Impact of X-ray Exposure on the Proliferation and Differentiation of Human Pre-Adipocytes

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
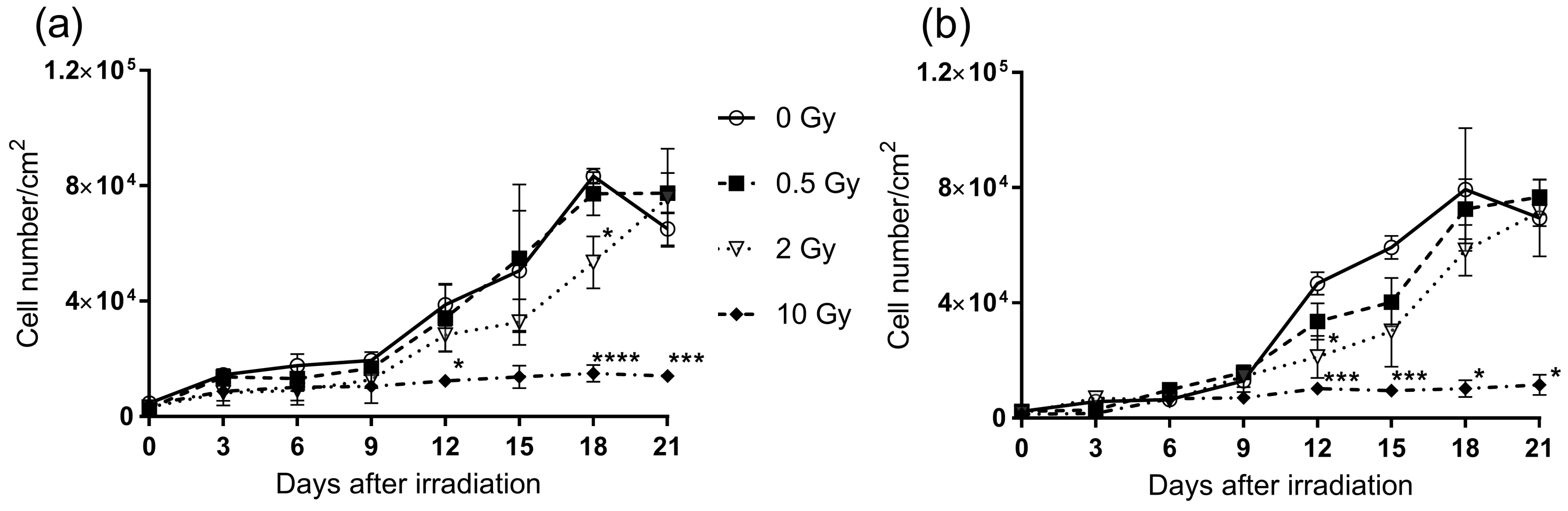
2.1. X-ray Irradiation Inhibits Proliferation and Clonogenic Survival of Pre-Adipocytes
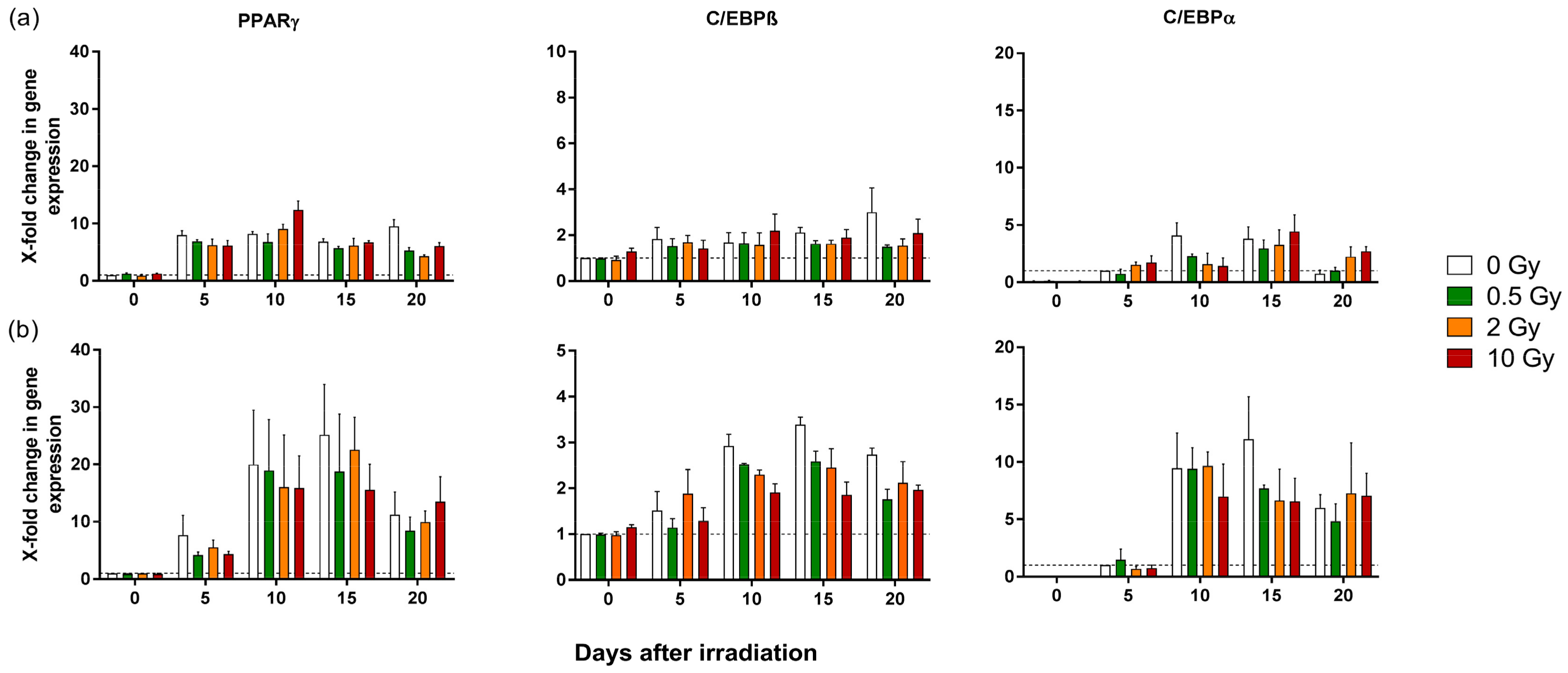
2.2. Differentiation Potential of Pre-Adipocytes is Mostly Unaffected by X-ray Irradiation
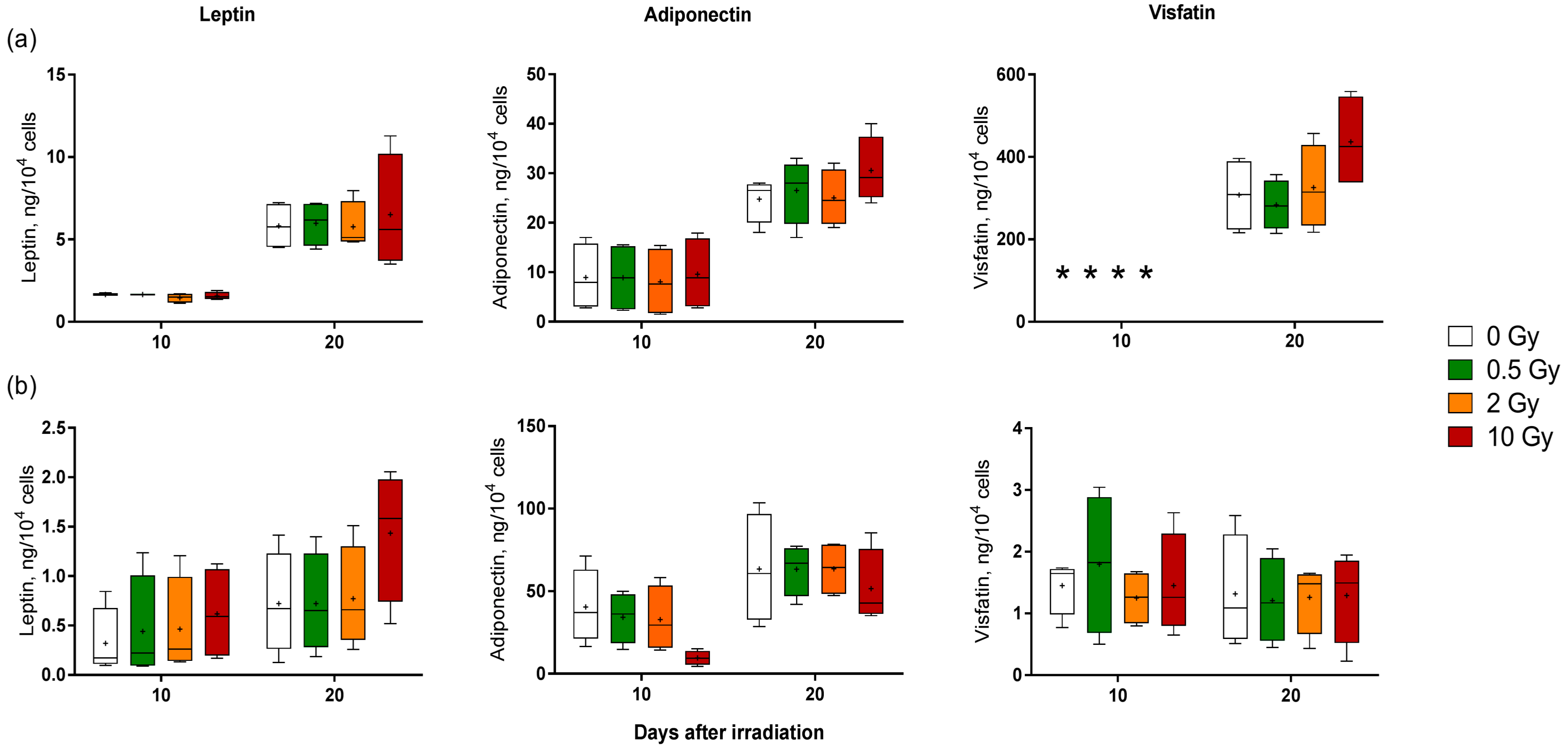
2.3. X-ray Irradiation Does Not Significantly Affect Adipokine Release by Human Adipocytes
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Irradiation Procedure
4.3. Crystal Violet Cell Proliferation Assay
4.4. Analysis of Adipocyte Differentiation
4.5. Quantification of Adipokine Release in Cell Culture Supernatants
4.6. Gene Expression Analysis During Differentiation
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| C/EBP | CCAAT/enhancer-binding proteins |
| CCL-2 | CC-chemokine ligand 2 |
| cDNA | complementary DNA |
| CFU | colony-forming unit |
| DAPI | 4′,6-Diamidin-2-phenylindol |
| ELISA | Enzyme-linked immunosorbent assay |
| IBMX | 3-Isobutyl-1-methylxanthin |
| IL | Interleukin |
| MMP-1 | Matrix metalloproteinase-1 |
| MSC | mesenchymal stem cells |
| MSD | musculoskeletal diseases |
| PPARγ | Perixosome proliferator-activated receptor gamma |
| qPCR | quantitative PCR |
| RA | rheumatoid arthritis |
| SGBS | Simpson–Golabi–Behmel Syndrome |
| WAT | white adipose tissue |
References
- Toussirot, É.; Grandclément, É.; Gaugler, B.; Michel, F.; Wendling, D.; Saas, P.; Dumoulin, G. Serum adipokines and adipose tissue distribution in rheumatoid arthritis and ankylosing spondylitis. A comparative study. Front. Immunol. 2013, 4, 453. [Google Scholar] [CrossRef] [PubMed]
- Schäffler, A.; Müller-Ladner, U.; Schölmerich, J.; Büchler, C. Role of adipose tissue as an inflammatory organ in human diseases. Endocr. Rev. 2006, 27, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Ràfols, M.E. Adipose tissue: Cell heterogeneity and functional diversity. Endocrinol. Nutr. 2014, 61, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Ioan-Facsinay, A.; Kloppenburg, M. An emerging player in knee osteoarthritis: The infrapatellar fat pad. Arthritis Res. Ther. 2013, 15, 225. [Google Scholar] [CrossRef] [PubMed]
- Caspar-Bauguil, S.; Cousin, B.; Galinier, A.; Segafredo, C.; Nibbelink, M.; André, M.; Casteilla, L.; Pénicaud, L. Adipose tissues as an ancestral immune organ: Site-specific change in obesity. FEBS Lett. 2005, 579, 3487–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divella, R.; de Luca, R.; Abbate, I.; Naglieri, E.; Daniele, A. Obesity and cancer: The role of adipose tissue and adipo-cytokines-induced chronic inflammation. J. Cancer 2016, 7, 2346–2359. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, J.M.; Stern, J.H.; Scherer, P.E. The cell biology of fat expansion. J. Cell Biol. 2015, 208, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Fasshauer, M.; Blüher, M. Adipokines in health and disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-kB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.; Choi, M.-S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Turer, A.T.; Hill, J.A.; Elmquist, J.K.; Scherer, P.E. Adipose tissue biology and cardiomyopathy: Translational implications. Circ. Res. 2012, 111, 1565–1577. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, A.S.; Obin, M.S. Obesity and the role of adipose tissue in inflammation and metabolism 1–4. Am. J. Clin. Nutr. 2006, 83, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Nieman, K.; Romero, I.; van Houten, B.; Lengyen, E. Adipose tissue and adipocytes supports tumorigenesis and metastasis. Biochim. Biophys. Acta 2013, 1831, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Vona-Davis, L.; Rose, D.P. Angiogenesis, adipokines and breast cancer. Cytokine Growth Factor Rev. 2009, 20, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Picon-Ruiz, M.; Pan, C.; Drews-elger, K.; Jang, K.; Besser, A.H.; Zhao, D.; Morata-tarifa, C.; Kim, M.; Ince, T.A.; Azzam, D.J.; et al. Interactions between adipocytes and breast cancer cells ctimulate cytokine production and drive Src/Sox2/miR-302b—Mediated malignant progression. Cancer Res. 2016, 76, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Duong, M.N.; Geneste, A.; Fallone, F.; Li, X.; Dumontet, C.; Muller, C. The fat and the bad: Mature adipocytes, key actors in tumor progression and resistance. Oncotarget 2017, 8, 57622–57641. [Google Scholar] [CrossRef] [PubMed]
- Neumann, E.; Junker, S.; Schett, G.; Frommer, K.; Müller-Ladner, U. Adipokines in bone disease. Nat. Rev. Rheumatol. 2016, 12, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Kusunoki, N.; Tanaka, N.; Kaneko, K.; Kusunoki, Y.; Endo, H.; Hasunuma, T.; Kawai, S. Elevated serum levels of resistin, leptin, and adiponectin are associated with C-reactive protein and also other clinical conditions in rheumatoid arthritis. Intern. Med. 2011, 50, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Gómez, R.; Conde, J.; Scotece, M.; Gómez-Reino, J.J.; Lago, F.; Gualillo, O. What’s new in our understanding of the role of adipokines in rheumatic diseases? Nat. Rev. Rheumatol. 2011, 7, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Abella, V.; Scotece, M.; Conde, J.; López, V.; Lazzaro, V.; Pino, J.; Gómez-Reino, J.J.; Gualillo, O. Adipokines, metabolic syndrome and rheumatic diseases. J. Immunol. Res. 2014, 2014, 343746. [Google Scholar] [CrossRef] [PubMed]
- Ehling, A.; Schäffler, A.; Herfarth, H.; Tarner, I.H.; Anders, S.; Distler, O.; Paul, G.; Distler, J.; Gay, S.; Schölmerich, J.; et al. The potential of adiponectin in driving arthritis. J. Immunol. 2006, 176, 4468–4478. [Google Scholar] [CrossRef] [PubMed]
- Frey, B.; Hehlgans, S.; Rödel, F.; Gaipl, U.S. Modulation of inflammation by low and high doses of ionizing radiation: Implications for benign and malign diseases. Cancer Lett. 2015, 368, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Yang, G.; Pan, Z.; Zhao, Y.; Liang, X.; Li, W. Hormetic response to low-dose radiation: Focus on the immune system and its clinical implications. Int. J. Mol. Sci. 2017, 18, 280. [Google Scholar] [CrossRef] [PubMed]
- Poglio, S.; Galvani, S.; Bour, S.; André, M.; Prunet-Marcassus, B.; Pénicaud, L.; Casteilla, L.; Cousin, B. Adipose tissue sensitivity to radiation exposure. Am. J. Pathol. 2009, 174, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.K.; Seol, M.A.; Park, H.R.; Jung, U.; Roh, C. Ionising radiation triggers fat accumulation in white adipose tissue. Int. J. Radiat. Biol. 2011, 87, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Posovszky, P.; Newell, F.; Wabitsch, M.; Tornqvist, H. Human SGBS cells—A unique tool for studies of human fat cell biology. Obes. Facts 2008, 1, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ojeda, F.J.; Ruperez, A.I.; Gomez-Llorente, C.; Gil, A.; Aguilera, C.M. Cell models and their application for studying adipogenic differentiation in relation to obesity: A review. Int. J. Mol. Sci. 2016, 17, 1040. [Google Scholar] [CrossRef] [PubMed]
- Wabitsch, M.; Brenner, R.E.E.; Melzner, I.; Braun, M.; Möller, P.; Heinze, E.; Debatin, K.-M.M.; Hauner, H. Characterization of a human preadipocyte cell strain with high capacity for adipose differentiation. Int. J. Obes. 2001, 25, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef] [PubMed]
- Armani, A.; Mammi, C.; Marzolla, V.; Calanchini, M.; Antelmi, A.; Rosano, G.M.C.; Fabbri, A.; Caprio, M. Cellular models for understanding adipogenesis, adipose dysfunction, and obesity. J. Cell. Biochem. 2010, 110, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Bochet, L.; Meulle, A.; Imbert, S.; Salles, B.; Valet, P.; Muller, C. Cancer-associated adipocytes promotes breast tumor radioresistance. Biochem. Biophys. Res. Commun. 2011, 411, 102–106. [Google Scholar] [CrossRef] [PubMed]
- Schäffler, A.; Schölmerich, J.; Buechler, C. Mechanisms of Disease: Adipokines and breast cancer—Endocrine and paracrine mechanisms that connect adiposity and breast cancer. Nat. Clin. Pract. Endocrin. Metab. 2007, 3, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Seegenschmiedt, M.H.; Micke, O.; Muecke, R. Radiotherapy for non-malignant disorders: State of the art and update of the evidence-based practice guidelines. Br. J. Radiol. 2015, 88, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rödel, F.; Fournier, C.; Wiedemann, J.; Merz, F.; Gaipl, U.S.; Frey, B.; Keilholz, L.; Seegenschmiedt, M.H.; Rödel, C.; Hehlgans, S. Basics of Radiation Biology When Treating Hyperproliferative Benign Diseases. Front. Immunol. 2017, 8, 519. [Google Scholar] [CrossRef] [PubMed]
- Cucu, A.; Shreder, K.; Kraft, D.; Rühle, P.F.; Klein, G.; Thiel, G.; Frey, B.; Gaipl, U.S.; Fournier, C. Decrease of Markers related to Bone erosion in serum of Patients with Musculoskeletal Disorders after serial low-Dose radon spa Therapy. Front. Immunol. 2017, 8, 882. [Google Scholar] [CrossRef] [PubMed]
- Multhoff, G.; Radons, J. Radiation, inflammation, and immune responses in cancer. Front. Oncol. 2012, 2, 58. [Google Scholar] [CrossRef] [PubMed]
- Rühle, A.; Xia, O.; Perez, R.L.; Trinh, T.; Richter, W.; Sarnowska, A.; Wuchter, P.; Debus, J.; Saffrich, R.; Huber, P.E.; et al. The Radiation Resistance of Human Multipotent Mesenchymal Stromal Cells Is Independent of Their Tissue of Origin. Int. J. Radiat. Oncol. Biol. Phys. 2018, 100, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Muthna, D.; Mokry, J.; Jiroutova, A.; Soukup, T. Irradiation of Adult Human Dental Pulp Stem Cells Provokes Activation of p53, Cell Cycle Arrest and Senescence but not Apoptosis. Stem Cells Dev. 2010, 19, 1855–1862. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-F.; Lin, C.T.; Chen, W.; Yang, C.; Chen, C.; Liao, S.; Liu, J.; Lu, C.; Lee, K. The Sensitivity of human mesenchymal stem cells to ionizing radiation. Int. J. Radiat. Oncol. 2006, 66, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Maria, O.M.; Kumala, S.; Heravi, M.; Syme, A.; Eliopoulos, N.; Muanza, T. Adipose mesenchymal stromal cells response to ionizing radiation. Cytotherapy 2016, 18, 384–401. [Google Scholar] [CrossRef] [PubMed]
- Nicolay, N.H.; Perez, R.L.; Saffrich, R.; Huber, P.E. Radio-resistant mesenchymal stem cells: Mechanisms of resistance and potential implications for the clinic. Oncotarget 2015, 6, 19366–19380. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.; Yang, X.; Lee, J.; Ryoo, Y.; Kim, J.; Oh, Y.; Kwon, S.; Liu, D.; Son, D. Serial changes in the proliferation and differentiation of adipose-derived stem cells after ionizing radiation. Stem Cell Res. Ther. 2016, 7, 117. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Kwong, D.L.W.; Chan, G.C.F. The effects of various irradiation doses on the growth and differentiation of marrow-derived human mesenchymal stromal cells. Pediatr. Transplant. 2007, 11, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Gal, T.J.; Munoz-Antonia, T.; Muro-Cacho, C.; Klotch, D.W. Radiation Effects on Osteoblasts In Vitro. Arch. Otolaryngol. Head Neck Surg. 2000, 126, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Van Dongen-Melman, J.E.W.M.; Hokken-Koelega, A.C.S.; Hahlen, K.; de Groot, A.; Tromp, C.G.; Egeler, R.M. Obesity after successful treatment of acute lymphoblastic leukemia in childhood. Pediatr. Res. 1995, 38, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.T.; Stovall, M.; Robinson, L.L. Long-term effects of radiation exposure among adult survivors of Childhood cancer: Results from the CCSS. Radiat. Res. 2011, 174, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; Mccann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [PubMed]
- Nicolay, N.H.; Liang, Y.; Perez, R.L.; Bostel, T.; Trinh, T.; Sisombath, S.; Weber, K.-J.; Ho, A.D.; Debus, J.; Saffrich, R.; et al. Mesenchymal stem cells are resistant to carbon ion radiotherapy. Oncotarget 2014, 6, 2076–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, C.R.; Agrawal, M.; Hoon, S.; Shabbir, A.; Shrivastava, M.K. SGBS cells as a model of human adipocyte browning: A comprehensive comparative study with primary human white subcutaneous adipocytes. Sci. Rep. 2017, 7, 4031. [Google Scholar] [CrossRef] [PubMed]
- Rühle, P.F.; Wunderlich, R.; Deloch, L.; Fournier, C.; Maier, A.; Klein, G.; Fietkau, R.; Gaipl, U.S.; Frey, B. Modulation of the peripheral immune system after low-dose radon spa therapy: Detailed longitudinal immune monitoring of patients within the RAD-ON01 study. Autoimmunity 2017, 50, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Moder, A.; Hufnagl, C.; Lind-Albrecht, G.; Hitzl, W.; Hartl, A.; Jakab, M.; Ritter, M. Effect of combined Low-Dose Radon-and Hyperthermia Treatment (LDRnHT) of patients with ankylosing spondylitis on serum levels of cytokines and bone metabolism markers: A pilot study. Int. J. Low Radiat. 2010, 7, 423–435. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shreder, K.; Rapp, F.; Tsoukala, I.; Rzeznik, V.; Wabitsch, M.; Fischer-Posovszky, P.; Fournier, C. Impact of X-ray Exposure on the Proliferation and Differentiation of Human Pre-Adipocytes. Int. J. Mol. Sci. 2018, 19, 2717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092717
Shreder K, Rapp F, Tsoukala I, Rzeznik V, Wabitsch M, Fischer-Posovszky P, Fournier C. Impact of X-ray Exposure on the Proliferation and Differentiation of Human Pre-Adipocytes. International Journal of Molecular Sciences. 2018; 19(9):2717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092717
Chicago/Turabian StyleShreder, Kateryna, Felicitas Rapp, Ioanna Tsoukala, Vanessa Rzeznik, Martin Wabitsch, Pamela Fischer-Posovszky, and Claudia Fournier. 2018. "Impact of X-ray Exposure on the Proliferation and Differentiation of Human Pre-Adipocytes" International Journal of Molecular Sciences 19, no. 9: 2717. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092717