Role of Nephronectin in Pathophysiology of Silicosis

 and
and
Abstract
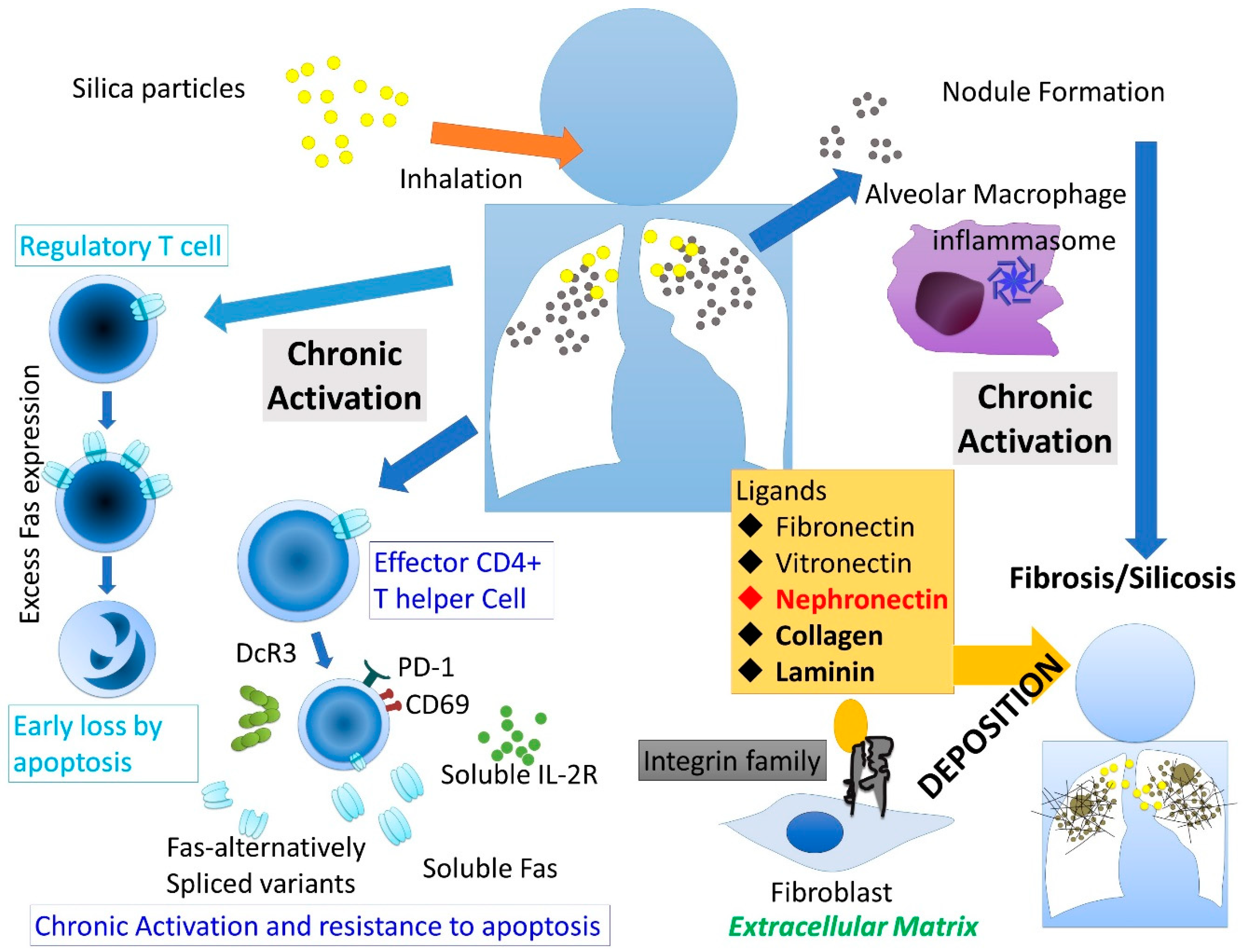
:1. Introduction
2. Silicosis and Integrin
3. Silicosis and Ligands for Integrin Family
3.1. Fibronectin (FN)
3.2. Vitronectin
3.3. Laminin
3.4. Collagen
4. Npnt in Silicosis
4.1. Patients and Healthy Volunteers
4.2. Measurement of Serum Npnt and Other Cytokines
4.3. Statistical Analyses
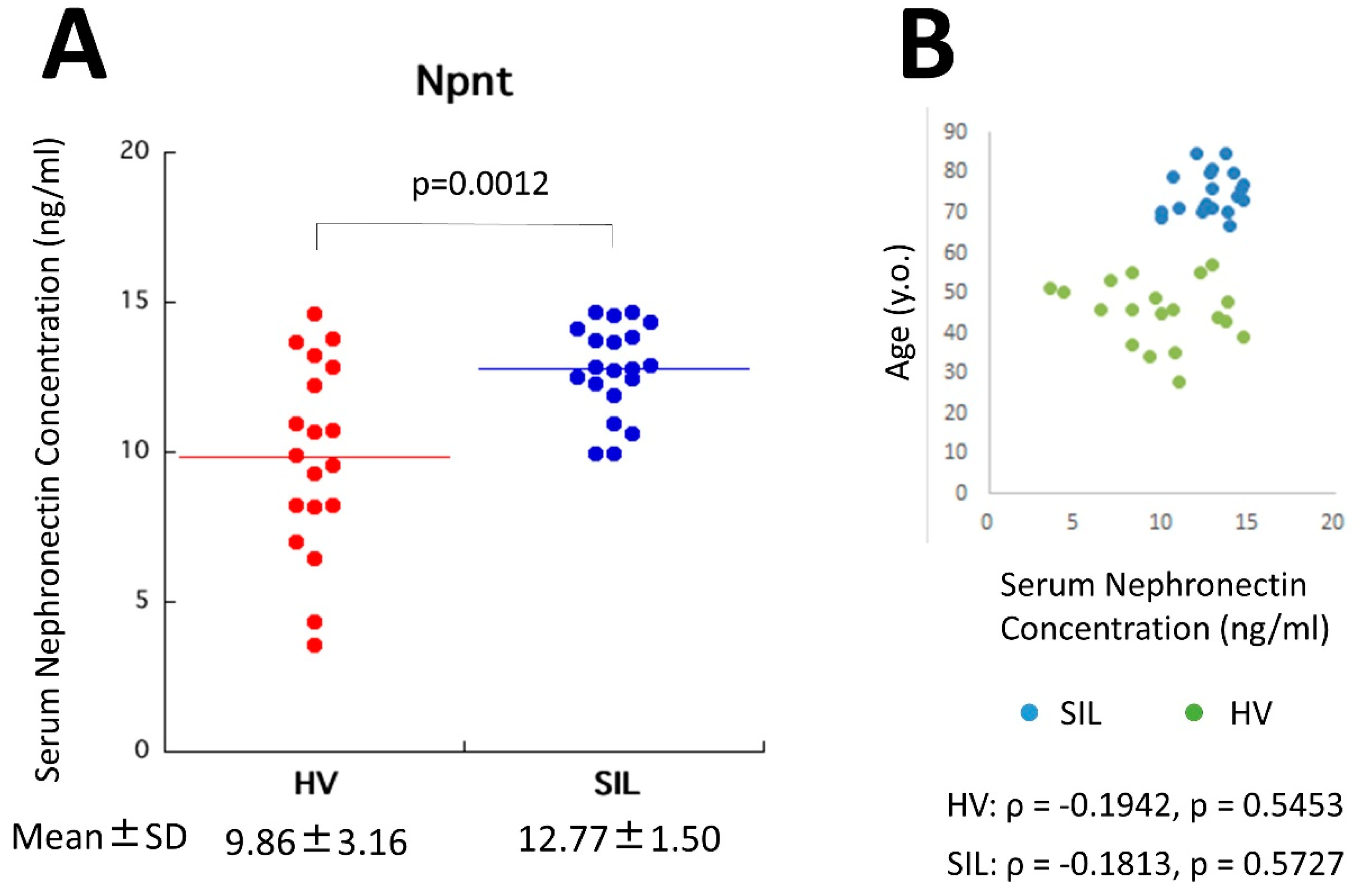
4.4. Serum Npnt in Silicosis Patients and Healthy Volunteers
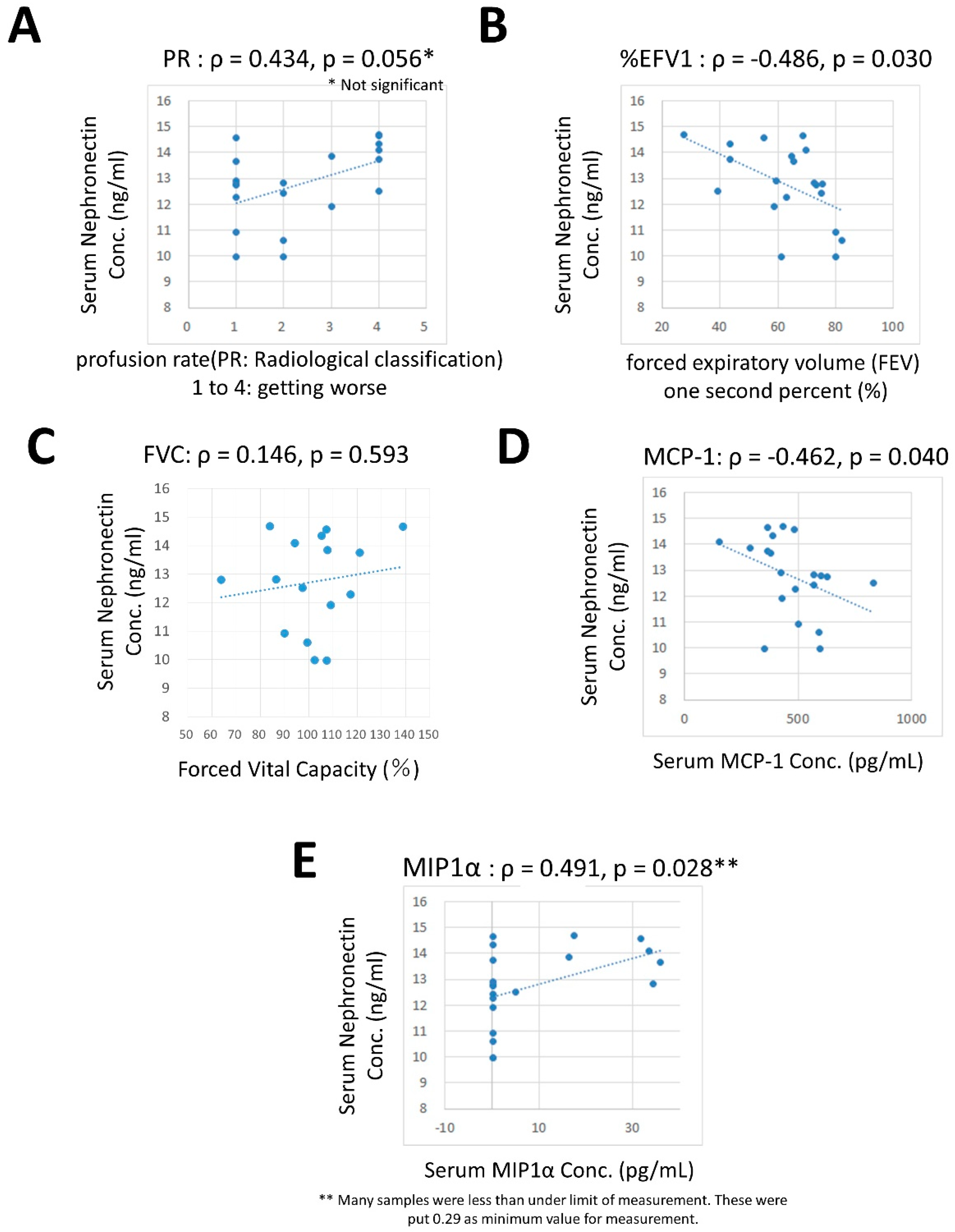
4.5. Correlation between Npnt and Other Cytokines, Growth Factors, and Respiratory Factors.
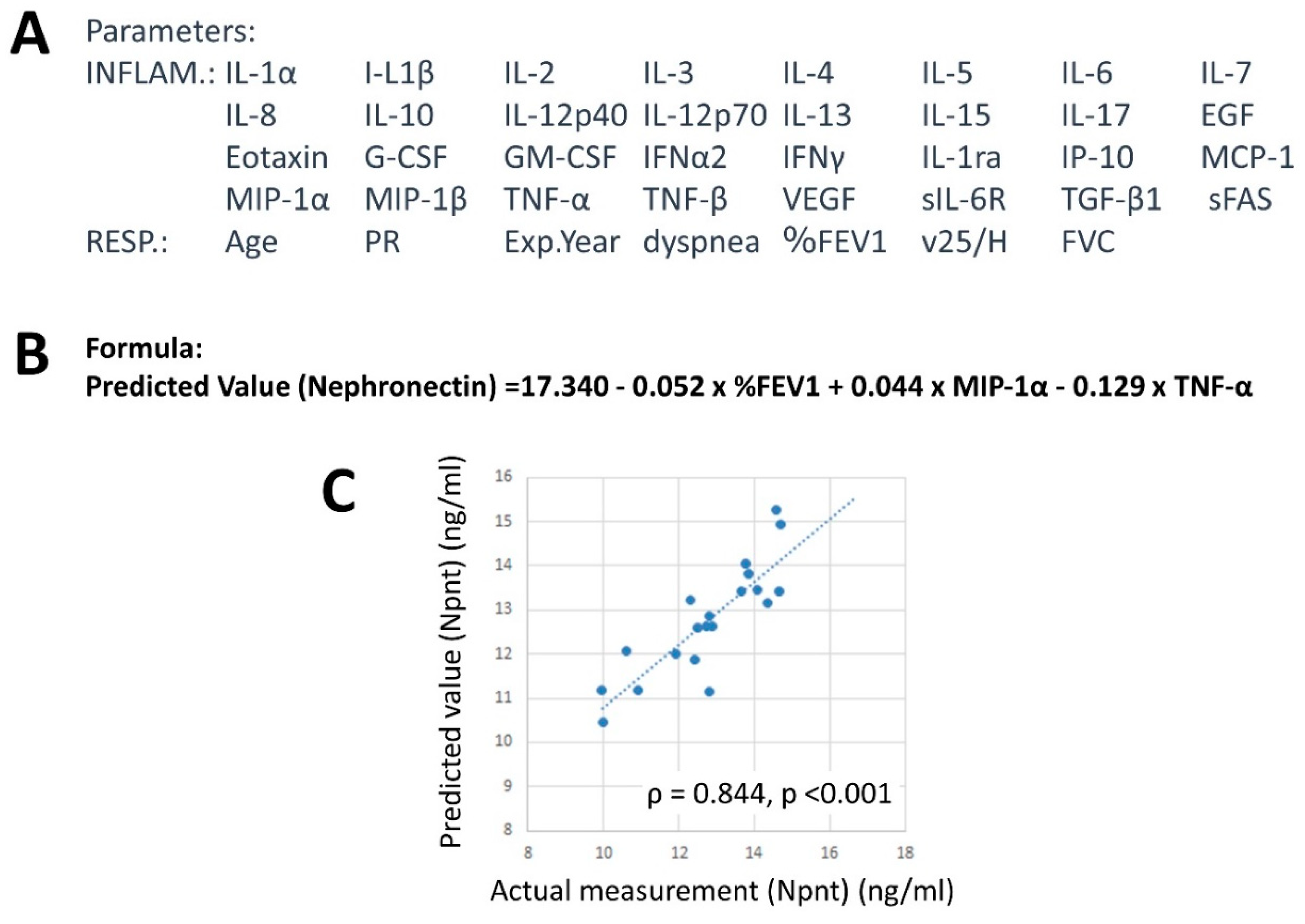
4.6. Factor Analysis
4.7. Factors Related to Npnt Levels
4.8. Summary of NPNT in Silicosis
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| MARCO | macrophage receptor with collagenous structure |
| IL | interleukin |
| Treg | regulatory T |
| NLR | NOD-like receptor |
| NLRP3 | pyrin domain-containing 3 |
| ASC | Apoptosis-associated Speck-like protein containing a CARD |
| CARD | caspase activation and recruitment domain |
| TGF | transforming growth factor |
| TNF | tumor necrosis factor |
| MCP-1 | Monocyte Chemotactic Protein-1 |
| CCL2 | chemokine (C-C motif) ligand 2 |
| MIP | Macrophage Inflammatory Proteins |
| Teff | effector T |
| ANCA | anti-neutrophil cytoplasmic antibody |
| PD | program death |
| sIL-2R | soluble IL-2 receptor |
| sFas | soluble Fas |
| DcR3 | decoy receptor 3 |
| TRAIL | TNF-related apoptosis-inducing ligand |
| ECM | extracellular matrix |
| Npnt | Nephronectin |
| MMP | Matrix metalloproteinase |
| ICAM | intercellular adhesion molecule |
| miR | micro RNA |
| FAK | Focal adhesion kinase |
| PI3K | Phosphoinositide 3-kinase |
| PKB | protein kinase B |
| FN | Fibronectin |
| BALF | bronchoalveolar lavage fluids |
| BNP | bone morphogenetic protein |
| BMSCs | bone marrow mesenchymal stem cells |
| SSc | systemic sclerosis |
| FGF | fibroblast growth factor |
| Ac-SDKP | N-acetyl-seryl-asoarthyl-lysyl-proline |
| SIL | silicosis patients |
| SD | standard deviation |
| HV | healthy volunteers |
| ILO | International Labor Office |
| EGF | epidermal growth factor |
| G-CSF | granulocyte colony stimulating factor |
| GM-CSF | granulocyte-macrophage-colony stimulating factor |
| IFN | Interferon |
| VEGF | vascular endothelial growth factor |
| sIL-6R | soluble IL-6 receptor |
| PR | profusion rate |
| Exp.Year | exposure years |
| %FEV1 | forced expiratory volume in 1 second |
| PFR | peak flow rate |
| FVC | percent forced vital capacity |
| v25H | 25% forced vital capacity (FVC)/Height |
| FVC | percent volume capacity |
| INFLAM | inflammatory |
| RESP | Respiratory |
| JAK | Janus kinases |
| STAT | signal transducer and activator of transcription |
| SP-D | surfactant protein–D |
References
- Cullinan, P.; Reid, P. Pneumoconiosis. Prim. Care Respir. J. 2013, 22, 249–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, C.C.; Yu, I.T.; Chen, W. Silicosis. Lancet 2012, 379, 2008–2018. [Google Scholar] [CrossRef]
- Hamilton, R.F., Jr.; Thakur, S.A.; Holian, A. Silica binding and toxicity in alveolar macrophages. Free Radic. Biol. Med. 2008, 44, 1246–1258. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, H. A mechanistic review of silica-induced inhalation toxicity. Inhal. Toxicol. 2015, 27, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Hornung, V.; Bauernfeind, F.; Halle, A.; Samstad, E.O.; Kono, H.; Rock, K.L.; Fitzgerald, K.A.; Latz, E. Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat. Immunol. 2008, 9, 847–856. [Google Scholar] [CrossRef]
- Kuroda, E.; Ishii, K.J.; Uematsu, S.; Ohata, K.; Coban, C.; Akira, S.; Aritake, K.; Urade, Y.; Morimoto, Y. Silica crystals and aluminum salts regulate the production of prostaglandin in macrophages via NALP3 inflammasome-independent mechanisms. Immunity 2011, 34, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.F., Jr.; Thakur, S.A.; Mayfair, J.K.; Holian, A. MARCO mediates silica uptake and toxicity in alveolar macrophages from C57BL/6 mice. J. Biol. Chem. 2006, 281, 34218–34226. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.A.; Hamilton, R., Jr.; Pikkarainen, T.; Holian, A. Differential binding of inorganic particles to MARCO. Toxicol. Sci. 2009, 107, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.M.; Swindle, E.J.; Kushnir-Sukhov, N.M.; Holian, A.; Metcalfe, D.D. Silica-directed mast cell activation is enhanced by scavenger receptors. Am. J. Respir. Cell Mol. Biol. 2007, 36, 43–52. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Weng, D.; Song, L.; Tang, W.; Dai, W.; Yu, Y.; Liu, F.; Zhao, M.; Lu, C.; et al. Neutralization of interleukin-17A delays progression of silica-induced lung inflammation and fibrosis in C57BL/6 mice. Toxicol. Appl. Pharmacol. 2014, 275, 62–72. [Google Scholar] [CrossRef]
- Song, L.; Weng, D.; Dai, W.; Tang, W.; Chen, S.; Li, C.; Chen, Y.; Liu, F.; Chen, J. Th17 can regulate silica-induced lung inflammation through an IL-1β-dependent mechanism. J. Cell Mol. Med. 2014, 18, 1773–1784. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Liu, F.; Li, C.; Lu, Y.; Lu, X.; Du, S.; Chen, Y.; Weng, D.; Chen, J. Blockade of Wnt/β-Catenin Pathway Aggravated Silica-Induced Lung Inflammation through Tregs Regulation on Th Immune Responses. Mediators Inflamm. 2016, 2016, 6235614. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Dai, W.; Li, C.; Liu, F.; Chen, Y.; Weng, D.; Chen, J. Baicalin Alleviates Silica-Induced Lung Inflammation and Fibrosis by Inhibiting the Th17 Response in C57BL/6 Mice. J. Nat. Prod. 2015, 78, 3049–3057. [Google Scholar] [CrossRef]
- Pétrilli, V.; Dostert, C.; Muruve, D.A.; Tschopp, J. The inflammasome: A danger sensing complex triggering innate immunity. Curr. Opin. Immunol. 2007, 19, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Sayan, M.; Mossman, B.T. The NLRP3 inflammasome in pathogenic particle and fibre-associated lung inflammation and diseases. Part. Fibre Toxicol. 2016, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Pollard, K.M. Silica, Silicosis, and Autoimmunity. Front. Immunol. 2016, 7, 97. [Google Scholar]
- Corsini, E.; Giani, A.; Peano, S.; Marinovich, M.; Galli, C.L. Resistance to silica-induced lung fibrosis in senescent rats: Role of alveolar macrophages and tumor necrosis factor-alpha (TNF). Mech. Ageing Dev. 2004, 125, 145–146. [Google Scholar] [CrossRef]
- Fazzi, F.; Njah, J.; Di Giuseppe, M.; Winnica, D.E.; Go, K.; Sala, E.; St Croix, C.M.; Watkins, S.C.; Tyurin, V.A.; Phinney, D.G.; et al. TNFR1/phox interaction and TNFR1 mitochondrial translocation Thwart silica-induced pulmonary fibrosis. J. Immunol. 2014, 192, 3837–3846. [Google Scholar] [CrossRef]
- van den Brûle, S.; Misson, P.; Bühling, F.; Lison, D.; Huaux, F. Overexpression of cathepsin K during silica-induced lung fibrosis and control by TGF-beta. Respir. Res. 2005, 6, 84. [Google Scholar] [CrossRef]
- Pryhuber, G.S.; Huyck, H.L.; Baggs, R.; Oberdörster, G.; Finkelstein, J.N. Induction of chemokines by low-dose intratracheal silica is reduced in TNFR I (p55) null mice. Toxicol. Sci. 2003, 72, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Hata, J.; Aoki, K.; Mitsuhashi, H.; Uno, H. Change in location of cytokine-induced neutrophil chemoattractants (CINCs) in pulmonary silicosis. Exp. Mol. Pathol. 2003, 75, 68–73. [Google Scholar] [CrossRef]
- Stockinger, B.; Veldhoen, M. Differentiation and function of Th17 T cells. Curr. Opin. Immunol. 2007, 19, 281–286. [Google Scholar] [CrossRef]
- Cosmi, L.; Maggi, L.; Santarlasci, V.; Liotta, F.; Annunziato, F. T helper cells plasticity in inflammation. Cytometry A 2014, 85, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hayashi, H.; Mastuzaki, H.; Kumagai-Takei, N.; Otsuki, T. Silicosis and autoimmunity. Curr. Opin. Allergy Clin. Immunol. 2017, 17, 78–84. [Google Scholar] [CrossRef]
- Ghahramani, N. Silica nephropathy. Int. J. Occup. Environ. Med. 2010, 1, 108–115. [Google Scholar]
- Schreiber, J.; Koschel, D.; Kekow, J.; Waldburg, N.; Goette, A.; Merget, R. Rheumatoid pneumoconiosis (Caplan’s syndrome). Eur. J. Intern. Med. 2010, 21, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdelghani, K.; Fazaa, A.; Souabni, L.; Zakraoui, L. Association of pulmonary silicosis and systemic sclerosis. BMJ Case Rep. 2015, 2015, bcr2013202509. [Google Scholar] [CrossRef]
- Hayashi, H.; Miura, Y.; Maeda, M.; Murakami, S.; Kumagai, N.; Nishimura, Y.; Kusaka, M.; Urakami, K.; Fujimoto, W.; Otsuki, T. Reductive alteration of the regulatory function of the CD4(+)CD25(+) T cell fraction in silicosis patients. Int. J. Immunopathol. Pharmacol. 2010, 23, 1099–1109. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Maeda, M.; Murakami, S.; Kumagai, N.; Chen, Y.; Hatayama, T.; Katoh, M.; Miyahara, N.; Yamamoto, S.; Yoshida, Y.; et al. Soluble interleukin-2 receptor as an indicator of immunological disturbance found in silicosis patients. Int. J. Immunopathol. Pharmacol. 2009, 22, 53–62. [Google Scholar] [CrossRef]
- Lee, S.; Matsuzaki, H.; Kumagai-Takei, N.; Yoshitome, K.; Maeda, M.; Chen, Y.; Kusaka, M.; Urakami, K.; Hayashi, H.; Fujimoto, W.; et al. Silica exposure and altered regulation of autoimmunity. Environ. Health Prev. Med. 2014, 19, 322–329. [Google Scholar] [CrossRef]
- Fernandes, D.J.; Bonacci, J.V.; Stewart, A.G. Extracellular matrix, integrins, and mesenchymal cell function in the airways. Curr. Drug Targets. 2006, 7, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Kadler, K.E.; Hill, A.; Canty-Laird, E.G. Collagen fibrillogenesis: Fibronectin, integrins, and minor collagens as organizers and nucleators. Curr. Opin. Cell Biol. 2008, 20, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Layseca, P.; Streuli, C.H. Signalling pathways linking integrins with cell cycle progression. Matrix Biol. 2014, 34, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Manzanares, M.; Sánchez-Madrid, F. Targeting the integrin interactome in human disease. Curr. Opin. Cell Biol. 2018, 55, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Brandenberger, R.; Schmidt, A.; Linton, J.; Wang, D.; Backus, C.; Denda, S.; Müller, U.; Reichardt, L.F. Identification and characterization of a novel extracellular matrix protein nephronectin that is associated with integrin alpha8beta1 in the embryonic kidney. J. Cell Biol. 2001, 154, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Uemura, T.; Morimitsu, K.; Sato-Nishiuchi, R.; Manabe, R.; Takagi, J.; Yamada, M.; Sekiguchi, K. Molecular basis of the recognition of nephronectin by integrin alpha8beta1. J. Biol. Chem. 2009, 284, 14524–14536. [Google Scholar] [CrossRef]
- Linton, J.M.; Martin, G.R.; Reichardt, L.F. The ECM protein nephronectin promotes kidney development via integrin alpha8beta1-mediated stimulation of Gdnf expression. Development 2007, 134, 2501–2509. [Google Scholar] [CrossRef] [PubMed]
- Toraskar, J.; Magnussen, S.N.; Chawla, K.; Svineng, G.; Steigedal, T.S. Nephronectin mediates p38 MAPK-induced cell viability via its integrin-binding enhancer motif. FEBS Open Bio. 2018, 8, 1992–2001. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Yamada, A.; Ikehata, M.; Yoshida, Y.; Sasa, K.; Morimura, N.; Sakashita, A.; Iijima, T.; Chikazu, D.; Ogata, H.; et al. FGF-2 suppresses expression of nephronectin via JNK and PI3K pathways. FEBS Open Bio. 2018, 8, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Bodo, M.; Lilli, C.; Calvitti, M.; Rosati, E.; Luca, G.; Lumare, A.; Gambelunghe, A.; Murgia, N.; Muzi, G.; Bellucci, C. Silica particle size and shape: In vitro effects on extracellular matrix metabolism and viability of human bronchial epithelial cells. J. Biol. Regul. Homeost. Agents. 2012, 26, 681–692. [Google Scholar]
- Nardi, J.; Nascimento, S.; Göethel, G.; Gauer, B.; Sauer, E.; Fão, N.; Cestonaro, L.; Peruzzi, C.; Souza, J.; Garcia, S.C. Inflammatory and oxidative stress parameters as potential early biomarkers for silicosis. Clin. Chim. Acta. 2018, 484, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Li, P.; Pan, H.; Li, Y.; Xu, Q.; Xu, T.; Ji, X.; Liu, Y.; Yao, W.; Han, L.; et al. miR-542-5p Attenuates Fibroblast Activation by Targeting Integrin α6 in Silica-Induced Pulmonary Fibrosis. Int. J. Mol. Sci. 2018, 19, pii: E3717. Int. J. Mol. Sci. 2018, 19, 3717. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.C.; Burns, J.; Munday, D.E.; McGee, J.O. Presence of fibronectin in pneumoconiotic lesions. Thorax 1982, 37, 54–56. [Google Scholar] [CrossRef]
- Bégin, R.; Massé, S.; Rola-Pleszczynski, M.; Martel, M.; Desmarais, Y.; Geoffroy, M.; LeBouffant, L.; Daniel, H.; Martin, J. Aluminum lactate treatment alters the lung biological activity of quartz. Exp. Lung Res. 1986, 10, 385–399. [Google Scholar] [CrossRef]
- Brown, G.M.; Donaldson, K.; Brown, D.M. Bronchoalveolar leukocyte response in experimental silicosis: Modulation by a soluble aluminum compound. Toxicol. Appl. Pharmacol. 1989, 101, 95–105. [Google Scholar] [CrossRef]
- Donaldson, K.; Brown, G.M.; Brown, D.M.; Slight, J.; Li, X.Y. Epithelial and extracellular matrix injury in quartz-inflamed lung: Role of the alveolar macrophage. Environ. Health Perspect. 1992, 97, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; An, G.; Wang, Y.; Liang, D.; Zhu, Z.; Lian, X.; Niu, P.; Guo, C.; Tian, L. Anti-fibrotic effects of bone morphogenetic protein-7-modified bone marrow mesenchymal stem cells on silica-induced pulmonary fibrosis. Exp. Mol. Pathol. 2017, 102, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, M.; Zardi, L.; Neri, D. Fibronectin as target for tumor therapy. Int. J. Cancer. 2006, 118, 1331–1339. [Google Scholar] [CrossRef]
- Labat-Robert, J. Fibronectin in malignancy. Semin. Cancer Biol. 2002, 12, 187–195. [Google Scholar] [CrossRef]
- Wisniowski, P.E.; Spech, R.W.; Wu, M.; Doyle, N.A.; Pasula, R.; Martin, W.J., 2nd. Vitronectin protects alveolar macrophages from silica toxicity. Am. J. Respir. Crit. Care Med. 2000, 162, 733–739. [Google Scholar] [CrossRef]
- Zetterberg, G.; Elmberger, G.; Johansson, A.; Lundahl, J.; Lundborg, M.; Sköld, C.M.; Tornling, G.; Camner, P.; Eklund, A. Rat alveolar and interstitial macrophages in the fibrosing stage following quartz exposure. Hum. Exp. Toxicol. 2000, 19, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Haustein, U.F.; Ziegler, V.; Herrmann, K.; Mehlhorn, J.; Schmidt, C. Silica-induced scleroderma. J. Am. Acad. Dermatol. 1990, 22, 444–448. [Google Scholar] [CrossRef]
- Herrmann, K.; Schulze, E.; Heckmann, M.; Schubert, I.; Meurer, M.; Ziegler, V.; Haustein, U.F.; Mehlhorn, J.; Krieg, T. Type III collagen aminopropeptide and laminin P1 levels in serum of patients with silicosis-associated and idiopathic systemic scleroderma. Br. J. Dermatol. 1990, 123, 1–7. [Google Scholar] [CrossRef]
- Bodo, M.; Muzi, G.; Bellucci, C.; Lilli, C.; Calvitti, M.; Lumare, A.; Dell’Omo, M.; Gambelunghe, A.; Baroni, T.; Murgia, N. Comparative in vitro studies on the fibrogenic effects of two samples of silica on epithelial bronchial cells. J. Biol. Regul. Homeost. Agents. 2007, 21, 97–104. [Google Scholar]
- Davis, G.S.; Holmes, C.E.; Pfeiffer, L.M.; Hemenway, D.R. Lymphocytes, lymphokines, and silicosis. J. Environ. Pathol. Toxicol. Oncol. 2001, 20, 53–65. [Google Scholar] [CrossRef]
- Xu, H.; Yang, F.; Sun, Y.; Yuan, Y.; Cheng, H.; Wei, Z.; Li, S.; Cheng, T.; Brann, D.; Wang, R. A new antifibrotic target of Ac-SDKP: Inhibition of myofibroblast differentiation in rat lung with silicosis. PLoS ONE 2012, 7, e40301. [Google Scholar] [CrossRef] [PubMed]
- Shifeng, L.; Hong, X.; Xue, Y.; Siyu, N.; Qiaodan, Z.; Dingjie, X.; Lijuan, Z.; Zhongqiu, W.; Xuemin, G.; Wenchen, C.; et al. Ac-SDKP increases α-TAT 1 and promotes the apoptosis in lung fibroblasts and epithelial cells double-stimulated with TGF-β1 and silica. Toxicol. Appl. Pharmacol. 2019, 369, 17–29. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Yang, Z.; Jia, Q.; Bo, C.; Shao, H.; Zhang, Z. Pirfenidone inhibits epithelial-mesenchymal transition and pulmonary fibrosis in the rat silicosis model. Toxicol. Lett. 2019, 300, 59–66. [Google Scholar] [CrossRef]
- Li, X.; An, G.; Wang, Y.; Liang, D.; Zhu, Z.; Tian, L. Targeted migration of bone marrow mesenchymal stem cells inhibits silica-induced pulmonary fibrosis in rats. Stem Cell Res. Ther. 2018, 9, 335. [Google Scholar] [CrossRef] [PubMed]
- El-Kashef, D.H. Nicorandil ameliorates pulmonary inflammation and fibrosis in a rat model of silicosis. Int. Immunopharmacol. 2018, 64, 289–297. [Google Scholar] [CrossRef]
- Guidelines for the use of the ILO International Classification of Radiographs of Pneumoconioses, Revised edition 2011. Available online: http://www.ilo.org/safework/info/publications/WCMS_168260/lang--en/index.htm (accessed on 25 May 2019).
- Watany, M.M.; El-Horany, H.E. Nephronectin (NPNT) and the prediction of nephrotic syndrome response to steroid treatment. Eur. J. Hum. Genet. 2018, 26, 1354–1360. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; DiGioacchino, M. MCP-1 and RANTES are mediators of acute and chronic inflammation. Allergy Asthma Proc. 2001, 22, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Gillitzer, R.; Goebeler, M. Chemokines in cutaneous wound healing. J. Leukoc. Biol. 2001, 69, 513–521. [Google Scholar]
- Menten, P.; Wuyts, A.; Van Damme, J. Macrophage inflammatory protein-1. Cytokine Growth Factor Rev. 2002, 13, 455–481. [Google Scholar] [CrossRef]
- Maurer, M.; von Stebut, E. Macrophage inflammatory protein-1. Int. J. Biochem. Cell Biol. 2004, 36, 1882–1886. [Google Scholar] [CrossRef]
- Lee, S.; Hayashi, H.; Kumagai-Takei, N.; Matsuzaki, H.; Yoshitome, K.; Nishimura, Y.; Uragami, K.; Kusaka, M.; Yamamoto, S.; Ikeda, M.; et al. Clinical evaluation of CENP-B and Scl-70 autoantibodies in silicosis patients. Exp. Ther. Med. 2017, 13, 2616–2622. [Google Scholar] [CrossRef] [Green Version]
- Otsuki, T.; Ichihara, K.; Tomokuni, A.; Sakaguchi, H.; Aikoh, T.; Matsuki, T.; Isozaki, Y.; Hyodoh, F.; Kusaka, M.; Kita, S.; et al. Evaluation of cases with silicosis using the parameters related to Fas-mediated apoptosis. Int. J. Mol. Med. 1999, 4, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Miyazono, A.; Yamada, A.; Morimura, N.; Takami, M.; Suzuki, D.; Kobayashi, M.; Tezuka, K.; Yamamoto, M.; Kamijo, R. TGF-beta suppresses POEM expression through ERK1/2 and JNK in osteoblasts. FEBS Lett. 2007, 581, 5321–5326. [Google Scholar] [CrossRef]
- Kurosawa, T.; Yamada, A.; Takami, M.; Suzuki, D.; Saito, Y.; Hiranuma, K.; Enomoto, T.; Morimura, N.; Yamamoto, M.; Iijima, T.; et al. Expression of nephronectin is inhibited by oncostatin M via both JAK/STAT and MAPK pathways. FEBS Open Biol. 2015, 5, 303–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracted Factors | |||
|---|---|---|---|
| Item | Factor 1 | Factor 2 | Factor 3 |
| Exp. Year | 0.424 | −0.040 | 0.392 |
| PR | −0.145 | 0.125 | −0.099 |
| FVC | 0.111 | −0.051 | −0.236 |
| %FEV1 | −0.063 | −0.158 | −0.126 |
| IL-1α | 0.478 | 0.635 | 0.136 |
| IL-1β | −0.159 | 0.012 | −0.055 |
| IL-2 | −0.134 | 0.669 | −0.091 |
| IL-4 | 0.791 | 0.047 | 0.032 |
| IL-8 | 0.929 | −0.047 | −0.276 |
| IL-10 | 0.552 | −0.235 | 0.237 |
| IL-17 | −0.064 | 1.003 | −0.074 |
| IFN-α2 | 0.717 | 0.124 | 0.029 |
| IFN-γ | −0.056 | 0.968 | −0.029 |
| IP-10/CXCL10 | 0.929 | −0.098 | 0.192 |
| MCP-1/CCL2 | 0.129 | −0.025 | 1.010 |
| MIP-1α | 0.643 | 0.057 | −0.526 |
| TNFα | 0.128 | 0.067 | 0.469 |
| TGFβ | 0.047 | −0.288 | −0.202 |
| Npnt | 0.132 | 0.089 | −0.456 |
| Contribution (%) | 17.973 | 17.736 | 15.192 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Honda, M.; Yamamoto, S.; Kumagai-Takei, N.; Yoshitome, K.; Nishimura, Y.; Sada, N.; Kon, S.; Otsuki, T. Role of Nephronectin in Pathophysiology of Silicosis. Int. J. Mol. Sci. 2019, 20, 2581. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102581
Lee S, Honda M, Yamamoto S, Kumagai-Takei N, Yoshitome K, Nishimura Y, Sada N, Kon S, Otsuki T. Role of Nephronectin in Pathophysiology of Silicosis. International Journal of Molecular Sciences. 2019; 20(10):2581. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102581
Chicago/Turabian StyleLee, Suni, Machiko Honda, Shoko Yamamoto, Naoko Kumagai-Takei, Kei Yoshitome, Yasumitsu Nishimura, Nagisa Sada, Shigeyuki Kon, and Takemi Otsuki. 2019. "Role of Nephronectin in Pathophysiology of Silicosis" International Journal of Molecular Sciences 20, no. 10: 2581. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102581