Molecular Evidence for Precursors of Sjögren’s Foci in Histologically Normal Lacrimal Glands

Abstract
:1. Introduction
2. Results
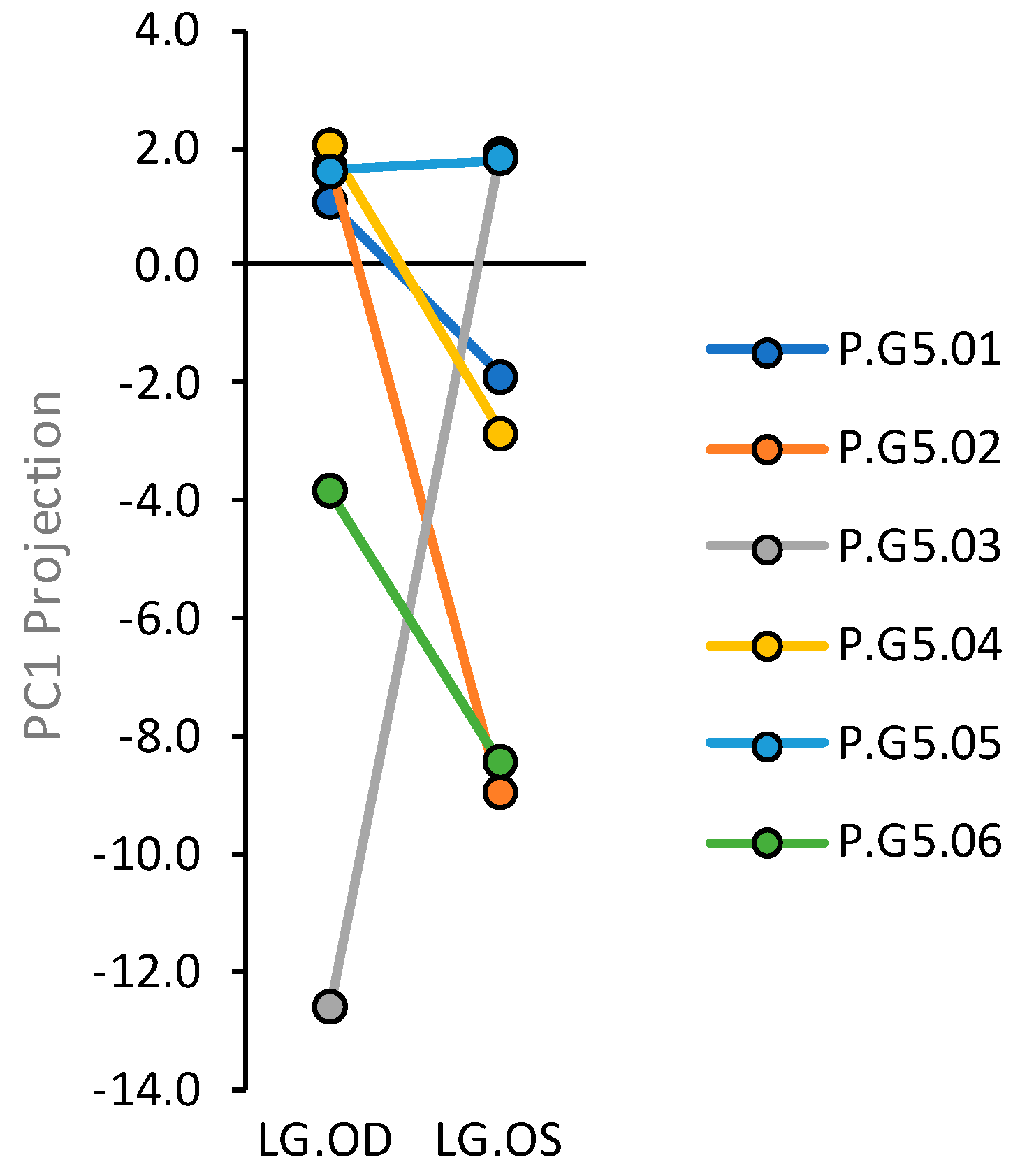
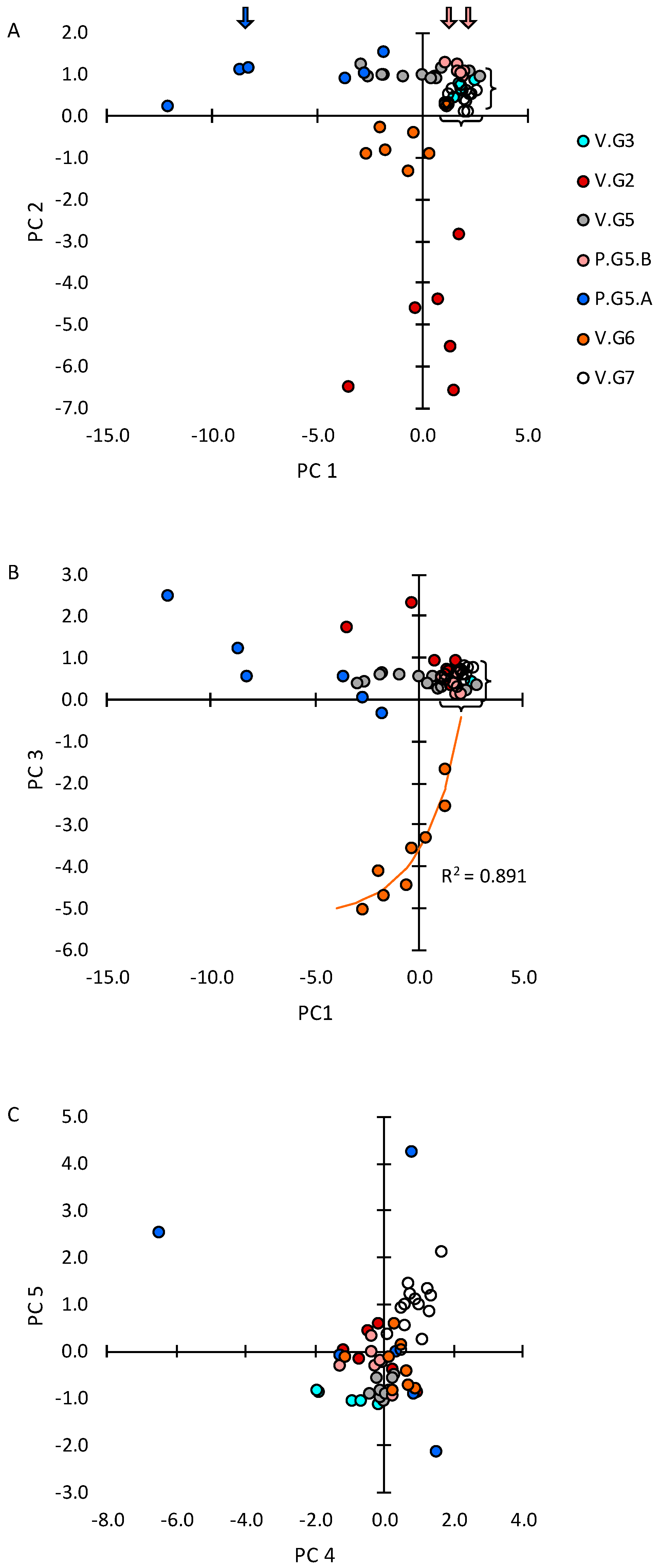
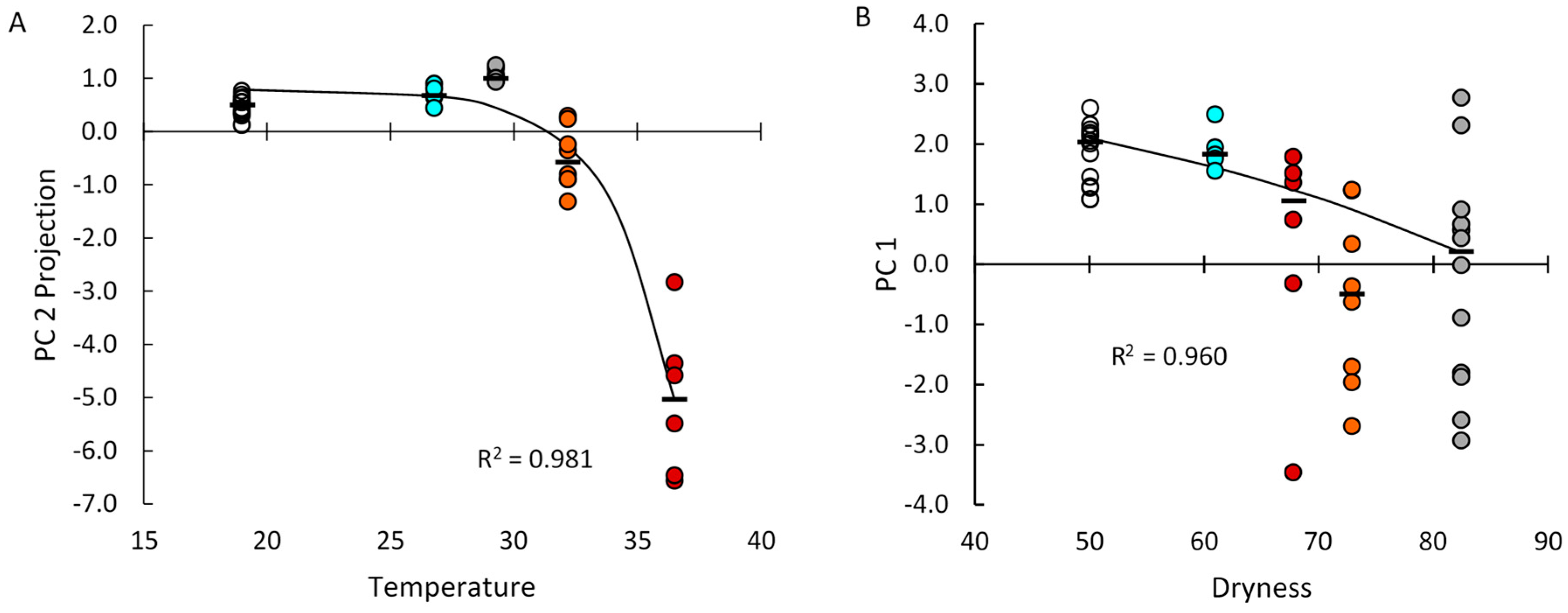
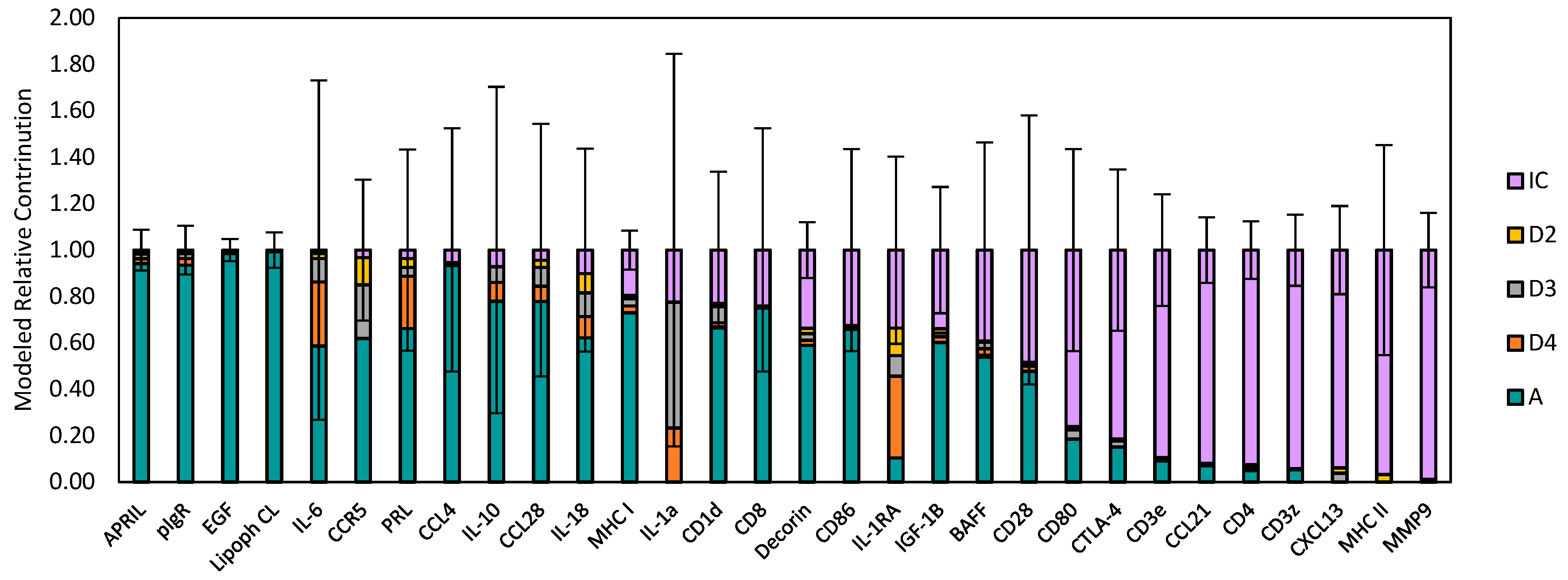
2.1. Systemic and Stochastic Variations in Immune Response-Related Gene Transcript Expression
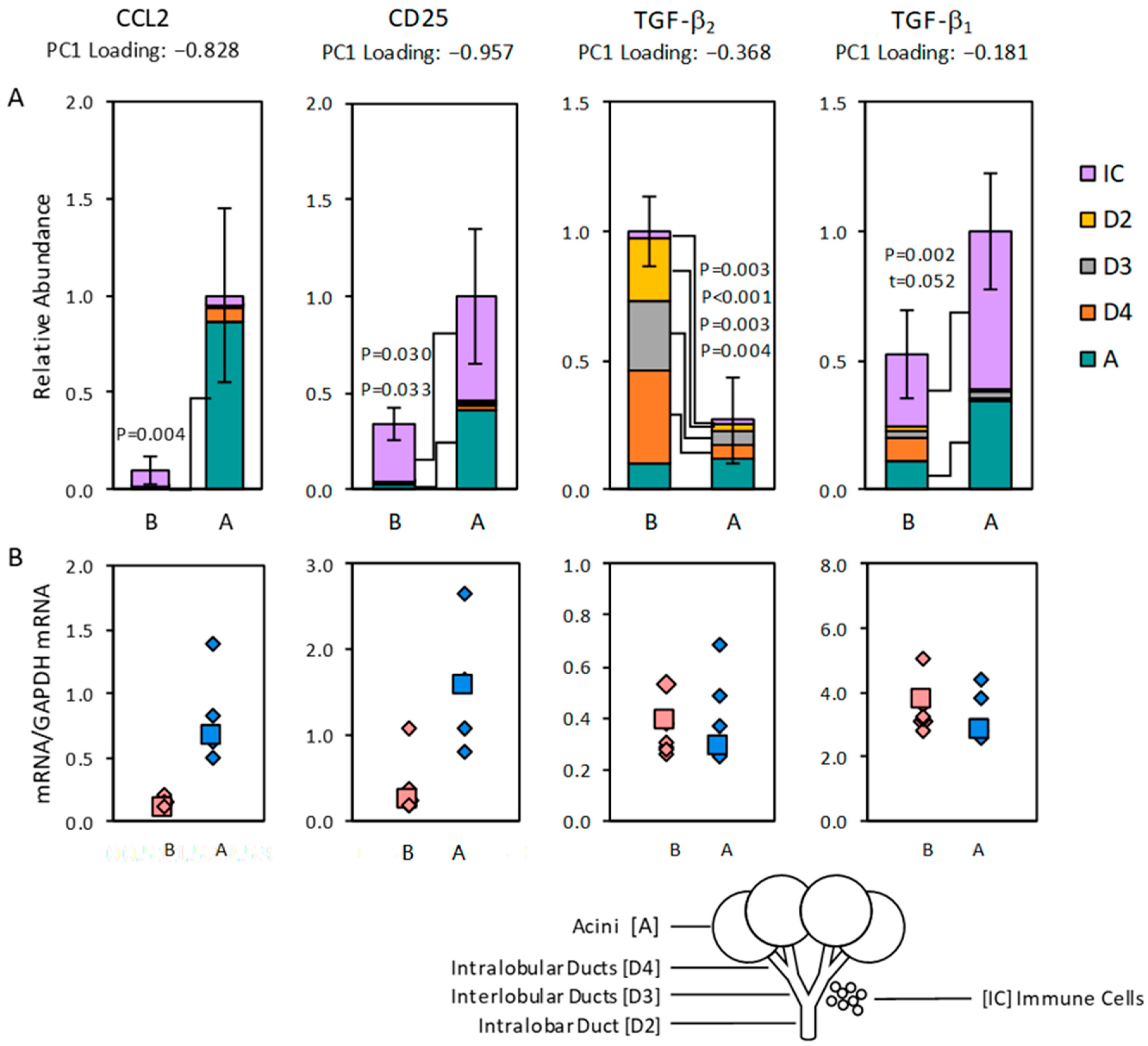
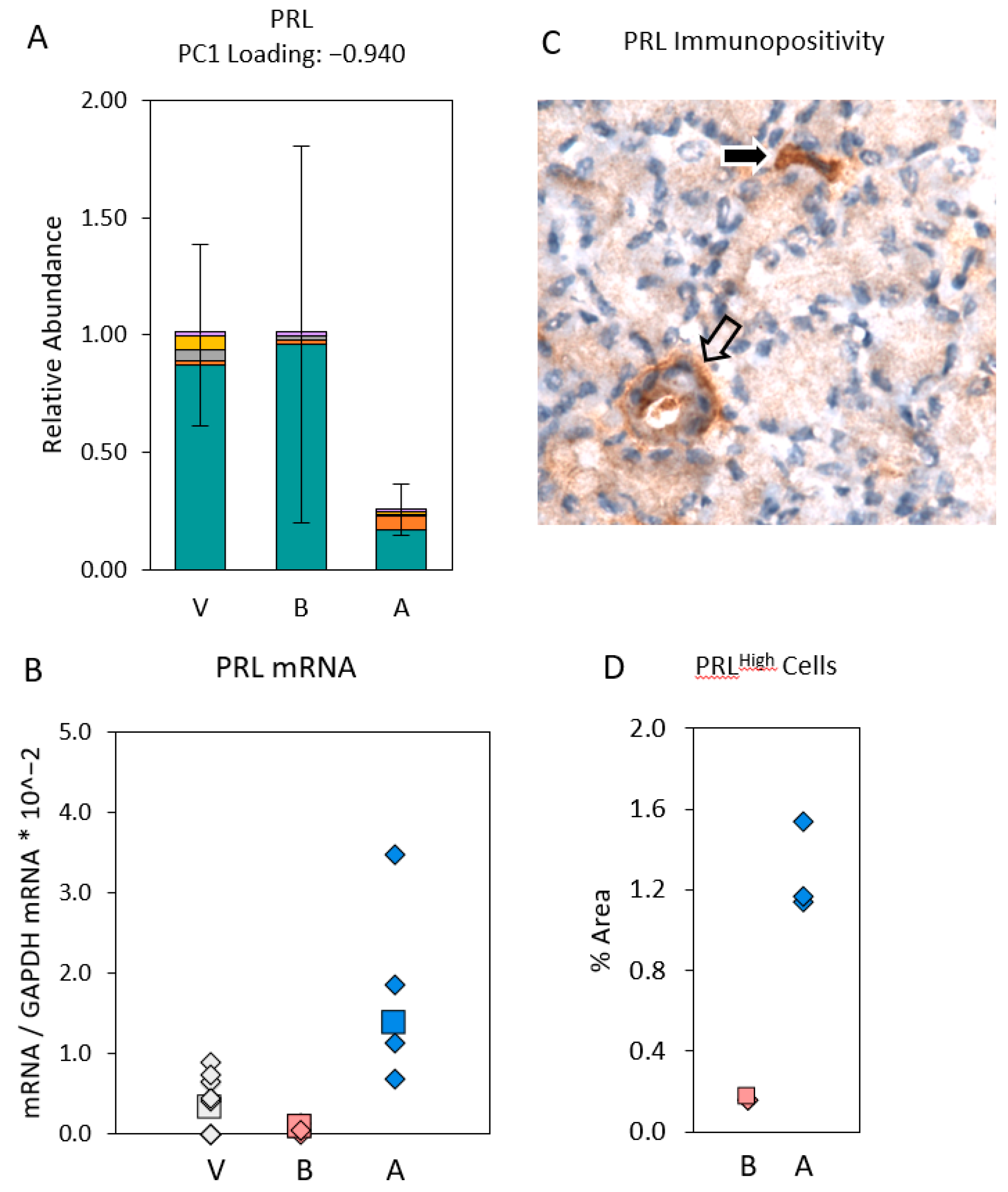
2.2. Distribution of Transcript Expression among Epithelial Segments and Immune Cells
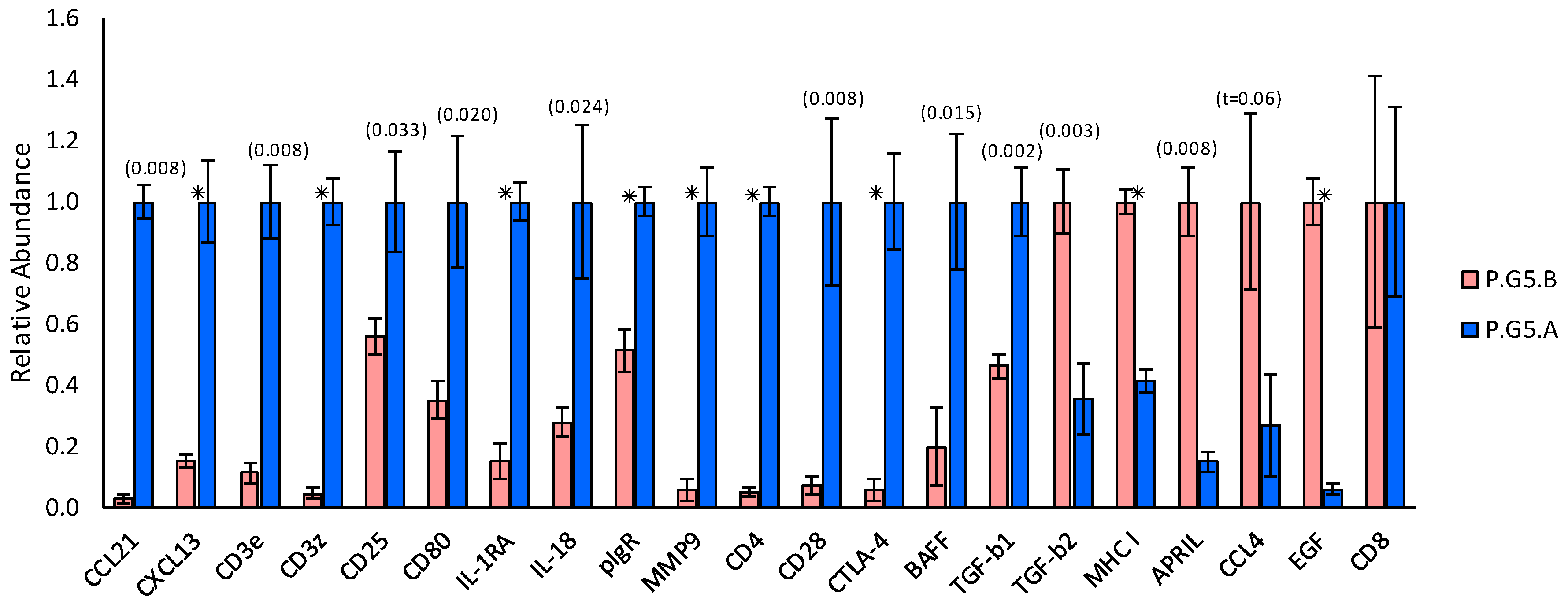
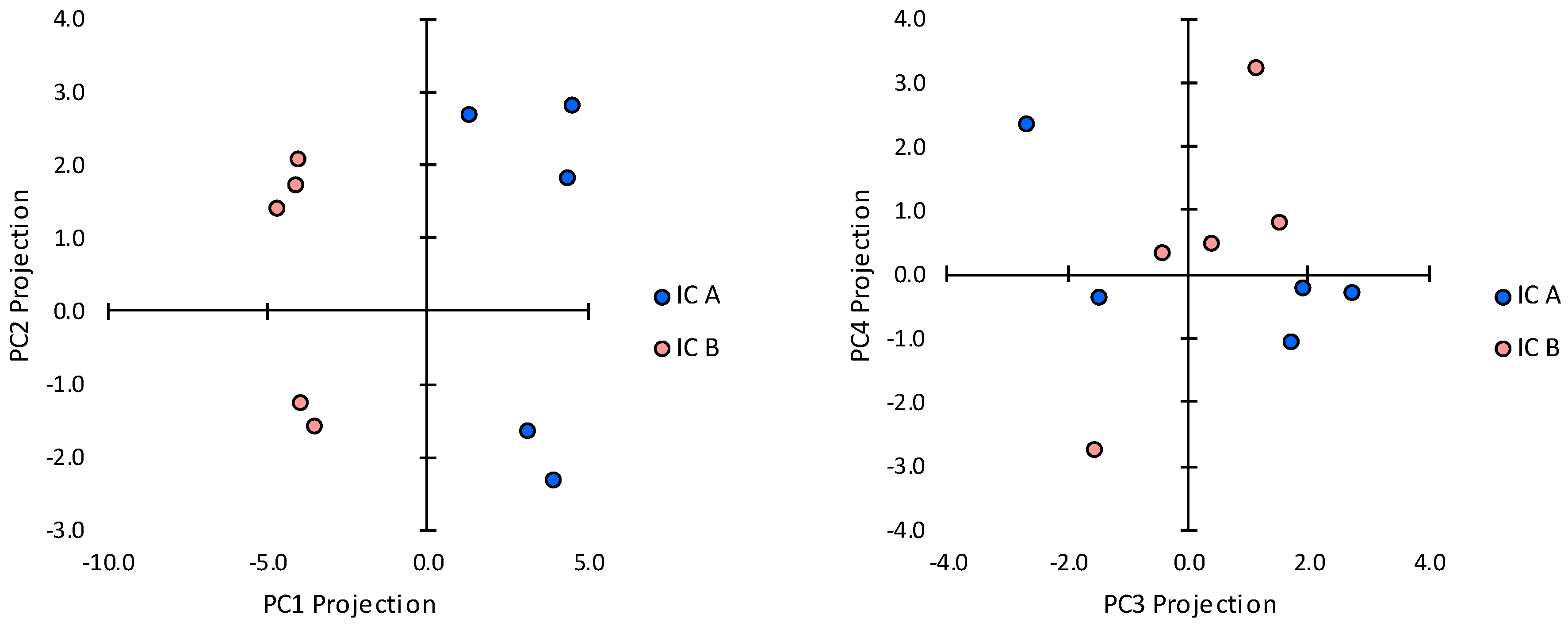
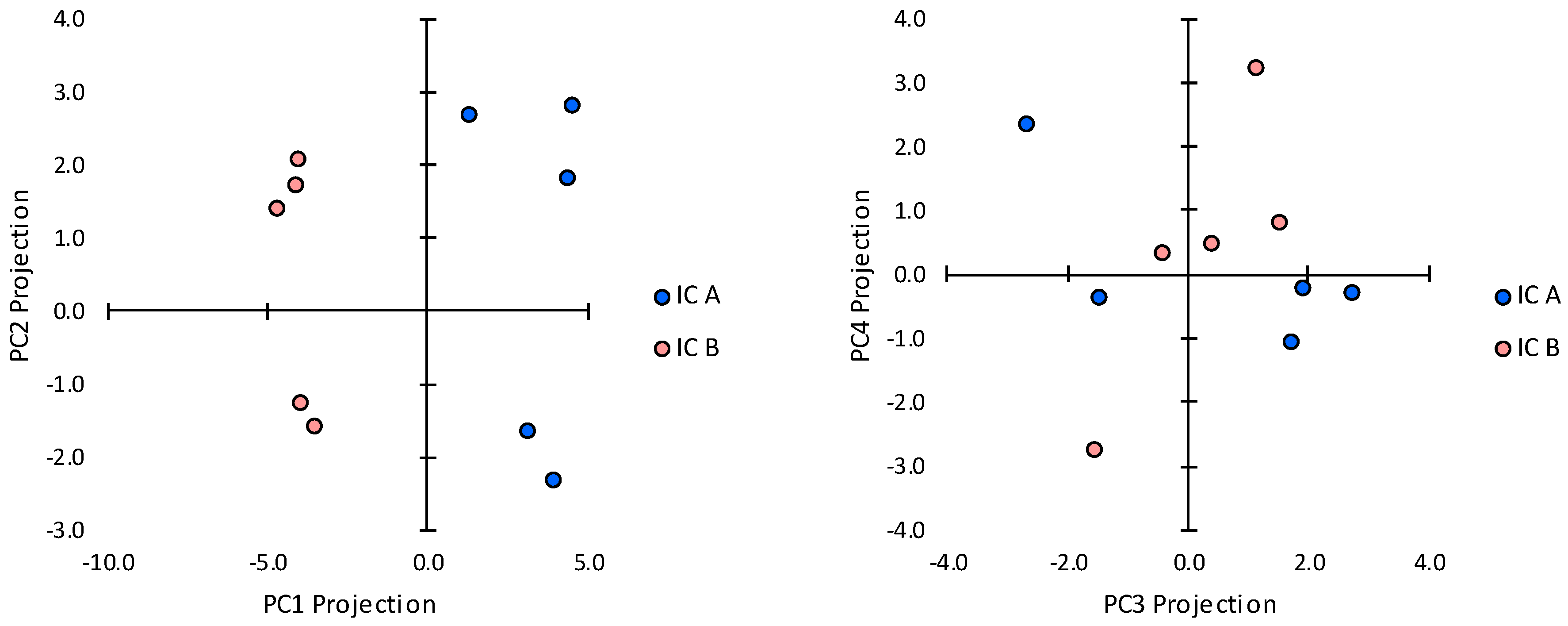
2.3. Transcript Expression in Immune Cell Clusters
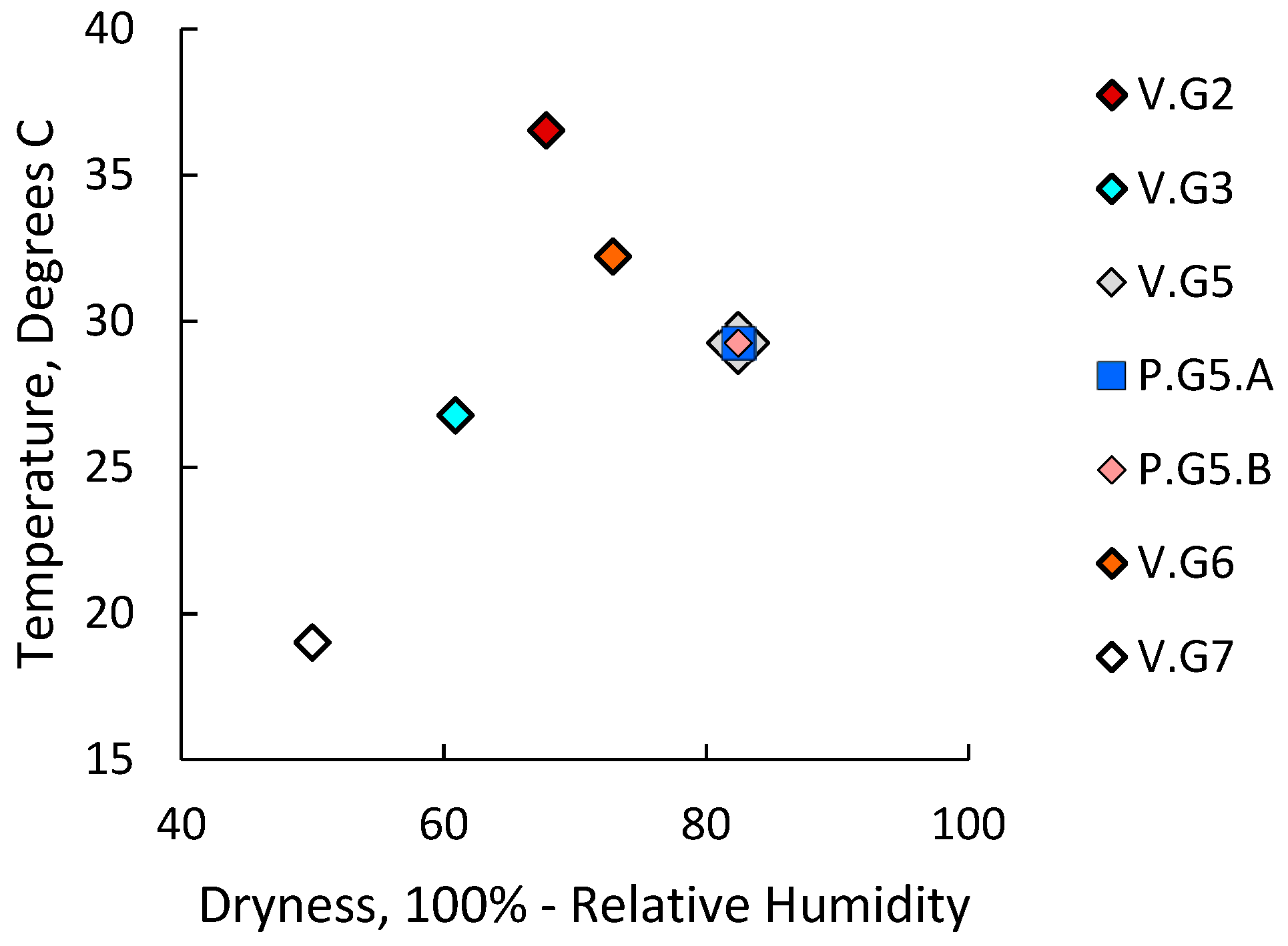
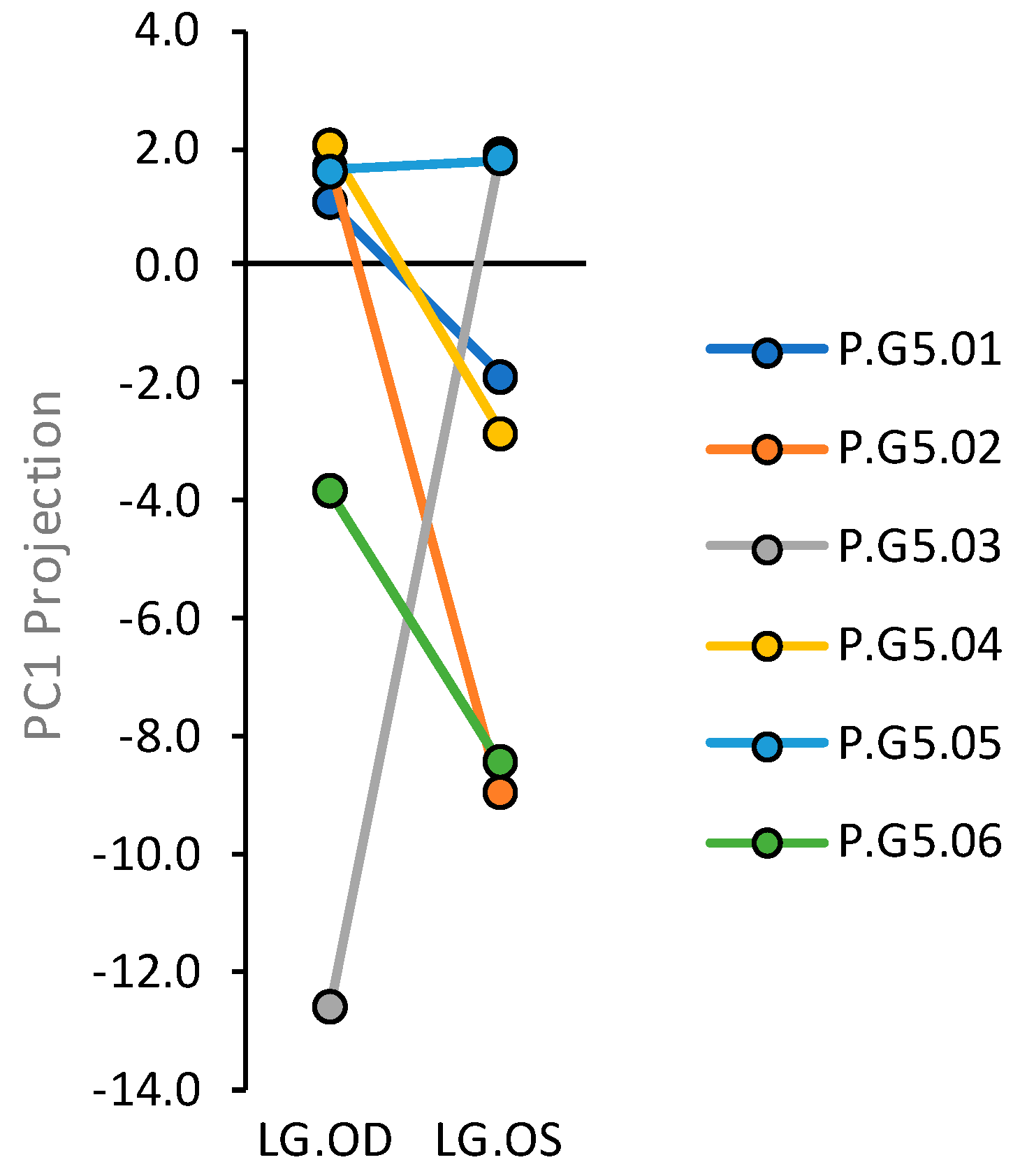
2.4. Cells Outside the Acinus Duct/Immune Cell Cluster Axis
3. Discussion
4. Methods
4.1. Animals
4.2. Tissue Collection
4.3. Laser Capture Microdissection
4.4. Real-Time Reverse-Transcriptase Polymerase Chain Reaction (Real Time RT-PCR)
4.5. Immunohistochemical Staining
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moutsopoulos, H.M.; Webber, B.L.; Vlagopoulos, T.P.; Chused, T.M.; Decker, J.L. Differences in the clinical manifestations of sicca syndrome in the presence and absence of rheumatoid arthritis. Am. J. Med. 1979, 66, 733–736. [Google Scholar] [CrossRef]
- Christodoulou, M.I.; Kapsogeorgou, E.K.; Moutsopoulos, H.M. Characteristics of the minor salivary gland infiltrates in Sjögren’s syndrome. J. Autoimmun. 2010, 34, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Maria, N.I.; Vogelsang, P.; Versnel, M.A. The clinical relevance of animal models in Sjögren’s syndrome: The interferon signature from mouse to man. Arthritis Res. Ther. 2015, 17, 172. [Google Scholar] [CrossRef] [PubMed]
- Kapsogeorgou, E.K.; Christodoulou, M.I.; Panagiotakos, D.B.; Paikos, S.; Tassidou, A.; Tzioufas, A.G.; Moutsopoulos, H.M. Minor salivary gland inflammatory lesions in Sjögren syndrome: Do they evolve? J. Rheumatol. 2013, 40, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Chused, T.M.; Kassan, S.S.; Opelz, G.; Moutsopoulos, H.M.; Terasaki, P.I. Sjögren’s syndrome association with HLA-Dw3. N. Engl. J. Med. 1977, 296, 895–897. [Google Scholar] [CrossRef]
- Moutsopoulos, H.M.; Chused, T.M.; Johnson, A.H.; Khudsen, B.; Mann, D.L. B lymphocyte antigens in sicca syndrome. Science 1978, 199, 1441–1442. [Google Scholar] [CrossRef] [PubMed]
- Gottenberg, J.E.; Busson, M.; Loiseau, P.; Cohen-Solal, J.; Lepage, V.; Charron, D.; Sibilia, J.; Mariette, X. In primary Sjögren’s syndrome, HLA class II is associated exclusively with autoantibody production and spreading of the autoimmune response. Arthritis Rheum. 2003, 48, 2240–2245. [Google Scholar] [CrossRef] [PubMed]
- Font, J.; Garcia-Carrasco, M.; Ramos-Casals, M.; Aldea, A.I.; Cervera, R.; Ingelmo, M.; Vives, J.; Yague, J. The role of interleukin-10 promoter polymorphisms in the clinical expression of primary Sjögren’s syndrome. Rheumatology 2002, 41, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.M.; Correa, P.A.; Herrera, M.; Eskdale, J.; Gallagher, G. Interleukin 10 (IL-10) influences autoimmune response in primary Sjogren’s syndrome and is linked to IL-10 gene polymorphism. J. Rheumatol. 2002, 29, 1874–1876. [Google Scholar] [CrossRef] [PubMed]
- Hulkkonen, J.; Pertovaara, M.; Antonen, J.; Lahdenpohja, N.; Pasternack, A.; Hurme, M. Genetic association between interleukin-10 promoter region polymorphisms and primary Sjögren’s syndrome. Arthritis Rheum. 2001, 44, 176–179. [Google Scholar] [CrossRef]
- Gottenberg, J.E.; Busson, M.; Loiseau, P.; Dourche, M.; Cohen-Solal, J.; Lepage, V.; Charron, D.; Miceli, C.; Sibilia, J.; Mariette, X. Association of transforming growth factor beta1 and tumor necrosis factor alpha polymorphisms with anti-SSB/La antibody secretion in patients with primary Sjogren’s syndrome. Arthritis Rheum. 2004, 50, 570–580. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, S.; Kanagawa, S.; Morinobu, A.; Takada, M.; Nakamura, K.; Sugai, S.; Maruya, E.; Saji, H. Association of a new allele of the TAP2 gene, TAP2*Bky2 (Val577), with susceptibility to Sjogren’s syndrome. Arthritis Rheum. 1997, 40, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Reksten, T.R.; Ice, J.A.; Kelly, J.A.; Adrianto, I.; Rasmussen, A.; Wang, S.; He, B.; Grundahl, K.M.; Glenn, S.B.; et al. Identification of a Sjögren’s syndrome susceptibility locus at OAS1 that influences isoform switching, protein expression, and responsiveness to type I. interferons. PLoS Genet. 2017, 13, e1006820. [Google Scholar] [CrossRef] [PubMed]
- Helmick, C.G.; Felson, D.T.; Lawrence, R.C.; Gabriel, S.; Hirsch, R.; Kwoh, C.K.; Liang, M.H.; Kremers, H.M.; Mayes, M.D.; Merkel, P.A.; et al. National Arthritis Data Workgroup. Estimates of the prevalence of arthritis and other rheumatic conditions in the United States. Part, I. Arthritis Rheum. 2008, 58, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Priori, R.; Medda, E.; Conti, F.; Cassarà, E.A.; Sabbadini, M.G.; Antonioli, C.M.; Gerli, R.; Danieli, M.G.; Giacomelli, R.; Pietrogrande, M.; et al. Risk factors for Sjögren’s syndrome: A case-control study. Clin. Exp. Rheumatol. 2007, 25, 378–384. [Google Scholar] [PubMed]
- Jørgensen, K.T.; Pedersen, B.V.; Nielsen, N.M.; Jacobsen, S.; Frisch, M. Childbirths and risk of female predominant and other autoimmune diseases in a population based Danish cohort. J. Autoimmun. 2012, 38, J81–J87. [Google Scholar] [CrossRef] [PubMed]
- Anaya, J.M.; Delgado-Vega, A.M.; Castiblanco, J. Genetic basis of Sjögren’s syndrome. How strong is the evidence? Clin. Dev. Immunol. 2006, 13, 209–222. [Google Scholar] [CrossRef]
- Hanafusa, T.; Pujol-Borrell, R.; Chiovato, L.; Russell, R.C.G.; Doniach, D.; Bottazzo, G.F. Aberrant expression of HLA-DR antigen on thyrocytes in Graves’ disease: Relevance for autoimmunity. Lancet 1983, 2, 1111–1115. [Google Scholar] [CrossRef]
- Bottazzo, G.F.; Pujol-Borrell, R.; Hanafusa, T. Role of aberrant HLA-DR expression and antigen presentation in induction of endocrine autoimmunity. Lancet 1983, 2, 1115–1119. [Google Scholar] [CrossRef]
- Lindahl, G.; Hedfors, E.; Klareskog, L.; Forsum, U. Epithelial HLA-DR expression and T lymphocyte subsets in salivary glands in Sjögren’s syndrome. Clin. Exp. Immunol. 1985, 61, 475–482. [Google Scholar]
- Fox, R.I.; Bumol, T.; Fantozzi, R.; Bone, R.; Schreiber, R. Expression of histocompatibility antigen HLA-DR by salivary gland epithelial cells in Sjogren’s syndrome. Arthritis Rheum. 1986, 29, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Moutsopoulos, H.M.; Hooks, J.J.; Chan, C.C.; Dalavanga, Y.A.; Skopouli, F.N.; Detrick, B. HLA-DR expression by labial minor salivary gland tissues in Sjögren’s syndrome. Ann. Rheum. Dis. 1986, 45, 677–683. [Google Scholar] [CrossRef]
- Rowe, D.; Griffiths, M.; Stewart, J.; Novick, D.; Beverley, P.C.; Isenberg, D.A. HLA class I and II, interferon, interleukin 2, and the interleukin 2 receptor expression on labial biopsy specimens from patients with Sjögren’s syndrome. Ann. Rheum. Dis. 1987, 46, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Manoussakis, M.N.; Dimitriou, I.D.; Kapsogeorgou, E.K.; Xanthou, G.; Paikos, S.; Polihronis, M.; Moutsopoulos, H.M. Expression of B7 costimulatory molecules by salivary gland epithelial cells in patients with Sjögren’s syndrome. Arthritis Rheum. 1999, 42, 229–239. [Google Scholar] [CrossRef]
- Dimitriou, I.D.; Kapsogeorgou, E.K.; Moutsopoulos, H.M.; Manoussakis, M.N. CD40 on salivary gland epithelial cells: High constitutive expression by cultured cells from Sjögren’s syndrome patients indicating their intrinsic activation. Clin. Exp. Immunol. 2002, 127, 386–392. [Google Scholar] [CrossRef]
- Cuello, C.; Palladinetti, P.; Tedla, N.; Di Girolamo, N.; Lloyd, A.R.; McCluskey, P.J.; Wakefield, D. Chemokine expression and leucocyte infiltration in Sjögren’s syndrome. Br. J. Rheumatol. 1998, 37, 779–783. [Google Scholar] [CrossRef]
- Xanthou, G.; Polihronis, M.; Tzioufas, A.G.; Paikos, S.; Sideras, P.; Moutsopoulos, H.M. “Lymphoid” chemokine messenger RNA expression by epithelial cells in the chronic inflammatory lesion of the salivary glands of Sjögren’s syndrome patients: Possible participation in lymphoid structure formation. Arthritis Rheum. 2001, 44, 408–418. [Google Scholar] [CrossRef]
- Ruffilli, I. Sjögren’s syndrome and chemokines. Clin. Ther. 2014, 165, e464–e469. [Google Scholar] [CrossRef]
- Ogawa, N.; Ping, L.; Zhenjun, L.; Takada, Y.; Sugai, S. Involvement of the interferon-gamma-induced T cell-attracting chemokines, interferon-gamma-inducible 10-kd protein (CXCL10) and monokine induced by interferon-gamma (CXCL9), in the salivary gland lesions of patients with Sjögren’s syndrome. Arthritis Rheum. 2002, 46, 2730–2741. [Google Scholar] [CrossRef]
- Latchney, L.R.; Fallon, M.A.; Culp, D.J.; Gelbard, H.A.; Dewhurst, S. Immunohistochemical assessment of fractalkine, inflammatory cells, and human herpesvirus 7 in human salivary glands. J. Histochem. Cytochem. 2004, 52, 671–681. [Google Scholar] [CrossRef]
- Lee, J.H.; Kwok, S.K.; Jung, S.M.; Lee, J.; Lee, J.S.; Baek, S.Y.; Kim, E.K.; Ju, J.H.; Park, S.H.; Kim, H.Y. Role of fractalkine in the pathogenesis of primary Sjögren syndrome: Increased serum levels of fractalkine, its expression in labial salivary glands, and the association with clinical manifestations. J. Rheumatol. 2014, 41, 2425–2438. [Google Scholar] [CrossRef] [PubMed]
- Barone, F.; Bombardieri, M.; Rosado, M.M.; Morgan, P.R.; Challacombe, S.J.; De Vita, S.; Carsetti, R.; Spencer, J.; Valesini, G.; Pitzalis, C. CXCL13, CCL21, and CXCL12 expression in salivary glands of patients with Sjogren’s syndrome and MALT lymphoma: Association with reactive and malignant areas of lymphoid organization. J. Immunol. 2008, 180, 5130–5140. [Google Scholar] [CrossRef] [PubMed]
- Berri, M.; Meurens, F.; Lefevre, F.; Chevaleyre, C.; Zanello, G.; Gerdts, V.; Salmon, H. Molecular cloning and functional characterization of porcine CCL28: Possible involvement in homing of IgA antibody secreting cells into the mammary gland. Mol. Immunol. 2008, 45, 271–277. [Google Scholar] [CrossRef]
- Cauli, A.; Yanni, G.; Pitzalis, C.; Challacombe, S.; Panayi, G.S. Cytokine and adhesion molecule expression in the minor salivary glands of patients with Sjögren’s syndrome and chronic sialoadenitis. Ann. Rheum. Dis. 1995, 54, 209–215. [Google Scholar] [CrossRef]
- Oxholm, P.; Daniels, T.E.; Bendtzen, K. Cytokine expression in labial salivary glands from patients with primary Sjögren’s syndrome. Autoimmunity 1992, 12, 185–191. [Google Scholar] [CrossRef]
- Bombardieri, M.; Barone, F.; Pittoni, V.; Alessandri, C.; Conigliaro, P.; Blades, M.C.; Priori, R.; McInnes, I.B.; Valesini, G.; Pitzalis, C. Increased circulating levels and salivary gland expression of interleukin-18 in patients with Sjögren’s syndrome: Relationship with autoantibody production and lymphoid organization of the periductal inflammatory infiltrate. Arthritis Res. Ther. 2004, 6, R447–R456. [Google Scholar] [CrossRef]
- Daridon, C.; Devauchelle, V.; Hutin, P.; Le Berre, R.; Martins-Carvalho, C.; Bendaoud, B.; Dueymes, M.; Saraux, A.; Youinou, P.; Pers, J.O. Aberrant expression of BAFF by B lymphocytes infiltrating the salivary glands of patients with primary Sjögren’s syndrome. Arthritis Rheum. 2007, 56, 1134–1144. [Google Scholar] [CrossRef]
- Sun, D.; Emmert-Buck, M.R.; Fox, P.C. Differential cytokine mRNA expression in human labial minor salivary glands in primary Sjögren’s syndrome. Autoimmunity 1998, 28, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Ha, Y.J.; Choi, Y.S.; Kang, E.H.; Chung, J.H.; Cha, S.; Song, Y.W.; Lee, Y.J. Increased expression of interferon-λ in minor salivary glands of patients with primary Sjögren’s syndrome and its synergic effect with interferon-α on salivary gland epithelial cells. Clin. Exp. Rheumatol. 2018, 36 (Suppl. 112), 31–40. [Google Scholar]
- Frey, W.H.; Nelson, J.D.; Frick, M.L.; Eide, R.P. Prolactin immunoreactivity in human tears and lacrimal gland: Possible implications for tear production. In The Preocular Tear Film in Health, Disease, and Contact Lens Wear; Holly, F.J., Ed.; The Dry Eye Institute: Lubbock, TX, USA, 1986; pp. 798–807. [Google Scholar]
- Hartmann, D.P.; Holaday, J.W.; Bernton, E.W. Inhibition of lymphocyte proliferation by antibodies to prolactin. FASEB J. 1989, 3, 2194–2202. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, D.W.; LeFevre, J.A.; Ulrich, E.D.; Adamson, C.R.; Zukowski, C.F. Identification of prolactin-like proteins synthesized by normal murine lymphocytes. Endocrinology 1990, 125, 2601–2603. [Google Scholar] [CrossRef]
- Peeva, E.; Venkatesh, J.; Michael, D.; Diamond, B. Prolactin as a modulator of B cell function: Implications for SLE. Biomed. Pharmacother. 2004, 58, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Jara, L.J.; Medina, G.; Saavedra, M.A.; Vera-Lastra, O.; Torres-Aguilar, H.; Navarro, C.; Vazquez Del Mercado, M.; Espinoza, L.R. Prolactin has a pathogenic role in systemic lupus erythematosus. Immunol. Res. 2017, 65, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Schechter, J.; Carey, J.; Wallace, M.; Wood, R. Distribution of growth factors and immune cells are altered in the lacrimal gland during pregnancy and lactation. Exp. Eye Res. 2000, 71, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Ding, C.; Chang, N.; Fong, Y.C.; Wang, Y.; Trousdale, M.D.; Mircheff, A.K.; Schechter, J.E. Interacting influences of pregnancy and corneal injury on rabbit lacrimal gland immunoarchitecture and function. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Mircheff, A.K.; Wang, Y.; Ding, C.; Warren, D.W.; Schechter, J.E. Potentially pathogenic immune cells and networks in apparently healthy lacrimal glands. Ocul. Surf. 2015, 13, 47–81. [Google Scholar] [CrossRef]
- Mircheff, A.K.; Wang, Y.; Thomas, P.B.; Nakamura, T.; Samant, D.; Trousdale, M.D.; Warren, D.W.; Ding, C.; Schechter, J.E. Systematic variations in immune response-related gene transcript abundance suggest new questions about environmental influences on lacrimal gland immunoregulation. Curr. Eye Res. 2011, 36, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Mircheff, A.K.; Wang, Y.; Li, M.; Pan, B.X.; Ding, C. Pregnancy probabilistically augments potential precursors to chronic, immune-mediated or autoimmune lacrimal gland infiltrates. Ocul. Surf. 2018, 16, 120–131. [Google Scholar] [CrossRef]
- Mircheff, A.K.; Wang, Y.; Schechter, J.E.; Li, M.; Tong, W.; Attar, M.; Chengalvala, M.; Harmuth, J.; Prusakiewicz, J.J. Multiple natural and experimental inflammatory rabbit lacrimal gland phenotypes. Ocul. Surf. 2016, 14, 460–483. [Google Scholar] [CrossRef]
- Tang, M.W.; Garcia, S.; Malvar Fernandez, B.; Gerlag, D.M.; Tak, P.P.; Reedquist, K.A. Rheumatoid arthritis and psoriatic arthritis synovial fluids stimulate prolactin production by macrophages. J. Leukoc. Biol. 2017, 102, 897–904. [Google Scholar] [CrossRef]
- Jonsson, R.; Theander, E.; Sjöström, B.; Brokstad, K.; Henriksson, G. Autoantibodies present before symptom onset in primary Sjögren syndrome. JAMA 2013, 310, 1854–1855. [Google Scholar] [CrossRef] [PubMed]
- Konttinen, Y.; Halinen, S.; Hanemaaijer, R.; Sorsa, T.; Hietanen, J.; Ceponis, A.; Xu, J.W.; Manthorpe, R.; Whittington, J.; Larsson, Å.; et al. Matrix metalloproteinase (MMP)-9 type IV collagenase/gelatinase implicated in the pathogenesis of Sjögren’s syndrome. Matrix Biol. 1998, 17, 335–347. [Google Scholar] [CrossRef]
- Macal, M.; Tam, M.A.; Hesser, C.; Di Domizio, J.; Leger, P.; Gilliet, M.; Zuniga, E.I. CD28 deficiency enhances type I IFN production by murine plasmacytoid dendritic cells. J. Immunol. 2016, 196, 1900–1909. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Z.X.; Yin, Z.; Liu, W.; Garcia, B.; Huang, X.; Acott, P.; Jevnikar, A.M. Anti-IL-2 receptor antibody decreases cytokine-induced apoptosis of human renal tubular epithelial cells (TEC). Nephrol. Dial. Transplant. 2011, 26, 2144–2153. [Google Scholar] [CrossRef] [PubMed]
- Tejman-Yarden, N.; Zlotnik, M.; Lewis, E.; Etzion, O.; Chaimovitz, C.; Douvdevani, A. Renal cells express a functional interleukin-15 receptor. Nephrol. Dial. Transplant. 2005, 20, 516–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanzavecchia, A.; Sallusto, F. Toll-like receptors and innate immunity in B-cell activation and antibody responses. Curr. Opin. Immunol. 2007, 19, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Guerrier, T.; Le Pottier, L.; Devauchelle, V.; Pers, J.O.; Jamin, C.; Youinou, P. Role of Toll-like receptors in primary Sjögren’s syndrome with a special emphasis on B-cell maturation within exocrine tissues. J. Autoimmun. 2012, 39, 69–76. [Google Scholar] [CrossRef]
- Nocturne, G.; Mariette, X. B cells in the pathogenesis of primary Sjögren syndrome. Nat. Rev. Rheumatol. 2018, 14, 133–145. [Google Scholar] [CrossRef]
- Maciel, G.; Crowson, C.S.; Matteson, E.L.; Cornec, D. Prevalence of primary Sjögren’s syndrome in a US population-based cohort. Arthritis Care Res. 2017, 69, 1612–1616. [Google Scholar] [CrossRef]
- Paschides, C.A.; Stefaniotou, M.; Papageorgiou, J.; Skourtis, P.; Psilas, K. Ocular surface and environmental changes. Acta Ophthalmol. Scand. 1998, 76, 74–77. [Google Scholar] [CrossRef] [Green Version]
- Fox, R.I.; Chilton, T.; Scott, S.; Benton, L.; Howell, F.V.; Vaughan, J.H. Potential role of Epstein–Barr virus in Sjögren’s syndrome. Rheum. Dis. Clin. North Am. 1987, 13, 275–292. [Google Scholar] [PubMed]
- Triantafyllopoulou, A.; Tapinos, N.; Moutsopoulos, H.M. Evidence for coxsakie virus infection in primary Sjögren’s syndrome. Arthritis Rheum. 2004, 50, 2897–2902. [Google Scholar] [CrossRef]
- Wang, Y.; Dou, H.; Liu, G.; Yu, L.; Chen, S.; Min, Y.; Zhao, K.; Wang, X.; Hu, C. Hepatitis C virus infection and the risk of Sjögren or sicca syndrome: A meta-analysis. Microbiol. Immunol. 2014, 58, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.L.; Gardener, M.R.; Bogus, Z.C.; Smith, M.A.; Astorri, E.; Michael, D.G.; Michael, D.A.; Zheng, C.; Burbelo, P.D.; Lai, Z.; et al. Hepatitis Delta virus detected in salivary glands of Sjögren’s syndrome patients and recapitulates a Sjögren’s syndrome-like phenotype in vivo. Pathog. Immun. 2016, 1, 12–40. [Google Scholar] [CrossRef]
- Terada, K.; Katamine, S.; Eguchi, K.; Moriuchi, R.; Kita, M.; Shimada, H.; Yamashita, I.; Iwata, K.; Tsuji, Y.; Nagataki, S.; et al. Prevalence of serum and salivary antibodies to HTLV-1 in Sjogren’s syndrome. Lancet 1994, 344, 1116–1119. [Google Scholar] [CrossRef]
- Ittah, M.; Miceli-Richard, C.; Lebon, P.; Pallier, C.; Lepajolec, C.; Mariette, X. Induction of B cell-activating factor by viral infection is a general phenomenon, but the types of viruses and mechanisms depend on cell type. J. Innate Immun. 2011, 3, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Croia, C.; Astorri, E.; Murray-Brown, W.; Willis, A.; Brokstad, K.A.; Sutcliffe, N.; Piper, K.; Jonsson, R.; Tappuni, A.R.; Pitzalis, C.; et al. Implication of Epstein-Barr virus infection in disease-specific autoreactive B cell activation in ectopic lymphoid structures of Sjögren’s syndrome. Arthritis Rheumatol. 2014, 66, 2545–2557. [Google Scholar] [CrossRef]
- Chaigne, B.; Lasfargues, G.; Marie, I.; Hüttenberger, B.; Lavigne, C.; Marchand-Adam, S.; Maillot, F.; Diot, E. Primary Sjögren’s syndrome and occupational risk factors: A. case-control study. J. Autoimmun. 2015, 60, 80–85. [Google Scholar] [CrossRef]
- Skopouli, F.N.; Katsiougiannis, S. How stress contributes to autoimmunity-lessons from Sjögren’s syndrome. FEBS Lett. 2018, 592, 5–14. [Google Scholar] [CrossRef]
- Ding, C.; Nandoskar, P.; Lu, M.; Thomas, P.; Trousdale, M.D.; Wang, Y. Changes of aquaporins in the lacrimal glands of a rabbit model of Sjögren’s syndrome. Curr. Eye Res. 2011, 36, 571–578. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | PC 1 | Variable | PC 2 | Variable | PC 3 | Variable | PC 4 | Variable | PC 5 |
|---|---|---|---|---|---|---|---|---|---|
| CCL28 | 0.2729 | CCL21 | −0.9465 | CCL2 | −0.8706 | TNF-a | −0.7861 | CCL4 | −0.2317 |
| MMP-9 | 0.0677 | IL-18 | −0.9337 | MHC II | −0.8296 | CD4 | −0.5162 | IL-6 | −0.1182 |
| IL-18 | 0.0516 | MMP-9 | −0.9093 | CCL4 | −0.5254 | CD8 | −0.2194 | IL-10 | −0.1129 |
| CCL21 | 0.0250 | PRL | −0.7501 | TNF-a | −0.3396 | CD28 | −0.2127 | BAFF | −0.1065 |
| TNF-a | −0.0210 | BAFF | −0.4072 | CTLA-4 | −0.2869 | IL-18 | −0.1159 | PRL | −0.0890 |
| CXCL13 | −0.3782 | MHC II | −0.1641 | IL-1a | −0.2599 | CCL21 | −0.1053 | CCR5 | −0.0661 |
| CCL2 | −0.3901 | CCL2 | −0.1145 | MMP-9 | −0.0866 | CTLA-4 | −0.0938 | CD4 | −0.0614 |
| MHC II | −0.4494 | CD4 | −0.0912 | CXCL13 | −0.0763 | CXCL13 | −0.0133 | IL-1a | −0.0458 |
| PRL | −0.5794 | CD28 | −0.0561 | CCL21 | −0.0381 | MMP-9 | 0.0682 | CCL21 | −0.0397 |
| BAFF | −0.6719 | CTLA-4 | −0.0027 | PRL | 0.1012 | BAFF | 0.0720 | CCL2 | 0.0190 |
| CD4 | −0.7205 | CXCL13 | 0.0184 | IL-1b | 0.1161 | MHC II | 0.0929 | MHC II | 0.0314 |
| CCL4 | −0.7319 | IL-10 | 0.0496 | CCR5 | 0.1804 | IL-6 | 0.1006 | IL-18 | 0.0832 |
| CTLA-4 | −0.8066 | CCL4 | 0.0552 | IL-18 | 0.1815 | PRL | 0.1062 | CD8 | 0.0882 |
| CD8 | −0.8348 | CCL28 | 0.0898 | CCL28 | 0.1888 | IL-1a | 0.1293 | CTLA-4 | 0.1373 |
| CD28 | −0.8400 | IL-1a | 0.0956 | IL-6 | 0.2037 | CCL4 | 0.1308 | CD28 | 0.1912 |
| IL-10 | −0.8485 | CCR5 | 0.1113 | CD4 | 0.2227 | CCL2 | 0.1448 | MMP-9 | 0.2257 |
| IL-1b | −0.8651 | IL-6 | 0.1369 | CD8 | 0.2370 | CCR5 | 0.1518 | TNF-a | 0.2653 |
| CCR5 | −0.9154 | TNF-α | 0.1684 | CD28 | 0.2759 | IL-1b | 0.1657 | IL-1b | 0.2748 |
| IL-6 | −0.9262 | IL-1b | 0.2006 | BAFF | 0.3343 | IL-10 | 0.1798 | CXCL13 | 0.6024 |
| IL-1a | −0.9291 | CD8 | 0.2979 | IL-10 | 0.3915 | CCL28 | 0.4592 | CCL28 | 0.6365 |
| Variable | PC 1 | Variable | PC 2 | Variable | PC 3 | Variable | PC 4 | Variable | PC 5 | Variable | PC 6 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| EGF | −0.9669 | CD86 | −0.8548 | MHC II | −0.8895 | Lipoph CL | −0.7080 | IGF-1B | −0.7061 | IL-10 | −0.5272 |
| MHC I | −0.9306 | CD1d | −0.8354 | IL-1a | −0.6157 | IL-6 | −0.5981 | TGF-b2 | −0.5759 | CCL28 | −0.5047 |
| APRIL | −0.9263 | CD8 | −0.7284 | PRL | −0.5481 | IL-10 | −0.5372 | CD80 | −0.5058 | Decorin | −0.3251 |
| TGF-b2 | −0.7619 | Decorin | −0.6752 | Decorin | −0.4931 | IGF-1B | −0.3973 | IL-18 | −0.5025 | CCL2 | −0.3127 |
| CCL4 | −0.6012 | Lipoph CL | −0.4969 | CD25 | −0.4266 | CD28 | −0.3191 | TGF-b1 | −0.3276 | IL-6 | −0.2676 |
| IL-6 | −0.5577 | CCL4 | −0.4626 | CCL28 | −0.4064 | Decorin | −0.2763 | CD28 | −0.1887 | BAFF | −0.2245 |
| IGF-1B | −0.4630 | CCL28 | −0.4518 | Lipoph CL | −0.3398 | CCR5 | −0.2163 | Decorin | −0.1561 | IL-18 | −0.1994 |
| CCL2 | −0.2098 | BAFF | −0.4488 | IL-6 | −0.2717 | APRIL | −0.1796 | MMP9 | −0.1298 | CCL4 | −0.1507 |
| CD86 | −0.1466 | IGF-1B | −0.3372 | CTLA-4 | −0.2486 | IL-1RA | −0.1577 | MHC I | −0.1260 | CD3z | −0.1246 |
| IL-10 | −0.0836 | CCL2 | −0.3210 | APRIL | −0.2257 | MHC II | −0.1448 | EGF | −0.1161 | CD80 | −0.1122 |
| Decorin | −0.0128 | CCR5 | −0.2810 | EGF | −0.1710 | CCL21 | −0.1441 | CD25 | −0.1106 | MHC II | −0.0921 |
| CD8 | −0.0095 | MHC I | −0.2544 | CD86 | −0.1617 | CD4 | −0.1188 | APRIL | −0.0994 | CTLA-4 | −0.0863 |
| CCR5 | 0.0799 | PRL | −0.2342 | IL-18 | −0.1556 | CD86 | −0.1145 | BAFF | −0.0947 | MMP9 | −0.0648 |
| MHC II | 0.1667 | CD80 | −0.1728 | CD1d | −0.1345 | CD8 | −0.1125 | Lipoph CL | −0.0768 | CD25 | −0.0532 |
| Lipoph CL | 0.2042 | pIgR | −0.1648 | MHC I | −0.1339 | TGF-b1 | −0.1108 | CCL28 | −0.0418 | CCR5 | −0.0466 |
| IL-1a | 0.2462 | TGF-b1 | −0.1478 | CXCL13 | −0.1124 | CD3z | −0.0994 | CTLA-4 | −0.0407 | APRIL | −0.0448 |
| CCL28 | 0.3279 | MMP9 | −0.1086 | TGF-b2 | −0.1007 | EGF | −0.0816 | CD3e | −0.0385 | CD1d | −0.0284 |
| CD1d | 0.4557 | APRIL | −0.0702 | pIgR | −0.0436 | MMP9 | −0.0549 | CCL4 | −0.0275 | CD28 | −0.0282 |
| PRL | 0.5574 | CCL21 | −0.0211 | CD3z | −0.0419 | pIgR | −0.0253 | CD3z | −0.0120 | IL-1RA | −0.0239 |
| CD28 | 0.7298 | EGF | −0.0137 | CD3e | 0.0378 | BAFF | −0.0240 | CXCL13 | −0.0042 | CD4 | 0.0144 |
| CD25 | 0.7883 | CD3z | 0.0071 | IL-10 | 0.0487 | TGF-b2 | −0.0121 | pIgR | −0.0002 | TGF-b2 | 0.0481 |
| IL-18 | 0.7890 | IL-10 | 0.0082 | MMP9 | 0.0510 | CXCL13 | −0.0049 | IL-1a | 0.0355 | TGF-b1 | 0.0605 |
| BAFF | 0.7992 | CXCL13 | 0.0388 | IL-1RA | 0.0624 | CD80 | −0.0029 | CD4 | 0.0361 | CCL21 | 0.0724 |
| CD80 | 0.8063 | CD25 | 0.0636 | IGF-1B | 0.0842 | MHC I | 0.0216 | CD1d | 0.0551 | IGF-1B | 0.0754 |
| TGF-β1 | 0.9121 | MHC II | 0.0790 | CCL21 | 0.0901 | IL-18 | 0.0328 | CCL2 | 0.0847 | CXCL13 | 0.0889 |
| IL-1RA | 0.9199 | CD3e | 0.1110 | TGF-b1 | 0.0920 | CD3e | 0.0533 | IL-6 | 0.0866 | MHC I | 0.0939 |
| pIgR | 0.9225 | TGF-b2 | 0.1111 | CCL4 | 0.1049 | CTLA-4 | 0.1141 | CD8 | 0.1309 | EGF | 0.0973 |
| CTLA-4 | 0.9261 | CD4 | 0.1112 | CD4 | 0.1178 | CD25 | 0.1266 | CD86 | 0.1542 | CD3e | 0.1127 |
| CCL21 | 0.9589 | CD28 | 0.1157 | BAFF | 0.1920 | CD1d | 0.1957 | CCL21 | 0.1991 | CD86 | 0.1171 |
| CXCL13 | 0.9673 | CTLA-4 | 0.1425 | CD80 | 0.2099 | PRL | 0.2016 | IL-1RA | 0.2002 | Lipoph CL | 0.1818 |
| CD3e | 0.9734 | IL-18 | 0.2005 | CD8 | 0.3040 | IL-1a | 0.4265 | PRL | 0.2299 | pIgR | 0.1999 |
| CD3z | 0.9749 | IL-1RA | 0.2385 | CCR5 | 0.3410 | CCL28 | 0.4501 | MHC II | 0.2489 | IL-1a | 0.3058 |
| CD4 | 0.9761 | IL-6 | 0.3726 | CCL2 | 0.3749 | CCL4 | 0.4738 | IL-10 | 0.4431 | PRL | 0.4001 |
| MMP9 | 0.9795 | IL-1a | 0.4557 | CD28 | 0.5472 | CCL2 | 0.7207 | CCR5 | 0.7383 | CD8 | 0.5382 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mircheff, A.K.; Wang, Y.; Pan, B.X.; Parsa, L.; Nandoskar, P.; Ding, C. Molecular Evidence for Precursors of Sjögren’s Foci in Histologically Normal Lacrimal Glands. Int. J. Mol. Sci. 2019, 20, 223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010223
Mircheff AK, Wang Y, Pan BX, Parsa L, Nandoskar P, Ding C. Molecular Evidence for Precursors of Sjögren’s Foci in Histologically Normal Lacrimal Glands. International Journal of Molecular Sciences. 2019; 20(1):223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010223
Chicago/Turabian StyleMircheff, Austin K., Yanru Wang, Billy X. Pan, Leili Parsa, Prachi Nandoskar, and Chuanqing Ding. 2019. "Molecular Evidence for Precursors of Sjögren’s Foci in Histologically Normal Lacrimal Glands" International Journal of Molecular Sciences 20, no. 1: 223. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20010223