Salt Tolerance Improvement in Rice through Efficient SNP Marker-Assisted Selection Coupled with Speed-Breeding


 , , ,
, , ,
Abstract
:
1. Introduction
2. Results
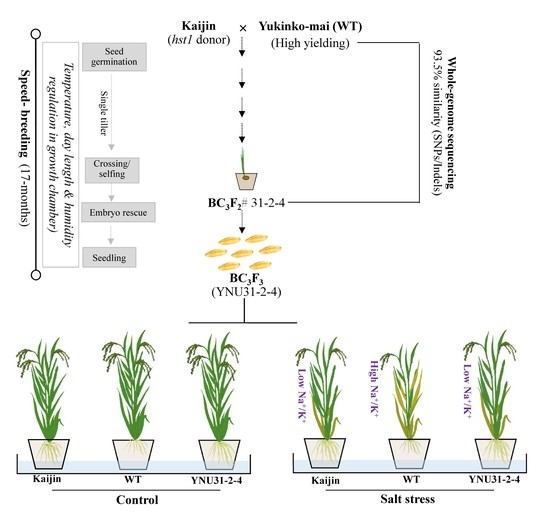
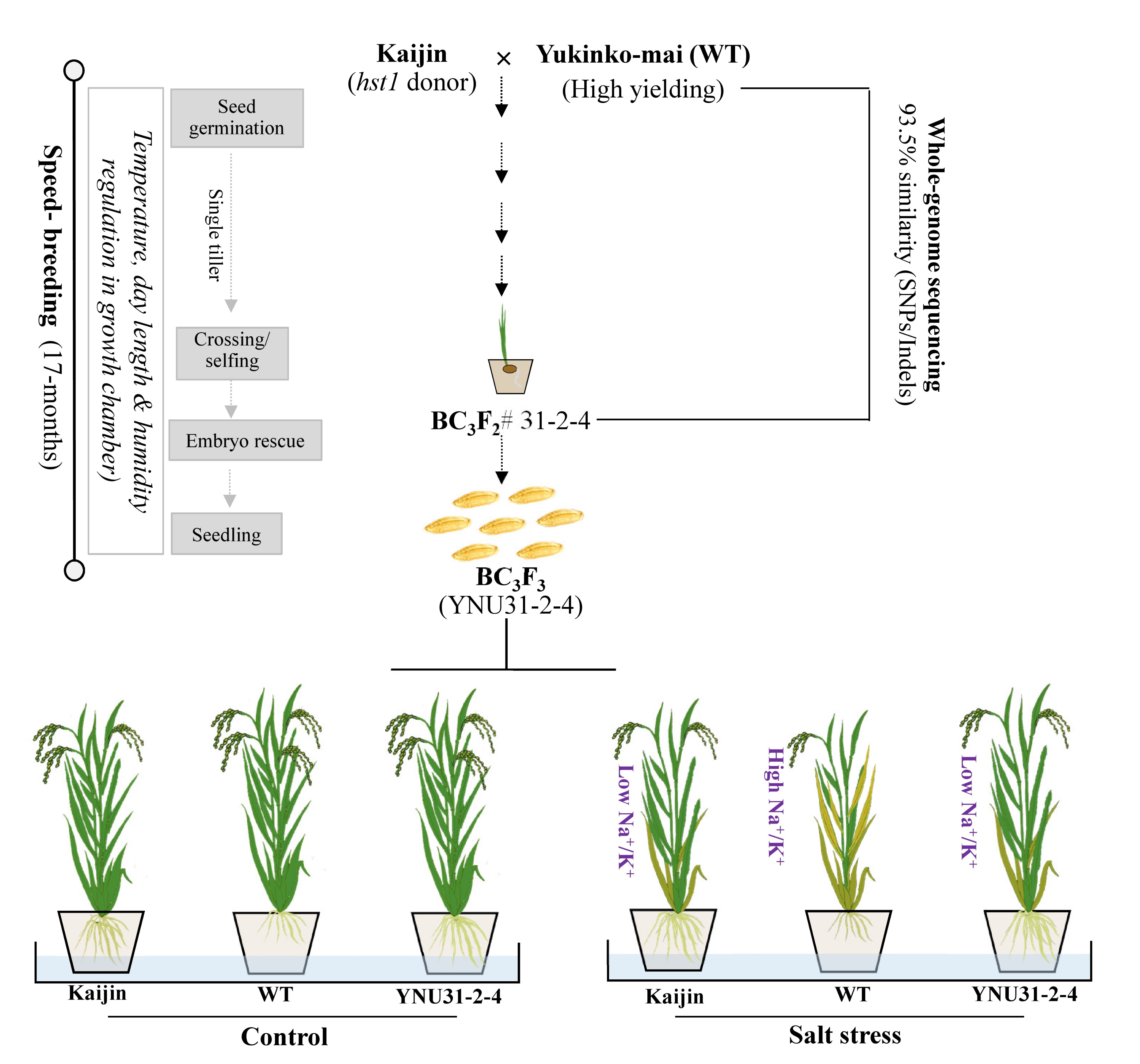
2.1. Breeding Scheme for Development of Advanced Plant Material
2.2. Introgression of hst1 into “Yukinko-mai” (WT) and Genotyping of Advanced Progeny Using SNP Marker
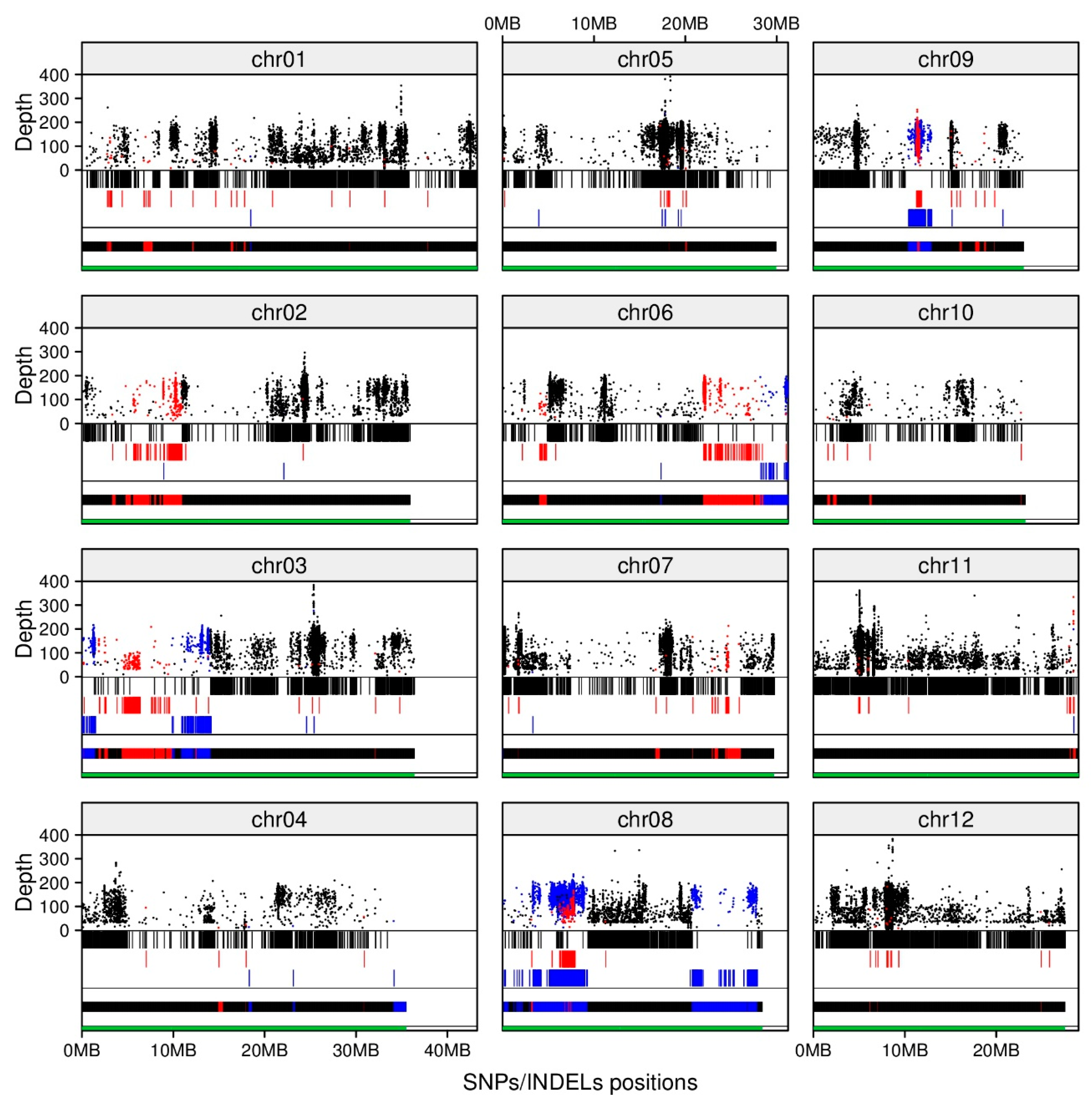
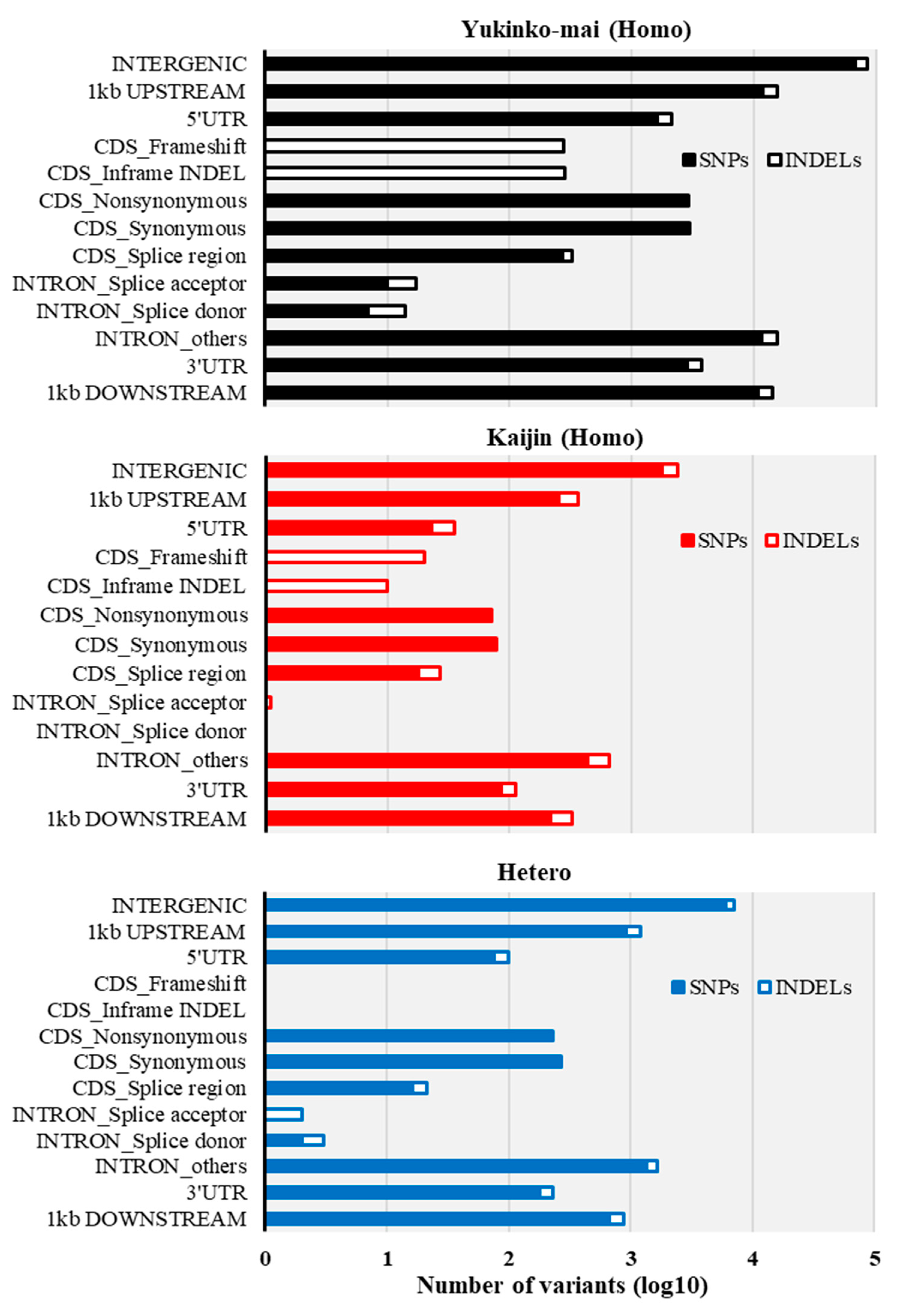
2.3. Recovery Rate and Characterization of BC3F2 #31-2-4 Genome
2.4. Field Evaluation of the “YNU31-2-4” for Main Agronomic Traits
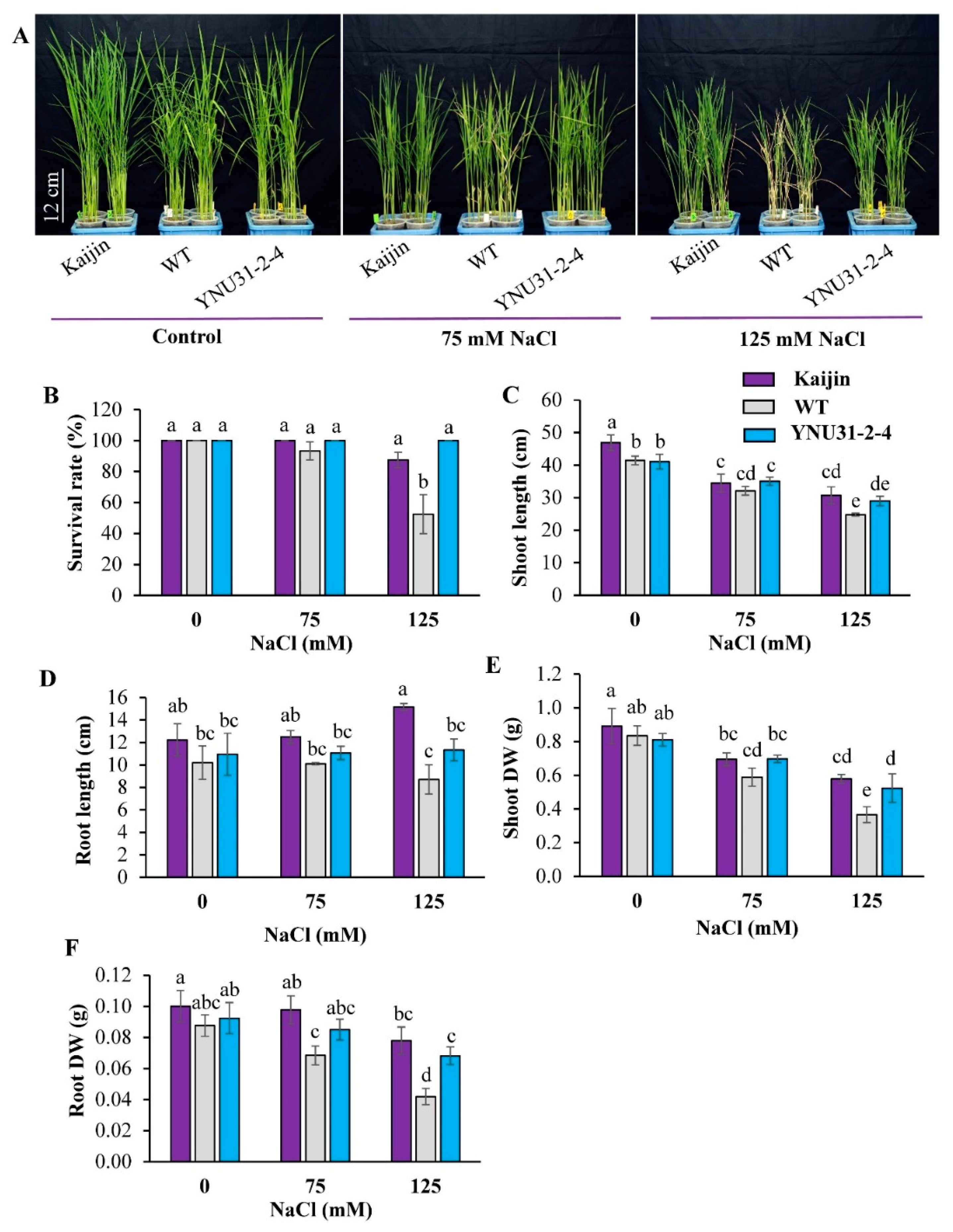
2.5. Salinity Tolerance of the “YNU31-2-4” Line at Seedling Stage
2.6. Na+ and K+ Content in Shoot and Roots of the Tested Genotypes under Salt Stress
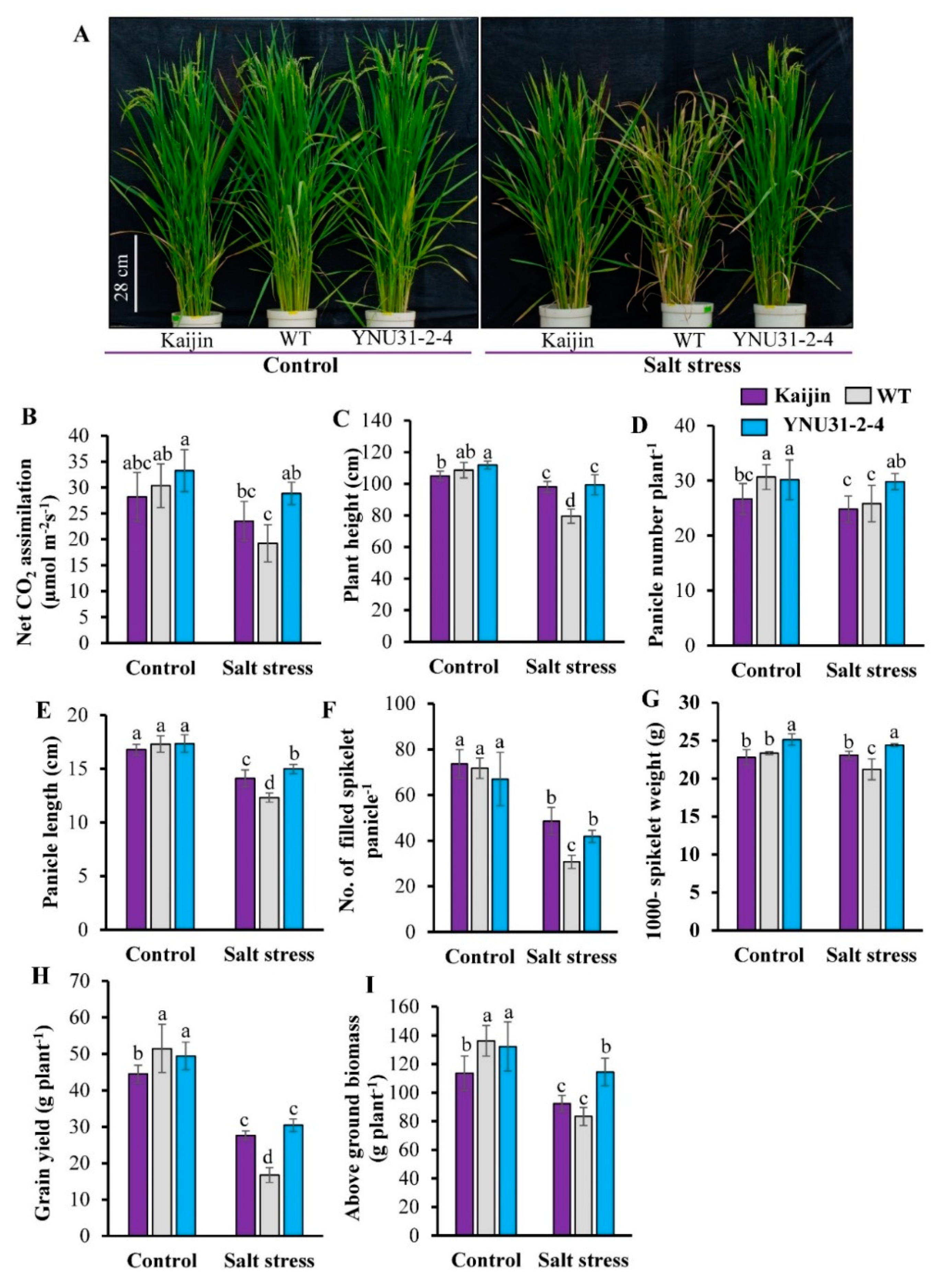
2.7. Salinity Tolerance and Yield Assessment at Reproductive Stage
3. Discussion
4. Materials and Methods
4.1. Planting Materials
4.2. Speed-Breeding–Modified Controlled-Biotron Breeding Conditions
4.3. Developing Salt-Tolerant Line by Backcrossing “Kaijin” to “Yukinko-mai”
4.4. Confirmation of Genotypes by Sanger Sequencing
4.5. DNA Library Construction and Whole-Genome Sequencing
4.6. Read Mapping, Variant Calling, and Variant Annotation
4.7. Estimation of Genome Recovery Rate
4.8. Phenotypic Evaluation under Field Condition
4.9. Growth Conditions and Evaluation of Salinity Tolerance at Seedling Stage
4.10. Determination of Leaf Relative Water Content, Chlorophyll, and Proline
4.11. Measurement of Na+ and K+ Concentrations
4.12. Evaluation of Salt-Stress Tolerance at Reproductive Stage
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wassmann, R.; Jagadish, S.V.K.; Heuer, S.; Ismail, A.; Redona, E.; Serraj, R.; Singh, R.K.; Howell, G.; Pathak, H.; Sumfleth, K. Chapter 2 Climate Change Affecting Rice Production. The Physiological and Agronomic Basis for Possible Adaptation Strategies. Adv. Agron. 2009, 101, 59–122. [Google Scholar]
- Welch, J.R.; Vincent, J.R.; Auffhammer, M.; Moya, P.F.; Dobermann, A.; Dawe, D. Rice yields in tropical/subtropical Asia exhibit large but opposing sensitivities to minimum and maximum temperatures. Proc. Natl. Acad. Sci. USA 2010, 107, 14562–14567. [Google Scholar] [CrossRef] [Green Version]
- Sreenivasulu, N.; Butardo, V.M.; Misra, G.; Cuevas, R.P.; Anacleto, R.; Kavi Kishor, P.B. Designing climate-resilient rice with ideal grain quality suited for high-temperature stress. J. Exp. Bot. 2015, 66, 1737–1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calanca, P.P. Effects of Abiotic Stress in Crop Production. In Quantification of Climate Variability, Adaptation and Mitigation for Agricultural Sustainability; Springer International Publishing: Cham, Switzerland, 2017; pp. 165–180. [Google Scholar]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef]
- United Nations. World Population Prospects: The 2012 Revision. Highlights and Advance Tables. Popul. Dev. Rev. 2013, 36, 775–801. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, P.S.; Mishra, B.; Gupta, S.R.; Rathore, A. Reproductive stage tolerance to salinity and alkalinity stresses in rice genotypes. Plant Breed. 2008, 127, 256–261. [Google Scholar] [CrossRef]
- Negrão, S.; Courtois, B.; Ahmadi, N.; Abreu, I.; Saibo, N.; Oliveira, M.M. Recent updates on salinity stress in rice: From physiological to molecular responses. CRC Crit. Rev. Plant Sci. 2011, 30, 329–377. [Google Scholar] [CrossRef]
- Marcos, M.; Sharifi, H.; Grattan, S.R.; Linquist, B.A. Spatio-temporal salinity dynamics and yield response of rice in water-seeded rice fields. Agric. Water Manag. 2018, 195, 37–46. [Google Scholar] [CrossRef]
- Todaka, D.; Nakashima, K.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Toward understanding transcriptional regulatory networks in abiotic stress responses and tolerance in rice. Rice 2012, 5, 6. [Google Scholar] [CrossRef]
- Horie, T.; Karahara, I.; Katsuhara, M. Salinity tolerance mechanisms in glycophytes: An overview with the central focus on rice plants. Rice 2012, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.J.; Negrão, S.; Tester, M. Salt resistant crop plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, I.N.B.L.; Kim, B.-K.; Yoon, I.-S.; Kim, K.-H.; Kwon, T.-R. Salt Tolerance in Rice: Focus on Mechanisms and Approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Chen, Z.C.; Yamaji, N.; Horie, T.; Che, J.; Li, J.; An, G.; Ma, J.F. A Magnesium Transporter OsMGT1 Plays a Critical Role in Salt Tolerance in Rice. Plant Physiol. 2017, 174, 1837–1849. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, D.; Mackill, D.; Deal, K.; Gregorio, G. RFLP and SSLP mapping of salinity tolerance genes in chromosome 1 of rice (Oryza sativa L.) using recombinant inbred lines. J. Agric. Food Chem. 2002, 85, 68–76. [Google Scholar]
- Thomson, M.J.; de Ocampo, M.; Egdane, J.; Rahman, M.A.; Sajise, A.G.; Adorada, D.L.; Tumimbang-Raiz, E.; Blumwald, E.; Seraj, Z.I.; Singh, R.K.; et al. Characterizing the Saltol quantitative trait locus for salinity tolerance in rice. Rice 2010, 3, 148–160. [Google Scholar] [CrossRef]
- Lin, H.X.; Zhu, M.Z.; Yano, M.; Gao, J.P.; Liang, Z.W.; Su, W.A.; Hu, X.H.; Ren, Z.H.; Chao, D.Y. QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 2004, 108, 253–260. [Google Scholar] [CrossRef]
- Ren, Z.-H.; Gao, J.-P.; Li, L.-G.; Cai, X.-L.; Huang, W.; Chao, D.-Y.; Zhu, M.-Z.; Wang, Z.-Y.; Luan, S.; Lin, H.-X. A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nat. Genet. 2005, 37, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Vu, H.T.T.; Le, D.D.; Ismail, A.M.; Le, H.H. Marker-assisted backcrossing (MABC) for improved salinity tolerance in rice (Oryza sativa L.) to cope with climate change in Vietnam. Aust. J. Crop Sci. 2012, 6, 1649–1654. [Google Scholar]
- Linh, L.H.; Linh, T.H.; Xuan, T.D.; Ham, L.H.; Ismail, A.M.; Khanh, T.D. Molecular Breeding to Improve Salt Tolerance of Rice (Oryza sativa L.) in the Red River Delta of Vietnam. Int. J. Plant Genom. 2012, 2012, 949038. [Google Scholar] [CrossRef]
- Gregorio, G.B.; Islam, M.R.; Vergara, G.V.; Thirumeni, S. Recent advances in rice science to design salinity and other abiotic stress tolerant rice varieties. Sabrao J. Breed. Genet. 2013, 45, 31–41. [Google Scholar]
- Babu, N.N.; Krishnan, S.G.; Vinod, K.K.; Krishnamurthy, S.L.; Singh, V.K.; Singh, M.P.; Singh, R.; Ellur, R.K.; Rai, V.; Bollinedi, H.; et al. Marker Aided Incorporation of Saltol, a Major QTL Associated with Seedling Stage Salt Tolerance, into Oryza sativa “Pusa Basmati 1121”. Front. Plant Sci. 2017, 8, 41. [Google Scholar] [CrossRef] [PubMed]
- Quan, R.; Wang, J.; Hui, J.; Bai, H.; Lyu, X.; Zhu, Y.; Zhang, H.; Zhang, Z.; Li, S.; Huang, R. Improvement of Salt Tolerance Using Wild Rice Genes. Front. Plant Sci. 2018, 8, 2269. [Google Scholar] [CrossRef] [Green Version]
- Negrão, S.; Cecília Almadanim, M.; Pires, I.S.; Abreu, I.A.; Maroco, J.; Courtois, B.; Gregorio, G.B.; McNally, K.L.; Margarida Oliveira, M. New allelic variants found in key rice salt-tolerance genes: An association study. Plant Biotechnol. J. 2013, 11, 87–100. [Google Scholar] [CrossRef]
- Jain, M.; Moharana, K.C.; Shankar, R.; Kumari, R.; Garg, R. Genomewide discovery of DNA polymorphisms in rice cultivars with contrasting drought and salinity stress response and their functional relevance. Plant Biotechnol. J. 2014, 12, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Thomson, M.J.; Shah-E-Alam, M.; De Ocampo, M.; Egdane, J.; Ismail, A.M. Exploring novel genetic sources of salinity tolerance in rice through molecular and physiological characterization. Ann. Bot. 2016, 117, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Sl, K.; Kumar, V.; Singh, B.; Rao, A.R.; Sv, A.M.; Rai, V.; Singh, A.K.; Singh, N.K. Mapping QTLs for Salt Tolerance in Rice (Oryza sativa L.) by Bulked Segregant Analysis of Recombinant Inbred Lines Using 50K SNP Chip. PLoS ONE 2016, 11, e0153610. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, B.; Panda, K.; Singh, B.P.; Singh, N.; Misra, P.; Rai, V.; Singh, N.K. Association of SNP Haplotypes of HKT Family Genes with Salt Tolerance in Indian Wild Rice Germplasm. Rice 2016, 9, 8. [Google Scholar] [CrossRef]
- Takagi, H.; Tamiru, M.; Abe, A.; Yoshida, K.; Uemura, A.; Yaegashi, H.; Obara, T.; Oikawa, K.; Utsushi, H.; Kanzaki, E.; et al. MutMap accelerates breeding of a salt-tolerant rice cultivar. Nat. Biotechnol. 2015, 33, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Takagi, H.; Nakajo, S.; Terauchi, R.; Kuroda, E. Yielding ability and physicochemical characteristics related to eating quality in salt-tolerant rice variety “Kaijin” under normal salt-free cultivated condition. Tohoku J. Crop Sci. 2016, 59, 19–20. [Google Scholar] [CrossRef]
- Ishizaki, K.; Matsui, T.; Kaneda, S.; Kobayashi, K.; Kasaneyama, H.; Abe, S.; Azuma, S.; Hoshi, T.; Sasaki, Y.; Hirao, K.; et al. A New Rice Cultivar “Yukinko-mai”. J. Niigata Agric. Res. Inst. 2008, 9, 89–98. [Google Scholar]
- Shiraya, T.; Mori, T.; Maruyama, T.; Sasaki, M.; Takamatsu, T.; Oikawa, K.; Itoh, K.; Kaneko, K.; Ichikawa, H.; Mitsui, T. Golgi/plastid-type manganese superoxide dismutase involved in heat-stress tolerance during grain filling of rice. Plant Biotechnol. J. 2015, 13, 1251–1263. [Google Scholar] [CrossRef]
- Lee, K.-S.; Choi, W.-Y.; Ko, J.-C.; Kim, T.-S.; Gregorio, G.B. Salinity tolerance of japonica and indica rice (Oryza sativa L.) at the seedling stage. Planta 2003, 216, 1043–1046. [Google Scholar] [CrossRef]
- Roy, K.; Sasada, K.; Kohno, E. Salinity status of the 2011 Tohoku-oki tsunami affected agricultural lands in northeast Japan. Int. Soil Water Conserv. Res. 2014, 2, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Kurotani, K.; Yamanaka, K.; Toda, Y.; Ogawa, D.; Tanaka, M.; Kozawa, H.; Nakamura, H.; Hakata, M.; Ichikawa, H.; Hattori, T.; et al. Stress Tolerance Profiling of a Collection of Extant Salt-Tolerant Rice Varieties and Transgenic Plants Overexpressing Abiotic Stress Tolerance Genes. Plant Cell Physiol. 2015, 56, 1867–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collard, B.C.Y.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 557–572. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Latif, M.A. Recurrent parent genome recovery analysis in a marker-assisted backcrossing program of rice (Oryza sativa L.). C. R. Biol. 2015, 338, 83–94. [Google Scholar] [CrossRef]
- Fukuoka, S.; Ebana, K.; Yamamoto, T.; Yano, M. Integration of Genomics into Rice Breeding. Rice 2010, 3, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Ma, T.; Zhang, A.; Ong, K.H.; Li, Z.; Yang, J.; Yin, Z. Marker-assisted breeding of the rice restorer line Wanhui 6725 for disease resistance, submergence tolerance and aromatic fragrance. Rice 2016, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Das, G.; Patra, J.K.; Baek, K.-H. Insight into MAS: A Molecular Tool for Development of Stress Resistant and Quality of Rice through Gene Stacking. Front. Plant Sci. 2017, 8, 985. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, T.; Yoshino, M.; Yamakawa, H.; Kinoshita, T. The biotron breeding system: A rapid and reliable procedure for genetic studies and breeding in rice. Plant Cell Physiol. 2011, 52, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, J.; Hayashi, T.; Iwata, H. A practical, rapid generation-advancement system for rice breeding using simplified biotron breeding system. Breed. Sci. 2016, 66, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Collard, B.C.Y.; Beredo, J.C.; Lenaerts, B.; Mendoza, R.; Santelices, R.; Lopena, V.; Verdeprado, H.; Raghavan, C.; Gregorio, G.B.; Vial, L.; et al. Revisiting rice breeding methods—Evaluating the use of rapid generation advance (RGA) for routine rice breeding. Plant Prod. Sci. 2017, 20, 337–352. [Google Scholar] [CrossRef]
- Watson, A.; Ghosh, S.; Williams, M.J.; Cuddy, W.S.; Simmonds, J.; Rey, M.D.; Asyraf Md Hatta, M.; Hinchliffe, A.; Steed, A.; Reynolds, D.; et al. Speed breeding is a powerful tool to accelerate crop research and breeding. Nat. Plants 2018, 4, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Nagatoshi, Y.; Fujita, Y. Accelerating Soybean Breeding in a CO2-Supplemented Growth Chamber. Plant Cell Physiol. 2019, 60, 77–84. [Google Scholar] [CrossRef] [PubMed]
- De Haas, L.S.; Koopmans, R.; Lelivelt, C.L.C.; Ursem, R.; Dirks, R.; Velikkakam James, G. Low-coverage resequencing detects meiotic recombination pattern and features in tomato RILs. DNA Res. 2017, 24, 549–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Si, W.; Yuan, Y.; Huang, J.; Zhang, X.; Zhang, Y.; Zhang, Y.; Tian, D.; Wang, C.; Yang, Y.; Yang, S. Widely distributed hot and cold spots in meiotic recombination as shown by the sequencing of rice F2 plants. New Phytol. 2015, 206, 1491–1502. [Google Scholar] [CrossRef]
- Yamamoto, E.; Yonemaru, J.-I.; Yamamoto, T.; Yano, M. OGRO: The Overview of functionally characterized Genes in Rice online database. Rice 2012, 5, 26. [Google Scholar] [CrossRef]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.C.; Hu, L.; Yamasaki, M.; Yoshida, S.; Kitano, H.; Hirano, K.; et al. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and Salt Tolerance. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef] [PubMed]
- Inui, T.; Yasutaka, T.; Endo, K.; Katsumi, T. Geo-environmental issues induced by the 2011 off the Pacific Coast of Tohoku Earthquake and tsunami. Soils Found. 2012, 52, 856–871. [Google Scholar] [CrossRef] [Green Version]
- Iftekharuddaula, K.M.; Salam, M.A.; Newaz, M.A.; Ahmed, H.U.; Collard, B.C.Y.; Septiningsih, E.M.; Sanchez, D.L.; Pamplona, A.M.; Mackill, D.J. Comparison of phenotypic versus marker-assisted background selection for the SUB1 QTL during backcrossing in rice. Breed. Sci. 2012, 62, 216–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, M.M.; Rafii, M.Y.; Ismail, M.R.; Mahmood, M.; Rahim, H.A.; Alam, M.A.; Ashkani, S.; Malek, M.A.; Latif, M.A. Marker-assisted backcrossing: A useful method for rice improvement. Biotechnol. Biotechnol. Equip. 2015, 29, 237–254. [Google Scholar] [CrossRef]
- Tanweer, F.A.; Rafii, M.Y.; Sijam, K.; Rahim, H.A.; Ahmed, F.; Ashkani, S.; Latif, M.A. Introgression of Blast Resistance Genes (Putative Pi-b and Pi-kh) into Elite Rice Cultivar MR219 through Marker-Assisted Selection. Front. Plant Sci. 2015, 6, 1002. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Septiningsih, E.M.; Kohli, A.; Thomson, M.J.; Ye, C.; Redoña, E.; Kumar, A.; Gregorio, G.B.; Wassmann, R.; Ismail, A.M.; et al. Genetic Advances in Adapting Rice to a Rapidly Changing Climate. J. Agron. Crop Sci. 2012, 198, 360–373. [Google Scholar] [CrossRef]
- Ma, Y.-C.; Augé, R.M.; Dong, C.; Cheng, Z.-M.M. Increased salt tolerance with overexpression of cation/proton antiporter 1 genes: A meta-analysis. Plant Biotechnol. J. 2017, 15, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Saxena, S.C.; Kaur, H.; Verma, P.; Petla, B.P.; Andugula, V.R.; Majee, M. Osmoprotectants: Potential for Crop Improvement Under Adverse Conditions. In Plant Acclimation to Environmental Stress; Springer: New York, NY, USA, 2013; pp. 197–232. [Google Scholar]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Zeng, L.; Shannon, M.C. Salinity Effects on Seedling Growth and Yield Components of Rice. Crop Sci. 2000, 40, 996. [Google Scholar] [CrossRef]
- Walia, H.; Wilson, C.; Zeng, L.; Ismail, A.M.; Condamine, P.; Close, T.J. Genome-wide transcriptional analysis of salinity stressed japonica and indica rice genotypes during panicle initiation stage. Plant Mol. Biol. 2007, 63, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-B.; Liu, C.; Tang, D.-Y.; Yan, L.; Wang, D.; Yang, Y.-Z.; Gui, J.-S.; Zhao, X.-Y.; Li, L.-G.; Tang, X.-D.; et al. The Receptor-Like Cytoplasmic Kinase STRK1 Phosphorylates and Activates CatC, Thereby Regulating H2O2 Homeostasis and Improving Salt Tolerance in Rice. Plant Cell 2018, 30, 1100–1118. [Google Scholar] [CrossRef]
- Murchie, E.H.; Yang, J.; Hubbart, S.; Horton, P.; Peng, S. Are there associations between grain-filling rate and photosynthesis in the flag leaves of field-grown rice? J. Exp. Bot. 2002, 53, 2217–2224. [Google Scholar] [CrossRef]
- Zhai, H.; Cao, S.; Wan, J.; Zhang, R.; Lu, W.; Li, L.; Kuang, T.; Min, S.; Zhu, D.; Cheng, S. Relationship between leaf photosynthetic function at grain filling stage and yield in super high-yielding hybrid rice (Oryza sativa L). Sci. China Ser. C Life Sci. 2002, 45, 637–646. [Google Scholar] [CrossRef]
- Kasai, M. Regulation of leaf photosynthetic rate correlating with leaf carbohydrate status and activation state of Rubisco under a variety of photosynthetic source/sink balances. Physiol. Plant. 2008, 134, 216–226. [Google Scholar] [CrossRef]
- Sanchez-Bragado, R.; Elazab, A.; Zhou, B.; Serret, M.D.; Bort, J.; Nieto-Taladriz, M.T.; Araus, J.L. Contribution of the ear and the flag leaf to grain filling in durum wheat inferred from the carbon isotope signature: Genotypic and growing conditions effects. J. Integr. Plant Biol. 2014, 56, 444–454. [Google Scholar] [CrossRef]
- Zhou, W.; Lv, T.; Yang, Z.; Wang, T.; Fu, Y.; Chen, Y.; Hu, B.; Ren, W. Morphophysiological mechanism of rice yield increase in response to optimized nitrogen management. Sci. Rep. 2017, 7, 17226. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, R.-J.; Han, T.-T.; Cai, W.; Fu, Z.-W.; Lu, Y.-T. Salt stress reduces root meristem size by nitric oxide-mediated modulation of auxin accumulation and signaling in Arabidopsis. Plant Physiol. 2015, 168, 343–356. [Google Scholar] [CrossRef]
- Yu, J.; Zhao, W.; Tong, W.; He, Q.; Yoon, M.-Y.; Li, F.-P.; Choi, B.; Heo, E.-B.; Kim, K.-W.; Park, Y.-J.; et al. A Genome-Wide Association Study Reveals Candidate Genes Related to Salt Tolerance in Rice (Oryza sativa) at the Germination Stage. Int. J. Mol. Sci. 2018, 19, 3145. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Maggio, A.; Bressan, R.; Yun, D.-J.; Ali, A.; Maggio, A.; Bressan, R.A.; Yun, D.-J. Role and Functional Differences of HKT1-Type Transporters in Plants under Salt Stress. Int. J. Mol. Sci. 2019, 20, 1059. [Google Scholar] [CrossRef]
- Okada, T.; Nakayama, H.; Shinmyo, A.; Yoshida, K. Expression of OsHAK genes encoding potassium ion transporters in rice. Plant Biotechnol. 2008, 25, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Hauser, F.; Horie, T. A conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant. Cell Environ. 2010, 33, 552–565. [Google Scholar] [CrossRef]
- Chen, Z.; Pottosin, I.I.; Cuin, T.A.; Fuglsang, A.T.; Tester, M.; Jha, D.; Zepeda-Jazo, I.; Zhou, M.; Palmgren, M.G.; Newman, I.A.; et al. Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 2007, 145, 1714–1725. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Walbot, V.; Warren, C. Regulation of Mu element copy number in maize lines with an active or inactive Mutator transposable element system. Mol. Gen. Genet. 1988, 211, 27–34. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Gtrellis: An R/Bioconductor package for making genome-level Trellis graphics. BMC Bioinform. 2016, 17, 169. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd ed.; International Rice Research Institute: Manila, Philippines, 1976; pp. 61–66. [Google Scholar]
- Sade, N.; Vinocur, B.J.; Diber, A.; Shatil, A.; Ronen, G.; Nissan, H.; Wallach, R.; Karchi, H.; Moshelion, M. Improving plant stress tolerance and yield production: Is the tonoplast aquaporin SlTIP2;2 a key to isohydric to anisohydric conversion? New Phytol. 2009, 181, 651–661. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Pequerul, A.; Pérez, C.; Madero, P.; Val, J.; Monge, E. A rapid wet digestion method for plant analysis. In Optimization of Plant Nutrition; Springer: Dordrecht, The Netherlands, 1993; pp. 3–6. [Google Scholar]
- Mitsui, T.; Loboda, T.; Itoh, A.; Ikarashi, T. Sugar-Controlled Ca2+ Uptake and ɑ-Amylase Secretion in Cultured Cells of Rice (Oryza sativa L.). Plant Cell Physiol. 1999, 40, 884–893. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Number of Variants | Genome % | ||
|---|---|---|---|---|
| SNPs | Indels | Total | ||
| ‘Yukinko-mai” (homo) | 84 927 | 21 361 | 106 288 | 89.7 |
| “Kaijin” (homo) | 2329 | 828 | 3157 | 2.7 |
| Hetero | 7556 | 1453 | 9009 | 7.6 |
| Total | 94 812 | 23 642 | 118 454 | 100.0 |
| Chr./Position | Ref. SNP/Indel | Genotype | Alteration Type | RAP ID | Protein Encoded | Dataset 1 | Dataset 2 | ||
|---|---|---|---|---|---|---|---|---|---|
| BC3F2 | “Kaijin” | “Yukinko-mai” | |||||||
| Chr06/26277010 | G | A | A | G | missense SNP | Os06g0644200 | Vacuolar HD-translocating inorganic pyrophosphatase 1 | Cold tolerance | – |
| Chr08/6268486 | GCACGGCCACGGC | Hetero | GCACGGCCACGGC | G | in-frame deletion | Os08g0207500 | Zn-regulated transporter, iron (Fe)-regulated transporter–like protein 4 | Other soil stress tolerance | – |
| Chr08/26913261 | G | Hetero | G | A | missense SNP | Os08g0538300 | Chitin elicitor receptor kinase 1 | Blast resistance | – |
| Chr09/11449688 | G | Hetero | G | A | missense SNP | Os09g0356200 | Malectin-like carbohydrate-binding domain–containing protein | – | Days to heading |
| Chr09/11449800 | AGG | Hetero | AGG | AC | frameshift deletion | – | |||
| Agronomic Traits | Genotype | ||
|---|---|---|---|
| “Kaijin” | WT | “YNU31-2-4” | |
| Flag leaf greenness (SPAD value) | 42.93 ± 2.63 a | 43.49 ± 1.82 a | 43.24 ± 2.42 a |
| Flag leaf length (cm) | 27.21 ± 2.53 b | 29.85 ± 2.13 a | 29.12 ± 1.85 a |
| Flag leaf width (cm) | 1.11 ± 0.06 b | 1.23 ± 0.10 a | 1.21 ± 0.04 a |
| Days-to-heading (day) | 105.17 ± 1.40 a | 105.08 ± 1.38 a | 103.75 ± 1.76 b |
| Plant height (cm) | 98.42 ± 3.56 a | 94.75 ± 3.49 b | 95.83 ± 2.41 ab |
| Tiller number per plant | 20.40 ± 3.31 b | 23.80 ± 3.22 a | 24.40 ± 3.57 a |
| Panicle number per plant | 20.40 ± 3.31 b | 23.80 ± 3.22 a | 24.20 ± 3.12 a |
| Panicle length (cm) | 18.71 ± 0.69 b | 19.79 ± 0.52 a | 19.11 ± 0.97 ab |
| Spikelet number per panicle | 71.99 ± 8.91 a | 73.55 ± 6.27 a | 72.66 ± 5.80 a |
| Grain number per panicle | 63.61 ± 8.20 a | 67.36 ± 5.81 a | 60.59 ± 7.29 a |
| 1000-grain weight (g) | 22.42 ± 0.14 b | 21.56 ± 0.22 c | 23.29 ± 0.43 a |
| Seed setting rate (%) | 88.34 ± 3.71 a | 91.61 ± 2.52 a | 83.43 ± 7.58 b |
| Grain yield (g per plant) | 33.77 ± 4.24 b | 40.07 ± 4.59 a | 37.84 ± 4.46 a |
| Aboveground biomass (g per plant) | 61.80 ± 8.50 b | 72.30 ± 7.63 a | 67.13 ± 9.14 ab |
| Grain length (mm) | 5.05 ± 0.03 b | 5.17 ± 0.01 a | 5.21 ± 0.06 a |
| Grain width (mm) | 2.78 ± 0.01 b | 2.66 ± 0.01 c | 2.86 ± 0.04 a |
| Grain thickness (mm) | 2.14 ± 0.01 a | 2.07 ± 0.01 c | 2.10 ± 0.01 b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rana, M.M.; Takamatsu, T.; Baslam, M.; Kaneko, K.; Itoh, K.; Harada, N.; Sugiyama, T.; Ohnishi, T.; Kinoshita, T.; Takagi, H.; et al. Salt Tolerance Improvement in Rice through Efficient SNP Marker-Assisted Selection Coupled with Speed-Breeding. Int. J. Mol. Sci. 2019, 20, 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102585
Rana MM, Takamatsu T, Baslam M, Kaneko K, Itoh K, Harada N, Sugiyama T, Ohnishi T, Kinoshita T, Takagi H, et al. Salt Tolerance Improvement in Rice through Efficient SNP Marker-Assisted Selection Coupled with Speed-Breeding. International Journal of Molecular Sciences. 2019; 20(10):2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102585
Chicago/Turabian StyleRana, Md Masud, Takeshi Takamatsu, Marouane Baslam, Kentaro Kaneko, Kimiko Itoh, Naoki Harada, Toshie Sugiyama, Takayuki Ohnishi, Tetsu Kinoshita, Hiroki Takagi, and et al. 2019. "Salt Tolerance Improvement in Rice through Efficient SNP Marker-Assisted Selection Coupled with Speed-Breeding" International Journal of Molecular Sciences 20, no. 10: 2585. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102585