Molecular Cloning and Sexually Dimorphic Expression Analysis of nanos2 in the Sea Urchin, Mesocentrotus nudus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
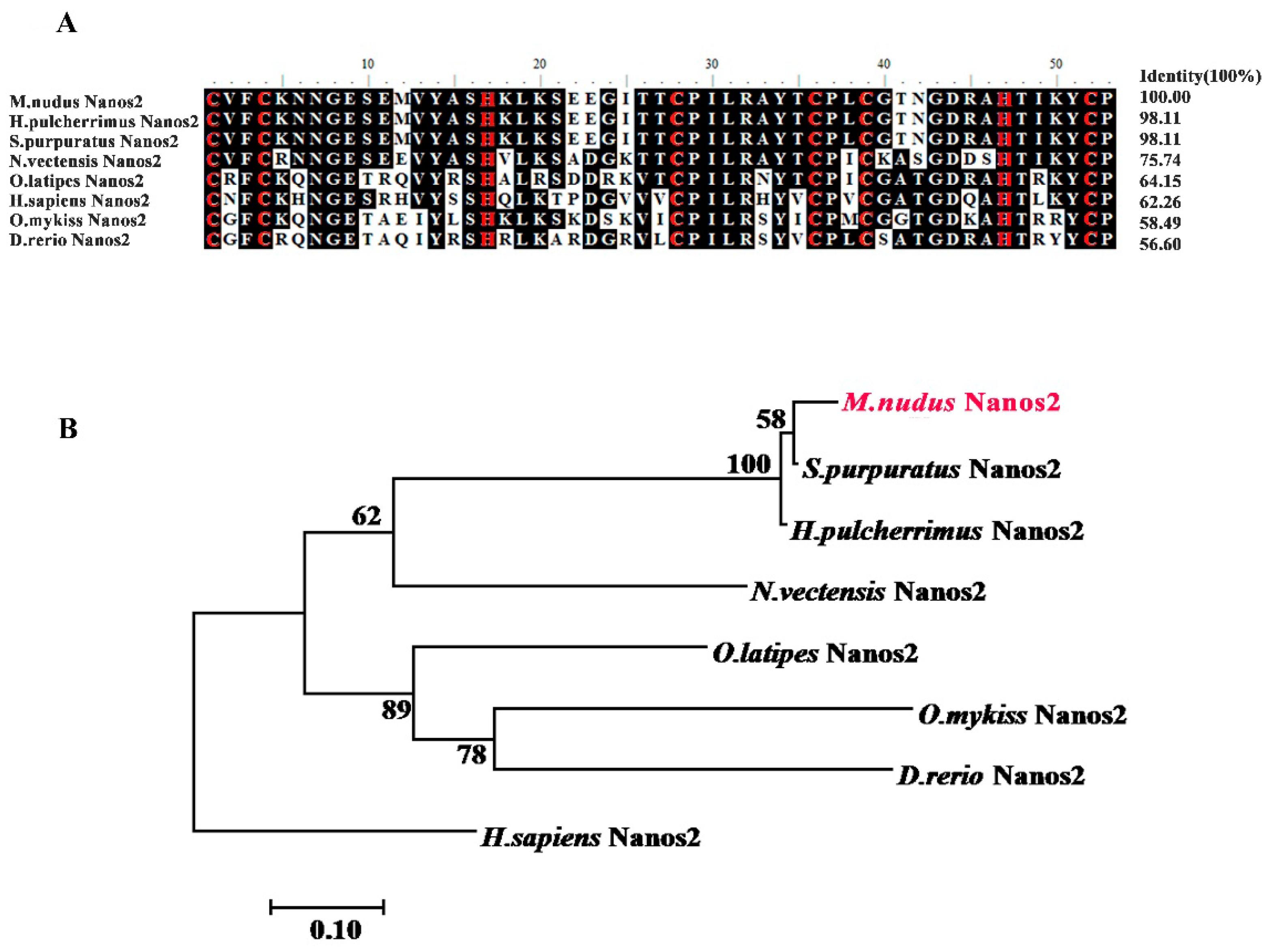
2.1. Isolation and Characterization of M. nudus nanos2
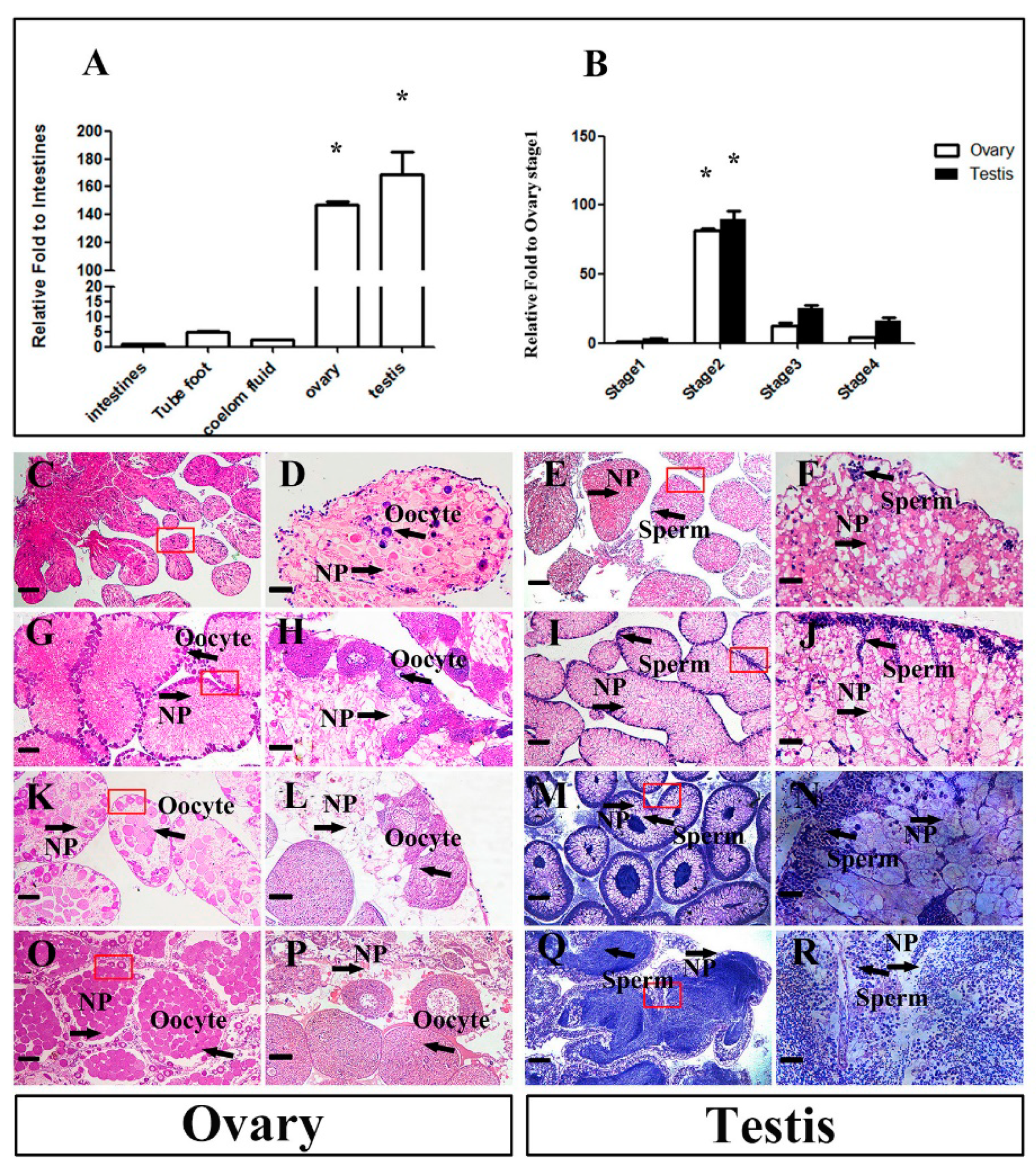
2.2. Mnnanos2 mRNA Expression Pattern in Different Adult Tissues and Dynamic Changes during Sex Development
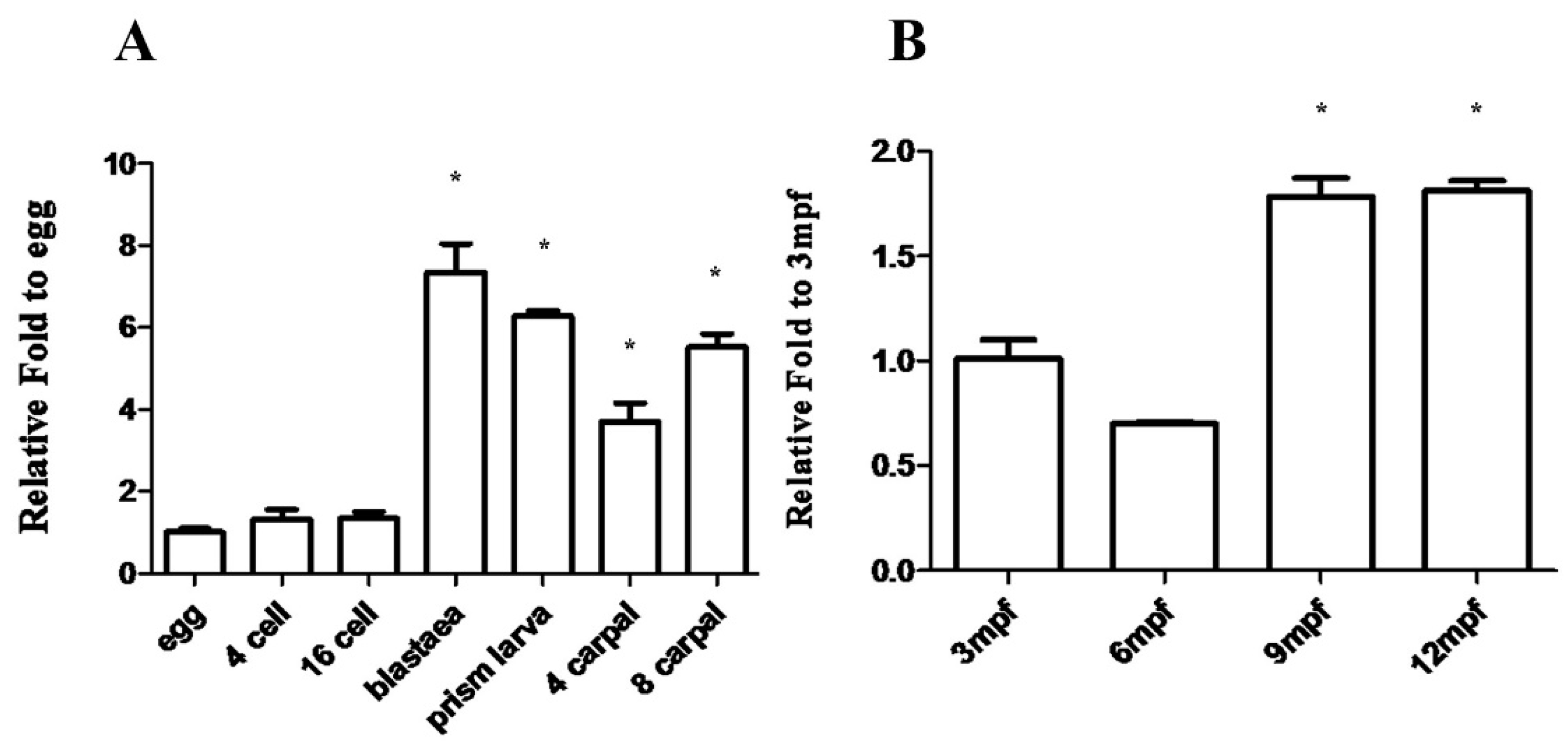
2.3. Mnnanos2 mRNA Expression Pattern during Embryogenesis and Early Ontogeny
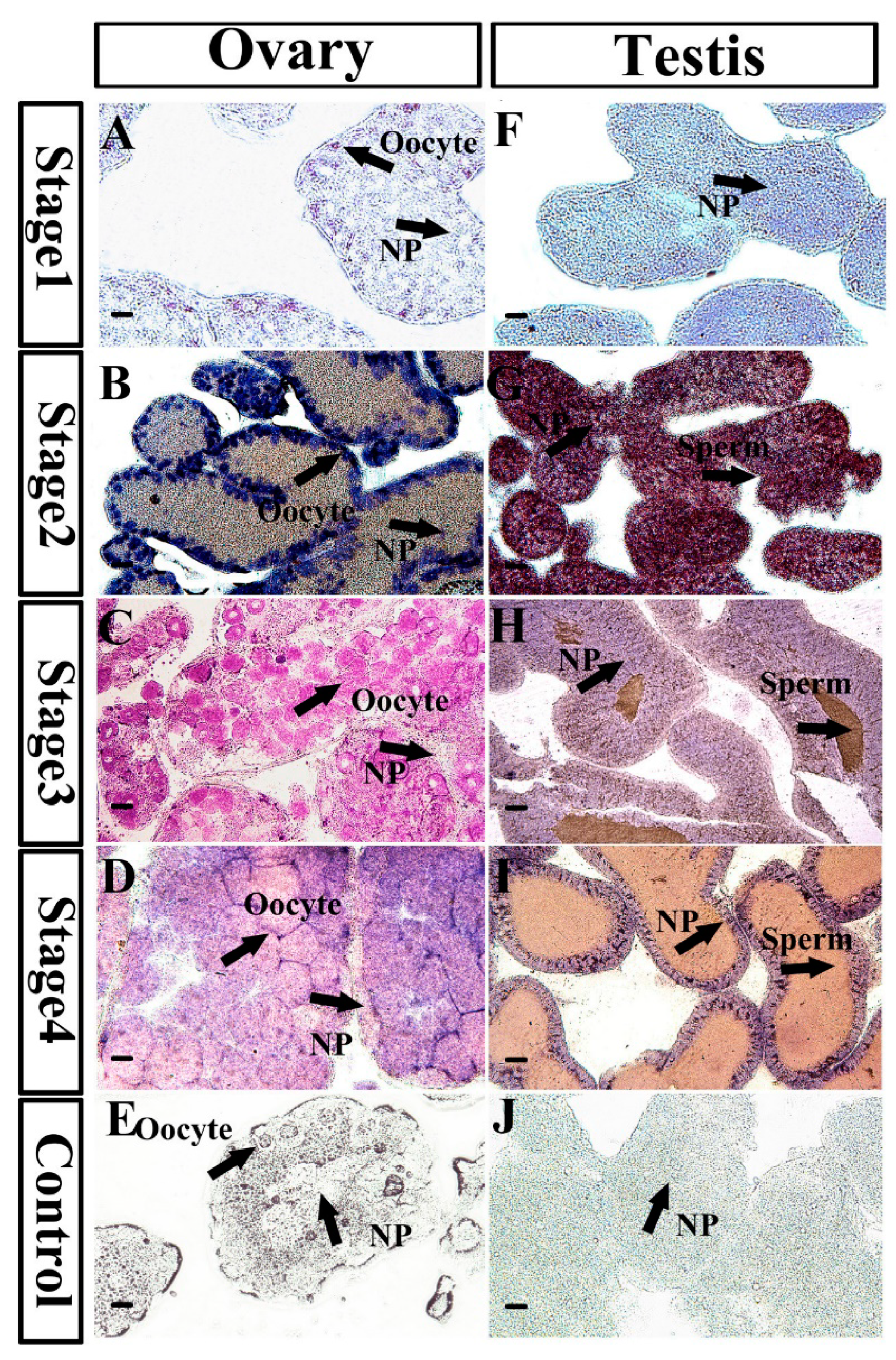
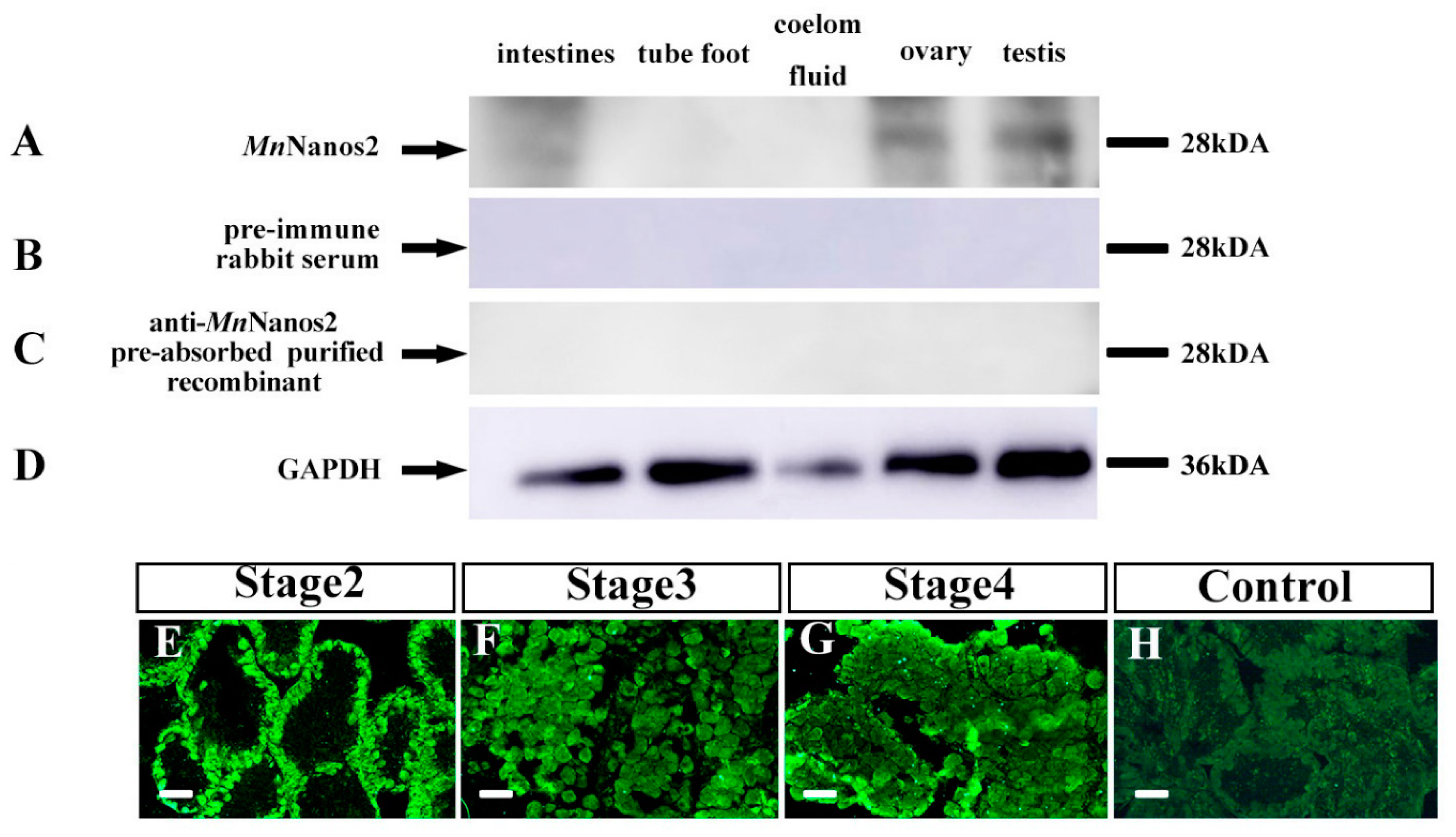
2.4. Characterization of MnNanos2 Protein Expression Pattern in Gonads
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Gonadal Histology
4.3. RNA Extraction and Real-Time Quantitative PCR (RT-qPCR)
4.4. Sequence and Phylogenetic Analyses
4.5. In Situ Hybridization
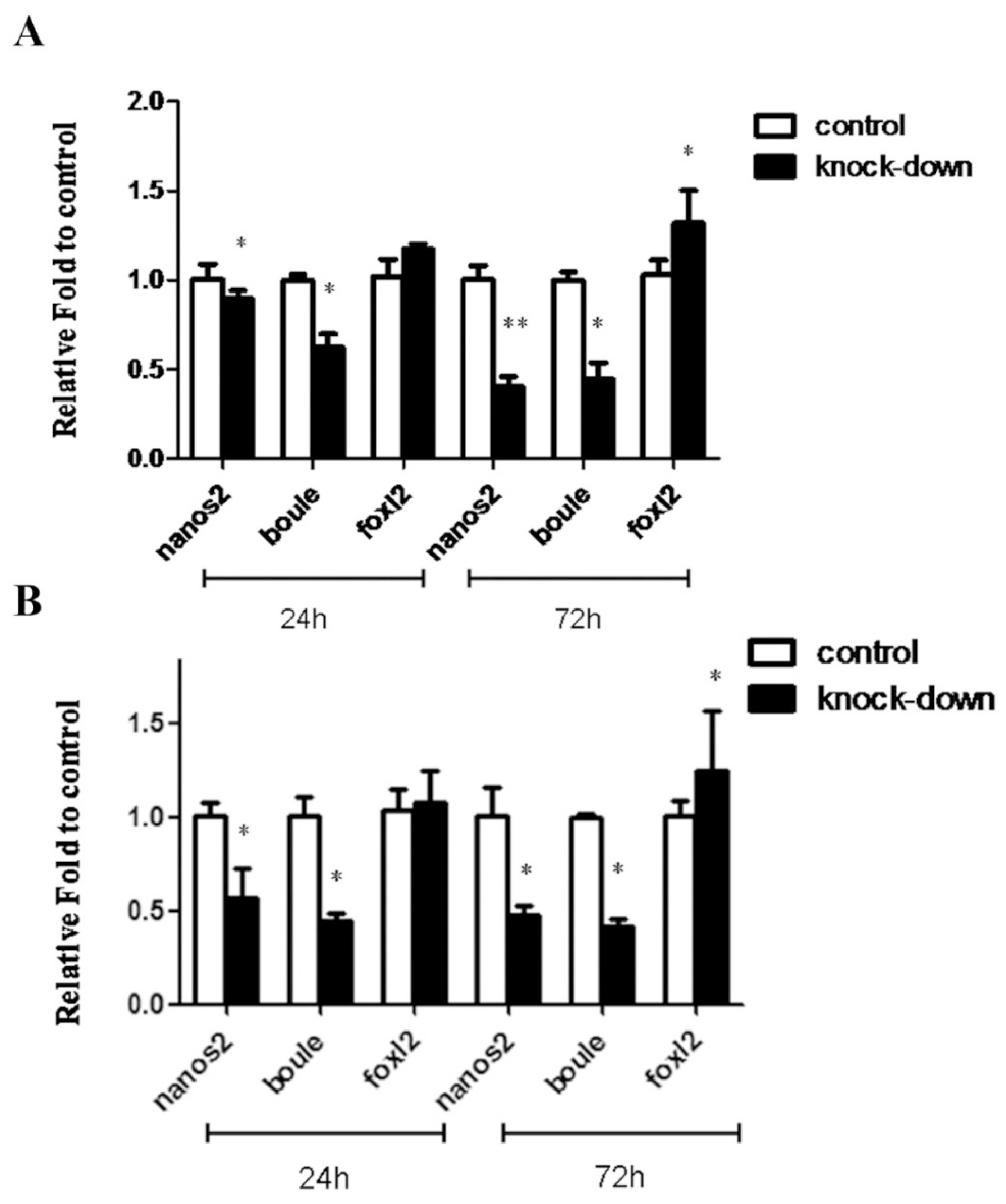
4.6. dsRNA Synthesis and Injection
4.7. Western Blot Analysis
4.8. Immunofluorescence Localization
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wang, C.; Lehmann, R. Nanos is the localized posterior determinant in Drosophila. J. Cell 1991, 66, 637–647. [Google Scholar] [CrossRef]
- Gribouval, L.; Sourdaine, P.; Lareyre, J.J.; Bellaiche, J.; Le Gac, F.; Mazan, S.; Guiardiere, C.; Auvray, P.; Gautier, A. The nanos1 gene was duplicated in early Vertebrates and the two paralogs show different gonadal expression profiles in a shark. Sci. Rep. 2018, 8, 6942. [Google Scholar] [CrossRef]
- Han, K.; Chen, S.; Cai, M.; Jiang, Y.; Zhang, Z.; Wang, Y. Nanos3 not nanos1 and nanos2 is a germ cell marker gene in large yellow croaker during embryogenesis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2018, 218, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Q.; Luo, A.L.; Zhao, P.W.; Li, T.T.; Geng, S.S.; Liang, X.W.; Xu, H.Y.; Lu, Y.Q.; Lu, S.S.; Yang, X.G.; et al. Nanos2 is a molecular marker of inchoate buffalo spermatogonia. Anim. Reprod. Sci. 2017, 186, 44–51. [Google Scholar] [CrossRef]
- Sun, Z.-H.; Wang, Y.; Lu, W.-J.; Li, Z.; Liu, X.-C.; Li, S.-S.; Zhou, L.; Gui, J.-F. Divergent expression patterns and function implications of four nanos genes in a hermaphroditic fish, epinephelus coioides. Int. J. Mol. Sci. 2017, 18, 685. [Google Scholar] [CrossRef]
- De Keuckelaere, E.; Hulpiau, P.; Saeys, Y.; Berx, G.; van Roy, F. Nanos genes and their role in development and beyond. Cell. Mol. Life Sci. 2018, 75, 1929–1946. [Google Scholar] [CrossRef]
- Asaoka-Taguchi, M.; Yamada, M.; Nakamura, A.; Hanyu, K.; Kobayashi, S. Maternal Pumilio acts together with Nanos in germline development in Drosophila embryos. Nat. Cell Biol. 1999, 1, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, M.; Sasaoka, Y.; Kiso, M.; Abe, K.; Haraguchi, S.; Kobayashi, S.; Saga, Y. Conserved Role of nanos Proteins in Germ Cell Development. Science 2003, 301, 1239–1241. [Google Scholar] [CrossRef]
- Beer, R.L.; Draper, B.W. Nanos3 maintains germline stem cells and expression of the conserved germline stem cell gene Nanos2 in the zebrafish ovary. Dev. Biol. 2013, 374, 308–318. [Google Scholar] [CrossRef]
- Yu, M.; Mu, H.; Niu, Z.; Chu, Z.; Zhu, H.; Hua, J. miR-34c enhances mouse spermatogonial stem cells differentiation by targeting Nanos2. J. Cell. Biochem. 2013, 115, 232–242. [Google Scholar] [CrossRef]
- Zhou, Z.; Shirakawa, T.; Ohbo, K.; Sada, A.; Wu, Q.; Hasegawa, K.; Saba, R.; Saga, Y. RNA Binding Protein Nanos2 Organizes Post-transcriptional Buffering System to Retain Primitive State of Mouse Spermatogonial Stem. J. Cells Dev. Cell 2015, 34, 96–107. [Google Scholar] [CrossRef]
- Zhang, W.; Bi, Y.; Wang, Y.; Li, D.; He, N.; Wang, M.; Jin, J.; Zuo, Q.; Zhang, Y.; Li, B. Nanos2 promotes differentiation of chicken (Gallus gallus) embryonic stem cells to male germ cells. J. Cell. Biochem. 2018, 119, 4435–4446. [Google Scholar] [CrossRef]
- Bellaiche, J.; Lareyre, J.J.; Cauty, C.; Yano, A.; Allemand, I.; Le Gac, F. Spermatogonial stem cell quest: nanos2, marker of a subpopulation of undifferentiated A spermatogonia in trout testis. Biol. Reprod. 2014, 90, 79. [Google Scholar] [CrossRef]
- Fresques, T.; Zazueta-Novoa, V.; Reich, A.; Wessel, G.M. Selective accumulation of germ-line associated gene products in early development of the sea star and distinct differences from germ-line development in the sea urchin. Dev. Dyn. 2014, 243, 568–587. [Google Scholar] [CrossRef] [PubMed]
- Juliano, C.E.; Yajima, M.; Wessel, G.M. Nanos functions to maintain the fate of the small micromere lineage in the sea urchin embryo. Dev. Biol. 2010, 337, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Oulhen, N.; Wessel, G.M. Differential Nanos 2 protein stability results in selective germ cell accumulation in the sea urchin. Dev. Biol. 2016, 418, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Liu, X.; Feng, T.; Chang, Y. Novel and conserved micrornas in Dalian purple urchin (Strongylocentrotus nudus) identified by next generation sequencing. Int. J. Biol. Sci. 2011, 7, 180. [Google Scholar] [CrossRef]
- Phillips, K.; Bremer, P.; Silcock, P.; Hamid, N.; Delahunty, C.; Barker, M.; Kissick, J. Effect of gender, diet and storage time on the physical properties and sensory quality of sea urchin (Evechinus chloroticus) gonads. Aquaculture 2009, 288, 205–215. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, J.; Zhang, W.; Chang, Y. Gonadal transcriptomic analysis and identification of candidate sex-related genes in Mesocentrotus nudus. Gene 2019, 698, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Wang, Q.; Wu, K.; Wei, Z.; Zhou, Z.; Liu, X. De novo transcriptome sequencing and comparative analysis to discover genes involved in ovarian maturity in Strongylocentrotus nudus. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 23, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Jiang, Y.; Shang, Y.; Zhang, Y.; Geng, C.; Wang, Q.; Chang, Y.Q. Establishment of lysozyme gene RNA interference systemand its involvement in salinity tolerance in sea cucumber (Apostichopus japonicus). Fish Shellfish Immunol. 2017, 65, 71–79. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Z.; Liang, S.; Yang, Q.; Wang, Y.; Qin, Z. A novel role of Kruppel-like factor 4 in Zhikong scallop Chlamys farreri during spermatogenesis. PLoS ONE 2017, 12, e0180351. [Google Scholar] [CrossRef]
- Yu, Y.; Ma, W.; Zeng, Q.; Qian, Y.; Yang, J.; Yang, W. Molecular Cloning and Sexually Dimorphic Expression of Two Dmrt Genes in the Giant Freshwater Prawn, Macrobrachium rosenbergii. Agric. Res. 2014, 3, 181–191. [Google Scholar] [CrossRef]
- Walker, C.W.; Tatsuya, U.; Michael, P.L. Gametogenesis and reproduction of sea urchins. Dev. Aquac. Fish. Sci. 2007, 37, 11–33. [Google Scholar]
- Walker, R.A. Quantification of immunohistochemistry--issues concerning methods, utility and semiquantitative assessment. Histopathology 2006, 49, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Dwarakanath, M.; Lim, M.; Xu, H.; Hong, Y. Differential expression of boule and dazl in adult germ cells of the Asian seabass. Gene 2014, 549, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, Y.; Li, Z.; Zhou, L.; Gui, J. Sequential, Divergent, and Cooperative Requirements of Foxl2a and Foxl2b in Ovary Development and Maintenance of Zebrafish. Genetics 2017, 205, 1551–1572. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Kobayashi, K.; Imura, T.; Higashijima, S.; Tanaka, M. Identification of Germline Stem Cells in the Ovary of the Teleost Medaka. Science 2010, 328, 1561–1563. [Google Scholar] [CrossRef]
- Saba, R.; Kato, Y.; Saga, Y. Nanos2 promotes male germ cell development independent of meiosis suppression. Dev. Biol. 2014, 385, 32–40. [Google Scholar] [CrossRef]
- Seydoux, K. Nos-1 and Nos-2, two genes related to Drosophila nanos, regulate primordial germ cell development and survival in Caenorhabditis elegans. Development 1999, 126, 4861–4871. [Google Scholar]
- D’Agostino, I.; Merritt, C.; Chen, P.L.; Seydoux, G.; Subramaniam, K. Translational repression restricts expression of the C. elegans Nanos homolog NOS-2 to the embryonic germline. Dev. Biol. 2006, 292, 244–252. [Google Scholar]
- Saga, Y. Function of Nanos2 in the male germ cell lineage in mice. Cell. Mol. Life Sci. 2010, 67, 3815–3822. [Google Scholar] [CrossRef] [PubMed]
- Oulhen, N.; Yoshida, T.; Yajima, M.; Song, J.L.; Sakuma, T.; Sakamoto, N.; Yamamoto, T.; Wessel, G.M. The 3′ UTR of nanos2 directs enrichment in the germ cell lineage of the sea urchin. Dev. Biol. 2013, 377, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.Y.; Su, Y.H. Genome editing in sea urchin embryos by using a CRISPR/Cas9 system. Dev. Biol. 2016, 409, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Xie, J.; Zhou, L.; Hong, Y.; Gui, J. Evolutionary conservation of Dazl genomic organization and its continuous and dynamic distribution throughout germline development in gynogenetic gibel carp. J. Exp. Zool. B Mol. Dev. Evol. 2019, 312, 855–871. [Google Scholar]
- Teaniniuraitemoana, V.; Huvet, A.; Levy, P.; Klopp, C.; Lhuillier, E.; Gaertner-Mazouni, N.; Gueguen, Y.; Le Moullac, G. Gonad transcriptome analysis of pearl oyster Pinctada margaritifera: Identification of potential sex differentiation and sex determining genes. BMC Genomics 2014, 15, 491. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liu, P.; Jia, F.; Li, J.; Gao, B. De novo Transcriptome Analysis of Portunus trituberculatus Ovary and Testis by RNA-Seq: Identification of Genes Involved in Gonadal Development. PLoS ONE 2015, 10, e0128659. [Google Scholar] [CrossRef]
- Schmidt, D.; Ovitt, C.E.; Anlag, K.; Fehsenfeld, S.; Gredsted, L.; Treier, A.C.; Treier, M. The murine winged-helixtranscription factor Foxl2 is required for granulosa celldifferentiation and ovary maintenance. Development 2004, 131, 933–942. [Google Scholar] [CrossRef]
- Li, Q.; Xie, J.; He, L.; Wang, Y.; Yang, H.; Duan, Z.; Wang, Q. FOXL2 down-regulates vitellogenin expression at mature stage in Eriocheir sinensis. Biosci. Rep. 2015, 35, e00278. [Google Scholar] [CrossRef] [Green Version]
- Govoroun, M.S.; Pannetier, M.; Pailhoux, E.; Cocquet, J.; Brillard, J.P.; Couty, I.; Batellier, F.; Cotinot, C. Isolation of chicken homolog of the FOXL2 gene and comparisonof its expression patterns with those of aromatase during ovarian development. Dev. Dyn. 2004, 231, 859–870. [Google Scholar] [CrossRef]
- Konstantin, V.; Yakovlev, N.R.; Battulin, O.L.; Serov, N.A.; Odintsova, N.A. Isolation of oogonia from ovaries of the sea urchin Strongylocentrotus Nudus. Cell Tissue Res. 2010, 342, 479–490. [Google Scholar]
- Wang, Y.; Sun, Z.; Zhou, L.; Li, Z.; Gui, J. Grouper Tshβ promoter-driven transgenic zebrafish marks proximal kidney tubule development. PLoS ONE 2014, 9, e97806. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Han, X.; Wang, J.; Liu, B.-Z.; Wei, J.-L.; Zhang, W.-J.; Sun, Z.-H.; Chang, Y.-Q. Molecular Cloning and Sexually Dimorphic Expression Analysis of nanos2 in the Sea Urchin, Mesocentrotus nudus. Int. J. Mol. Sci. 2019, 20, 2705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112705
Zhang J, Han X, Wang J, Liu B-Z, Wei J-L, Zhang W-J, Sun Z-H, Chang Y-Q. Molecular Cloning and Sexually Dimorphic Expression Analysis of nanos2 in the Sea Urchin, Mesocentrotus nudus. International Journal of Molecular Sciences. 2019; 20(11):2705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112705
Chicago/Turabian StyleZhang, Jian, Xiao Han, Jin Wang, Bing-Zheng Liu, Jin-Liang Wei, Wei-Jie Zhang, Zhi-Hui Sun, and Ya-Qing Chang. 2019. "Molecular Cloning and Sexually Dimorphic Expression Analysis of nanos2 in the Sea Urchin, Mesocentrotus nudus" International Journal of Molecular Sciences 20, no. 11: 2705. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20112705