Proteomic Landscape of the Mature Roots in a Rubber-Producing Grass Taraxacum Kok-saghyz

 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
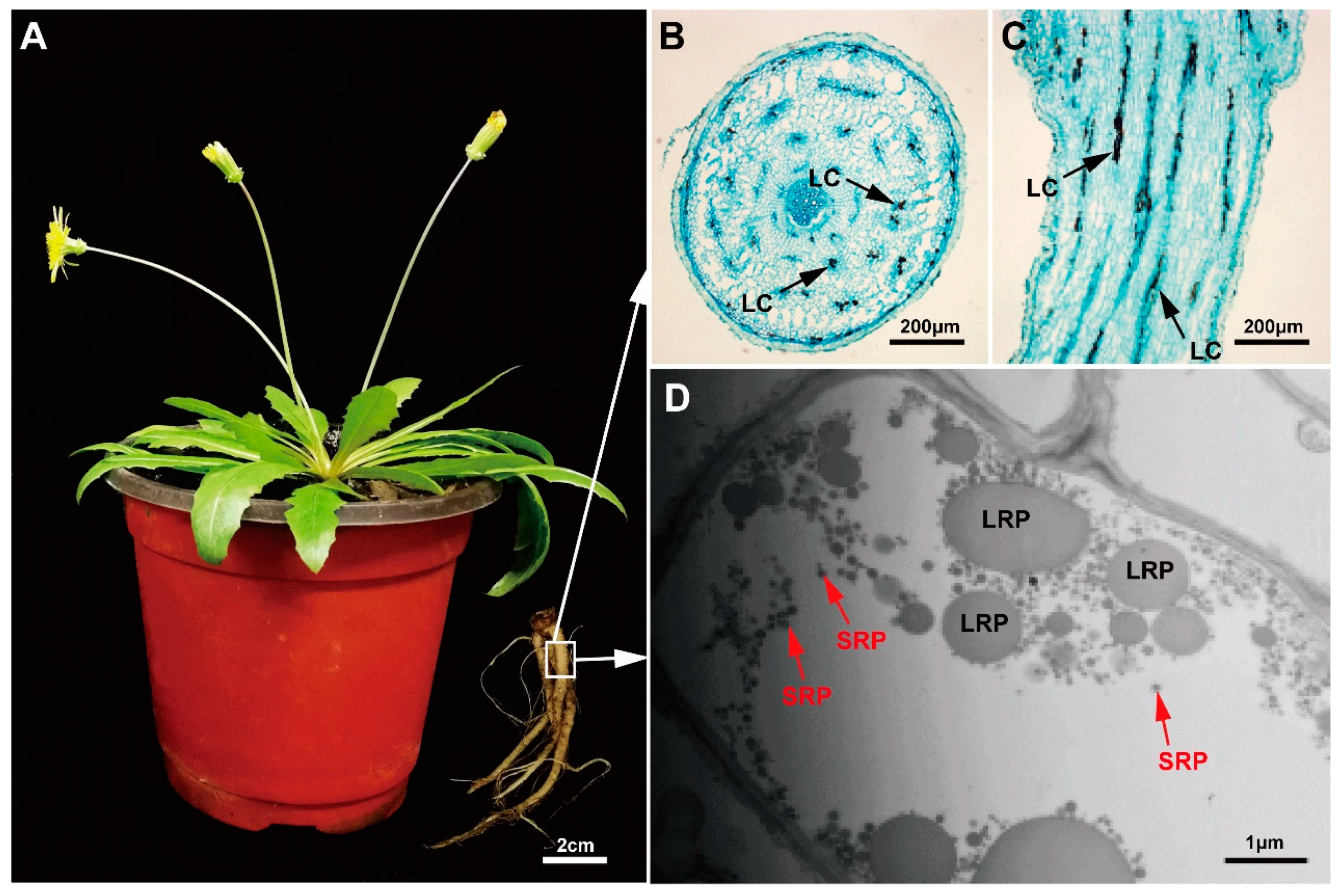
2.1. Morphological Observation of Rubber Particles in the Main Roots of TKS
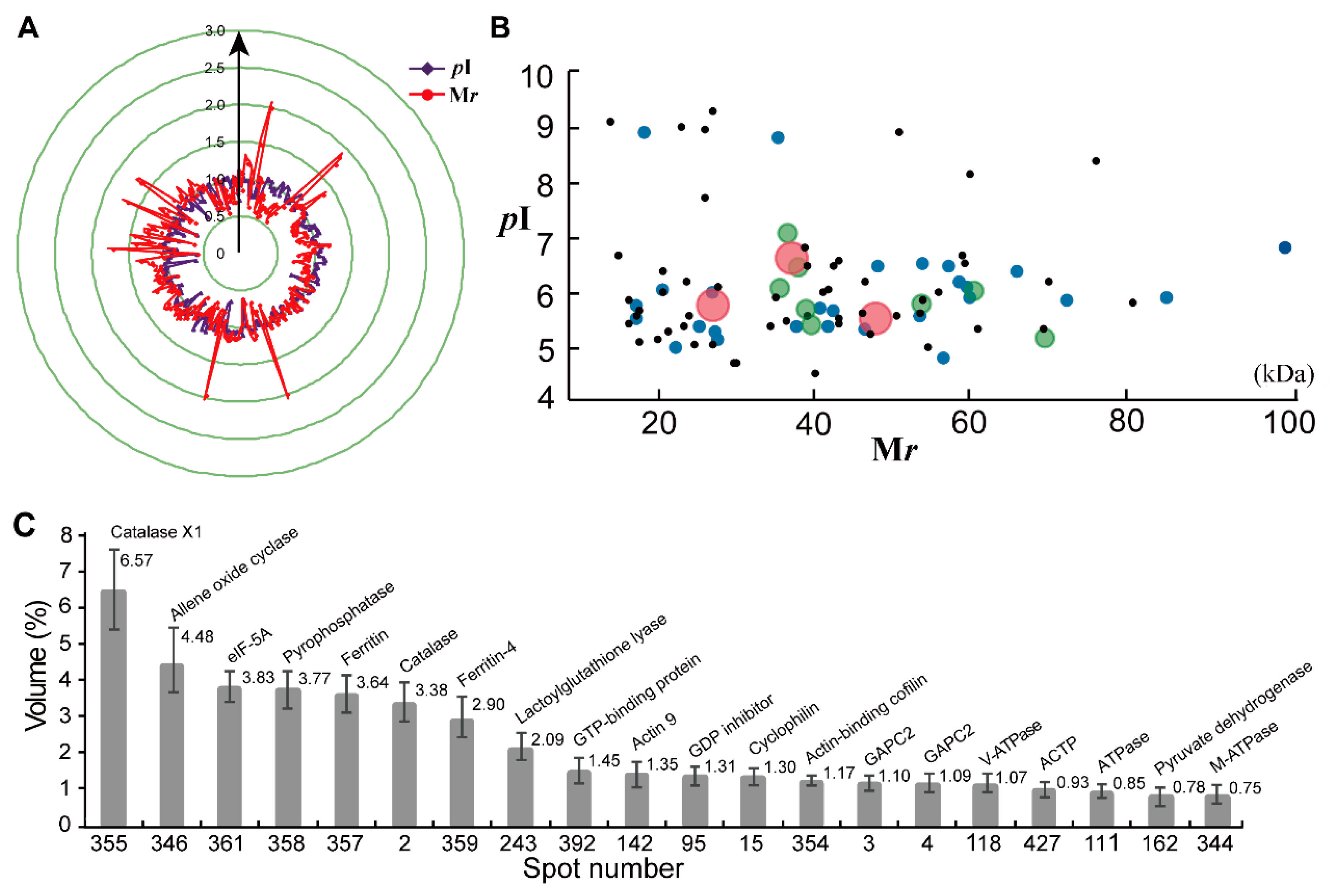
2.2. Establishment of a Visual Two-Dimensional Gel Electrophoresis (2-DE) Proteome Map and Mass Spectrometry (MS) Identification of High Abundance Proteins in Taraxacum Kok-saghyz (TKS) Roots
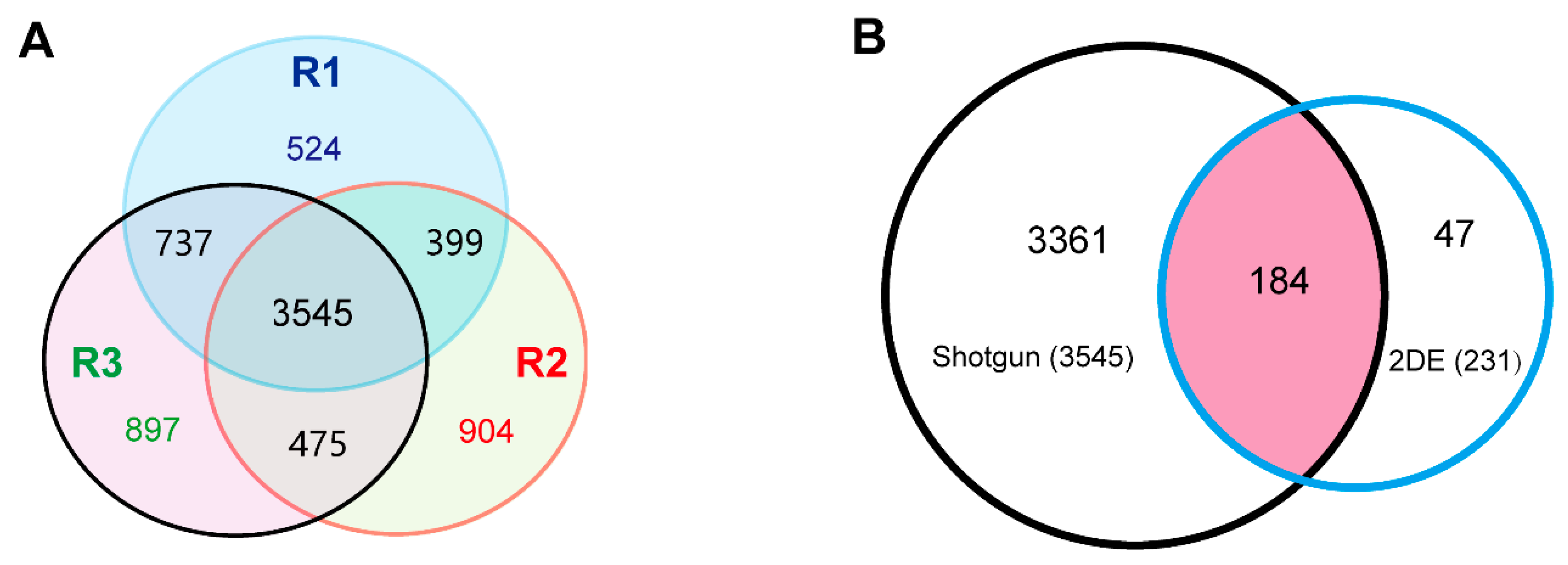
2.3. High-Throughput Shotgun Proteomic Analysis of the 6M Roots of TKS
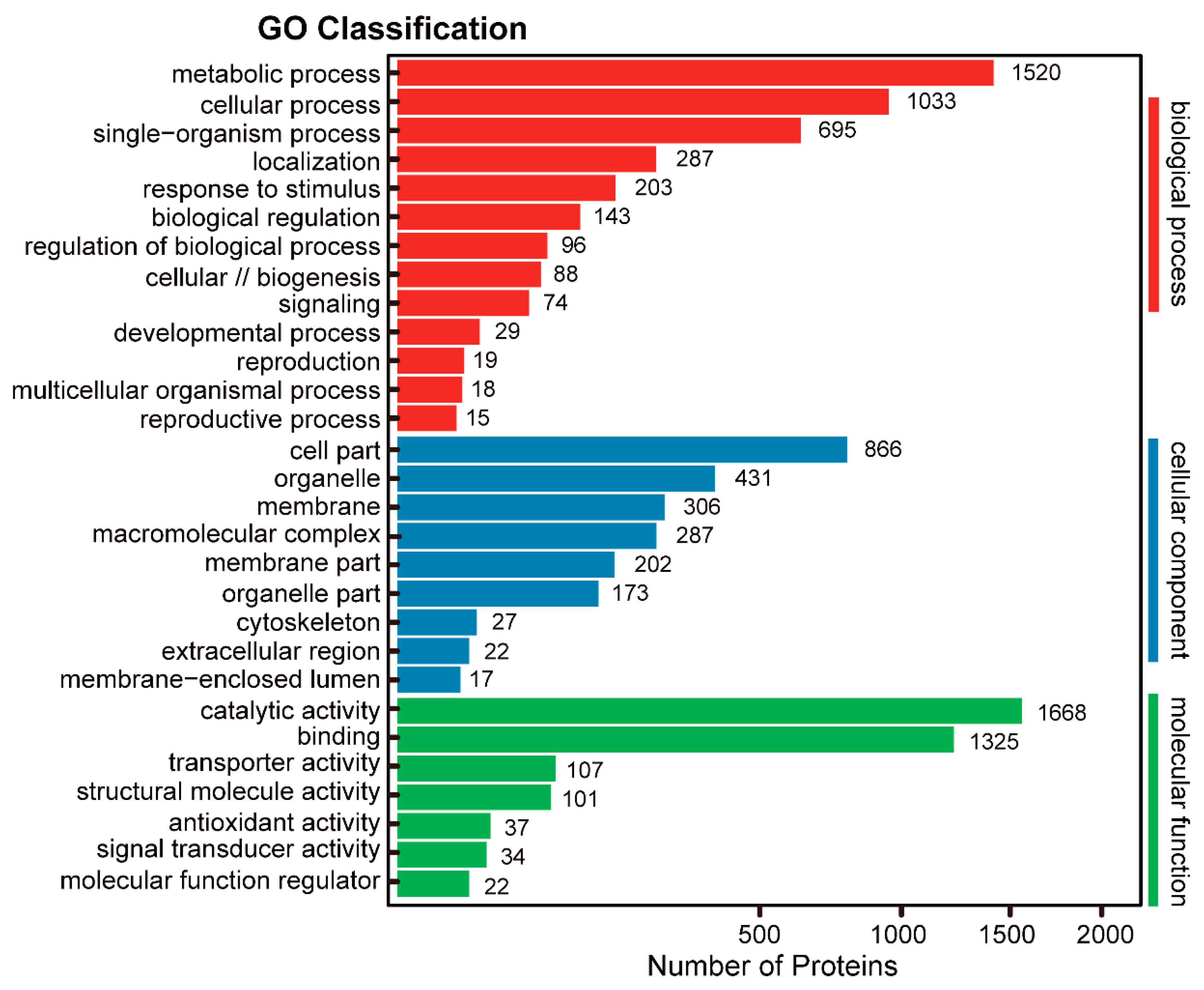
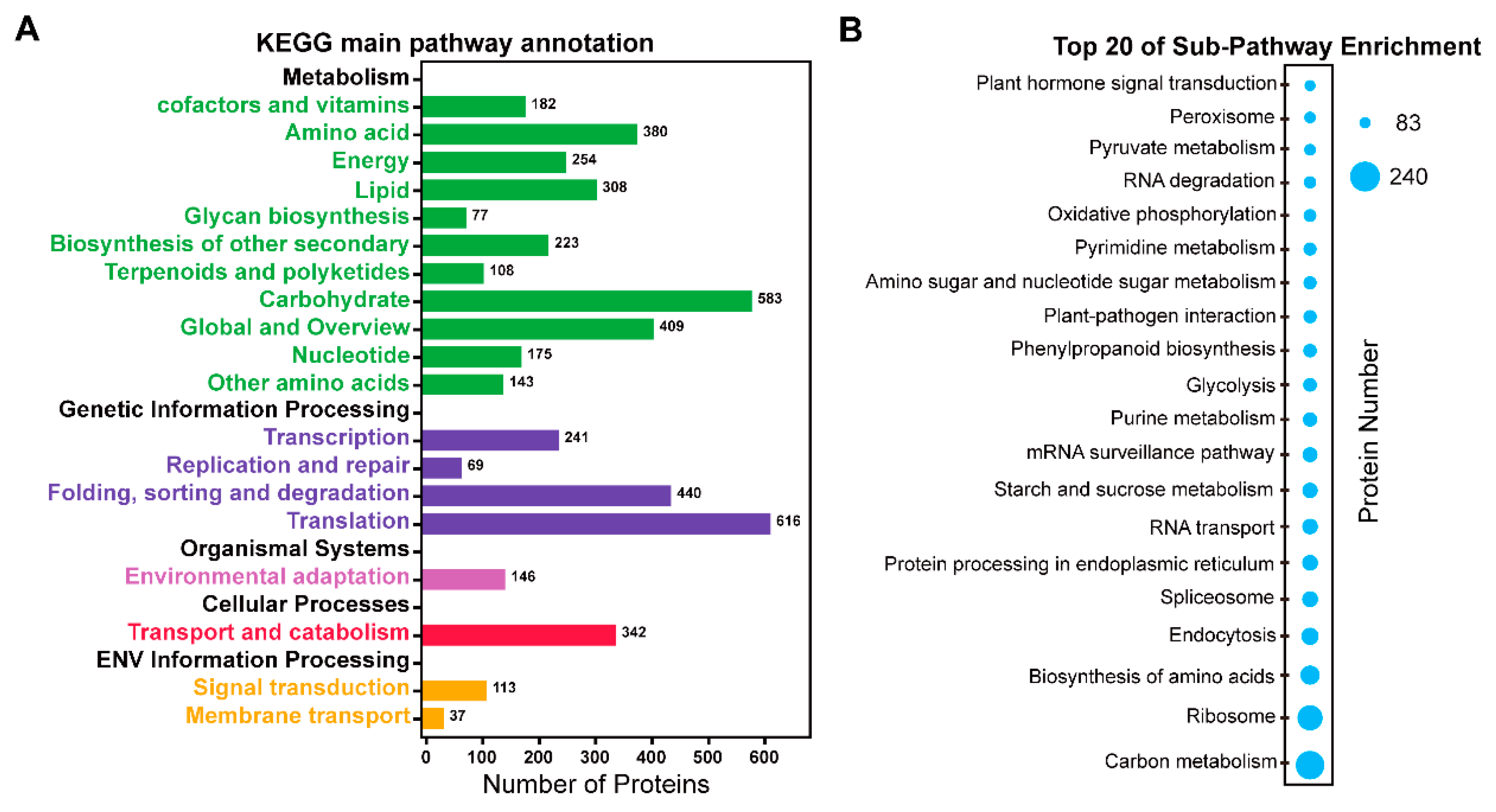
2.4. Pathway Analysis of the Identified Proteins in 6M TKS Roots
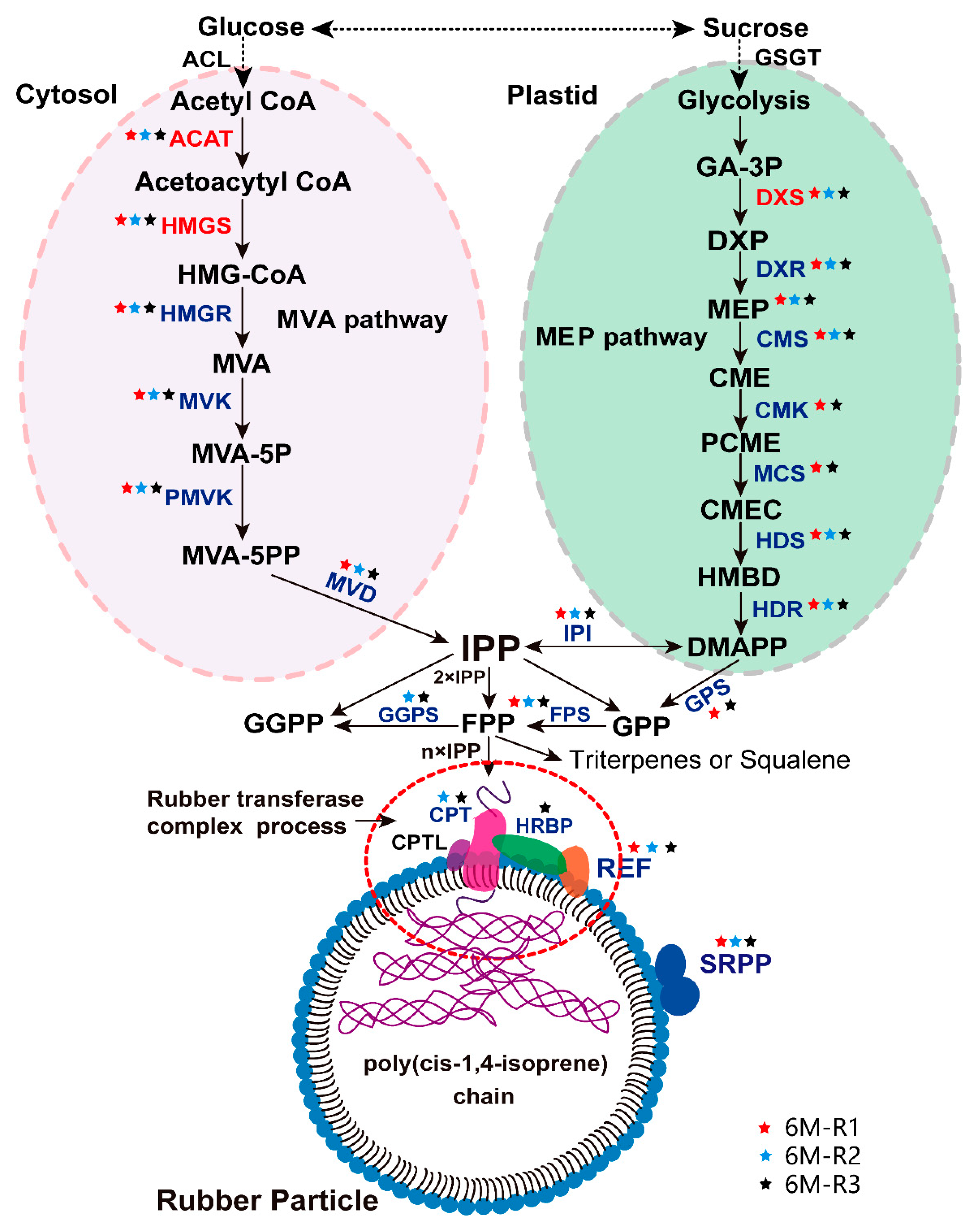
2.5. The Proteins Involved in Natural Rubber Biosynthesis in TKS Roots
3. Discussion
3.1. The First Visual Proteome Based on 2-DE Gel Demonstrated Many Protein Isoforms in the Mature TKS Roots
3.2. Large-Scale Shotgun Proteomics Landscape Revealed both Mevalonate Acid (MVA) and Methylerythritol Phosphate (MEP) Pathways are Important for Natural Rubber Biosynthesis (NRB) in the Mature TKS Roots
3.3. Almost All NRB-Related Proteins Can Be Identified From the Mature TKS Roots
4. Materials and Methods
4.1. Plant Growth Conditions and Root Collection
4.2. Morphological Observation of Rubber Particles in TKS Roots
4.3. Protein Extraction and Two-Dimensional Gel Electrophoresis
4.4. Identification of Protein Spots via MALDI-TOF/TOF-MS
4.5. Large-Scale Shotgun Proteomics of TKS Roots
4.6. Functional Classification, Hierarchical Clustering and Pathway Analysis of All the Identified Proteins From TKS Roots
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACL | ATP citrate lyase |
| ACAT | acetoacetyl-CoA thiolase |
| CHAPS | 3-[(3-Cholamidopropyl) dimethylammonio] propanesulfonate |
| CPT | cis-isoprene transferase |
| CPTL | cis-isoprene transferase like |
| DMAPP | dimethylallyl diphosphate |
| DXS | 1-deoxy-D-xylulose-5-phosphate synthase |
| eIF | eukaryotic translation initiation factor |
| FPS | farnesyl diphosphate synthase |
| GAPC | glyceraldehyde-3-phosphate dehydrogenase |
| GGPPS | geranylgeranyl diphosphate synthase |
| GPP | geranyl pyrophosphate |
| GPS | geranylgeranyl diphosphate synthase |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| HMGS | 3-hydroxy-glutaryl-CoA reductase |
| HMGR | hydroxy-methyl-glutaryl-CoA reductase |
| IPI | isopentenyl-diphosphate delta-isomerase |
| IPG | immobilized pH gradient |
| iTRAQ | isobaric tags for relative and absolute quantification |
| LRP | large rubber particle protein |
| MEP | methyl-D-erythritol-4-phosphate |
| MVA | mevalonate acid |
| MVD | diphosphomevalonate decarboxylase |
| MVK | mevalonate kinase |
| MS | mass spectrometry |
| NR | natural rubber |
| NRB | natural rubber biosynthesis |
| PMVK | mevalonate disphosphate decarboxylase |
| REF | rubber elongation factor |
| RTA | rubber transferase activating factor |
| SRPP | small rubber particle protein |
| 2-DE | two-dimensional gel electrophoresis |
References
- Mooibroek, H.; Cornish, K. Alternative sources of natural rubber. Appl. Microbiol. Biotechnol. 2000, 53, 355–365. [Google Scholar] [CrossRef]
- Wang, X.C.; Wang, D.; Sun, Y.; Yang, Q.; Chang, L.L.; Wang, L.M.; Meng, X.R.; Huang, Q.X.; Jin, X.; Tong, Z. Comprehensive proteomics analysis of laticifer latex reveals new insights into ethylene stimulation of natural rubber production. Sci. Rep. 2015, 5, 13778. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.; Hillebrand, A.; Wurbs, D.; Wahler, D.; Lenders, M.; Gronover, C.; Prüfer, D. Molecular cloning and characterization of rubber biosynthetic genes from Taraxacum kok-saghyz. Plant Mol. Biol. Rep. 2010, 28, 277–284. [Google Scholar] [CrossRef]
- Cornish, K.; Kopicky, S.L.; Mcnulty, S.K.; Amstutz, N.; Chanon, A.M.; Walker, S.; Kleinhenz, M.D.; Miller, A.R.; Streeter, J.G. Temporal diversity of Taraxacum kok-saghyz plants reveals high rubber yield phenotypes. Biodiversitas 2016, 17, 847–856. [Google Scholar] [CrossRef]
- Edathil, T.T. South American leaf blight: a potential threat to the natural rubber industry in Asia and Africa. Trop. Pest Manag. 1986, 4, 296–303. [Google Scholar] [CrossRef]
- Ramirez-Cadavid, D.A.; Cornish, K.; Michel, J. Taraxacum kok-saghyz (TK): compositional analysis of a feedstock for natural rubber and other bioproducts. Ind. Crop. Prod. 2017, 107, 624–640. [Google Scholar] [CrossRef]
- Collins-Silva, J.; Nural, A.T.; Skaggs, A.; Scott, D.; Hathwaik, U.; Woolsey, R.; Schegg, K.; McMahan, C.; Whalen, M.; Cornish, K.; et al. Altered levels of the Taraxacum kok-saghyz (Russian dandelion) small rubber particle protein, TKSRPP3, result in qualitative and quantitative changes in rubber metabolism. Phytochemistry 2012, 79, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.J.; Ryu, S.B.; Kwak, Y.S.; Kang, H. A novel cDNA from Parthenium argentatum Gray enhances the rubber biosynthetic activity in vitro. J. Exp. Bot. 2004, 396, 377–385. [Google Scholar] [CrossRef]
- Ikeda, Y.; Junkong, P.; Ohashi, T.; Phakkeeree, T.; Sakaki, Y.; Tohsan, A.; Kohjiya, S.; Cornish, K. Strain-induced crystallization behaviour of natural rubbers from guayule and rubber dandelion revealed by simultaneous time-resolved WAXD/tensile measurements: indispensable function for sustainable resources. RSC Adv. 2016, 6, 95601–95610. [Google Scholar] [CrossRef]
- Tang, C.R.; Yang, M.; Fang, Y.J.; Luo, Y.F.; Gao, S.H.; Xiao, X.H.; An, Z.W.; Zhou, B.H.; Zhang, B.; Tan, X.Y.; et al. The rubber tree genome reveals new insights into rubber production and species adaptation. Nat. Plants 2016, 6, 16073. [Google Scholar] [CrossRef] [PubMed]
- Whaley, W.G.; Bowen, J.S. Russian dandelion (kok-saghyz). An emergency source of natural rubber. Misc. Publ. US Dept. Agric. 1947, 618, 1–212. [Google Scholar]
- van Beilen, J.B.; Poirier, Y. Establishment of new crops for the production of natural rubber. Trends Biotechnol. 2007, 25, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Kirschner, J.; Stepanek, J.; Cerny, T.; Heer, P.; Dijk, P.J. Available ex situ germplasm of the potential rubber crop Taraxacum kok-saghyz belongs to a poor rubber producer, T. brevicorniculatum (Compositae–Crepidinae). Genet. Resour. Crop Evol. 2013, 2, 455–471. [Google Scholar] [CrossRef]
- Kreuzberger, M.; Hahn, T.; Zibek, S.; Schiemann, J.; Thiele, K. Seasonal pattern of biomass and rubber and inulin of wild Russian dandelion (Taraxacum kok-saghyz L. Rodin) under experimental field conditions. Eur. J. Agron. 2016, 80, 66–77. [Google Scholar] [CrossRef]
- Stolze, A.; Wanke, A.; van Deenen, N.; Geyer, R.; Prüfer, D.; Gronover, C.S. Development of rubber-enriched dandelion varieties by metabolic engineering of the inulin pathway. Plant Biotechnol. J. 2017, 6, 740–753. [Google Scholar] [CrossRef]
- Lin, T.; Xu, X.; Ruan, J.; Liu, S.Z.; Wu, S.G.; Shao, X.J.; Wang, X.B.; Gan, L.; Qin, B.; Yang, Y.S.; et al. Genome analysis of Taraxacum kok-saghyz Rodin provides new insights into rubber biosynthesis. Natl. Sci. Rev. 2017, 5, 78–87. [Google Scholar] [CrossRef]
- Ramirez-Cadavid, D.A.; Valles-Ramirez, S.; Cornish, K.; Michel, F.C. Simultaneous quantification of rubber, inulin, and resins in Taraxacum kok-saghyz (TK) roots by sequential solvent extraction. Plant Biotechnol. J. 2018, 122, 647–656. [Google Scholar] [CrossRef]
- Bach, T.J. Some new aspects of isoprenoid biosynthesis in plants a review. Lipids 1995, 3, 191–202. [Google Scholar] [CrossRef]
- Walther, T.C.; Farese, R.V. The life of lipid droplets. Biochim. Biophys. Acta 2009, 6, 459–466. [Google Scholar] [CrossRef]
- Seetang-Nun, Y.; Sharkey, T.D.; Suvachittanont, W. Molecular cloning and characterization of two cDNAs encoding 1-deoxy-D-xylulose 5-phosphate reductoisomerase from Hevea brasiliensis. J. Plant Physiol. 2007, 9, 991–1002. [Google Scholar] [CrossRef]
- Yagami, T.; Haishima, Y.; Tsuchiya, T.; Tomitaka-Yagami, A.; Kano, H.; Matsunaga, K. Proteomic analysis of putative latex allergens. Int. Arch. Allergy Immunol. 2004, 135, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Shi, M.J.; Lu, X.L.; Ma, R.F.; Wu, C.G.; Guo, A.P.; Peng, M.; Tian, W.M. A method for protein extraction from different subcellular fractions of laticifer latex in Hevea brasiliensis compatible with 2-DE and MS. Proteome Sci. 2010, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, A.; Bachi, A.; Fasoli, E.; Boschetti, E.; Peltre, G.; Sénéchal, H.; Sutrad, J.P.; Citteriob, A.; Righettib, P.G. In-depth exploration of Hevea brasiliensis latex proteome and hidden allergens via combinatorial peptide ligand libraries. J. Proteom. 2010, 73, 1368–1380. [Google Scholar] [CrossRef]
- Habib, M.H.; Yuen, G.C.; Othman, F.; Zainudin, N.N.; Latiff, A.A.; Ismail, M.N. Proteomics analysis of latex from Hevea brasiliensis (clone RRIM 600). Biochem. Cell Biol. 2017, 95, 232–242. [Google Scholar] [CrossRef]
- Tong, Z.; Wang, D.; Sun, Y.; Yang, Q.; Meng, X.R.; Wang, L.M.; Feng, W.Q.; Li, L.; Wurtele, E.S.; Wang, X.C. Comparative proteomics of rubber latex revealed multiple protein species of REF/SRPP family respond diversely to ethylene stimulation among different rubber tree clones. Int. J. Mol. Sci. 2017, 18, 958. [Google Scholar] [CrossRef] [PubMed]
- Rojruthai, P.; Sakdapipanich, J.T.; Takahashi, S.; Hyegin, L.; Noike, M.; Koyama, T.; Tanaka, Y. In vitro synthesis of high molecular weight rubber by Hevea small rubber particles. J. Biosci. Bioeng. 2010, 109, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.L.; Xia, K.C.; Dai, L.J.; Kang, G.J.; Li, Y.; Nie, Z.Y.; Duan, C.F.; Zeng, R.Z. Proteome analysis of the large and the small rubber particles of Hevea brasiliensis using 2D-DIGE. Plant Phys. Biochem. 2012, 60, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.J.; Kang, G.J.; Li, Y.; Nie, Z.Y.; Duan, C.F.; Zeng, R.Z. In-depth proteome analysis of the rubber particle of Hevea brasiliensis (para rubber tree). Plant Mol. Biol. 2013, 82, 155–168. [Google Scholar] [CrossRef]
- Dai, L.J.; Kang, G.J.; Nie, Z.Y.; Li, Y.; Zeng, R.Z. Comparative proteomic analysis of latex from Hevea brasiliensis treated with ethrel and methyl jasmonate using iTRAQ-coupled two-dimensional LC-MS/MS. J. Proteom. 2016, 132, 167–175. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Y.; Tong, Z.; Yang, Q.; Chang, L.L.; Meng, X.R.; Wang, L.M.; Tian, W.M.; Wang, X.C. A protein extraction method for low protein concentration solutions compatible with the proteomic analysis of rubber particles. Electrophoresis 2016, 37, 2930–2939. [Google Scholar] [CrossRef]
- Yamashita, S.; Yamaguchi, H.; Waki, T.; Aoki, Y.; Mizuno, M.; Yanbe, F.; Ishii, T.; Funaki, A.; Tozawa, Y.; Miyagi-Inoue, Y.; et al. Identification and reconstitution of the rubber biosynthetic machinery on rubber particles from Hevea brasiliensis. eLife 2016, 5, e19022. [Google Scholar] [CrossRef]
- Wahler, D.; Colby, T.; Kowalski, N.A.; Harzen, A.; Wotzka, S.Y.; Hillebrand, A.; Fischer, R.; Helsper, J.; Schmidt, J.; Gronover, C.S.; et al. Proteomic analysis of latex from the rubber-producing plant Taraxacum brevicorniculatum. Proteomics 2012, 12, 901–905. [Google Scholar] [CrossRef]
- Singh, A.P.; Wi, S.G.; Chung, G.C.; Kim, Y.S.; Kang, H. The micromorphology and protein characterization of rubber particles in Ficus carica, Ficus benghalensis and Hevea brasiliensis. J. Exp. Bot. 2003, 54, 985–992. [Google Scholar] [CrossRef]
- Jorrin-Novo, V.J.; Pascual, J.; Sanchez-Lucas, R.; Romero-Rodriguez, M.C.; Rodriguez-Ortega, M.J.; Lenz, C.; Valledor, L. Fourteen years of plant proteomics reflected in proteomics: moving from model species and 2DE-based approaches to orphan species and gel-free platforms. Proteomics 2015, 15, 1089–1112. [Google Scholar] [CrossRef]
- Wang, D.; Sun, Y.; Chang, L.L.; Tong, Z.; Xie, Q.L.; Jin, X.; Zhu, L.P.; He, P.; Li, H.B.; Wang, X.C. Subcellular proteome profiles of different latex fractions revealed washed solutions from rubber particles contain crucial enzymes for natural rubber biosynthesis. J. Proteom. 2018, 182, 53–64. [Google Scholar] [CrossRef]
- Cho, W.K.; Jo, Y.; Chu, H.; Park, S.H.; Kim, K.H. Integration of latex protein sequence data provides comprehensive functional overview of latex proteins. Mol. Biol. Rep. 2014, 41, 1469–1481. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Feeney, M.; Ahmadi, M.; Lonoce, C.; Sajari, R.; Di Cola, A.; Frigerio, L. Subcellular localization and interactions among rubber particle proteins from Hevea brasiliensis. J. Exp. Bot. 2017, 68, 5045–5055. [Google Scholar] [CrossRef] [PubMed]
- Sando, T.; Takeno, S.; Watanabe, N.; Okumoto, H.; Kuzuyama, T.; Yamashita, A.; Hattori, M.; Ogasawara, N.; Fukusaki, E.; Kobayashi, A. Cloning and characterization of the 2-C-methyl-D-erythritol 4-phosphate (MEP) pathway genes of a natural-rubber producing plant, Hevea brasiliensis. Biosci. Biotechnol. Biochem. 2008, 11, 2903–2917. [Google Scholar] [CrossRef]
- Wu, C.T.; Li, Y.; Nie, Z.Y.; Dai, L.J.; Kang, G.J.; Zeng, R.Z. Molecular cloning and expression analysis of the mevalonate diphosphate decarboxylase gene from the latex of Hevea brasiliensis. Tree Genet. Genomes 2017, 13, 22. [Google Scholar] [CrossRef]
- Lau, N.S.; Makita, Y.; Kawashima, M.; Taylor, T.D.; Kondo, S.; Othman, A.S.; Shu-Chien, A.C.; Matsui, M. The rubber tree genome shows expansion of gene family associated with rubber biosynthesis. Sci. Rep. 2016, 6, 28594. [Google Scholar] [CrossRef]
- Puskas, J.E.; Gautriaud, E.; Deffieux, A.; Kennedy, J.P. Natural rubber biosynthesis-a living carbocationic polymerization. Prog. Polym. Sci. 2006, 31, 533–548. [Google Scholar] [CrossRef]
- Cornish, K. Similarities and differences in rubber biochemistry among plant species. Phytochemistry 2001, 57, 1123–1134. [Google Scholar] [CrossRef]
- Kirby, J.; Keasling, J.D. Biosynthesis of plant isoprenoids perspectives for microbial engineering. Annu. Rev. Plant Biol. 2009, 60, 335–355. [Google Scholar] [CrossRef]
- Putter, K.M.; van Deenen, N.; Unland, K.; Prufer, D.; Gronover, C.S. Isoprenoid biosynthesis in dandelion latex is enhanced by the overexpression of three key enzymes involved in the mevalonate pathway. BMC Plant Biol. 2017, 17, 88. [Google Scholar] [CrossRef]
- Sirinupong, N.; Suwanmanee, P.; Doolittle, R.F.; Suvachitanont, W. Molecular cloning of a new cDNA and expression of 3-hydroxy-3-methylglutaryl-CoA synthase gene from Hevea brasiliensis. Planta 2005, 221, 502–512. [Google Scholar] [CrossRef] [PubMed]
- Grabinska, K.A.; Park, E.J.; Sessa, W.C. cis-Prenyltransferase: new insights into protein glycosylation, rubber synthesis, and human diseases. J. Biol. Chem. 2016, 291, 18582–18590. [Google Scholar] [CrossRef]
- Schaller, H.; Crausem, B.; Benveniste, P.; Chye, M.L.; Tan, C.T.; Song, Y.H.; Chua, N.H. Expression of the Hevea brasiliensis Müll. Arg. 3-Hydroxy-3-Methylglutaryl-Coenzyme A Reductase 1 in tobacco results in sterol overproduction. Plant Physiol. 1995, 109, 761–770. [Google Scholar] [CrossRef]
- Sando, T.; Hayashi, T.; Takeda, T.; Akiyama, Y.; Nakazawa, Y.; Fukusaki, E.; Kobayashi, A. Histochemical study of detailed laticifer structure and rubber biosynthesis-related protein localization in Hevea brasiliensis using spectral confocal laser scanning microscopy. Planta 2009, 230, 215–225. [Google Scholar] [CrossRef]
- Chye, M.L.; Tan, C.T.; Chua, N.H. Three genes encode 3-hydroxy-3-methylglutaryl-coenzyme A reductase in Hevea brasiliensis: hmg1 and hmg3 are differentially expressed. Plant Mol. Biol. 1992, 19, 473–484. [Google Scholar] [CrossRef]
- Williamson, B.Y.; Kekwick, R.G. The enzymes forming isopentenyl pyrophosphate from 5-phosphomevalonate (mevalonate 5-phosphate) in the latex of Hevea brasiliensis. Biochem. J. 1971, 124, 407–417. [Google Scholar]
- Ko, J.H.; Chow, K.S.; Han, K.H. Transcriptome analysis reveals novel features of the molecular events occurring in the laticifers of Hevea brasiliensis (para rubber tree). Plant Mol. Biol. 2003, 53, 479–492. [Google Scholar] [CrossRef]
- Swiezewska, E.; Danikiewicz, W. Polyisoprenoids: Structure, biosynthesis and function. Prog. Lipid Res. 2005, 44, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.S.; Wan, K.L.; Isa, M.N.; Bahari, A.; Tan, S.H.; Harikrishna, K.; Yeang, H.Y. Insights into rubber biosynthesis from transcriptome analysis of Hevea brasiliensis latex. J. Exp. Bot. 2007, 58, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.S.; Mat-Isa, M.N.; Bahari, A.; Ghazali, A.K.; Alias, H.; Mohd-Zainuddin, Z.; Hoh, C.C.; Wan, K.L. Metabolic routes affecting rubber biosynthesis in Hevea brasiliensis latex. J. Exp. Bot. 2012, 63, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Makita, Y.; Ng, K.K.; Veera Singham, G.; Kawashima, M.; Hirakawa, H.; Sato, S.; Othman, A.S.; Matsui, M. Large-scale collection of full-length cDNA and transcriptome analysis in Hevea brasiliensis. DNA Res. 2017, 24, 159–167. [Google Scholar]
- Uthup, T.K.; Saha, T.; Ravindran, M.; Bini, K. Impact of an intragenic retrotransposon on the structural integrity and evolution of a major isoprenoid biosynthesis pathway gene in Hevea brasiliensis. Plant Physiol. Biochem. 2013, 73, 176–188. [Google Scholar] [CrossRef]
- Epping, J.; Deenen, N.V.; Niephaus, E.; Stolze, A.; Fricke, J.; Huber, C.; Eisenreich, W.; Twyman, R.M.; Prufer, D.; Gronover, C.S. A rubber transferase activator is necessary for natural rubber biosynthesis in dandelion. Nat. Plants 2015, 1, 15048. [Google Scholar] [CrossRef]
- Berthelot, K.; Lecomte, S.; Estevez, Y.; Peruch, F. Hevea brasiliensis REF (Hev b 1) and SRPP (Hev b 3): An overview on rubber particle proteins. Biochimie 2014, 106, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gidda, S.K.; Park, S.; Pyc, M.; Yurchenko, O.; Cai, Y.; Wu, P.; Andrews, D.W.; Chapman, K.D.; Dyer, J.M.; Mullen, R.T. Lipid drop let associated proteins (LDAPs) are required for the dynamic regulation of neutral lipid compartmentation in plant cells. Plant Physiol. 2016, 170, 2052–2071. [Google Scholar] [CrossRef]
- Kim, E.Y.; Park, K.Y.; Seo, Y.S.; Kim, W.T. Arabidopsis Small rubber particle protein homolog SRPs play dual roles as positive factors for tissue growth and development and in drought stress responses. Plant Physiol. 2016, 4, 2494–2510. [Google Scholar] [CrossRef]
- Post, J.; van Deenen, N.; Fricke, J.; Kowalski, N.; Wurbs, D.; Schaller, H.; Eisenreich, W.; Huber, C.; Twyman, R.M.; Prüfer, D.; et al. Laticifer-specific cis-prenyltransferase silencing affects the rubber, triterpene, and inulin content of Taraxacum brevicorniculatum. Plant Physiol. 2012, 158, 1406–1417. [Google Scholar] [CrossRef]
- Hillebrand, A.; Post, J.J.; Wurbs, D.; Wahler, D.; Lenders, M.; Krzyzanek, V.; Prufer, D.; Gronover, C.S. Down-regulation of small rubber particle protein expression affects integrity of rubber particles and rubber content in Taraxacum brevicorniculatum. PLoS ONE 2012, 7, e41874. [Google Scholar] [CrossRef]
- Tata, S.K.; Choi, J.Y.; Jung, J.Y.; Lim, K.Y.; Shin, J.S.; Ryu, S.B. Laticifer tissue-specific activation of the Hevea SRPP promoter in Taraxacum brevicorniculatum and its regulation by light, tapping and cold stress. Ind. Crop. Prod. 2012, 40, 219–224. [Google Scholar] [CrossRef]
- Wang, X.C.; Shi, M.; Wang, D.; Chen, Y.; Cai, F.; Zhang, S.; Wang, L.; Tong, Z.; Tian, W.M. Comparative proteomics of primary and secondary lutoids reveals that chitinase and glucanase play a crucial combined role in rubber particle aggregation in Hevea brasiliensis. J. Proteome Res. 2013, 12, 5146–5159. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Wu, J. Laticifer differentiation in Hevea brasiliensis induction by exogenous jasmonic acid and linolenic acid. Ann. Bot. 2000, 85, 37–43. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; García-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Q.; Ding, G.; Zhu, L.; Yu, L.; Yuan, B.; Gao, X.; Wang, D.; Sun, Y.; Liu, Y.; Li, H.; et al. Proteomic Landscape of the Mature Roots in a Rubber-Producing Grass Taraxacum Kok-saghyz. Int. J. Mol. Sci. 2019, 20, 2596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102596
Xie Q, Ding G, Zhu L, Yu L, Yuan B, Gao X, Wang D, Sun Y, Liu Y, Li H, et al. Proteomic Landscape of the Mature Roots in a Rubber-Producing Grass Taraxacum Kok-saghyz. International Journal of Molecular Sciences. 2019; 20(10):2596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102596
Chicago/Turabian StyleXie, Quanliang, Guohua Ding, Liping Zhu, Li Yu, Boxuan Yuan, Xuan Gao, Dan Wang, Yong Sun, Yang Liu, Hongbin Li, and et al. 2019. "Proteomic Landscape of the Mature Roots in a Rubber-Producing Grass Taraxacum Kok-saghyz" International Journal of Molecular Sciences 20, no. 10: 2596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20102596