The Biology of the Nuclear Envelope and Its Implications in Cancer Biology


1
Molecular Pathology, Department of Translational Medicine, Lund University, Skåne University Hospital, 20502 Malmö, Sweden
2
Laboratory of Molecular Cell Biomedicine, University of the Balearic Islands, 07121 Palma de Mallorca, Spain
3
Lipopharma Therapeutics, Isaac Newton, 07121 Palma de Mallorca, Spain
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2019, 20(10), 2586; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102586
Submission received: 23 March 2019
/
Revised: 7 May 2019
/
Accepted: 25 May 2019
/
Published: 27 May 2019
(This article belongs to the Special Issue Molecular Regulation of the Endomembrane System)
{kind=link}
Abstract
:The formation of the nuclear envelope and the subsequent compartmentalization of the genome is a defining feature of eukaryotes. Traditionally, the nuclear envelope was purely viewed as a physical barrier to preserve genetic material in eukaryotic cells. However, in the last few decades, it has been revealed to be a critical cellular component in controlling gene expression and has been implicated in several human diseases. In cancer, the relevance of the cell nucleus was first reported in the mid-1800s when an altered nuclear morphology was observed in tumor cells. This review aims to give a current and comprehensive view of the role of the nuclear envelope on cancer first by recapitulating the changes of the nuclear envelope during cell division, second, by reviewing the role of the nuclear envelope in cell cycle regulation, signaling, and the regulation of the genome, and finally, by addressing the nuclear envelope link to cell migration and metastasis and its use in cancer prognosis.
Keywords:
nucleus; envelope; γ-tubulin; mitosis; cell cycle; signaling; genome; cancer; diagnosis; metastasis1. Introduction
Papanicolaou’s smear test [1] is a routine check for uterine and cervical cancer. In addition, it has been implemented for a variety of specimens, such as fine needle aspiration biopsies, cerebrospinal fluid, pleural fluid, and urine samples, among others [2,3]. This cancer diagnosis test is based on a combination of changes in the staining, size, and shape of the nuclear chromatin. The eukaryotic nucleus encloses and regulates this chromatin by using a double membrane (nuclear envelope, NE), which, in turn, provides a unique molecular and biochemical environmental protective mechanism against potentially damaging cytoplasmic enzymatic activities, such as oxidative metabolism.
NE includes three interconnected domains with morphological differences: The inner nuclear membrane (INM) and outer nuclear membrane (ONM) and the pore membranes. These two individual lipid bilayers are separated by a luminal space of 30–50 nm in human cells, named the lumen or perinuclear space. For the transport of macromolecules in and out of the nucleus, both NM connect to form pore membranes where the nuclear pore complexes (NPCs) responsible for transport regulation are inserted [4,5,6]. Indeed, INM and ONM form discrete domains of a single membrane system separated by the NPCs [4,5]. The ONM is contiguous with the rough endoplasmic reticulum (ER) and contains ribosomes on its outer surface [4,5]. Furthermore, the ONM is connected to the cytoskeleton through its integral proteins, and in turn, these proteins connect to the luminal parts of INM proteins at the luminal space, all in all connecting the cytoskeleton to the nucleoskeleton and chromatin. Although INM, ONM, pore membranes, and ER originate from a continuous structure, they maintain their identities to a large extent through unique profiles of integral-membrane and other associated proteins, together with specific compositions of lipids and cholesterol [7,8,9]. In addition, the protein composition of the nuclear envelope membranes is variable across different mammalian tissues (reviewed in Reference [10]).
In metazoan organisms, the primary scaffold of NE is provided by a fibrous layer called the nuclear lamina. This thick protein layer that underlies the INM has been described in studies with isolated NEs, since nuclear lamina is resistant to most chemical extractions used in biology. The most abundant proteins of the nuclear lamina are, by far, three polypeptides of around 65–70 kDa that correspond to type V intermediate filament proteins and are named lamins [11,12]. It is estimated that there are roughly 3,000,000 copies of lamins in a typical mammalian nucleus [13]. The specific lamin nucleoskeleton is distinct from the nuclear matrix that supports chromatin inside the nucleus. Four lamin proteins are expressed in mammalian somatic cells. A-type lamins (lamins A and C) are produced by alternative mRNA splicing of LMNA gene, whereas B-type lamins (lamin B1 and B2) are encoded, respectively, by LMNB1 and LMNB2 genes. Lamins C2 and B3 are germ-cell-specific isoforms produced by alternative splicing of LMNA and LMNB1, respectively. B-type lamin expression occurs early in embryonic development and persists ubiquitously through adult life. In contrast, the A-type lamins are expressed in an asynchronous and developmentally regulated manner and are only detected after tissue differentiation (reviewed in [14]). Indeed, before day 10 of murine embryonic development, no A-type lamins are detected in the embryo proper [15,16]. The last constituent of the nuclear lamina is a collection of integral and associated proteins of the INM.
The gatekeeper function necessary for the translocation of proteins in and out of the nucleus is controlled principally by the NPC. The NPCs are large assemblies of >60 MDa in mammals. Several copies of a set of 30 diverse proteins, termed nucleoporins (NUPs), are found in these structures (reviewed in [6]). Only a restricted number of structural domains are found in the nucleoporins sequence. Those domains are transmembrane domains, Phe-Gly (FG) repeats, WD domains, α-helices, and β-propellers [17,18,19]. Around 2000–3000 NPC units are found in an average mammalian nucleus. FG-NUPs, in particular, play the most relevant role in defining the NPC diffusion limit and the list of nuclear transport receptors that might be shuttled through the NPC ([20] and reviewed in Ref. [21]). Indeed, NPCs are not a mere gateway to the nucleus. First, the size and maturation of nuclear pores is a crucial event to the nuclear import and nuclear growth and size at the end of mitosis [22,23], reviewed in Ref. [24] and during interphase [23,25,26], as shown in several vertebrate models. Second, NPCs might alter nuclear morphology through the physical link with lamins [27]. Third, nucleoporins are a model of long-term protein endurance: NPCs are sustained over the lifetime of a cell by means of a slow but finite interchange of its steadily more stable subcomplexes [28]. In tumorigenesis, the incidental finding of Nup88 as a biomarker of cancer [29,30] opened the door to detection of high levels of Nup88 in several types of tumors [30,31,32,33,34,35,36]. Beyond this protein, only a small number of nucleoporins have been associated with tumorigenesis. These include proteins Nup62, Nup88, Nup98, Nup214, and Nup358/RanBP2, all of them elements of the trafficking pathway and are specifically related to the export of mRNA (reviewed in Ref. [37]). In the literature, increases in the number of NPCs were related with more aggressive tumors [38,39,40,41].
In addition, nuclear envelope transmembrane (NET) proteins are also found both INM and ONM. This host of integral proteins might be tissue-specific for up to 60% of their protein elements [42]. Most NETs bind lamins and interact with chromatin (reviewed in [43]). The most known NET is emerin, which is encoded by the EMD gene in humans. Emerin is a founding member of the LEM domain-containing integral proteins of the inner nuclear membrane in vertebrates, where LEM is named for LAP2, emerin, and MAN1 (reviewed in [44]). Emerin is highly expressed in cardiac and skeletal muscle and several mutations affecting this gene cause X-linked recessive Emery–Dreifuss muscular dystrophy (EDMD).
More than preserving genetic material and providing architecture and mechanical support in eukaryotic cells, the NE is a key cellular hub that plays a dynamic role in the control of cell cycle regulation, mitosis, apoptosis, DNA repair, ageing, nuclear architecture, signaling, chromatin organization, gene expression regulation, and cell migration [45,46,47,48,49,50,51]. All of these various functions are critical for the processes of tumorigenesis, tumor growth, and metastasis. All in all, it is reasonable that the diagnosis of cancer relies on morphologically distinctive alterations in the NE that are only recognizable by the eye of a well-trained pathologist.
2. The Nuclear Envelope in Cell Division
Cancer is the result of uncontrolled cell division. NE proteins can mostly affect the cell cycle in higher eukaryotes when the cells undergo open mitosis and the nucleus architecture is dismantled to allow the partitioning of the genetic material between the daughter cells. Indeed, the finding in mammalian cells of the depolymerization of lamin polymers upon hyperphosphorylation of lamin A at the onset of mitosis was the first clue in NE regulating cell division [52,53,54,55].
2.1. Nuclear Envelope Disassembly at the Onset of Cell Division
Phosphorylation of NE proteins and of their binding partners drives the coordinated disruption of NE interactions and structures at the beginning of mitosis. Together with lamins, several NPC proteins and NETs are also phosphorylated by mitotic kinases (gp210, LAP2β, and lamin B receptor –LBR), as shown in human, murine, or avian models [56,57,58,59]. In human and Caenorhabditis elegans cells, the same occurs with barrier-to-autointegration-factor (BAF), a chromatin binding partner of several NETs connecting chromatin to NE [60,61] (reviewed in [62]).
Disassembly of the NE needs close coordination with the generation of the bipolar mitotic spindle. In prophase, NPC-attached dynein motors assist in the separation of the centrosomes [63,64].
Disassembly of NPCs is not a straight reversal of the assembly steps (reviewed in [65,66]). In many cases, components of the NE and NPCs actively participate in mitotic events when released from their interphase organization [67]. At the G2/M cell cycle transition, two nucleoporins participate in tethering centrosomes to the NE [68,69]. During prophase, these interactions might help microtubules in their function for NE breakdown [70] and for moving of sister centrosomes to opposite sides of the nucleus [68,69,71]. At the end of prophase, the NPC is dismantled releasing elements with important regulatory functions during mitosis: NUP358 at kinetochore functioning [72,73,74], NUP88 and other nucleoporins interfering with microtubule dynamics to promote spindle assembly, NUP98 in regulating the adenomatous polyposis coli (APC)/C [67,75,76,77] (reviewed in [65]).
During mitosis in animal cells, remodeled nuclear membranes intermix in a large part with the tubulo-vesicular mitotic ER [78], while NE vesiculation also occurs [79,80,81,82,83,84,85,86]. Regarding the over 100 different NETs in any given cell, several of them go into a storage form and others exert critical functions, such as RanGTP, the transport receptor importin/karyopherinβ, and RepoMan ([87], reviewed in Ref. [6,88,89]).
Several features of cancer cells, such as lagging chromosomes, aneuploidy, and polyploidy, might occur after a failed NE breakdown at the onset of mitosis and the subsequent blocking of spindle assembly.
2.2. Nuclear Assembly After Cell Division
2.2.1. Chromatin Enclosing, INM Protein Recruitment, and NPC Formation
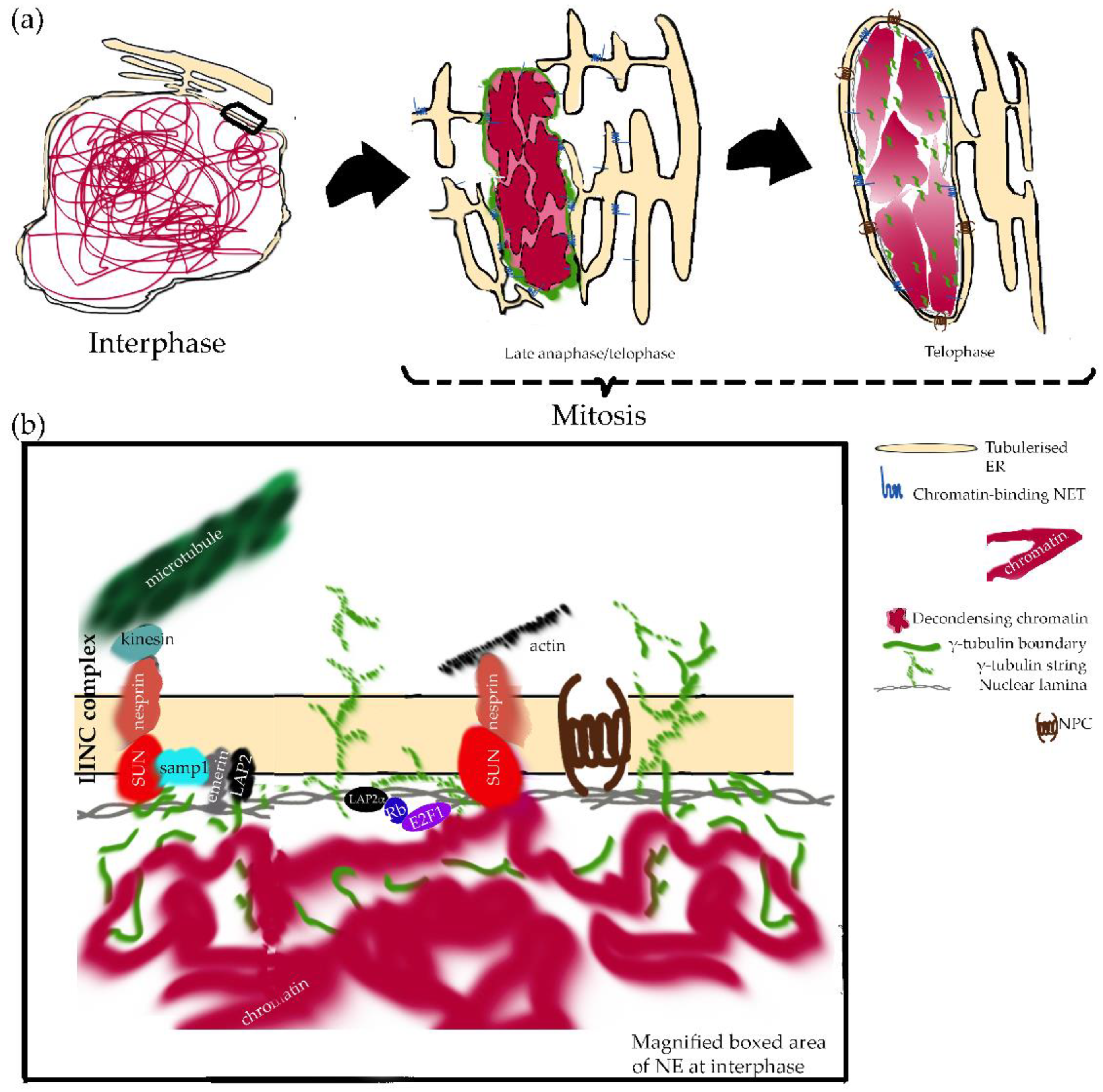
During metazoan anaphase, chromosomes cluster compactly together in a disc-like configuration whose surface drives nuclear assembly. During early telophase, NE reassembly is initiated by changes at this chromatin surface (i.e., removal of mitotic histone marks by phosphatases) and the dephosphorylation-induced binding of NETs and their associated membranes to chromatin (reviewed in [78]).
Several mechanisms combine to recruit ONM and INM proteins, constituents of NPCs, and lamins. In metazoa, INM proteins are attracted both by both specific interactions and by the general affinity of many INM proteins for chromatin/DNA, for example, LBR binds heterochromatin protein 1 (HP1) [90,91] and histone H3 [92,93,94]. Importantly, NET and NPC binding to mitotic chromosomes in early telophase seems to drive NE reassembly [95,96,97,98,99], implicating phenylalanine and glycine (FG)-rich nucleoporins and the AT-hook-domain containing protein, ELYS/Mel-28 (Figure 1). ELYS localizes not only to NPCs, but is also associated with chromosomal kinetochores during cell division. At the end of mitosis, ELYS recruits the NUP107-160 subcomplex, which is required for the correct segregation of mitotic chromosomes [100,101]. In addition, in NPC assembly and chromatin decondensation, lysine demethylase LSD1 is required [102], while the Repo-Man-promoted dephosphorylation of histone H3 seems indispensable for targeting importin-b to mitotic chromatin [103] (Figure 1).
Engulfment of recently separated sister chromatids by the NE occurs in an astonishingly short timeframe thanks to deposits of membrane fragments on the chromatin surface that trigger the enveloping process (reviewed in [78,104]). It is still a matter of debate whether ER moves toward chromatin in the way of membrane sheets or tubules (reviewed in [105]). Regardless of the case, defects in any of these mitotic functions could affect the quality of cell division and lead to aneuploidy, a common feature of tumors [106]. In the metazoa, NPC formation occurs in two different phases of the cell cycle and through different assembly mechanisms: Post-mitotic NPC assembly and interphase NPC formation [18,107,108,109,110]. Two models for post-mitotic NPC assembly have been proposed: The insertion model claims that NPCs are reassembled into an intact nuclear envelope, while the enclosure model proposes that NPC assembly starts before the NE encloses the chromatin (reviewed in [111,112,113]). In any case, post-mitotic NPC assembly happens in a step-wise manner and it is subjected to fine surveillance mechanisms (reviewed in [65]). NPC assembly begins in early anaphase with soluble NPC proteins (Nup107-160 scaffold) positioning on the chromatin, mediated by Elys/Mel28, before membrane reformation. Then, it might be followed by the recruitment of transmembrane nucleoporins (reviewed in [24,108,109]). In the case of NUP153, it might even participate in the biogenesis of the lamina [114].
Surveillance mechanisms ensure correct post-mitotic reformation of NPCs (reviewed in [65]) and the assembly of the basket-like feature is particularly necessary to complete cytokinesis in a timely manner [114]. Aurora kinase B links the basket-like feature with cytokinesis, and this is currently explored as a chemotherapeutic approach in clinical trials against cancer [115].
The number of NPCs formed during interphase doubles prior re-entry into mitosis (reviewed in [111]). However, little is known about NPC formation during interphase, although it is differentially regulated compared to post-mitotic NPC assembly. Interphase NPC formation is dependent upon cyclin-dependent kinase (CDK) activity, but not upon ELYS/Mel-28 [116,117].
Kinetochores and microtubules are also essential in NE reassembly, particularly, in the recruitment of BAF to the chromatin template [118]. Acetylated Lem4 (ANKLE2) participates in this process by promoting the dephosphorylation of BAF [96,97,119]. The binding of BAF to chromatin is indispensable for most of the integral-membrane LEM-domain containing proteins to connect to chromatin through their interaction with BAF itself [118,120,121]. A recent elegant report using human cells showed that the role of BAF in nuclear assembly depends upon its ability to link distant DNA sites [122]. Microtubule organizer γ-tubulin may play a noncanonical and distinct role in promoting NE assembly [123]. A local suppression of microtubules during nuclear formation, fulfilled by chromatin-bound microtubule regulators, is required in X. laevis for proper pronuclear assembly and regular morphology of the nucleus [124]. An association of γ-tubulin with the nucleoporin ELYS/Mel-28 and the NE reassembling GTPase, Ran, has been described in X. laevis [125]. In both X. laevis and mammalian cells, a γ-tubulin boundary made of γ-strings is formed around chromatin during NE assembly, and this γ-tubulin boundary ensures the formation of the lamina around chromatin by recruiting of lamin B (Figure 1) [123]. The formation of fibrillar aggregates of γ-tubulin was further confirmed upon chaperonin containing TCP-1 (CCT) action in vitro [126]. Shaping the nucleus and achieving a regular distribution of NPCs has been shown to depend upon the γ-tubulin complex protein 3-interacting proteins in Arabidopsis thaliana [127]. Furthermore, in human cells, expression of a γ-tubulin mutant that lacks the DNA-binding domain forms chromatin-empty nuclear-like structures, demonstrating that a persistent interplay between the chromatin-associated and the cytosolic pools of γ-tubulin is required for proper NE assembly [123].
The finding that several NETs (NET5/Samp1/Tmem201, WFS1, Tmem214, and otefin) partially concentrate on or around the mitotic spindle, and in the case of the latter, the centrosome during mitosis together with the tissue specificity of many of the NETs affecting the cell cycle, suggests that further implications of these proteins in mitosis might come in the future (reviewed in [14]).
2.2.2. Spatial Distribution of NE Elements
Chromatin discs involve different areas: the “inner core” (the central region of the disc that faces the midzone), the “outer core” (the central region that faces away), and the “non-core” region (the peripheral edge of the disc) [16,118,132,133].
In metazoa, INM proteins and membrane destined for the NE make initial contacts at the non-core region with the γ-tubulin boundary before spreading around and engulfing chromatin [123,134]. Telomeric regions of sister chromatids are bound with unique proteins essential to nuclear architecture, as is LAP2α [98,99]. Furthermore, in newly forming nuclei, telomeres localize to the periphery of the nucleus, suggesting that these regions are involved in the initial seed of nuclear assembly [135].
The core region (inner and outer) is the target for ESCRT (endosomal sorting complexes required for transport) pathway proteins to recruit the microtubule severing factor spastin and seal annular gaps in the newly formed NE [136,137,138,139]. This core is initially deficient in NPC formation, but the process begins at this site soon after membrane closure [133].
The relevance of NPCs in anchoring interactions necessary for nuclear shape maintenance and structural integrity is illustrated, first, by interactions between nucleoporins NUP53, NUP88 and NUP153 and lamins [140,141,142], second, by the finding of polymorphic, lobular nuclear shapes after the depletions of these nucleoporins [140,143,144], and third, by SUN domain-containing protein 1 (SUN1) preferential location in the vicinity of NPCs [145].
All in all, defects in NE proteins might cause an inability to disassemble the NE at mitosis onset (generating partially maintained connections between NE fragments and chromatin) and to reassemble NE at the end of mitosis, blocking proper chromosome segregation and resulting in micronuclei and aneuploidy [106]. The wrapping of all chromosomes into a sole nucleus is thus essential for preserving the integrity of the genome and preventing the development of tumors.
3. The Nuclear Envelope in Cell Cycle Regulation and Signaling
3.1. Nuclear Envelope in Cell Cycle Regulation
Several elements of the NE (lamin A, lamin B, LAP2α, γ-tubulin, and emerin) have been shown to interfere with the function of the main effectors of cell cycle regulation (retinoblastoma protein–RB, E2Fs, c-Myc), as reviewed below.
In the mammalian cell cycle, normal cells exert a tight regulation of the G1-to-S phase transition, whereas in cancer cells, this transition is a main objective for dysregulation. RB is one of the earlier identified tumor suppressors [146]. Hence, RB activity is deregulated in a broad spectrum of tumors [147]. RB has abundant binding partners [148], the most important of which is the transcriptional factor E2F, which controls a range of genes important for entry into the S phase of the cell cycle. Hypophosphorylated RB binds to E2F complexes and represses the expression of S-phase genes, retaining cells in G1. CDK-dependent phosphorylation promotes the release of RB from E2F and cell cycle progression [149].
In mammals, lamin A regulates G1-to-S phase transition by affecting the RB pathway [150,151,152,153,154], since A-type lamins are required for proper RB function. In detail, A-type lamins promote RB-dependent transcriptional repression of E2F target genes. Furthermore, A-type lamins influence three other machineries regulating RB function: RB phosphorylation, RB localization, and RB protein stability [155,156]. The effect of A-type lamins in RB protein stability, together with the altered activity of ubiquitin ligase components detected in cells expressing mutant forms of lamin A, raise the possibility that A-type lamins work as coordinators of nuclear proteasome function [157].
The RB pathway is further implicated in telomere regulation and cell senescence and cell differentiation in multiple lineage, DNA replication, mitosis, and DNA-damage-activated checkpoint pathways (among others) [147], further linking A-type lamins to all of these processes. Supporting the implication of lamins in the regulation of DNA replication, intranuclear A-type lamins have been shown to associate with initial sites of DNA synthesis upon S-phase entry [158]. In immortalized cells, lamin B was localized to intranuclear sites of late S-phase replication [159], and disruption of the lamin structure impairs initiation of DNA synthesis [160,161,162].
More than A-type lamins, nuclear γ-tubulin also regulate the transcriptional activity of E2F [163]. Nuclear γ-tubulin and E2F concur in a DNA-binding complex isolated from E2F-regulated promoters [163]. In addition, RB1 and γ-tubulin proteins mutually control their expression, and, in several tumors, an inverse correlation in their expression levels was reported for γ-tubulin and RB1 [164]. Interestingly, γ-tubulin also interacts with lamin B recruitment at post-mitotic NE reassembly, as previously mentioned [123].
Other A-type lamin functions may promote G1 maintenance, since RB–lamin A/C and extracellular signal-regulated kinase (ERK)1/2–lamin A/C complexes are mutually exclusive. When G1 arrested cells are stimulated with serum, c-Fos protein is phosphorylated by mitogen activated protein kinase (MAPK) ERK1/2. Phosphorylated c-Fos associates with c-Jun- to form a dimeric Activating Protein 1 (AP-1) transcription activator complex that mediates cell cycle progression [165]. ERK1/2-dependent lamin A/C binding upon serum stimulation releases RB from the RB–lamin A/C complex, thereby promoting cell cycle progression.
The NPC-associated sentrin-specific protease 1 (SENP1) is also reported to influence cell cycle progression by regulating the expression of CDK inhibitors [166,167].
Regarding c-Myc-encoded proteins, their association with the nuclear matrix was first described in avian nuclei [168], and more recently, it was shown that the stabilized and active form of the MYC protein (pS62MYC) is enriched at the nuclear periphery of mammalian cell lines in proximity with lamin A/C [169], and precisely localizes to the nuclear pore basket [170]. However, how this regulates transcription and cellular functions remains to be elucidated.
Concerning cell senescence, the NE and the RB pathway have been implicated in an oncogenic signaling that triggers a cell cycle arrest program, i.e., oncogene-induced senescence (OIS). A dramatic reorganization of heterochromatin occurs in OIS. OIS cells lose heterochromatin interactions with lamin B1 through lamina-associated domains (LADs) [171,172], therefore, heterochromatin leaves the nuclear periphery and appears as internal senescence-associated heterochromatin foci (SAHFs) [173]. The activation of the pRB pathway is implicated in the appearance of SAHFs [173], while the NE is also implicated via laminB1 and LBR [174,175] and nuclear pore density [176]. In addition, the composition and density of the NPC changes during differentiation and tumorigenesis [19,46,176,177,178].
With respect to the maintenance of telomere metabolism [179] and DNA damage, in human cells, mutant LMNA has been connected to p53 engagement due to enhanced DNA damage (reviewed in [180]). Indeed, retinoblastoma independent regulation of cell proliferation and senescence by the p53-p21 axis was reported in lamin A/C-depleted cells [181].
In apoptosis, both via the intrinsic and extrinsic pathways, lamins have been described as cleaved by caspases 3 and 6. Indeed, the cleavage of lamin proteins by caspases is a necessary step in apoptosis that allows for nuclear membrane degradation to proceed, followed by chromatin condensation in a murine model [182]. In human and avian cells, A-type lamins are cleaved at their conserved VEID site by caspase 6, while B-type are cleaved at their VEVD site by caspase 3 [183,184,185]. In contrast, an active role of lamins in the induction, but also the prevention of apoptosis is beginning to emerge (reviewed in [186]). In cancer, apoptosis is usually reported. Strikingly, apoptosis levels are increased in the most highly proliferative tumors compared to lowly proliferative tumors. The role of lamins, if any, behind these altered levels is still unclear. One possibility would be that the amount of lamins present and the accessibility of lamins for caspases could delay the onset of apoptosis in certain tumors [187].
In metazoan, an estimated 10% of total A-type lamins localize throughout the nucleoplasm in a mobile and dynamic pool, most likely in association with LAP2α [98,162,188]. Studies on the role of A-type lamins and the RB pathway do not discriminate between these two lamin pools. However, the LAP2α promoter was reported to bind E2F1 and c-Myc [65], E2F1 and E2F4 [66], E2F3b [67], and E2F7 [68], as assessed by chromatin immunoprecipitation and microarray techniques. Indeed, LAP2α expression aligns with the phase of the cell cycle, and its overexpression has been described in various human tumor samples and cancer-derived cell lines (reviewed in [189]).
A last example is the INM protein emerin, which has been linked to cell cycle misregulation in microarray studies in X-linked EDMD patient samples where the lack of emerin disrupts the RB pathway [190].
In summary, several elements of the NE interact with the regulators of cell cycle progression, cell senescence, telomer metabolism and apoptosis. Therefore, perturbations in NE elements affecting the strict control of these interactions can lead to the development of cancer.
3.2. Nuclear Envelope in Cell Signaling
Extracellular or cytoplasmic stimuli reach the nuclear interior through signal transduction with the aim of inducing a cellular response, resolved mainly through variations in gene expression. The following signaling cascades from the plasma membrane count with an additional layer of regulation at the NE: MAPK signaling, AKT-Mammalian Target of Rapamycin signaling, Notch signaling, Wnt signaling, NF-κB signaling, and transforming growth factor-β (TGFβ) signaling. This control comes from the fact that signaling cascades need to get into the nucleus through the NPCs and that several effectors (β-catenin and smads) are sequestered at the NE by multiple NETs, as described below.
3.2.1. Lamins in Cell Signaling
The MAPK pathway dysregulation has been shown to be a driving factor in oncogenesis [191,192,193]. This pathway involves three main arms: ERK1/2, c-Jun NH2-terminal kinase (JNK), and p38 [194,195]. Phosphorylated MAPKs transit to specific subcellular compartments, such as the nucleus, to elicit their function. The localization of phosphorylated MAPKs to the nucleus is predominantly mediated by binding interactions with sequestering anchors and components of the nuclear transport machinery. In the nucleus, phosphorylated MAPKs regulate various cellular processes from growth to apoptosis, passing through differentiation, inflammation, metabolism, stress response, and autophagy [194,195,196]. An ERK1/2-activated transcription factor promoting cell cycle progression is, as previously introduced, c-Fos. c-Fos activity is suppressed by a sequestering interaction with lamin A that localizes this transcription factor to the NE [197]. Furthermore, ERK1/2 colocalizes with c-Fos at the NE by means of binding lamin A and this leads to the phosphorylation and release of c-Fos from the NE in mammalian models [198]. In addition, enhanced nuclear accumulation of ERK1/2 and JNK was reported in mice carrying a missense mutation that causes autosomal dominant EDMD in humans [199].
The AKT- mammalian target of rapamycin (mTOR) signaling pathway is frequently co-activated along with ERK1/2 in response to growth factor signaling and in various forms of cancer [200,201]. Alterations in A-type lamins have been shown to trigger AKT-mTOR signaling in the above-mentioned mice model of autosomal dominant EDMD and in mice expressing a truncated form of lamin A (lamin AΔ8-11). In mammals, lamin A itself can be phosphorylated by AKT, by which its expression can be regulated [202,203].
Hutchinson–Gilford progeria syndrome (HGPS) is mainly linked to a silent mutation LMNA. This G608G mutation in LMNA triggers a cryptic splice site and as a result, the progerin protein is produced. Progerin is a truncated form of prelamin A where the last C-terminal 50 amino acids are missing [204,205]. Several signaling pathways are imbalanced in HPGS due to the presence of progerin:
- One of them is Notch signaling, which is altered in many cancers and is thought to maintain cancer stem cells [206]. This highly conserved juxtacrine signaling is involved in regulating cell fate specification and it is altered in the mesenchymal stem cell lineage in HPGS.
- Additionally, the deposition of extracellular matrix (ECM) is altered in children with HGPS, mainly due to the reduced activity of the TCF4/LEF1 complex, a key downstream effector of the Wnt signaling pathway. Progerin expression decreases the expression and nuclear accumulation of LEF1 [207].
3.2.2. LEM Proteins in Cell Signaling
Several LEM proteins have been shown to recruit and regulate the transcriptional co-activators of the Wnt and the TGFβ signaling pathways: β-catenin and Smads, respectively:
- Emerin is a binding partner of β-catenin. Upon activation of Wnt signaling, β-catenin escapes proteasomal degradation and accumulates in the nucleus. Emerin binding to β-catenin inhibits its activity by facilitating nuclear export, thereby preventing accumulation in the nucleus in human fibroblasts [210].
- MAN1 binds to receptor-mediated Smads (rSmads), intracellular mediators of the TGF-β, and bone morphogenic protein (BMP) signaling. rSmads play an intimate role in cancer metastasis [211]. The C-terminus of MAN1 sequesters rSmads at the inner nuclear membrane, thereby preventing their ability to migrate to gene enhancer regions and activate transcription [212,213,214].
In brief, several elements of the NE provide an additional stage of regulation of signaling pathways that control proliferation and, in turn, potential points whose dysregulation may generate the unrestrained proliferation typical of tumor transformation.
4. Nuclear Envelope Regulation of the Genome
Tumor cells are characterized by massive changes in both the pattern of gene expression and in genome organization, the most critical of these being chromosome translocations and DNA damage/breaks. Since the link between chromosome translocations and tumorigenesis has been established in humans [215], checking karyotypes for translocations has become a standard diagnostic tool for many cancer types. Physically, chromosome translocations need DNA damage and repair systems to occur, but the course of chromosome translocations varies upon aspects of higher order chromosome structure in the nucleus. Particularly, this refers to chromosome positioning patterns. These patterns are preserved in a tissue-specific manner and might explain the preference for certain chromosomal translocations in certain tumor types. Indeed, in a particular tissue where the tumor arises, chromosomes adjacent one to another in the interphase nucleus will be involved in the chromosomal translocations distinctive of this tumor type [216].
4.1. Spatial Genome Organization Directed from the Nuclear Periphery
A direct physical contact between the NE and the chromatin was first suggested by microscopy observations [217,218]. Later biochemical experiments further supported that contact. In detail, chromatin components were retained and detected when extracting and purifying the NE using high ionic strength buffers [219].
More recently, new evidence in support of this view has come from specific chromatin epigenetic modifications. Heterochromatin is modified with histone H3 lysine 9 dimethylation and trimethylation (H3K9me2 and H3K9me3). In higher eukaryotes, antibodies against H3K9me2 reveal the distribution of heterochromatin close to the NE. Antibodies against H3K9me3, in contrast, stain internal heterochromatic domains in accordance with the presence of H3K9me3 in telomeres and centromeres [220]. These data agree with earlier biochemical and microscopic observations describing distinct subpopulations of HP1α at the NE in both mouse and human cells [221].
In addition, several chromatin-binding partners of NETs precisely colocalize with transcriptionally silenced chromatin [90,222,223]. Indeed, these partners are indispensable for heterochromatin distribution at the nuclear periphery in a specific pattern [50,224]. Mostly, the NET SUN proteins have been reported to mediate tethering to the NE of telomere and centromere in several systems [225,226,227,228,229].
4.2. Tethering of Chromosomes and Loci to the NE
NETs tissue-specific expression was previously described above. The specific collection of NETs in a cell type seems to be crucial in determining the position of chromosomes to the NE. Hence, when NETs preferentially expressed in muscle cells, fat, or liver are exogenously expressed in fibroblasts, chromosomes reposition to the NE. In liver cells, knock-down of liver-specific NETs 45 and 47 triggered the release of several chromosomes from the NE [230]. Strikingly, different NETs might affect the location of different subsets of chromosomes, raising the tempting possibility that NETs are the actual endogenous players behind a tissue-specific spatial genome organization [230].
4.3. Chromatin Lipid Fraction
The presence of phospholipids as a component of chromatin is well-documented and many enzymes, such as sphingomyelin-synthase (SMS) and sphingomyelinase (SMase), have been located in the INM [231]. Many different roles have been attributed to the intranuclear lipid fraction in relation to cell proliferation and differentiation.
Lipid microdomains, rich in Sphingomyelin (SM) and cholesterol (CHO), present in the INM are called nuclear lipid microdomains (NLMs). In the mammal liver, NLMs act as a resting place for active chromatin and transcription factors by regulating DNA [232] and RNA [8,233] synthesis.
SM is highly represented inside the nucleus and changes in its amount in different cell physiological states. These variations might reflect the nuclear presence of a SMase, which hydrolyzes SM to ceramide and phosphorylcholine (PPC), and a SMS, which synthesizes SM from ceramide and PPC derived from PC (reviewed by [231]). The prognostic value of SMS levels in certain types of human cancer is just beginning to be understood [234,235]. In the S-phase, the decondensation of chromatin is favored by a decrease of SM due to the increased activity of SMase. The decrease of SM favors the increase of the CHO free fraction. In turn, CHO enhances the activities of cyclin-dependent kinases needed for the entrance into the S-phase. In the contrast, inhibition of CHO synthesis induces cell cycle arrest in the S-phase with decreased expression of cdk2 and cdk4. At the end of the S-phase, an increase in SM is observed when SMS increases its activity whereas that of SMase decreases. Indeed, the restoration of the amount of SM marks the transition moment to the G2 phase (reviewed by [231]).
To recapitulate, NE actively participates in the distribution and organization of the genome within the nucleus, by means of NETs and specific membrane lipids. In the early stages of cancer, alterations in this distribution can result in chromosomal translocations and altered gene expression.
5. The Nuclear Envelope Link to Cell Migration and Metastasis and Its Use in Cancer Prognosis
The differences in cell morphology in human tumor cells were first reported in the mid-1800s [236,237]. Diagnostic features include differences in cell size and shape, number, and size of the nuclei, and loss of adherence to adjacent cells in biopsies [236,238]. In the mid-1900s, Papanicolau’s smear test was established as a routine technique for cervical cancer detection and is used today in a wide range of specimens, as outlined above [2,3,239]. Interestingly, in current computer-assisted diagnostic protocols, the diagnostic parameters employed are still largely morphological and nucleus centric, as they were 160 years ago. Focusing on the nucleus, the trained eye of the cytopathologist observes those features: Karyoplasmic ratio, nuclear roundness, NE smoothness, chromatin distribution, and the presence of NE invaginations and grooves (reviewed in [240]).
5.1. Nuclear Mechanics
Remarkably, while different cancers arise through different mechanisms and from different tissues, the above-mentioned nuclear abnormalities are mostly still common in all cancers, suggesting that nuclear structural alterations are functionally relevant in carcinogenesis. Mechanistically, the nucleoskeleton–cytoskeleton connection has profound effects on cell polarization, nuclear positioning, nuclear migration, and cell migration [241,242].
In addition to the implication of oncogenes and tumor-suppressor genes, the role of the physical constraints of tumor cells and their microenvironment has been explored within the last 15 years. Cancer cells show reduced stiffness, generate increased contractile forces, and are strongly influenced by their biomechanical environment (reviewed in [243]). Physical measurement can be used to distinguish nontumorigenic cells from cancer cells, and highly invasive from less invasive cells among those later [244].
The nucleus is both elastic and viscoelastic. Both behaviors are due to the nuclear lamina and the nuclear interior, respectively. Typically, the nucleus is ~2–10 times stiffer when compared to the surrounding cytoplasm. Hence, both the nuclear lamina and the nuclear interior govern the mechanical deformability of the nucleus. In reference to the lamins, cells modulate their levels according to the environmental constraints. Numerous studies on mice and cells grown on different supports show a prevalence of type A lamins in situations requiring stiffness, compared to a predominance of type B lamins when elasticity is required [245,246,247,248]. Moreover, lamins participate in the transmission of mechanical forces from the cytoskeleton, as they are integrated in a membrane–protein–chromatin network that allows their physical connection with Linkers of the Nucleoskeleton to the Cytoskeleton (LINC complexes) [249]. Additionally, chromatin contributes to nuclear stiffness and viscoelastic behavior (reviewed in [250]).
The structure and composition of the NE is especially relevant in cellular mechanics and function, affecting nuclear deformability and fragility and participating in mechanotransduction signaling (reviewed in [250]). Regarding the nuclear interior, it contains more than DNA and histones. The composition of this nucleoskeleton, its function, and its relevance in cancer remain a matter of debate, while a plethora of structural proteins are present on this compartment (actin, myosin, spectrin, γ-tubulin) (reviewed in [129,250]). Nucleo-cytoskeletal coupling has been reported to occur through LINC complexes, SUN domain proteins, nesprins and other KASH domain proteins, and the INM protein Samp1 [251,252]. These nucleo-cytoskeletal coupling elements are critical to ensure the force transmission between the nucleus and cytoskeleton and the modulation of the protrusions needed for cell migration (reviewed in [250]). Moreover, direct connections of the cytoskeleton with the nucleoskeleton through the NE might be an alternative means to the NPC for signal transduction between the cytoplasm and nucleus (reviewed in [240]). Using these elements, the stimulation of integrins on the surface of intact endothelial cells, for instance, results in both the reorientation of cytoskeletal filaments, the distortion of the nucleus, and the spatial redistribution of subnuclear structures [253]. In addition, changes in nuclear organization might affect gene expression of DNA stability (reviewed in [250]). Still, one open question is whether this mechanically induced change in nuclear structure and chromatin configuration can activate specifically mechanosensitive genes [254].
The relevance of nuclear mechanics in cancer comes from the fact that the nucleus is the largest and stiffest organelle of the cell, dominating the overall cellular mechanical response when cells are subjected to large deformations, for instance, when squeezing through narrow constrictions imposed by ECM fibers and other cells [241,242,255]. The irregular nuclear morphology of cancer cells often resembles that of abnormal nuclear shapes in cells with altered NE proteins, such as A- and B-type lamins and LBR (reviewed in [250]). Presumably, changes in nuclear architecture modify the rigidity of the nucleus, and this might increase nuclear deformability to benefit metastatic processes where cells need to pass through narrow interstitial spaces or small capillaries. Transient nuclear deformations, resulting in hourglass- and cigar-shape nuclei as well as nuclear protrusions have been observed during cancer cell migration in vivo [241].
5.2. Nuclear Positioning, Nuclear Envelope Rupture and Repair, and Cancer Cell Migration
Nuclear positioning in the cytoplasm is a highly regulated process that is dynamic in space and time and is required for multiple cellular and developmental processes. Extreme examples are skeletal muscle cells with a nuclei position at the periphery and epithelial tissues where nuclei are usually away from the apical membrane (reviewed in [256]). Nuclear positioning might additionally be involved in metastatic tumor cell migration, since cell polarization is required as a prior step for proper cell migration. In the course of cell polarization, in multiple migrating cell types, a centrosome and Golgi reorientation phenomenon occurs by which these organelles become positioned between the nucleus and the future leading edge. This reorientation seems to be achieved by a rearward movement of the nucleus away from the future leading edge, whereas the centrosome stays mostly static in the center of the cell [257]. Inhibition of nuclear movement impairs cell migration (reviewed in [256]).
Several molecular entities of the nucleo-cytoskeletal coupling have been found to be required for nuclear movement prior to cell migration: Nesprins, SUN, Samp1, and transmembrane actin- associated nuclear (TAN) lines, a novel NE structures involved in force transduction during nuclear movement (reviewed in [256]). The reshaping of the NE and the rearrangement of the NE-associated cytoskeleton help to prepare the cells for directional motion [258,259]. Furthermore, recent reports in mammals have connected the movement and positioning of organelles in the cytoplasm to γ-tubulin protein, e.g., in the positioning of Golgi apparatus and mitochondria [260,261]. In line with this, the Leu387Pro mutation in γ-tubulin was shown to influence nuclear positioning in yeast cells [262]. This may be attributed to the association of nuclear and cytosolic γ-strings with other components of the γ-tubulin meshwork, such as γ-tubules [263], which may provide a supporting scaffold dictating the positioning of the nuclear compartment.
A long list of translocations involving nucleoporins have been described in tumors (reviewed in [65,75]). Several of them alter the normal function of the NPC and contribute to pathogenicity. NUP214–ABL fusion protein needs to be targeted to the NPC for its transforming activity [264], and NUP98 fusion proteins affect nuclear export [265]. Furthermore, nucleoporins interact with important players in cell migration [143,266,267,268,269,270,271]. For example, NUP153 contributes to cell motility and migration interacting with A-type lamins and SUN1, while NUP358 interacts with kinesin 2 to locate APC to the cell cortex [267].
Concerning NE rupture and repair, in normal cells, NE breakdown and reassembly is limited to mitosis and is precisely regulated [105]. Missegregated chromosomes during mitotic exit can recruit their own NE to form micronuclei (MN), resulting in a NE susceptible to loss of integrity [272] and might be an objective biomarker for genomic instability in solid tumors. The irreversible NE rupture of MN can cause extensive DNA damage and promote tumorigenesis. MNs contain less NPC than the nucleus, are defective in nucleocytoplasmic transport, and replicate their DNA in an ineffective and asynchronous manner to their primary nucleus [272,273]. Furthermore, in many cancer cells, transient rupture and resealing of the NE is a common event during interphase. As a result, the nucleus and cytoplasm temporarily exchange material between them, even occasionally entrapping cytoplasmic organelles inside the nucleus. NE rupture has been further linked to the appearance of micronuclei, to the mislocalization of nucleoplasmic and cytoplasmic proteins, and to the exit of chromatin portions from the nuclear interior [274].
5.3. Nuclear Envelope Invaginations and Cancer
The NE is mainly a smooth surface, but it also presents invaginations. Those invaginations can stretch deep into the nucleoplasm, eventually crossing the nucleus entirely. High-resolution microscopy observations have revealed that aberrations in nuclear shape and size used for cancer diagnosis are probably due to large-scale invaginations of the NE that are able to traverse most or even all the nucleus (reviewed in [275]). These invaginations have been named the nucleoplasmic reticulum (NR) for their morphological similarity to the ER. NR invaginations are called type I when the INM alone is involved, while NR invaginations are called type II when both the INM and ONM invaginate into the nucleoplasm. Type II invaginations may enclose microtubules, microfilaments, and even mitochondria in their cytoplasmic core [276,277]. Interestingly, a potential implication of γ-tubulin tubules in nuclear invagination may be further investigated, since a recent report showed that γ-tubules could also pass through the center of the nucleus [261]. The NR is observed in nuclei from various normal and abnormal tissues [278], as well as in cells grown in 2D and 3D cultures, including many tumor cell types, such as brain, breast, kidney, bladder, prostate, and ovary [279,280]. Appearance of the NR may occur both after mitosis, during NE reassembly, and without mitosis, i.e., de novo in interphase cell nuclei. Then, NR is commonly maintained throughout interphase and, in specific cell types, it can show heritable patterns [281,282,283]. The actual mechanisms by which this new nuclear structural component, the NR, affects cell function in normal and cancer cells are still under investigation. However, the diagnostic and prognostic significance of irregularities and invaginations of the NE in cancer cells are indisputable.
6. Conclusions
The specific traits of nuclear cell morphology in tumor cells have been used in the last 150 years to evaluate the diagnosis and prognosis of human cancer. Although the NM, attached NETs, and other proteins have been found to be implicated in this morphology (e.g., type A/C and type B lamins, nucleoporins, γ-tubulin), further research is needed to precisely define the molecular entities behind the alterations in the number, size, and shape of nuclei in tumor cells.
Author Contributions
M.A.-K. wrote and revised the paper, provided guidance and funding; C.A.R. conceived and designed the manuscript, collected the relevant references, wrote and revised the paper.
Funding
This research was funded by the Swedish Cancer Society (grant number CAN2016/369), Skane University Hospital in Malmö Cancer Research Fund (grant number 20151209), the Swedish Childhood Cancer Foundation (grant number PR2016-0084), the Crafoordska Foundation (grant number 20180670), the European Commission Seventh Framework Programme (agreement number PIEF-GA-2013-627441) and the Balearic Island Government and European Social Fund (grant number PD/036/2016).
Acknowledgments
We apologize to all authors whose work could not be cited due to space constraints. The authors thank C Hoffmann for editorial assistance.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| APC | Adenomatous polyposis coli (APC) |
| BAF | Barrier-to- autointegration-factor |
| BMP | Bone morphogenic protein |
| CCT | Chaperonin containing TCP-1 |
| CDK | Cyclin-dependent kinase |
| CHO | Cholesterol |
| ECM | Extracellular matrix |
| EDMD | Emery–Dreifuss muscular dystrophy |
| ER | Endoplasmic reticulum |
| ERK1/2 | Extracellular signal-regulated kinase 1/2 |
| ESCRT | Endosomal sorting complexes required for transport |
| FG | Phenylalanine and glycine |
| HGPS | Hutchinson–Gilford progeria syndrome |
| HP1 | Heterochromatin protein 1 |
| INM | Inner nuclear membrane |
| JNK | c-Jun NH2-terminal kinase |
| LADs | Lamina-associated domains |
| LBR | Lamin B receptor |
| MAPK | Mitogen Activated Protein Kinase |
| MN | Micronuclei |
| NE | Nuclear envelope |
| NETs | Nuclear envelope transmembrane proteins |
| NLMs | Nuclear lipid microdomains |
| NPCs | Nuclear pore complexes |
| NR | Nucleoplasmic reticulum |
| OIS | Oncogene-induced senescence |
| ONM | Outer nuclear membrane |
| PPC | Phosphorylcholine |
| RB | Retinoblastoma |
| SAHFs | Senescence-associated heterochromatin foci |
| SENP1 | Sentrin-specific protease 1 |
| SM | Sphingomyelin |
| SMS | Sphingomyelin-synthase |
| TAN lines | Transmembrane actin- associated nuclear lines |
| TGFβ | Transforming growth factor-β |
References
- Papanicolaou, G.N. A new procedure for staining vaginal smears. Science 1942, 95, 438–439. [Google Scholar] [CrossRef] [PubMed]
- Morrison, L.F.; Hopp, E.S.; Wu, R. II Diagnosis of Malignancy of the Nasopharynx. Cytological Studies by the Smear Technic. Ann. Otol. Rhinol. Laryngol. 1949, 58, 18–32. [Google Scholar] [CrossRef]
- Montgomery, P.W.; Von Haam, E. A Study of the Exfoliative Cytology in Patients with Carcinoma of the Oral Mucosa. J. Dent. Res. 1951, 30, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Callan, H.G.; Tomlin, S.G. Experimental studies on amphibian oocyte nuclei I. Investigation of the structure of the nuclear membrane by means of the electron microscope. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1950, 137, 367–378. [Google Scholar]
- Tapley, E.C.; Starr, D.A. Connecting the nucleus to the cytoskeleton by SUN–KASH bridges across the nuclear envelope. Curr. Opin. Cell Biol. 2013, 25, 57–62. [Google Scholar] [CrossRef]
- Suntharalingam, M.; Wente, S.R. Peering through the Pore: Nuclear Pore Complex Structure, Assembly, and Function. Dev. Cell 2003, 4, 775–789. [Google Scholar] [CrossRef]
- Schirmer, E.C.; Gerace, L. The nuclear membrane proteome: extending the envelope. Trends Biochem. Sci. 2005, 30, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Cascianelli, G.; Villani, M.; Tosti, M.; Marini, F.; Bartoccini, E.; Viola Magni, M.; Albi, E. Lipid Microdomains in Cell Nucleus. Mol. Biol. Cell 2008, 19, 5289–5295. [Google Scholar] [CrossRef] [Green Version]
- Ledeen, R.W.; Wu, G. Nuclear lipids: key signaling effectors in the nervous system and other tissues. J. Lipid Res. 2004, 45, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.C.; Worman, H.J. Nuclear Envelope Regulation of Signaling Cascades. Adv. Exp. Med. Biol. 2014, 773, 187–206. [Google Scholar]
- Aaronson, R.P.; Blobel, G. Isolation of nuclear pore complexes in association with a lamina. Proc. Natl. Acad. Sci. USA 1975, 72, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Maeno, H.; Sugimoto, K.; Nakajima, N. Genomic Structure of the Mouse Gene (Lmnb1) Encoding Nuclear Lamin B1. Genomics 1995, 30, 342–346. [Google Scholar] [CrossRef]
- Gerace, L.; Burke, B. Functional Organization of the Nuclear Envelope. Annu. Rev. Cell Biol. 1988, 4, 335–374. [Google Scholar] [CrossRef] [PubMed]
- Robson, M.I.; Le Thanh, P.; Schirmer, E.C. NETs and cell cycle regulation. Adv. Exp. Med. Biol. 2014, 773, 165–185. [Google Scholar]
- Ellenberg, J.; Lippincott-Schwartz, J. Dynamics and Mobility of Nuclear Envelope Proteins in Interphase and Mitotic Cells Revealed by Green Fluorescent Protein Chimeras. Methods 1999, 19, 362–372. [Google Scholar] [CrossRef]
- Haraguchi, T.; Koujin, T.; Hayakawa, T.; Kaneda, T.; Tsutsumi, C.; Imamoto, N.; Akazawa, C.; Sukegawa, J.; Yoneda, Y.; Hiraoka, Y. Live fluorescence imaging reveals early recruitment of emerin, LBR, RanBP2, and Nup153 to reforming functional nuclear envelopes. J. Cell Sci. 2000, 113, 779–794. [Google Scholar]
- Cronshaw, J.M.; Krutchinsky, A.N.; Zhang, W.; Chait, B.T.; Matunis, M.J. Proteomic analysis of the mammalian nuclear pore complex. J. Cell Biol. 2002, 158, 915–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strambio-De-Castillia, C.; Niepel, M.; Rout, M.P. The nuclear pore complex: bridging nuclear transport and gene regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 490–501. [Google Scholar] [CrossRef]
- Raices, M.; D’Angelo, M.A. Nuclear pore complex composition: a new regulator of tissue-specific and developmental functions. Nat. Rev. Mol. Cell Biol. 2012, 13, 687–699. [Google Scholar] [CrossRef]
- Terry, L.J.; Wente, S.R. Flexible Gates: Dynamic Topologies and Functions for FG Nucleoporins in Nucleocytoplasmic Transport. Eukaryot. Cell 2009, 8, 1814–1827. [Google Scholar] [CrossRef] [Green Version]
- Ungricht, R.; Kutay, U. Mechanisms and functions of nuclear envelope remodelling. Nat. Rev. Mol. Cell Biol. 2017, 18, 229–245. [Google Scholar] [CrossRef]
- D’Angelo, M.A.; Anderson, D.J.; Richard, E.; Hetzer, M.W. Nuclear pores form de novo from both sides of the nuclear envelope. Science 2006, 312, 440–443. [Google Scholar] [CrossRef]
- Newport, J.W.; Wilson, K.L.; Dunphy, W.G. A lamin-independent pathway for nuclear envelope assembly. J. Cell Biol. 1990, 111, 2247–2259. [Google Scholar] [CrossRef]
- Webster, M.; Witkin, K.L.; Cohen-Fix, O. Sizing up the nucleus: nuclear shape, size and nuclear-envelope assembly. J. Cell Sci. 2009, 122, 1477–1486. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Papagiannouli, F.; Wagner, N.; Wilsch-Bräuninger, M.; Braun, M.; Furlong, E.E.; Loserth, S.; Wenzl, C.; Pilot, F.; Vogt, N.; et al. Developmental control of nuclear size and shape by Kugelkern and Kurzkern. Curr. Biol. 2006, 16, 543–552. [Google Scholar] [CrossRef]
- Dittmer, T.A.; Stacey, N.J.; Sugimoto-Shirasu, K.; Richards, E.J. LITTLE NUCLEI genes affecting nuclear morphology in Arabidopsis thaliana. Plant Cell 2007, 19, 2793–2803. [Google Scholar] [CrossRef]
- Doucet, C.M.; Hetzer, M.W. Nuclear pore biogenesis into an intact nuclear envelope. Chromosoma 2010, 119, 469–477. [Google Scholar] [CrossRef]
- Toyama, B.H.; Savas, J.N.; Park, S.K.; Harris, M.S.; Ingolia, N.T.; Yates, J.R.; Hetzer, M.W. Identification of long-lived proteins reveals exceptional stability of essential cellular structures. Cell 2013, 154, 971–982. [Google Scholar] [CrossRef]
- Schneider, J.; Moragues, D.; Martínez, N.; Romero, H.; Jimenez, E.; Pontón, J. Cross-reactivity between Candida albicans and human ovarian carcinoma as revealed by monoclonal antibodies PA10F and C6. Br. J. Cancer 1998, 77, 1015–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, N.; Alonso, A.; Moragues, M.D.; Pontón, J.; Schneider, J. The nuclear pore complex protein Nup88 is overexpressed in tumor cells. Cancer Res. 1999, 59, 5408–5411. [Google Scholar]
- Gould, V.E.; Orucevic, A.; Zentgraf, H.; Gattuso, P.; Martinez, N.; Alonso, A. Nup88 (karyoporin) in human malignant neoplasms and dysplasias: correlations of immunostaining of tissue sections, cytologic smears, and immunoblot analysis. Hum. Pathol. 2002, 33, 536–544. [Google Scholar] [CrossRef]
- Agudo, D.; Gómez-Esquer, F.; Martínez-Arribas, F.; Núñez-Villar, M.J.; Pollán, M.; Schneider, J. Nup88 mRNA overexpression is associated with high aggressiveness of breast cancer. Int. J. Cancer 2004, 109, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Knoess, M.; Kurz, A.K.; Goreva, O.; Bektas, N.; Breuhahn, K.; Odenthal, M.; Schirmacher, P.; Dienes, H.P.; Bock, C.T.; Zentgraf, H.; et al. Nucleoporin 88 expression in hepatitis B and C virus-related liver diseases. World J. Gastroenterol. 2006, 12, 5870–5874. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Y.; Zhao, Z.-R.; Jiang, L.; Li, J.-C.; Gao, Y.-M.; Cui, D.-S.; Wang, C.-J.; Schneider, J.; Wang, M.-W.; Sun, X.-F. Nup88 expression in normal mucosa, adenoma, primary adenocarcinoma and lymph node metastasis in the colorectum. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2007, 28, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Brustmann, H.; Hager, M. Nucleoporin 88 expression in normal and neoplastic squamous epithelia of the uterine cervix. Ann. Diagn. Pathol. 2009, 13, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.; Martínez-Arribas, F.; Torrejón, R. Nup88 expression is associated with myometrial invasion in endometrial carcinoma. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 2010, 20, 804–808. [Google Scholar] [CrossRef]
- Simon, D.N.; Rout, M.P. Cancer and the Nuclear Pore Complex. Adv. Exp. Med. Biol. 2014, 773, 285–307. [Google Scholar]
- Johannessen, J.V.; Sobrinho-Simöes, M.; Finseth, I.; Pilström, L. Papillary carcinomas of the thyroid have pore-deficient nuclei. Int. J. Cancer 1982, 30, 409–411. [Google Scholar] [CrossRef]
- Czerniak, B.; Koss, L.G.; Sherman, A. Nuclear pores and DNA ploidy in human bladder carcinomas. Cancer Res. 1984, 44, 3752–3756. [Google Scholar]
- Sugie, S.; Yoshimi, N.; Tanaka, T.; Mori, H.; Williams, G.M. Alterations of nuclear pores in preneoplastic and neoplastic rat liver lesions induced by 2-acetylaminofluorene. Carcinogenesis 1994, 15, 95–98. [Google Scholar] [CrossRef]
- Lewin, J.M.; Lwaleed, B.A.; Cooper, A.J.; Birch, B.R. The direct effect of nuclear pores on nuclear chemotherapeutic concentration in multidrug resistant bladder cancer: the nuclear sparing phenomenon. J. Urol. 2007, 177, 1526–1530. [Google Scholar] [CrossRef]
- Korfali, N.; Wilkie, G.S.; Swanson, S.K.; Srsen, V.; de las Heras, J.; Batrakou, D.G.; Malik, P.; Zuleger, N.; Kerr, A.R.W.; Florens, L.; et al. The nuclear envelope proteome differs notably between tissues. Nucleus 2012, 3, 552–564. [Google Scholar] [CrossRef] [Green Version]
- Mattout-Drubezki, A.; Gruenbaum, Y. Dynamic interactions of nuclear lamina proteins with chromatin and transcriptional machinery. Cell. Mol. Life Sci. 2003, 60, 2053–2063. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.J.; Soshnev, A.A.; Geyer, P.K. Networking in the nucleus: a spotlight on LEM-domain proteins. Curr. Opin. Cell Biol. 2015, 34, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Heessen, S.; Fornerod, M. The inner nuclear envelope as a transcription factor resting place. Embo Rep. 2007, 8, 914–919. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Bravo, V.; Pippa, R.; Song, W.-M.; Carceles-Cordon, M.; Dominguez-Andres, A.; Fujiwara, N.; Woo, J.; Koh, A.P.; Ertel, A.; Lokareddy, R.K.; et al. Nuclear Pores Promote Lethal Prostate Cancer by Increasing POM121-Driven E2F1, MYC, and AR Nuclear Import. Cell 2018, 174, 1200–1215. [Google Scholar] [CrossRef]
- Dauer, W.T.; Worman, H.J. The nuclear envelope as a signaling node in development and disease. Dev. Cell 2009, 17, 626–638. [Google Scholar] [CrossRef]
- Kind, J.; van Steensel, B. Genome–nuclear lamina interactions and gene regulation. Curr. Opin. Cell Biol. 2010, 22, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Mattout, A.; Pike, B.L.; Towbin, B.D.; Bank, E.M.; Gonzalez-Sandoval, A.; Stadler, M.B.; Meister, P.; Gruenbaum, Y.; Gasser, S.M. An EDMD mutation in C. elegans lamin blocks muscle-specific gene relocation and compromises muscle integrity. Curr. Biol. 2011, 21, 1603–1614. [Google Scholar] [CrossRef]
- Solovei, I.; Wang, A.S.; Thanisch, K.; Schmidt, C.S.; Krebs, S.; Zwerger, M.; Cohen, T.V.; Devys, D.; Foisner, R.; Peichl, L.; et al. LBR and lamin A/C sequentially tether peripheral heterochromatin and inversely regulate differentiation. Cell 2013, 152, 584–598. [Google Scholar] [CrossRef]
- Burke, B.; Stewart, C.L. The nuclear lamins: flexibility in function. Nat. Rev. Mol. Cell Biol. 2013, 14, 13–24. [Google Scholar] [CrossRef]
- Gerace, L.; Blobel, G. The Nuclear Envelope Lamina is Reversibly Depolymerized during Mitosis: Cell. Available online: https://www.cell.com/cell/pdf/0092-8674(80)90409-2.pdf?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2F0092867480904092%3Fshowall%3Dtrue (accessed on 10 March 2019).
- Heald, R.; McKeon, F. Mutations of phosphorylation sites in lamin A that prevent nuclear lamina disassembly in mitosis. Cell 1990, 61, 579–589. [Google Scholar] [CrossRef]
- Peter, M.; Nakagawa, J.; Dorée, M.; Labbé, J.C.; Nigg, E.A. In vitro disassembly of the nuclear lamina and M phase-specific phosphorylation of lamins by cdc2 kinase. Cell 1990, 61, 591–602. [Google Scholar] [CrossRef]
- Ward, G.E.; Kirschner, M.W. Identification of cell cycle-regulated phosphorylation sites on nuclear lamin C. Cell 1990, 61, 561–577. [Google Scholar] [CrossRef]
- Pyrpasopoulou, A.; Meier, J.; Maison, C.; Simos, G.; Georgatos, S.D. The lamin B receptor (LBR) provides essential chromatin docking sites at the nuclear envelope. Embo J. 1996, 15, 7108–7119. [Google Scholar] [CrossRef]
- Foisner, R.; Gerace, L. Integral membrane proteins of the nuclear envelope interact with lamins and chromosomes, and binding is modulated by mitotic phosphorylation. Cell 1993, 73, 1267–1279. [Google Scholar] [CrossRef]
- Courvalin, J.C.; Segil, N.; Blobel, G.; Worman, H.J. The lamin B receptor of the inner nuclear membrane undergoes mitosis-specific phosphorylation and is a substrate for p34cdc2-type protein kinase. J. Biol. Chem. 1992, 267, 19035–19038. [Google Scholar] [PubMed]
- Tseng, L.-C.; Chen, R.-H. Temporal control of nuclear envelope assembly by phosphorylation of lamin B receptor. Mol. Biol. Cell 2011, 22, 3306–3317. [Google Scholar] [CrossRef]
- Nichols, R.J.; Wiebe, M.S.; Traktman, P. The Vaccinia-related Kinases Phosphorylate the N′ Terminus of BAF, Regulating Its Interaction with DNA and Its Retention in the Nucleus. Mol. Biol. Cell 2006, 17, 2451–2464. [Google Scholar] [CrossRef] [PubMed]
- Gorjánácz, M.; Klerkx, E.P.F.; Galy, V.; Santarella, R.; López-Iglesias, C.; Askjaer, P.; Mattaj, I.W. Caenorhabditis elegans BAF-1 and its kinase VRK-1 participate directly in post-mitotic nuclear envelope assembly. Embo J. 2007, 26, 132–143. [Google Scholar] [CrossRef] [PubMed]
- LaJoie, D.; Ullman, K.S. Coordinated events of nuclear assembly. Curr. Opin. Cell Biol. 2017, 46, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.A.; van Heesbeen, R.G.H.P.; Meaders, J.L.; Geers, E.F.; Fernandez-Garcia, B.; Medema, R.H.; Tanenbaum, M.E. Nuclear envelope-associated dynein drives prophase centrosome separation and enables Eg5-independent bipolar spindle formation. Embo J. 2012, 31, 4179–4190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Simone, A.; Nédélec, F.; Gönczy, P. Dynein Transmits Polarized Actomyosin Cortical Flows to Promote Centrosome Separation. Cell Rep. 2016, 14, 2250–2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, K.-H.; Factor, R.E.; Ullman, K.S. The nuclear envelope environment and its cancer connections. Nat. Rev. Cancer 2012, 12, 196–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabachinski, G.; Schwartz, T.U. The nuclear pore complex--structure and function at a glance. J. Cell Sci. 2015, 128, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Chatel, G.; Fahrenkrog, B. Nucleoporins: Leaving the nuclear pore complex for a successful mitosis. Cell. Signal. 2011, 23, 1555–1562. [Google Scholar] [CrossRef]
- Bolhy, S.; Bouhlel, I.; Dultz, E.; Nayak, T.; Zuccolo, M.; Gatti, X.; Vallee, R.; Ellenberg, J.; Doye, V. A Nup133-dependent NPC-anchored network tethers centrosomes to the nuclear envelope in prophase. J. Cell Biol. 2011, 192, 855–871. [Google Scholar] [CrossRef] [Green Version]
- Splinter, D.; Tanenbaum, M.E.; Lindqvist, A.; Jaarsma, D.; Flotho, A.; Yu, K.L.; Grigoriev, I.; Engelsma, D.; Haasdijk, E.D.; Keijzer, N.; et al. Bicaudal D2, Dynein, and Kinesin-1 Associate with Nuclear Pore Complexes and Regulate Centrosome and Nuclear Positioning during Mitotic Entry. PLoS Biol. 2010, 8, e1000350. [Google Scholar] [CrossRef]
- Burke, B.; Ellenberg, J. Remodelling the walls of the nucleus. Nat. Rev. Mol. Cell Biol. 2002, 3, 487. [Google Scholar] [CrossRef]
- Silkworth, W.T.; Nardi, I.K.; Paul, R.; Mogilner, A.; Cimini, D. Timing of centrosome separation is important for accurate chromosome segregation. Mol. Biol. Cell 2012, 23, 401–411. [Google Scholar] [CrossRef]
- Dawlaty, M.M.; Malureanu, L.; Jeganathan, K.B.; Kao, E.; Sustmann, C.; Tahk, S.; Shuai, K.; Grosschedl, R.; van Deursen, J.M. Resolution of Sister Centromeres Requires RanBP2-Mediated SUMOylation of Topoisomerase IIα. Cell 2008, 133, 103–115. [Google Scholar] [CrossRef]
- Jeganathan, K.B.; Malureanu, L.; van Deursen, J.M. The Rae1-Nup98 complex prevents aneuploidy by inhibiting securin degradation. Nature 2005, 438, 1036–1039. [Google Scholar] [CrossRef]
- Jeganathan, K.B.; Baker, D.J.; van Deursen, J.M. Securin associates with APCCdh1 in prometaphase but its destruction is delayed by Rae1 and Nup98 until the metaphase/anaphase transition. Cell Cycle Georget. Tex 2006, 5, 366–370. [Google Scholar] [CrossRef]
- Xu, S.; Powers, M.A. Nuclear pore proteins and cancer. Semin. Cell Dev. Biol. 2009, 20, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Sterling, H.; Burlingame, A.; McCormick, F. Tpr directly binds to Mad1 and Mad2 and is important for the Mad1–Mad2-mediated mitotic spindle checkpoint. Genes Dev. 2008, 22, 2926–2931. [Google Scholar] [CrossRef]
- Cross, M.K.; Powers, M.A. Nup98 regulates bipolar spindle assembly through association with microtubules and opposition of MCAK. Mol. Biol. Cell 2011, 22, 661–672. [Google Scholar] [CrossRef]
- Schellhaus, A.K.; De Magistris, P.; Antonin, W. Nuclear Reformation at the End of Mitosis. J. Mol. Biol. 2016, 428, 1962–1985. [Google Scholar] [CrossRef]
- Wilkie, G.S.; Korfali, N.; Swanson, S.K.; Malik, P.; Srsen, V.; Batrakou, D.G.; de las Heras, J.; Zuleger, N.; Kerr, A.R.W.; Florens, L.; et al. Several Novel Nuclear Envelope Transmembrane Proteins Identified in Skeletal Muscle Have Cytoskeletal Associations. Mol. Cell. Proteom. 2011, 10, M110.003129. [Google Scholar] [CrossRef]
- Audhya, A.; Desai, A.; Oegema, K. A role for Rab5 in structuring the endoplasmic reticulum. J. Cell Biol. 2007, 178, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Buch, C.; Lindberg, R.; Figueroa, R.; Gudise, S.; Onischenko, E.; Hallberg, E. An integral protein of the inner nuclear membrane localizes to the mitotic spindle in mammalian cells. J. Cell Sci. 2009, 122, 2100–2107. [Google Scholar] [CrossRef] [Green Version]
- Drummond, S.; Ferrigno, P.; Lyon, C.; Murphy, J.; Goldberg, M.; Allen, T.; Smythe, C.; Hutchison, C.J. Temporal differences in the appearance of NEP-B78 and an LBR-like protein during Xenopus nuclear envelope reassembly reflect the ordered recruitment of functionally discrete vesicle types. J. Cell Biol. 1999, 144, 225–240. [Google Scholar] [CrossRef]
- Hetzer, M.; Meyer, H.H.; Walther, T.C.; Bilbao-Cortes, D.; Warren, G.; Mattaj, I.W. Distinct AAA-ATPase p97 complexes function in discrete steps of nuclear assembly. Nat. Cell Biol. 2001, 3, 1086–1091. [Google Scholar] [CrossRef]
- Salpingidou, G.; Rzepecki, R.; Kiseleva, E.; Lyon, C.; Lane, B.; Fusiek, K.; Golebiewska, A.; Drummond, S.; Allen, T.D.; Ellis, J.A.; et al. NEP-A and NEP-B both contribute to nuclear pore formation in Xenopus eggs and oocytes. J. Cell Sci. 2008, 121, 706–716. [Google Scholar] [CrossRef] [Green Version]
- Vigers, G.P.A.; Lohka, M.J. A distinct vesicle population targets membranes and pore complexes to the nuclear envelope in Xenopus eggs. J. Cell Biol. 1991, 112, 545–556. [Google Scholar] [CrossRef]
- Vigers, G.P.; Lohka, M.J. Regulation of nuclear envelope precursor functions during cell division. J Cell Sci 1992, 102, 273–284. [Google Scholar]
- Vagnarelli, P.; Earnshaw, W.C. Repo-Man-PP1: A link between chromatin remodelling and nuclear envelope reassembly. Nucleus 2012, 3, 138–142. [Google Scholar] [CrossRef]
- Ohba, T.; Nakamura, M.; Nishitani, H.; Nishimoto, T. Self-organization of microtubule asters induced in Xenopus egg extracts by GTP-bound ran. Science 1999, 284, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Wiese, C.; Wilde, A.; Moore, M.S.; Adam, S.A.; Merdes, A.; Zheng, Y. Role of importin-β in coupling ran to downstream targets in microtubule assembly. Science 2001, 291, 653–656. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Worman, H.J. Interaction between an integral protein of the nuclear envelope inner membrane and human chromodomain proteins homologous to Drosophila HP1. J. Biol. Chem. 1996, 271, 14653–14656. [Google Scholar] [CrossRef]
- Ye, Q.; Callebaut, I.; Pezhman, A.; Courvalin, J.-C.; Worman, H.J. Domain-specific Interactions of Human HP1-type Chromodomain Proteins and Inner Nuclear Membrane Protein LBR. J. Biol. Chem. 1997, 272, 14983–14989. [Google Scholar] [CrossRef] [Green Version]
- Polioudaki, H.; Kourmouli, N.; Drosou, V.; Bakou, A.; Theodoropoulos, P.A.; Singh, P.B.; Giannakouros, T.; Georgatos, S.D. Histones H3/H4 form a tight complex with the inner nuclear membrane protein LBR and heterochromatin protein 1. Embo Rep. 2001, 2, 920–925. [Google Scholar] [CrossRef] [Green Version]
- Ulbert, S.; Platani, M.; Boue, S.; Mattaj, I.W. Direct membrane protein–DNA interactions required early in nuclear envelope assembly. J. Cell Biol. 2006, 173, 469–476. [Google Scholar] [CrossRef]
- Anderson, D.J.; Hetzer, M.W. Nuclear envelope formation by chromatin-mediated reorganization of the endoplasmic reticulum. Nat. Cell Biol. 2007, 9, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Molitor, T.P.; Traktman, P. Depletion of the protein kinase VRK1 disrupts nuclear envelope morphology and leads to BAF retention on mitotic chromosomes. Mol. Biol. Cell 2014, 25, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Asencio, C.; Davidson, I.F.; Santarella-Mellwig, R.; Ly-Hartig, T.B.N.; Mall, M.; Wallenfang, M.R.; Mattaj, I.W.; Gorjánácz, M. Coordination of Kinase and Phosphatase Activities by Lem4 Enables Nuclear Envelope Reassembly during Mitosis. Cell 2012, 150, 122–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, P.-Y.; Zhang, J.-B.; Zhu, X.-D.; Zhang, W.; Wu, W.-Z.; Tan, Y.-S.; Hou, J.; Tang, Z.-Y.; Qin, L.-X.; Sun, H.-C. Two pathologic types of hepatocellular carcinoma with lymph node metastasis with distinct prognosis on the basis of CK19 expression in tumor. Cancer 2008, 112, 2740–2748. [Google Scholar] [CrossRef] [PubMed]
- Dechat, T. LAP2 and BAF transiently localize to telomeres and specific regions on chromatin during nuclear assembly. J. Cell Sci. 2004, 117, 6117–6128. [Google Scholar] [CrossRef] [PubMed]
- Chojnowski, A.; Ong, P.F.; Wong, E.S.M.; Lim, J.S.Y.; Mutalif, R.A.; Navasankari, R.; Dutta, B.; Yang, H.; Liow, Y.Y.; Sze, S.K.; et al. Progerin reduces LAP2α-telomere association in hutchinson-gilford progeria. eLife 2015, 4, 1–21. [Google Scholar] [CrossRef]
- Zierhut, C.; Jenness, C.; Kimura, H.; Funabiki, H. Nucleosomal regulation of chromatin composition and nuclear assembly revealed by histone depletion. Nat. Struct. Mol. Biol. 2014, 21, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Zhang, Y. Nucleosome assembly is required for nuclear pore complex assembly in mouse zygotes. Nat. Struct. Mol. Biol. 2014, 21, 609–616. [Google Scholar] [CrossRef]
- Schooley, A.; Moreno-Andre, D.; De Magistris, P.; Vollmer, B.; Antonin, W. The lysine demethylase LSD1 is required for nuclear envelope formation at the end of mitosis. J. Cell Sci. 2015, 128, 3466–3477. [Google Scholar] [CrossRef]
- Vagnarelli, P.; Ribeiro, S.; Sennels, L.; Sanchez-Pulido, L.; de Lima Alves, F.; Verheyen, T.; Kelly, D.A.; Ponting, C.P.; Rappsilber, J.; Earnshaw, W.C. Repo-Man Coordinates Chromosomal Reorganization with Nuclear Envelope Reassembly during Mitotic Exit. Dev. Cell 2011, 21, 328–342. [Google Scholar] [CrossRef] [Green Version]
- Worman, H.J.; Ostlund, C.; Wang, Y. Diseases of the nuclear envelope. Cold Spring Harb. Perspect. Biol. 2010, 2, a000706. [Google Scholar] [CrossRef]
- Wandke, C.; Kutay, U. Enclosing Chromatin: Reassembly of the Nucleus after Open Mitosis. Cell 2013, 152, 1222–1225. [Google Scholar] [CrossRef] [Green Version]
- Qi, W.; Yu, H. The Spindle Checkpoint and Chromosomal Stability. Genome Dyn. 2006, 1, 116–130. [Google Scholar]
- Tran, E.J.; Wente, S.R. Dynamic Nuclear Pore Complexes: Life on the Edge. Cell 2006, 125, 1041–1053. [Google Scholar] [CrossRef] [Green Version]
- Antonin, W.; Ellenberg, J.; Dultz, E. Nuclear pore complex assembly through the cell cycle: regulation and membrane organization. Febs Lett. 2008, 582, 2004–2016. [Google Scholar] [CrossRef]
- D’Angelo, M.A.; Hetzer, M.W. Structure, dynamics and function of nuclear pore complexes. Trends Cell Biol. 2008, 18, 456–466. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.-H.; Baines, A.T.; Fiordalisi, J.J.; Shipitsin, M.; Feig, L.A.; Cox, A.D.; Der, C.J.; Counter, C.M. Activation of RalA is critical for Ras-induced tumorigenesis of human cells. Cancer Cell 2005, 7, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Maeshima, K.; Iino, H.; Hihara, S.; Imamoto, N. Nuclear size, nuclear pore number and cell cycle. Nucleus 2011, 2, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Schooley, A.; Vollmer, B.; Antonin, W. Building a nuclear envelope at the end of mitosis: coordinating membrane reorganization, nuclear pore complex assembly, and chromatin de-condensation. Chromosoma 2012, 121, 539–554. [Google Scholar] [CrossRef] [Green Version]
- Weberruss, M.; Antonin, W. Perforating the nuclear boundary – how nuclear pore complexes assemble. J. Cell Sci. 2016, 129, 4439–4447. [Google Scholar] [CrossRef]
- Mackay, D.R.; Makise, M.; Ullman, K.S. Defects in nuclear pore assembly lead to activation of an Aurora B-mediated abscission checkpoint. J. Cell Biol. 2010, 191, 923–931. [Google Scholar] [CrossRef]
- Dar, A.A.; Goff, L.W.; Majid, S.; Berlin, J.; El-Rifai, W. Aurora kinase inhibitors--rising stars in cancer therapeutics? Mol. Cancer Ther. 2010, 9, 268–278. [Google Scholar] [CrossRef]
- Doucet, C.M.; Talamas, J.A.; Hetzer, M.W. Cell cycle-dependent differences in nuclear pore complex assembly in metazoa. Cell 2010, 141, 1030–1041. [Google Scholar] [CrossRef]
- Maeshima, K.; Iino, H.; Hihara, S.; Funakoshi, T.; Watanabe, A.; Nishimura, M.; Nakatomi, R.; Yahata, K.; Imamoto, F.; Hashikawa, T.; et al. Nuclear pore formation but not nuclear growth is governed by cyclin-dependent kinases (Cdks) during interphase. Nat. Struct. Mol. Biol. 2010, 17, 1065–1071. [Google Scholar] [CrossRef]
- Haraguchi, T.; Kojidani, T.; Koujin, T.; Shimi, T.; Osakada, H.; Mori, C.; Yamamoto, A.; Hiraoka, Y. Live cell imaging and electron microscopy reveal dynamic processes of BAF-directed nuclear envelope assembly. J. Cell Sci. 2008, 121, 2540–2554. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, T.; Kukolj, E.; Brachner, A.; Beltzung, E.; Bruno, M.; Kostrhon, S.; Opravil, S.; Hudecz, O.; Mechtler, K.; Warren, G.; et al. SIRT2 regulates nuclear envelope reassembly through ANKLE2 deacetylation. J Cell Sci 2016, 129, 4607–4621. [Google Scholar] [CrossRef]
- Haraguchi, T.; Koujin, T.; Segura-Totten, M.; Lee, K.K.; Matsuoka, Y.; Yoneda, Y.; Wilson, K.L.; Hiraoka, Y. BAF is required for emerin assembly into the reforming nuclear envelope. J. Cell Sci. 2001, 114, 4575–4585. [Google Scholar]
- Margalit, A.; Segura-Totten, M.; Gruenbaum, Y.; Wilson, K.L. Barrier-to-autointegration factor is required to segregate and enclose chromosomes within the nuclear envelope and assemble the nuclear lamina. Proc. Natl. Acad. Sci. USA 2005, 102, 3290–3295. [Google Scholar] [CrossRef] [Green Version]
- Samwer, M.; Schneider, M.W.G.; Hoefler, R.; Schmalhorst, P.S.; Jude, J.G.; Zuber, J.; Gerlich, D.W. DNA Cross-Bridging Shapes a Single Nucleus from a Set of Mitotic Chromosomes. Cell 2017, 170, 956–972. [Google Scholar] [CrossRef]
- Rosselló, C.A.; Lindström, L.; Glindre, J.; Eklund, G.; Alvarado-Kristensson, M. Gamma-tubulin coordinates nuclear envelope assembly around chromatin. Heliyon 2016, 2, e00166. [Google Scholar] [CrossRef]
- Xue, J.Z.; Woo, E.M.; Postow, L.; Chait, B.T.; Funabiki, H. Chromatin-Bound Xenopus Dppa2 Shapes the Nucleus by Locally Inhibiting Microtubule Assembly. Dev. Cell 2013, 27, 47–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, H.; Koch, B.; Walczak, R.; Ciray-Duygu, F.; González-Sánchez, J.C.; Devos, D.P.; Mattaj, I.W.; Gruss, O.J. The nucleoporin MEL-28 promotes RanGTP-dependent γ-tubulin recruitment and microtubule nucleation in mitotic spindle formation. Nat. Commun. 2014, 5, 3270. [Google Scholar] [CrossRef]
- Pouchucq, L.; Lobos-Ruiz, P.; Araya, G.; Valpuesta, J.M.; Monasterio, O. The chaperonin CCT promotes the formation of fibrillar aggregates of γ-tubulin. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Batzenschlager, M.; Masoud, K.; Janski, N.; Houlné, G.; Herzog, E.; Evrard, J.-L.; Baumberger, N.; Erhardt, M.; Nominé, Y.; Kieffer, B.; et al. The GIP gamma-tubulin complex-associated proteins are involved in nuclear architecture in Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chumová, J.; Kourová, H.; Trögelová, L.; Halada, P.; Binarová, P. Microtubular and Nuclear Functions of γ-Tubulin: Are They LINCed? Cells 2019, 8, 259. [Google Scholar] [CrossRef]
- Rosselló, C.A.; Lindström, L.; Eklund, G.; Corvaisier, M.; Kristensson, M.A. γ-Tubulin–γ-Tubulin Interactions as the Basis for the Formation of a Meshwork. Int. J. Mol. Sci. 2018, 19, 3245. [Google Scholar] [CrossRef]
- Gimpel, P.; Lee, Y.L.; Sobota, R.M.; Calvi, A.; Koullourou, V.; Patel, R.; Mamchaoui, K.; Nédélec, F.; Shackleton, S.; Schmoranzer, J.; et al. Nesprin-1α-Dependent Microtubule Nucleation from the Nuclear Envelope via Akap450 Is Necessary for Nuclear Positioning in Muscle Cells. Curr. Biol. 2017, 27, 2999–3009. [Google Scholar] [CrossRef]
- Larsson, V.J.; Jafferali, M.H.; Vijayaraghavan, B.; Figueroa, R.A.; Hallberg, E. Mitotic spindle assembly and γ-tubulin localisation depend on the integral nuclear membrane protein Samp1. J Cell Sci 2018, 131, jcs211664. [Google Scholar] [CrossRef]
- Clever, M.; Funakoshi, T.; Mimura, Y.; Takagi, M.; Imamoto, N. The nucleoporin ELYS/Mel28 regulates nuclear envelope subdomain formation in HeLa cells. Nucleus 2012, 3, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, S.; Bui, K.H.; Schorb, M.; Julius Hossain, M.; Politi, A.Z.; Koch, B.; Eltsov, M.; Beck, M.; Ellenberg, J. Nuclear pore assembly proceeds by an inside-out extrusion of the nuclear envelope. eLife 2016, 5, e19071. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Ladinsky, M.S.; Kirchhausen, T. Formation of the postmitotic nuclear envelope from extended ER cisternae precedes nuclear pore assembly. J. Cell Biol. 2011, 194, 425–440. [Google Scholar] [CrossRef] [Green Version]
- Crabbe, L.; Cesare, A.J.; Kasuboski, J.M.; Fitzpatrick, J.A.J.; Karlseder, J. Human Telomeres Are Tethered to the Nuclear Envelope during Postmitotic Nuclear Assembly. Cell Rep. 2012, 2, 1521–1529. [Google Scholar] [CrossRef] [Green Version]
- Vietri, M.; Schink, K.O.; Campsteijn, C.; Wegner, C.S.; Schultz, S.W.; Christ, L.; Thoresen, S.B.; Brech, A.; Raiborg, C.; Stenmark, H. Spastin and ESCRT-III coordinate mitotic spindle disassembly and nuclear envelope sealing. Nature 2015, 522, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Olmos, Y.; Perdrix-Rosell, A.; Carlton, J.G. Membrane Binding by CHMP7 Coordinates ESCRT-III-Dependent Nuclear Envelope Reformation. Curr. Biol. 2016, 26, 2635–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olmos, Y.; Hodgson, L.; Mantell, J.; Verkade, P.; Carlton, J.G. ESCRT-III controls nuclear envelope reformation. Nature 2015, 522, 236–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M.; Bjorkman, P.J.; Nikolova, L.; Chen, O.S.; LaJoie, D.; Redd, M.J.; Sundquist, W.I.; Frost, A.; von Appen, A.; Ladinsky, M.S.; et al. LEM2 recruits CHMP7 for ESCRT-mediated nuclear envelope closure in fission yeast and human cells. Proc. Natl. Acad. Sci. USA 2017, 114, E2166–E2175. [Google Scholar] [CrossRef] [Green Version]
- Hawryluk-Gara, L.A.; Shibuya, E.K.; Wozniak, R.W. Vertebrate Nup53 interacts with the nuclear lamina and is required for the assembly of a Nup93-containing complex. Mol. Biol. Cell 2005, 16, 2382–2394. [Google Scholar] [CrossRef] [PubMed]
- Lussi, Y.C.; Hügi, I.; Laurell, E.; Kutay, U.; Fahrenkrog, B. The nucleoporin Nup88 is interacting with nuclear lamin A. Mol. Biol. Cell 2011, 22, 1080–1090. [Google Scholar] [CrossRef]
- Smythe, C.; Jenkins, H.E.; Hutchison, C.J. Incorporation of the nuclear pore basket protein nup153 into nuclear pore structures is dependent upon lamina assembly: evidence from cell-free extracts of Xenopus eggs. Embo J. 2000, 19, 3918–3931. [Google Scholar] [CrossRef]
- Zhou, L.; Panté, N. The nucleoporin Nup153 maintains nuclear envelope architecture and is required for cell migration in tumor cells. Febs Lett. 2010, 584, 3013–3020. [Google Scholar] [CrossRef] [Green Version]
- Mackay, D.R.; Elgort, S.W.; Ullman, K.S. The nucleoporin Nup153 has separable roles in both early mitotic progression and the resolution of mitosis. Mol. Biol. Cell 2009, 20, 1652–1660. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Pante, N.; Misteli, T.; Elsagga, M.; Crisp, M.; Hodzic, D.; Burke, B.; Roux, K.J. Functional association of Sun1 with nuclear pore complexes. J. Cell Biol. 2007, 178, 785–798. [Google Scholar] [CrossRef] [Green Version]
- Friend, S.H.; Bernards, R.; Rogelj, S.; Weinberg, R.A.; Rapaport, J.M.; Albert, D.M.; Dryja, T.P. A human DNA segment with properties of the gene that predisposes to retinoblastoma and osteosarcoma. Nature 1986, 323, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Dick, F.A.; Rubin, S.M. Molecular mechanisms underlying RB protein function. Nat. Rev. Mol. Cell Biol. 2013, 14, 297. [Google Scholar] [CrossRef]
- Morris, E.J.; Dyson, N.J. Retinoblastoma protein partners. Adv. Cancer Res. 2001, 82, 1–54. [Google Scholar]
- Hanahan, D.; Weinberg, R. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Boban, M.; Braun, J.; Foisner, R. Lamins: ‘structure goes cycling’. Biochem. Soc. Trans. 2010, 38, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Mancini, M.A.; Shan, B.; Nickerson, J.A.; Penman, S.; Lee, W.H. The retinoblastoma gene product is a cell cycle-dependent, nuclear matrix-associated protein. Proc. Natl. Acad. Sci. USA 1994, 91, 418–422. [Google Scholar] [CrossRef]
- Ozaki, T.; Saijo, M.; Murakami, K.; Enomoto, H.; Taya, Y.; Sakiyama, S. Complex formation between lamin A and the retinoblastoma gene product: identification of the domain on lamin A required for its interaction. Oncogene 1994, 9, 2649–2653. [Google Scholar]
- Templeton, D.J.; Park, S.H.; Lanier, L.; Weinberg, R.A. Nonfunctional mutants of the retinoblastoma protein are characterized by defects in phosphorylation, viral oncoprotein association, and nuclear tethering. Proc. Natl. Acad. Sci. USA 1991, 88, 3033–3037. [Google Scholar] [CrossRef]
- Mittnacht, S.; Weinberg, R.A. G1/S phosphorylation of the retinoblastoma protein is associated with an altered affinity for the nuclear compartment. Cell 1991, 65, 381–393. [Google Scholar] [CrossRef]
- Johnson, B.R.; Nitta, R.T.; Frock, R.L.; Mounkes, L.; Barbie, D.A.; Stewart, C.L.; Harlow, E.; Kennedy, B.K. A-type lamins regulate retinoblastoma protein function by promoting subnuclear localization and preventing proteasomal degradation. Proc. Natl. Acad. Sci. USA 2004, 101, 9677–9682. [Google Scholar] [CrossRef] [Green Version]
- Van Berlo, J.H.; Voncken, J.W.; Kubben, N.; Broers, J.L.V.; Duisters, R.; van Leeuwen, R.E.W.; Crijns, H.J.G.M.; Ramaekers, F.C.S.; Hutchison, C.J.; Pinto, Y.M. A-type lamins are essential for TGF-β1 induced PP2A to dephosphorylate transcription factors. Hum. Mol. Genet. 2005, 14, 2839–2849. [Google Scholar] [CrossRef]
- Muralikrishna, B.; Chaturvedi, P.; Sinha, K.; Parnaik, V.K. Lamin misexpression upregulates three distinct ubiquitin ligase systems that degrade ATR kinase in HeLa cells. Mol. Cell. Biochem. 2012, 365, 323–332. [Google Scholar] [CrossRef]
- Kennedy, B.K.; Barbie, D.A.; Classon, M.; Dyson, N.; Harlow, E. Nuclear organization of DNA replication in primary mammalian cells. Genes Dev. 2000, 14, 2855–2868. [Google Scholar] [CrossRef] [Green Version]
- Moir, R.D.; Montag-Lowy, M.; Goldman, R.D. Dynamic properties of nuclear lamins: lamin B is associated with sites of DNA replication. J. Cell Biol. 1994, 125, 1201–1212. [Google Scholar] [CrossRef]
- Ellis, D.J.; Jenkins, H.; Whitfield, W.G.F.; Hutchison, C.J. GST-lamin fusion proteins act as dominant negative mutants in Xenopus egg extract and reveal the function of the lamina in DNA replication. J Cell Sci 1997, 110, 2507–2518. [Google Scholar]
- Spann, T.P.; Moir, R.D.; Goldman, A.E.; Stick, R.; Goldman, R.D. Disruption of nuclear lamin organization alters the distribution of replication factors and inhibits DNA synthesis. J. Cell Biol. 1997, 136, 1201–1212. [Google Scholar] [CrossRef]
- Moir, R.D.; Spann, T.P.; Herrmann, H.; Goldman, R.D. Disruption of nuclear lamin organization blocks the elongation phase of DNA replication. J. Cell Biol. 2000, 149, 1179–1192. [Google Scholar] [CrossRef]
- Hoog, G.; Zarrizi, R.; von Stedingk, K.; Jonsson, K.; Alvarado-Kristensson, M. Nuclear localization of γ-tubulin affects E2F transcriptional activity and S-phase progression. Faseb J. 2011, 25, 3815–3827. [Google Scholar] [CrossRef]
- Ehlén, A.; Rosselló, C.A.; Von Stedingk, K.; Höög, G.; Nilsson, E.; Pettersson, H.M.; Jirström, K.; Alvarado-Kristensson, M. Tumors with nonfunctional retinoblastoma protein are killed by reduced γ-tubulin levels. J. Biol. Chem. 2012, 287, 17241–17247. [Google Scholar] [CrossRef]
- Verde, P.; Casalino, L.; Talotta, F.; Yaniv, M.; Weitzman, J.B. Deciphering AP-1 function in tumorigenesis: Fra-ternizing on target promoters. Cell Cycle 2007, 6, 2633–2639. [Google Scholar] [CrossRef]
- Palancade, B.; Doye, V. Sumoylating and desumoylating enzymes at nuclear pores: underpinning their unexpected duties? Trends Cell Biol. 2008, 18, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, J.; Zuo, Y.; Deng, J.; Wang, L.-S.; Chen, G.-Q. SUMO-specific protease 1 regulates the in vitro and in vivo growth of colon cancer cells with the upregulated expression of CDK inhibitors. Cancer Lett. 2011, 309, 78–84. [Google Scholar] [CrossRef]
- Eisenman, R.N.; Tachibana, C.Y.; Abrams, H.D.; Hann, S.R. V-myc- and c-myc-encoded proteins are associated with the nuclear matrix. Mol. Cell. Biol. 1985, 5, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Myant, K.; Qiao, X.; Halonen, T.; Come, C.; Laine, A.; Janghorban, M.; Partanen, J.I.; Cassidy, J.; Ogg, E.L.; Cammareri, P.; et al. Serine 62-phosphorylated MYC Associates with nuclear lamins and its regulation by CIP2A is essential for regenerative proliferation. Cell Rep. 2015. [Google Scholar] [CrossRef]
- Su, Y.; Pelz, C.; Huang, T.; Torkenczy, K.; Wang, X.; Cherry, A.; Daniel, C.J.; Liang, J.; Nan, X.; Dai, M.-S.; et al. Post-translational modification localizes MYC to the nuclear pore basket to regulate a subset of target genes involved in cellular responses to environmental signals. Genes Dev. 2018, 32, 1398–1419. [Google Scholar] [CrossRef]
- Guelen, L.; Pagie, L.; Brasset, E.; Meuleman, W.; Faza, M.B.; Talhout, W.; Eussen, B.H.; de Klein, A.; Wessels, L.; de Laat, W.; et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008, 453, 948–951. [Google Scholar] [CrossRef] [PubMed]
- Lenain, C.; De Graaf, C.A.; Pagie, L.; Visser, N.L.; De Haas, M.; De Vries, S.S.; Peric-Hupkes, D.; Van Steensel, B.; Peeper, D.S. Massive reshaping of genome-nuclear lamina interactions during oncogene-induced senescence. Genome Res. 2017, 27, 1634–1644. [Google Scholar] [CrossRef]
- Narita, M.; Nũnez, S.; Heard, E.; Narita, M.; Lin, A.W.; Hearn, S.A.; Spector, D.L.; Hannon, G.J.; Lowe, S.W. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell 2003, 113, 703–716. [Google Scholar] [CrossRef]
- Sadaie, M.; Salama, R.; Carroll, T.; Tomimatsu, K.; Chandra, T.; Young, A.R.J.; Narita, M.; Pérez-Mancera, P.A.; Bennett, D.C.; Chong, H.; et al. Redistribution of the Lamin B1 genomic binding profile affects rearrangement of heterochromatic domains and SAHF formation during senescence. Genes Dev. 2013, 27, 1800–1808. [Google Scholar] [CrossRef] [Green Version]
- Lukasova, E.; Kovařík, A.; Bačíková, A.; Falk, M.; Kozubek, S. Loss of lamin B receptor is necessary to induce cellular senescence. Biochem. J. 2017, 474, 281–300. [Google Scholar] [CrossRef]
- Boumendil, C.; Hari, P.; Olsen, K.C.F.; Acosta, J.C.; Bickmore, W.A. Nuclear pore density controls heterochromatin reorganization during senescence. Genes Dev. 2019, 33, 144–149. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, M.A.; Gomez-Cavazos, J.S.; Mei, A.; Lackner, D.H.; Hetzer, M.W. A Change in Nuclear Pore Complex Composition Regulates Cell Differentiation. Dev. Cell 2012, 22, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Sellés, J.; Penrad-Mobayed, M.; Guillaume, C.; Fuger, A.; Auvray, L.; Faklaris, O.; Montel, F. Nuclear pore complex plasticity during developmental process as revealed by super-resolution microscopy. Sci. Rep. 2017, 7, 14732. [Google Scholar] [CrossRef]
- Gonzalez-Suarez, I.; Gonzalo, S. Nurturing the genome: A-type lamins preserve genomic stability. Nucleus 2010, 1, 129–135. [Google Scholar] [CrossRef]
- Redwood, A.B.; Gonzalez-Suarez, I.; Gonzalo, S. Regulating the levels of key factors in cell cycle and DNA repair: New pathways revealed by lamins. Cell Cycle 2011, 10, 3652–3657. [Google Scholar] [CrossRef]
- Moiseeva, O.; Bourdeau, V.; Vernier, M.; Dabauvalle, M.-C.; Ferbeyre, G. Retinoblastoma-independent regulation of cell proliferation and senescence by the p53-p21 axis in lamin A /C-depleted cells. Aging Cell 2011, 10, 789–797. [Google Scholar] [CrossRef]
- Oberhammer, F.A.; Hochegger, K.; Fröschl, G.; Tiefenbacher, R.; Pavelka, M. Chromatin condensation during apoptosis is accompanied by degradation of lamin A+B, without enhanced activation of cdc2 kinase. J. Cell Biol. 1994, 126, 827–837. [Google Scholar] [CrossRef]
- Slee, E.A.; Adrain, C.; Martin, S.J. Executioner Caspase-3, -6, and -7 Perform Distinct, Non-redundant Roles during the Demolition Phase of Apoptosis. J. Biol. Chem. 2001, 276, 7320–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruchaud, S.; Korfali, N.; Villa, P.; Kottke, T.J.; Dingwall, C.; Kaufmann, S.H.; Earnshaw, W.C. Caspase-6 gene disruption reveals a requirement for lamin A cleavage in apoptotic chromatin condensation. Embo J. 2002, 21, 1967–1977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okinaga, T.; Kasai, H.; Tsujisawa, T.; Nishihara, T. Role of caspases in cleavage of lamin A/C and PARP during apoptosis in macrophages infected with a periodontopathic bacterium. J. Med. Microbiol. 2007, 56, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Broers, J.L.V.; Ramaekers, F.C.S. The role of the nuclear lamina in cancer and apoptosis. Adv. Exp. Med. Biol. 2014, 773, 27–48. [Google Scholar]
- Rao, L.; Perez, D.; White, E. Lamin proteolysis facilitates nuclear events during apoptosis. J. Cell Biol. 1996, 135, 1441–1455. [Google Scholar] [CrossRef]
- Naetar, N.; Korbei, B.; Kozlov, S.; Kerenyi, M.A.; Dorner, D.; Kral, R.; Gotic, I.; Fuchs, P.; Cohen, T.V.; Bittner, R.; et al. Loss of nucleoplasmic LAP2α–lamin A complexes causes erythroid and epidermal progenitor hyperproliferation. Nat. Cell Biol. 2008, 10, 1341–1348. [Google Scholar] [CrossRef]
- Brachner, A.; Foisner, R. Lamina-associated polypeptide (LAP)2α and other LEM proteins in cancer biology. Adv. Exp. Med. Biol. 2014, 773, 143–163. [Google Scholar]
- Bakay, M.; Wang, Z.; Melcon, G.; Schiltz, L.; Xuan, J.; Zhao, P.; Sartorelli, V.; Seo, J.; Pegoraro, E.; Angelini, C.; et al. Nuclear envelope dystrophies show a transcriptional fingerprint suggesting disruption of Rb–MyoD pathways in muscle regeneration. Brain 2006, 129, 996–1013. [Google Scholar] [CrossRef]
- Raman, M.; Chen, W.; Cobb, M.H. Differential regulation and properties of MAPKs. Oncogene 2007, 26, 3100. [Google Scholar] [CrossRef]
- McKay, M.M.; Morrison, D.K. Integrating signals from RTKs to ERK/MAPK. Oncogene 2007, 26, 3113. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, Á.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian Mitogen-Activated Protein Kinase Signal Transduction Pathways Activated by Stress and Inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [Green Version]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK Signal Transduction Pathways Activated by Stress and Inflammation: A 10-Year Update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [Green Version]
- Plotnikov, A.; Zehorai, E.; Procaccia, S.; Seger, R. The MAPK cascades: Signaling components, nuclear roles and mechanisms of nuclear translocation. Biochim. Biophys. Acta Mol. Cell Res. 2011, 1813, 1619–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivorra, C.; Kubicek, M.; González, J.M.; Sanz-González, S.M.; Álvarez-Barrientos, A.; O’Connor, J.E.; Burke, B.; Andrés, V. A mechanism of AP-1 suppression through interaction of c-Fos with lamin A/C. Genes Dev. 2006, 20, 307–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, J.M.; Navarro-Puche, A.; Casar, B.; Crespo, P.; Andrés, V. Fast regulation of AP-1 activity through interaction of lamin A/C, ERK1/2, and c-Fos at the nuclear envelope. J. Cell Biol. 2008, 183, 653–666. [Google Scholar] [CrossRef] [Green Version]
- Arimura, T.; Helbling-Leclerc, A.; Massart, C.; Varnous, S.; Niel, F.; Lacène, E.; Fromes, Y.; Toussaint, M.; Mura, A.M.; Kelle, D.I.; et al. Mouse model carrying H222P-Lmna mutation develops muscular dystrophy and dilated cardiomyopathy similar to human striated muscle laminopathies. Hum. Mol. Genet. 2004, 14, 155–169. [Google Scholar] [CrossRef]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar]
- Cantley, L.C.; Shaw, R.J. Ras, PI(3)K and mTOR signalling controls tumour cell growth. Nature 2006, 441, 424. [Google Scholar]
- Bertacchini, J.; Beretti, F.; Cenni, V.; Guida, M.; Gibellini, F.; Mediani, L.; Marin, O.; Maraldi, N.M.; De Pol, A.; Lattanzi, G.; et al. The protein kinase Akt/PKB regulates both prelamin A degradation and Lmna gene expression. Faseb J. 2013, 27, 2145–2155. [Google Scholar] [CrossRef]
- Cenni, V.; Bertacchini, J.; Beretti, F.; Lattanzi, G.; Bavelloni, A.; Riccio, M.; Ruzzene, M.; Marin, O.; Arrigoni, G.; Parnaik, V.; et al. Lamin A Ser404 Is a nuclear target of akt phosphorylation in C2C12 cells. J. Proteome Res. 2008, 7, 4727–4735. [Google Scholar] [CrossRef]
- De Sandre-Giovannoli, A.; Bernard, R.; Cau, P.; Navarro, C.; Amiel, J.; Boccaccio, I.; Lyonnet, S.; Stewart, C.L.; Munnich, A.; Le Merrer, M.; et al. Lamin A truncation in Hutchinson-Gilford progeria. Science 2003, 300, 2055. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.; Brown, W.T.; Gordon, L.B.; Glynn, M.W.; Singer, J.; Scott, L.; Erdos, M.R.; Robbins, C.M.; Moses, T.Y.; Berglund, P.; et al. Recurrent de novo point mutations in lamin A cause Hutchinson–Gilford progeria syndrome. Nature 2003, 423, 293. [Google Scholar] [CrossRef]
- Mounkes, L.C.; Kozlov, S.; Hernandez, L.; Sullivan, T.; Stewart, C.L. A progeroid syndrome in mice is caused by defects in A-type lamins. Nature 2003, 423, 298. [Google Scholar] [CrossRef]
- Hernandez, L.; Roux, K.J.; Wong, E.S.M.; Mounkes, L.C.; Mutalif, R.; Navasankari, R.; Rai, B.; Cool, S.; Jeong, J.W.; Wang, H.; et al. Functional coupling between the extracellular matrix and nuclear lamina by wnt signaling in progeria. Dev. Cell 2010, 19, 413–425. [Google Scholar] [CrossRef]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49. [Google Scholar] [CrossRef]
- Osorio, F.G.; Bárcena, C.; Soria-Valles, C.; Ramsay, A.J.; de Carlos, F.; Cobo, J.; Fueyo, A.; Freije, J.M.P.; López-Otín, C. Nuclear lamina defects cause ATM-dependent NF-κB activation and link accelerated aging to a systemic inflammatory response. Genes Dev. 2012, 26, 2311–2324. [Google Scholar] [CrossRef] [PubMed]
- Markiewicz, E.; Tilgner, K.; Barker, N.; Van De Wetering, M.; Clevers, H.; Dorobek, M.; Hausmanowa-Petrusewicz, I.; Ramaekers, F.C.S.; Broers, J.L.V.; Blankesteijn, W.M.; et al. The inner nuclear membrane protein Emerin regulates β-catenin activity by restricting its accumulation in the nucleus. Embo J. 2006, 25, 3275–3285. [Google Scholar] [CrossRef]
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Hellemans, J.; Preobrazhenska, O.; Willaert, A.; Debeer, P.; Verdonk, P.C.M.; Costa, T.; Janssens, K.; Menten, B.; Van Roy, N.; Vermeulen, S.J.T.; et al. Loss-of-function mutations in LEMD3 result in osteopoikilosis, Buschke-Ollendorff syndrome and melorheostosis. Nat. Genet. 2004, 36, 1213. [Google Scholar] [CrossRef]
- Lin, F.; Morrison, J.M.; Wu, W.; Worman, H.J. MAN1, an integral protein of the inner nuclear membrane, binds Smad2 and Smad3 and antagonizes transforming growth factor-β signaling. Hum. Mol. Genet. 2004, 14, 437–445. [Google Scholar] [CrossRef]
- Pan, D.; Estévez-Salmerón, L.D.; Stroschein, S.L.; Zhu, X.; He, J.; Zhou, S.; Luo, K. The integral inner nuclear membrane protein MAN1 physically interacts with the R-smad proteins to repress signaling by the transforming growth factor-β superfamily of cytokines. J. Biol. Chem. 2005, 280, 15992–16001. [Google Scholar] [CrossRef]
- Rowley, J.D. Identificaton of a translocation with quinacrine fluorescence in a patient with acute leukemia. Ann. Genet. 1973, 16, 109–112. [Google Scholar]
- Parada, L.A.; McQueen, P.G.; Munson, P.J.; Misteli, T. Conservation of relative chromosome positioning in normal and cancer cells. Curr. Biol. 2002, 12, 1692–1697. [Google Scholar] [CrossRef]
- Paddy, M.R.; Belmont, A.S.; Saumweber, H.; Agard, D.A.; Sedat, J.W. Interphase nuclear envelope lamins form a discontinuous network that interacts with only a fraction of the chromatin in the nuclear periphery. Cell 1990, 62, 89–106. [Google Scholar] [CrossRef]
- Belmont, A.S.; Zhai, Y.; Thilenius, A. Lamin B distribution and association with peripheral chromatin revealed by optical sectioning and electron microscopy tomography. J. Cell Biol. 1993, 123, 1671–1685. [Google Scholar] [CrossRef]
- Bouvier, D.; Hubert, J.; Seve, A.P.; Bouteille, M. Characterization of lamina-bound chromatin in the nuclear shell isolated from HeLa cells. Exp. Cell Res. 1985, 156, 500–512. [Google Scholar] [CrossRef]
- Kind, J.; Pagie, L.; Ortabozkoyun, H.; Boyle, S.; De Vries, S.S.; Janssen, H.; Amendola, M.; Nolen, L.D.; Bickmore, W.A.; Van Steensel, B. Single-cell dynamics of genome-nuclear lamina interactions. Cell 2013, 153, 178–192. [Google Scholar] [CrossRef]
- Minc, E.; Allory, Y.; Worman, H.J.; Courvalin, J.C.; Buendia, B. Localization and phosphorylation of HP1 proteins during the cell cycle in mammalian cells. Chromosoma 1999, 108, 220–234. [Google Scholar] [CrossRef]
- Makatsori, D.; Kourmouli, N.; Polioudaki, H.; Shultz, L.D.; McLean, K.; Theodoropoulos, P.A.; Singh, P.B.; Georgatos, S.D. The inner nuclear membrane protein lamin B receptor forms distinct microdomains and links epigenetically marked chromatin to the nuclear envelope. J. Biol. Chem. 2004, 279, 25567–25573. [Google Scholar] [CrossRef]
- Brown, C.R.; Kennedy, C.J.; Delmar, V.A.; Forbes, D.J.; Silver, P.A. Global histone acetylation induces functional genomic reorganization at mammalian nuclear pore complexes. Genes Dev. 2008, 22, 627–639. [Google Scholar] [CrossRef] [Green Version]
- Zullo, J.M.; Demarco, I.A.; Piqué-Regi, R.; Gaffney, D.J.; Epstein, C.B.; Spooner, C.J.; Luperchio, T.R.; Bernstein, B.E.; Pritchard, J.K.; Reddy, K.L.; et al. DNA sequence-dependent compartmentalization and silencing of chromatin at the nuclear lamina. Cell 2012, 149, 1474–1487. [Google Scholar] [CrossRef] [PubMed]
- Chikashige, Y.; Tsutsumi, C.; Yamane, M.; Okamasa, K.; Haraguchi, T.; Hiraoka, Y. Meiotic Proteins Bqt1 and Bqt2 Tether Telomeres to Form the Bouquet Arrangement of Chromosomes. Cell 2006, 125, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Alsheimer, M.; Benavente, R.; Hodzic, D.; Schmitt, J.; Hoog, C.; Stewart, C.L. Transmembrane protein Sun2 is involved in tethering mammalian meiotic telomeres to the nuclear envelope. Proc. Natl. Acad. Sci. USA 2007, 104, 7426–7431. [Google Scholar] [Green Version]
- Bupp, J.M.; Martin, A.E.; Stensrud, E.S.; Jaspersen, S.L. Telomere anchoring at the nuclear periphery requires the budding yeast Sad1-UNC-84 domain protein Mps3. J. Cell Biol. 2007, 179, 845–854. [Google Scholar] [CrossRef]
- Conrad, M.N.; Lee, C.-Y.; Wilkerson, J.L.; Dresser, M.E. MPS3 mediates meiotic bouquet formation in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2007, 104, 8863–8868. [Google Scholar] [CrossRef]
- Hou, H.; Zhou, Z.; Wang, Y.; Wang, J.; Kallgren, S.P.; Kurchuk, T.; Miller, E.A.; Chang, F.; Jia, S. Csi1 links centromeres to the nuclear envelope for centromere clustering. J. Cell Biol. 2012, 199, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Zuleger, N.; Boyle, S.; Kelly, D.A.; de las Heras, J.I.; Lazou, V.; Korfali, N.; Batrakou, D.G.; Randles, K.N.; Morris, G.E.; Harrison, D.J.; et al. Specific nuclear envelope transmembrane proteins can promote the location of chromosomes to and from the nuclear periphery. Genome Biol. 2013, 14, R14. [Google Scholar] [CrossRef]
- Albi, E.; Viola Magni, M.P. The role of intranuclear lipids. Biol. Cell 2004, 96, 657–667. [Google Scholar] [CrossRef]
- Albi, E.; Lazzarini, A.; Lazzarini, R.; Floridi, A.; Damaskopoulou, E.; Curcio, F.; Cataldi, S. Nuclear Lipid Microdomain as Place of Interaction between Sphingomyelin and DNA during Liver Regeneration. Int. J. Mol. Sci. 2013, 14, 6529–6541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albi, E.; Villani, M. Nuclear lipid microdomains regulate cell function. Commun. Integr. Biol. 2009, 2, 23–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-García, P.; Rosselló, C.A.; Rodríguez-Lorca, R.; Beteta-Göbel, R.; Fernández-Díaz, J.; Lladó, V.; Busquets, X.; Escribá, P.V.; Fernández-García, P.; Rosselló, C.A.; et al. The Opposing Contribution of SMS1 and SMS2 to Glioma Progression and Their Value in the Therapeutic Response to 2OHOA. Cancers 2019, 11, 88. [Google Scholar] [CrossRef]
- Casares, D.; Escribá, P.V.; Rosselló, C.A. Membrane Lipid Composition: Effect on Membrane and Organelle Structure, Function and Compartmentalization and Therapeutic Avenues. Int. J. Mol. Sci. 2019, 20, 2167. [Google Scholar] [CrossRef]
- Beale, L. Examination of sputum from a case of cancer of the pharynx and the adjacent parts. Arch Med. Lond. 1860, 2, 44. [Google Scholar]
- Beale, L.S. The Microscope in Its Application to Practical Medicine; Churchill: London, UK, 1858. [Google Scholar]
- Müller, J. Über den Feineren Bau und die Formen der krankhaften Geschwülste. On the Nature and Structural Characteristics of Cancer, and of Those Morbid Growths which May be Confounded with It; G. Reimer: Berlin, Germany, 1838. [Google Scholar]
- Papanicolaou, G.N. New cancer diagnosis. In Proceedings of the Third Race Betterment Conference, Battle Creek, MI, USA, 2–6 January 1928. [Google Scholar]
- De Las Heras, J.I.; Schirmer, E.C. The nuclear envelope and cancer: A diagnostic perspective and historical overview. Adv. Exp. Med. Biol. 2014, 773, 5–26. [Google Scholar]
- Friedl, P.; Wolf, K.; Lammerding, J. Nuclear mechanics during cell migration. Curr. Opin. Cell Biol. 2011, 23, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Wolf, K.; te Lindert, M.; Krause, M.; Alexander, S.; te Riet, J.; Willis, A.L.; Hoffman, R.M.; Figdor, C.G.; Weiss, S.J.; Friedl, P. Physical limits of cell migration: Control by ECM space and nuclear deformation and tuning by proteolysis and traction force. J. Cell Biol. 2013, 201, 1069–1084. [Google Scholar] [CrossRef] [Green Version]
- Suresh, S. Biomechanics and biophysics of cancer cells. Acta Mater. 2007, 55, 3989–4014. [Google Scholar] [CrossRef] [Green Version]
- Kraning-Rush, C.M.; Califano, J.P.; Reinhart-King, C.A. Cellular traction stresses increase with increasing metastatic potential. PLoS ONE 2012, 7, e32572. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.; Ivanovska, I.L.; Buxboim, A.; Harada, T.; Dingal, P.C.D.P.; Pinter, J.; Pajerowski, J.D.; Spinler, K.R.; Shin, J.-W.; Tewari, M.; et al. Nuclear Lamin-A Scales with Tissue Stiffness and Enhances Matrix-Directed Differentiation. Science 2013, 341, 1240104. [Google Scholar] [CrossRef]
- Harada, T.; Swift, J.; Irianto, J.; Shin, J.-W.; Spinler, K.R.; Athirasala, A.; Diegmiller, R.; Dingal, P.C.D.P.; Ivanovska, I.L.; Discher, D.E. Nuclear lamin stiffness is a barrier to 3D migration, but softness can limit survival. J Cell Biol 2014, 204, 669–682. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.-W.; Spinler, K.R.; Swift, J.; Chasis, J.A.; Mohandas, N.; Discher, D.E. Lamins regulate cell trafficking and lineage maturation of adult human hematopoietic cells. Proc. Natl. Acad. Sci. USA 2013, 110, 18892–18897. [Google Scholar] [CrossRef] [Green Version]
- Rowat, A.C.; Jaalouk, D.E.; Zwerger, M.; Ung, W.L.; Eydelnant, I.A.; Olins, D.E.; Olins, A.L.; Herrmann, H.; Weitz, D.A.; Lammerding, J. Nuclear Envelope Composition Determines the Ability of Neutrophil-type Cells to Passage through Micron-scale Constrictions. J. Biol. Chem. 2013, 288, 8610–8618. [Google Scholar] [CrossRef] [Green Version]
- Lombardi, M.L.; Jaalouk, D.E.; Shanahan, C.M.; Burke, B.; Roux, K.J.; Lammerding, J. The Interaction between Nesprins and Sun Proteins at the Nuclear Envelope Is Critical for Force Transmission between the Nucleus and Cytoskeleton. J. Biol. Chem. 2011, 286, 26743–26753. [Google Scholar] [CrossRef] [Green Version]
- Denais, C.; Lammerding, J. Nuclear mechanics in cancer. Adv. Exp. Med. Biol. 2014, 773, 435–470. [Google Scholar]
- Starr, D.A.; Fridolfsson, H.N. Interactions Between Nuclei and the Cytoskeleton Are Mediated by SUN-KASH Nuclear-Envelope Bridges. Annu. Rev. Cell Dev. Biol. 2010, 26, 421–444. [Google Scholar] [CrossRef] [Green Version]
- Sosa, B.A.; Kutay, U.; Schwartz, T.U. Structural insights into LINC complexes. Curr. Opin. Struct. Biol. 2013, 23, 285–291. [Google Scholar] [CrossRef]
- Maniotis, A.J.; Chen, C.S.; Ingber, D.E. Demonstration of mechanical connections between integrins, cytoskeletal filaments, and nucleoplasm that stabilize nuclear structure. Proc. Natl. Acad. Sci. USA 1997, 94, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Dahl, K.N.; Ribeiro, A.J.S.; Lammerding, J. Nuclear shape, mechanics, and mechanotransduction. Circ. Res. 2008, 102, 1307–1318. [Google Scholar] [CrossRef]
- Lammerding, J. Mechanics of the nucleus. Compr. Physiol. 2011, 1, 783–807. [Google Scholar] [PubMed]
- Osorio, D.S.; Gomes, E.R. Connecting the nucleus to the cytoskeleton for nuclear positioning and cell migration. Adv. Exp. Med. Biol. 2014, 773, 505–521. [Google Scholar] [PubMed]
- Gomes, E.R.; Jani, S.; Gundersen, G.G. Nuclear movement regulated by Cdc42, MRCK, myosin, and actin flow establishes MTOC polarization in migrating cells. Cell 2005, 121, 451–463. [Google Scholar] [CrossRef]
- Gundersen, G.G.; Worman, H.J. Nuclear Positioning. Cell 2013, 152, 1376–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luxton, G.W.G.; Gomes, E.R.; Folker, E.S.; Vintinner, E.; Gundersen, G.G. Linear Arrays of Nuclear Envelope Proteins Harness Retrograde Actin Flow for Nuclear Movement. Science 2010, 329, 956–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos, R.M.; Sanchís, A.; Tassin, A.M.; Fedriani, C.; Bornens, M. GMAP-210 recruits γ-tubulin complexes to cis-Golgi membranes and is required for Golgi ribbon formation. Cell 2004, 118, 323–335. [Google Scholar] [CrossRef]
- Lindström, L.; Li, T.; Malycheva, D.; Kancharla, A.; Nilsson, H.; Vishnu, N.; Mulder, H.; Johansson, M.; Rosselló, C.A.; Alvarado-Kristensson, M. The GTPase domain of gamma-tubulin is required for normal mitochondrial function and spatial organization. Commun. Biol. 2018, 1, 37. [Google Scholar] [CrossRef]
- Poirier, K.; Lebrun, N.; Broix, L.; Tian, G.; Saillour, Y.; Boscheron, C.; Parrini, E.; Valence, S.; Pierre, B.S.; Oger, M.; et al. Mutations in TUBG1, DYNC1H1, KIF5C and KIF2A cause malformations of cortical development and microcephaly. Nat. Genet. 2013, 45, 639. [Google Scholar] [CrossRef]
- Lindström, L.; Alvarado-Kristensson, M. Characterization of gamma-tubulin filaments in mammalian cells. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 158–171. [Google Scholar]
- De Keersmaecker, K.; Rocnik, J.L.; Bernad, R.; Lee, B.H.; Leeman, D.; Gielen, O.; Verachtert, H.; Folens, C.; Munck, S.; Marynen, P.; et al. Kinase activation and transformation by NUP214-ABL1 is dependent on the context of the nuclear pore. Mol. Cell 2008, 31, 134–142. [Google Scholar] [CrossRef]
- Takeda, A.; Sarma, N.J.; Abdul-Nabi, A.M.; Yaseen, N.R. Inhibition of CRM1-mediated nuclear export of transcription factors by leukemogenic NUP98 fusion proteins. J. Biol. Chem. 2010, 285, 16248–16257. [Google Scholar] [CrossRef]
- Joseph, J.; Dasso, M. The nucleoporin Nup358 associates with and regulates interphase microtubules. Febs Lett. 2008, 582, 190–196. [Google Scholar] [CrossRef]
- Murawala, P.; Tripathi, M.M.; Vyas, P.; Salunke, A.; Joseph, J. Nup358 interacts with APC and plays a role in cell polarization. J. Cell Sci. 2009, 122, 3113–3122. [Google Scholar] [CrossRef] [Green Version]
- Collin, L.; Schlessinger, K.; Hall, A. APC nuclear membrane association and microtubule polarity. Biol. Cell 2008, 100, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Hubert, T.; Vandekerckhove, J.; Gettemans, J. Exo70-mediated recruitment of nucleoporin Nup62 at the leading edge of migrating cells is required for cell migration. Traffic Cph. Den. 2009, 10, 1257–1271. [Google Scholar] [CrossRef]
- Gloerich, M.; Vliem, M.J.; Prummel, E.; Meijer, L.A.T.; Rensen, M.G.A.; Rehmann, H.; Bos, J.L. The nucleoporin RanBP2 tethers the cAMP effector Epac1 and inhibits its catalytic activity. J. Cell Biol. 2011, 193, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Takahashi, M.; Li, Y.; Dillon, T.J.; Kaech, S.; Stork, P.J.S. The interaction of Epac1 and Ran promotes Rap1 activation at the nuclear envelope. Mol. Cell. Biol. 2010, 30, 3956–3969. [Google Scholar] [CrossRef]
- Hatch, E.M.; Fischer, A.H.; Deerinck, T.J.; Hetzer, M.W. Catastrophic Nuclear Envelope Collapse in Cancer Cell Micronuclei. Cell 2013, 154, 48–60. [Google Scholar] [CrossRef]
- Crasta, K.; Ganem, N.J.; Dagher, R.; Lantermann, A.B.; Ivanova, E.V.; Pan, Y.; Nezi, L.; Protopopov, A.; Chowdhury, D.; Pellman, D. DNA breaks and chromosome pulverization from errors in mitosis. Nature 2012, 482, 53–58. [Google Scholar] [CrossRef]
- Vargas, J.D.; Hatch, E.M.; Anderson, D.J.; Hetzer, M.W. Transient nuclear envelope rupturing during interphase in human cancer cells. Nucleus 2012, 3, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Malhas, A.N.; Vaux, D.J. Nuclear envelope invaginations and cancer. Adv. Exp. Med. Biol. 2014, 773, 523–535. [Google Scholar]
- Clubb, B.H.; Locke, M. 3T3 cells have nuclear invaginations containing F-actin. Tissue Cell 1998, 30, 684–691. [Google Scholar] [CrossRef]
- Brandes, D.; Schofield, B.H.; Anton, E. Nuclear mitochondria? Science 1965, 149, 1373–1374. [Google Scholar] [CrossRef]
- Fricker, M.; Hollinshead, M.; White, N.; Vaux, D. Interphase nuclei of many mammalian cell types contain deep, dynamic, tubular membrane-bound invaginations of the nuclear envelope. J. Cell Biol. 1997, 136, 531–544. [Google Scholar] [CrossRef]
- Malhas, A.; Goulbourne, C.; Vaux, D.J. The nucleoplasmic reticulum: Form and function. Trends Cell Biol. 2011, 21, 362–373. [Google Scholar] [CrossRef]
- True, L.D.; Jordan, C.D. The cancer nuclear microenvironment: Interface between light microscopic cytology and molecular phenotype. J. Cell. Biochem. 2008, 104, 1994–2003. [Google Scholar] [CrossRef]
- Fricker, M.; Hollinshead, M.; White, N.; Vaux, D. The convoluted nucleus. Trends Cell Biol. 1997, 7, 181. [Google Scholar] [CrossRef]
- Gupton, S.L.; Collings, D.A.; Allen, N.S. Endoplasmic reticulum targeted GFP reveals ER organization in tobacco NT-1 cells during cell division. Plant Physiol. Biochem. 2006, 44, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Fischer, A.H.; Taysavang, P.; Jhiang, S.M. Nuclear envelope irregularity is induced by RET/PTC during interphase. Am. J. Pathol. 2003, 163, 1091–1100. [Google Scholar] [CrossRef]
Figure 1.
A model for the role of nuclear γ-tubulin in chromatin enclosing, inner nuclear membrane (INM) protein recruitment, and nuclear pore complex (NPC) formation in nuclear disassembly and reassembly upon cell division. (a) Schematic representation of the nucleus and nuclear envelope (NE) during interphase and subsequent mitosis. Post-mitotic reassembly of the NE is initiated by changes at the chromatin surface (i.e., removal of mitotic histone marks by phosphatases) and dephosphorylation-induced binding of nuclear envelope transmembrane proteins (NETs) and their associated membranes to chromatin. Engulfment of recently separated sister chromatids by the NE occurs via deposits of membrane fragments on the chromatin surface that trigger the enveloping process. The NE is reformed from endoplasmic reticulum (ER) membranes, which contact chromatin either as tubules or sheets. After membrane deposition, NPC assembly allows importation of the elements for nuclear lamina assembly. Microtubule organizer γ-tubulin plays a distinct role in promoting this NE formation by providing a γ-tubulin boundary indispensable for NE assembly. At the onset of mitosis, the lamina meshwork is disrupted, but the γ-tubulin boundary around the mitotic chromosomes is maintained. During mitosis, chromatin-associated γ-strings link the sister chromatids to the cytosolic γ-string pool. Finally, at anaphase/telophase, the γ-tubulin boundary composed of cytosolic and chromatin-associated γ-strings forms a supporting scaffold that assists the formation of the nuclear envelope. (b) Magnified boxed area of NE at interphase: During interphase, γ-tubulin bridges connect the cytosolic and the nuclear γ-tubulin pools ([123], reviewed in [128,129]). The LINC protein nesprins recruit centrosomal proteins and regulate the nucleation of microtubules from the NE in myotubes [130]. The INM protein Samp1 is in contact with both γ-tubulin and SUN1 [131]. At the beginning of mitosis, during the rupture of the NE to the spindle microtubules, Samp1 might recruit γ-tubulin from the fenestrated NE. In the spindle, γ-tubulin and Samp1 complex together with augmins would potentially assist in the nucleation of the microtubules. Not to scale.
Figure 1.
A model for the role of nuclear γ-tubulin in chromatin enclosing, inner nuclear membrane (INM) protein recruitment, and nuclear pore complex (NPC) formation in nuclear disassembly and reassembly upon cell division. (a) Schematic representation of the nucleus and nuclear envelope (NE) during interphase and subsequent mitosis. Post-mitotic reassembly of the NE is initiated by changes at the chromatin surface (i.e., removal of mitotic histone marks by phosphatases) and dephosphorylation-induced binding of nuclear envelope transmembrane proteins (NETs) and their associated membranes to chromatin. Engulfment of recently separated sister chromatids by the NE occurs via deposits of membrane fragments on the chromatin surface that trigger the enveloping process. The NE is reformed from endoplasmic reticulum (ER) membranes, which contact chromatin either as tubules or sheets. After membrane deposition, NPC assembly allows importation of the elements for nuclear lamina assembly. Microtubule organizer γ-tubulin plays a distinct role in promoting this NE formation by providing a γ-tubulin boundary indispensable for NE assembly. At the onset of mitosis, the lamina meshwork is disrupted, but the γ-tubulin boundary around the mitotic chromosomes is maintained. During mitosis, chromatin-associated γ-strings link the sister chromatids to the cytosolic γ-string pool. Finally, at anaphase/telophase, the γ-tubulin boundary composed of cytosolic and chromatin-associated γ-strings forms a supporting scaffold that assists the formation of the nuclear envelope. (b) Magnified boxed area of NE at interphase: During interphase, γ-tubulin bridges connect the cytosolic and the nuclear γ-tubulin pools ([123], reviewed in [128,129]). The LINC protein nesprins recruit centrosomal proteins and regulate the nucleation of microtubules from the NE in myotubes [130]. The INM protein Samp1 is in contact with both γ-tubulin and SUN1 [131]. At the beginning of mitosis, during the rupture of the NE to the spindle microtubules, Samp1 might recruit γ-tubulin from the fenestrated NE. In the spindle, γ-tubulin and Samp1 complex together with augmins would potentially assist in the nucleation of the microtubules. Not to scale.

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alvarado-Kristensson, M.; Rosselló, C.A. The Biology of the Nuclear Envelope and Its Implications in Cancer Biology. Int. J. Mol. Sci. 2019, 20, 2586. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102586
AMA Style
Alvarado-Kristensson M, Rosselló CA. The Biology of the Nuclear Envelope and Its Implications in Cancer Biology. International Journal of Molecular Sciences. 2019; 20(10):2586. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102586
Chicago/Turabian StyleAlvarado-Kristensson, Maria, and Catalina Ana Rosselló. 2019. "The Biology of the Nuclear Envelope and Its Implications in Cancer Biology" International Journal of Molecular Sciences 20, no. 10: 2586. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102586
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.