Innate immune control and regulation of influenza virus infections. J. Leukoc. Biol. 2009, 86, 803–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilder, M.S.; Sword, C.P. Mechanisms of pathogenesis in Listeria monocytogenes infection. II. Characterization of listeriosis in the CD-1 mouse and survey of biochemical lesions. J. Bacteriol. 1967, 93, 531–537. [Google Scholar] [PubMed]
- Calame, D.G.; Mueller-Ortiz, S.L.; Wetsel, R.A. Innate and adaptive immunologic functions of complement in the host response to Listeria monocytogenes infection. Immunobiology 2016, 221, 1407–1417. [Google Scholar] [CrossRef] [PubMed]
- McGuirk, P.; McCann, C.; Mills, K.H.G. Pathogen-specific T regulatory 1 cells induced in the respiratory tract by a bacterial molecule that stimulates interleukin 10 production by dendritic Cells. J. Exp. Med. 2002, 195, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Moreno, G.; Errea, A.; Van Maele, L.; Roberts, R.; Léger, H.; Sirard, J.C.; Benecke, A.; Rumbo, M.; Hozbor, D. Toll-like receptor 4 orchestrates neutrophil recruitment into airways during the first hours of Bordetella pertussis infection. Microbes Infect. 2013, 15, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Rolin, O.; Smallridge, W.; Henry, M.; Goodfield, L.; Place, D.; Harvill, E.T. Toll-like receptor 4 limits transmission of Bordetella bronchiseptica. PLoS ONE 2014, 9, e85229. [Google Scholar] [CrossRef] [PubMed]
- Zanin-Zhorov, A.; Tal-Lapidot, G.; Cahalon, L.; Cohen-Sfady, M.; Pevsner-Fischer, M.; Lider, O.; Cohen, I.R. Cutting edge: T cells respond to lipopolysaccharide innately via TLR4 signaling. J. Immunol. 2007, 179, 41–44. [Google Scholar] [CrossRef] [PubMed]
- McGuirk, P.; Mills, K.H. A regulatory role for interleukin 4 in differential inflammatory responses in the lung following infection of mice primed with Th1- or Th2-inducing pertussis vaccines. Infect. Immun. 2000, 68, 1383–1390. [Google Scholar] [CrossRef] [PubMed]
- Glinka, Y.; Prud’homme, G.J. Neuropilin-1 is a receptor for transforming growth factor beta-1, activates its latent form, and promotes regulatory T cell activity. J. Leukoc. Biol. 2008, 84, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Gerondakis, S.; Siebenlist, U. Roles of the NF-kappaB pathway in lymphocyte development and function. Cold Spring Harb. Perspect. Biol. 2010, 2, a000182. [Google Scholar] [CrossRef] [PubMed]
- Tiemessen, M.M.; Jagger, A.L.; Evans, H.G.; van Herwijnen, M.J.; John, S.; Taams, L.S. CD4+CD25+Foxp3+ regulatory T cells induce alternative activation of human monocytes/macrophages. Proc. Natl. Acad. Sci. USA 2007, 104, 19446–19451. [Google Scholar] [CrossRef] [PubMed]
- Przemska-Kosicka, A.; Childs, C.E.; Maidens, C.; Dong, H.; Todd, S.; Gosney, M.A.; Tuohy, K.M.; Yaqoob, P. Age-related changes in the natural killer cell response to seasonal influenza vaccination are not influenced by a synbiotic: A randomised controlled trial. Front. Immunol. 2018, 9, 591. [Google Scholar] [CrossRef] [PubMed]
- Betts, R.J.; Prabhu, N.; Ho, A.W.; Lew, F.C.; Hutchinson, P.E.; Rotzschke, O.; Macary, P.A.; Kemeny, D.M. Influenza A virus infection results in a robust, antigen-responsive, and widely disseminated Foxp3+ regulatory T cell response. J. Virol. 2012, 86, 2817–2825. [Google Scholar] [CrossRef] [PubMed]
- Vissing, N.H.; Chawes, B.L.; Bisgaard, H. Increased risk of pneumonia and bronchiolitis after bacterial colonization of the airways as neonates. Am. J. Respir. Crit. Care Med. 2013, 188, 1246–1252. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L. Prior infection with Bordetella bronchiseptica increases nasal colonization by Haemophilus parasuis in swine. Vet. Microbiol. 2004, 99, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Loving, C.L.; Brockmeier, S.L.; Vincent, A.L.; Palmer, M.V.; Sacco, R.E.; Nicholson, T.L. Influenza virus coinfection with Bordetella bronchiseptica enhances bacterial colonization and host responses exacerbating pulmonary lesions. Microb. Pathog. 2010, 49, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Basha, S.; Surendran, N.; Pichichero, M. Immune responses in neonates. Expert Rev. Clin. Immunol. 2014, 10, 1171–1184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, S.V. The lung microbiome and airway disease. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. 5), S462–S465. [Google Scholar] [CrossRef] [PubMed]
- Viegas, N.; Andzinski, L.; Wu, C.F.; Komoll, R.M.; Gekara, N.; Dittmar, K.E.; Weiss, S.; Jablonska, J. IFN-gamma production by CD27(+) NK cells exacerbates Listeria monocytogenes infection in mice by inhibiting granulocyte mobilization. Eur. J. Immunol. 2013, 43, 2626–2637. [Google Scholar] [CrossRef] [PubMed]
- Kirberg, J.; Baron, A.; Jakob, S.; Rolink, A.; Karjalainen, K.; von Boehmer, H. Thymic selection of CD8+ single positive cells with a class II major histocompatibility complex-restricted receptor. J. Exp. Med. 1994, 180, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, D.J.; Liblau, R.; Scott, B.; Fleck, S.; McDevitt, H.O.; Sarvetnick, N.; Lo, D.; Sherman, L.A. CD8(+) T cell-mediated spontaneous diabetes in neonatal mice. J. Immunol. 1996, 157, 978–983. [Google Scholar] [PubMed]
- Arico, B.; Rappuoli, R. Bordetella parapertussis and Bordetella bronchiseptica contain transcriptionally silent pertussis toxin genes. J. Bacteriol. 1987, 169, 2847–2853. [Google Scholar] [CrossRef] [PubMed]
- Stegemann, S.; Dahlberg, S.; Kroger, A.; Gereke, M.; Bruder, D.; Henriques-Normark, B.; Gunzer, M. Increased susceptibility for superinfection with Streptococcus pneumoniae during influenza virus infection is not caused by TLR7-mediated lymphopenia. PLoS ONE 2009, 4, e4840. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probe Set ID | Gene Symbol | Day 7 | Day 42 |
|---|---|---|---|
| 1438082_at | Tmem206 | 1.1 | 7.8 |
| 1437641_at | Rprd2 | −1.0 | 6.7 |
| 1433575_at | Sox4 | 1.6 | 6.4 |
| 1437668_at | Ccrl1 | 1.1 | 5.4 |
| 1436981_a_at | Ywhaz | −1.0 | 5.3 |
| 1452247_at | Fxr1 | −1.5 | 5.3 |
| 1421144_at | Rpgrip1 | 1.5 | 4.9 |
| 1418265_s_at | Irf2 | 1.2 | 4.7 |
| 1431037_a_at | Elavl1 | 1.2 | 4.5 |
| 1438908_at | Map3k12 | 1.1 | 4.4 |
| 1436567_a_at | Ndufa7 | 1.3 | 4.4 |
| 1419913_at | Strap | −1.1 | 4.3 |
| 1419918_at | Tmed7 | 1.2 | 4.3 |
| 1418199_at | Hemgn | 1.4 | 4.2 |
| 1426358_at | Taok1 | 1.3 | 4.2 |
| 1448011_at | Vps13c | −1.0 | 4.1 |
| 1419924_at | Fnip1 | 1.1 | 4.0 |
| 1422307_at | Sppl3 | −1.1 | 4.0 |
| 1420175_at | Tax1bp1 | −1.1 | 3.9 |
| 1422185_a_at | Cyb5r3 | −1.1 | 3.9 |
| 1454169_a_at | Epsti1 | 1.0 | 3.9 |
| 1438216_at | Rreb1 | −1.1 | 3.8 |
| 1431645_a_at | Gdi2 | −1.2 | 3.8 |
| 1429969_at | 4833403J16Rik | 1.0 | 3.8 |
| 1419866_s_at | Atxn2 | 1.0 | 3.8 |
| 1420535_a_at | Nub1 | −1.0 | 3.7 |
| 1437801_at | Morf4l1 | −1.0 | 3.7 |
| 1438009_at | Hist1h2ad | 1.6 | 3.6 |
| 1435749_at | Gda | 1.9 | 3.6 |
| 1439005_x_at | Ywhaz | −1.1 | 3.6 |
| Probe Set ID | Gene Symbol | CD25+ vs. CD25− | |||
|---|---|---|---|---|---|
| contr. | Bb inf. | ||||
| Day 7 | Day 42 | Day 7 | Day 42 | ||
| 1420692_at | Il2ra | 53.8 | 14.7 | 29.7 | 25.1 |
| 1420765_a_at | Foxp3 | 39.9 | 11.9 | 32.0 | 16.2 |
| 1439569_at | Gpr83 | 15.3 | 11.3 | 10.7 | 14.6 |
| 1429159_at | Itih5 | 16.5 | 7.0 | 8.3 | 13.9 |
| 1418507_s_at | Socs2 | 12.5 | 4.8 | 13.0 | 13.6 |
| 1447541_s_at | Itgae | 27.8 | 10.7 | 20.7 | 13.1 |
| 1434141_at | Gucy1a3 | 5.4 | 4.5 | 8.6 | 13.1 |
| 1457342_at | Ikzf4 | 10.5 | 9.9 | 8.5 | 13.0 |
| 1456956_at | Ikzf2 | 22.6 | 13.7 | 11.7 | 12.8 |
| 1419334_at | Ctla4 | 12.8 | 9.9 | 9.5 | 12.1 |
| 1448943_at | Nrp1 | 12.6 | 5.9 | 6.3 | 11.4 |
| 1429918_at | Arhgap20 | 9.2 | 8.8 | 9.0 | 10.9 |
| 1449216_at | Itgae | 16.1 | 8.4 | 15.0 | 10.5 |
| 1437542_at | Ikzf2 | 15.2 | 8.4 | 10.7 | 10.4 |
| 1423626_at | Dst | 9.0 | 7.4 | 5.9 | 9.4 |
| 1418084_at | Nrp1 | 11.8 | 5.6 | 8.2 | 8.8 |
| 1425145_at | Il1rl1 | 12.0 | 4.1 | 11.0 | 8.5 |
| 1448944_at | Nrp1 | 7.6 | 4.8 | 5.3 | 7.8 |
| 1426208_x_at | Plagl1 | 13.6 | 8.1 | 9.5 | 7.7 |
| 1438274_at | Ikzf4 | 8.0 | 7.5 | 6.3 | 7.7 |
| 1428329_a_at | Ift80 | 3.9 | 4.4 | 3.5 | 7.6 |
| 1428074_at | Tmem158 | 5.0 | 3.2 | 4.3 | 7.1 |
| 1449109_at | Socs2 | 7.9 | 4.2 | 8.4 | 6.8 |
| 1423415_at | Gpr83 | 8.3 | 11.6 | 8.0 | 6.8 |
| 1417601_at | Rgs1 | 2.7 | 4.9 | 3.0 | 6.4 |
| 1420788_at | Klrg1 | 13.2 | 5.3 | 10.0 | 6.4 |
| 1419219_at | Cyp4f18 | 12.2 | 10.3 | 8.6 | 6.2 |
| 1457198_at | Nrp1 | 7.7 | 6.7 | 6.1 | 5.9 |
| 1460469_at | Tnfrsf9 | 10.9 | 5.0 | 6.9 | 5.8 |
| 1418057_at | Tiam1 | 8.3 | 4.1 | 5.5 | 5.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeron, A.; Boehme, J.D.; Volckmar, J.; Gereke, M.; Yevsa, T.; Geffers, R.; Guzmán, C.A.; Schreiber, J.; Stegemann-Koniszewski, S.; Bruder, D. Respiratory Bordetella bronchiseptica Carriage is Associated with Broad Phenotypic Alterations of Peripheral CD4+CD25+ T Cells and Differentially Affects Immune Responses to Secondary Non-Infectious and Infectious Stimuli in Mice. Int. J. Mol. Sci. 2018, 19, 2602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092602
Jeron A, Boehme JD, Volckmar J, Gereke M, Yevsa T, Geffers R, Guzmán CA, Schreiber J, Stegemann-Koniszewski S, Bruder D. Respiratory Bordetella bronchiseptica Carriage is Associated with Broad Phenotypic Alterations of Peripheral CD4+CD25+ T Cells and Differentially Affects Immune Responses to Secondary Non-Infectious and Infectious Stimuli in Mice. International Journal of Molecular Sciences. 2018; 19(9):2602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092602
Chicago/Turabian StyleJeron, Andreas, Julia D. Boehme, Julia Volckmar, Marcus Gereke, Tetyana Yevsa, Robert Geffers, Carlos A. Guzmán, Jens Schreiber, Sabine Stegemann-Koniszewski, and Dunja Bruder. 2018. "Respiratory Bordetella bronchiseptica Carriage is Associated with Broad Phenotypic Alterations of Peripheral CD4+CD25+ T Cells and Differentially Affects Immune Responses to Secondary Non-Infectious and Infectious Stimuli in Mice" International Journal of Molecular Sciences 19, no. 9: 2602. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092602