Translation Control of HAC1 by Regulation of Splicing in Saccharomyces cerevisiae


Department of Biology, University of Ottawa, Marie-Curie Private, Ottawa, ON K1N 9A7, Canada
Int. J. Mol. Sci. 2019, 20(12), 2860; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122860
Submission received: 22 April 2019
/
Revised: 30 May 2019
/
Accepted: 10 June 2019
/
Published: 12 June 2019
(This article belongs to the Special Issue Translational Control)
Abstract
:Hac1p is a key transcription factor regulating the unfolded protein response (UPR) induced by abnormal accumulation of unfolded/misfolded proteins in the endoplasmic reticulum (ER) in Saccharomyces cerevisiae. The accumulation of unfolded/misfolded proteins is sensed by protein Ire1p, which then undergoes trans-autophosphorylation and oligomerization into discrete foci on the ER membrane. HAC1 pre-mRNA, which is exported to the cytoplasm but is blocked from translation by its intron sequence looping back to its 5’UTR to form base-pair interaction, is transported to the Ire1p foci to be spliced, guided by a cis-acting bipartite element at its 3’UTR (3’BE). Spliced HAC1 mRNA can be efficiently translated. The resulting Hac1p enters the nucleus and activates, together with coactivators, a large number of genes encoding proteins such as protein chaperones to restore and maintain ER homeostasis and secretary protein quality control. This review details the translation regulation of Hac1p production, mediated by the nonconventional splicing, in the broad context of translation control and summarizes the evolution and diversification of the UPR signaling pathway among fungal, metazoan and plant lineages.
Keywords:
HAC1; IRE1; ERN1; XBP1; unfolded protein response; UPR; non-spliceosome splicing; ER stress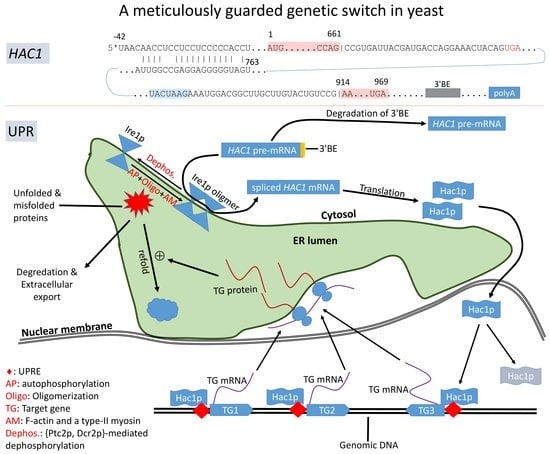
1. Introduction
Translation control occurs at different levels, with global and specific regulation as two extremes of the continuum. Global translation regulation impacts many genes. For example, dietary restriction in eukaryotes typically leads to a reduction of the ATP level and upregulation of translation repressor 4E-BP that binds to eukaryotic translation initiation factor 4E. This results in an overall reduction of cap-dependent translation. However, certain messages, e.g., translation of mitochondrion-targeted proteins, such as those in Complex I and IV of the electron transport chain, is increased [1]. In Saccharomyces cerevisiae, nutrient depletion also leads to a general reduction of cap-dependent translation, but a special set of genes that encourage invasive growth of yeast cells exhibits increased translation to take advantage of nutrients below the surface of the culture medium [2]. Another case of general translation control involves sulfur limitation, which reduces sulfur-containing amino acids such as methionine, which is needed for initiating translation [3]. Yeast cells respond to sulfur-limitation by using fewer proteins rich in sulfur-containing amino acid residues and replace the function of sulfur-rich proteins by sulfur-poor isozymes when possible [3].
Specific translation regulation involves a specific gene. Such specific regulation may be achieved through the action of trans-acting proteins [4] or microRNA [5], mediated by cis-acting nucleotide motifs. Autoregulation of several proteins represents prime examples of specific regulation. Eukaryotic genes encoding the poly(A)-binding protein (PABP) are known to have a poly(A) tract at 5’UTR, and PABP in high abundance will bind this poly(A) tract at 5’UTR to block translation by preventing the small ribosomal subunit from scanning to the initiation codon [6,7]. However, a poly(A) tract in 5’UTR that is shorter than the minimum binding size for PABP can recruit translation initiation factors and promote translation initiation [8]. This short negative feedback loop ensures optimal level of PABP production. Another example of translation autoregulation involves release factor 2 (RF2, encoded by prfB and decoding UGA and UAA stop codons) in many bacterial species [9,10]. The mRNA contains an inframe UGA stop codon followed by nucleotide C. UGAC represents a weak stop signal and is strongly avoided in highly expressed bacterial genes [11,12,13]. In Escherichia cli, when RF2 is abundant, this inframe UGA in prfB mRNA is decoded, leading to a truncated non-functional protein of only 25 amino acid residues. When RF2 is rare, this UGA is often not decoded, leading to a +1 frameshift by the ribosome and the production of a long functional RF2 of 366 residues. Thus, prfB mRNA is blocked from translation when RF2 is abundant, but allowed to be translated when RF2 is rare.
Translation regulation of HAC1 mRNA in yeast (Saccharomyces cerevisiae) belongs to specific translation regulation. It is special because the regulation is achieved through splicing which occurs only during ER (endoplasmic reticulum) stress sensed by transmembrane kinase/endonuclease Ire1p. This review details the regulation of HAC1 translation in the yeast, Saccharomyces cerevisiae, in the unfolded protein response (UPR) and outlines the diversity of the control mechanism in other evolutionary lineages.
2. HAC1 and Its Translation Control via Ire1p-Mediated Splicing
HAC1/YFL031W [14] is located on chromosome VI of the yeast, and encodes a transcription factor of 238 amino acids that plays a key role in transmitting the signal of unfolded/misfolded protein response (UPR) from ER to the nucleus [15,16,17,18], regulating downstream UPR genes such as KAR2, PDI1, EUG1, FKB2, and LHS1 that share a conserved UPRE motif [19]. Its pre-mRNA contains two exons and an intron of 252 nt (Figure 1). The intron has a peculiar 5’ss (G|CCGUGAU, where “|” marks the exon|intron boundary) instead of the consensus of G|GUAUGU in yeast. It ends with a CG dinucleotide (Figure 1) instead of the conventional AG dinucleotide. The intron is not processed by spliceosome, but is first cleaved by the protein kinase/endonuclease Ire1p [15,16], and then ligated by tRNA ligase Trl1p [20,21,22,23,24,25].
2.1. Intron-Mediated Translation Control of HAC1 mRNA
HAC1 is constitutively transcribed [15,16,28], and its spliced and unspliced forms are termed HAC1i and HAC1u, respectively, with the superscripted “i” and ”u” for induced and uninduced [17,29]. Unfortunately, this terminology is not universally adopted, and the spliced and unspliced forms of Xbp1 mRNA are represented as Xbp1s and Xbp1u, respectively, with the subscripted “s” and “u” for spliced and unspliced, respectively. HAC1u is exported to cytoplasm, contrary to most genes whose mRNAs are exported to the cytoplasm for translation only after splicing.
While HAC1u and HAC1i are equally stable [17], the presence of the intron in HAC1u inhibits its translation [16,17,30,31,32], whereas HAC1i is efficiently translated to generate functional Hac1ip. If HAC1u does get translated through experimental manipulation, e.g., by mutating the nucleotides involved in base-pairing in Figure 1 [33], the translation would stop at the stop codon highlighted in red in Figure 1. This results in a shorter protein (Hac1up) missing the activation domain encoded in the second exon. Hac1up exhibits much reduced transactivation activity on UPRE [28] than Hac1ip when assessed by using β-gal as a reporter gene [17,28,34].
The inhibitory effect of the HAC1 intron on translation is hypothesized and experimentally verified [33,35] to be through long-range base-pairing between intron and 5’UTR [26,32,33], as schematically depicted in Figure 1. This base-pair interaction would interfere with the cap-dependent scanning by the small ribosomal subunit to find the start codon. This interpretation of the base-pair interaction depicted in Figure 1, suggesting inhibition at translation initiation, which is consistent with recent experimental evidence [33,35,36], but not with earlier hypotheses of inhibition at translation elongation [17,32]. These early studies claimed that HAC1u is associated with polysomes but its translation fails to complete because of ribosome stalling [17,32]. There are three lines of support for inhibition of translation initiation of HAC1u instead of ribosome stalling during translation elongation [35]. First, mutating sites involved in base-pair interactions increases HAC1u associated with polysomes [33]. Second, adding an inframe AUG at the 5’ end of the base-pair interaction releases translation block. Third, increasing RNA helicase eIF4A expression, which likely would facilitate removal of the base-pair interaction, reduces translation block. Three additional experimental studies [33,36,37] demonstrated that efficient ribosomal loading occurs only on HAC1i, but not HAC1u, consistent with the interpretation that translation block occurs during the initiation stage. In non-UPR cells where HAC1 mRNA should almost all be in unspliced form, there are few HAC1 mRNA in polysomes [33,38]. Di Santo et al. [33] provided direct experimental evidence to demonstrate that the polysome-like sedimentation of HAC1u is not due to ribosomes translating HAC1u, but is due to non-specific association between HAC1u and other actively translated mRNAs.
2.2. Ire1p-Mediated Splicing of HAC1 mRNA
As suggested in Figure 1, removing the HAC1 intron would eliminate the translation block. The HAC1 intron is not spliced by the regular spliceosome pathway, but is cleaved by the protein kinase/endonuclease Ire1p [15,16], first discovered in budding yeast cells experiencing accumulation of unfolded proteins [39,40]. The cleavage does not exhibit the regular order of step 1 reaction of cleaving the 5’ end of intron followed by the step 2 reaction of cleaving the 3’ end intron. Instead, the 5’ and 3’ ends are cleaved independently in random order [27]. The two exons are then ligated by tRNA ligase Trl1p [20,21,22,23,24,25]. This splicing not only removes the inhibitory effect of the intron on translation, but also replaces the last 10 amino acids (aa) encoded in HAC1u by the 18 aa encoded in the second exon. The last 18 aa function as a transcriptional activation domain [28] so HAC1ip features much higher transactivation activity than HAC1up [17,28,34].
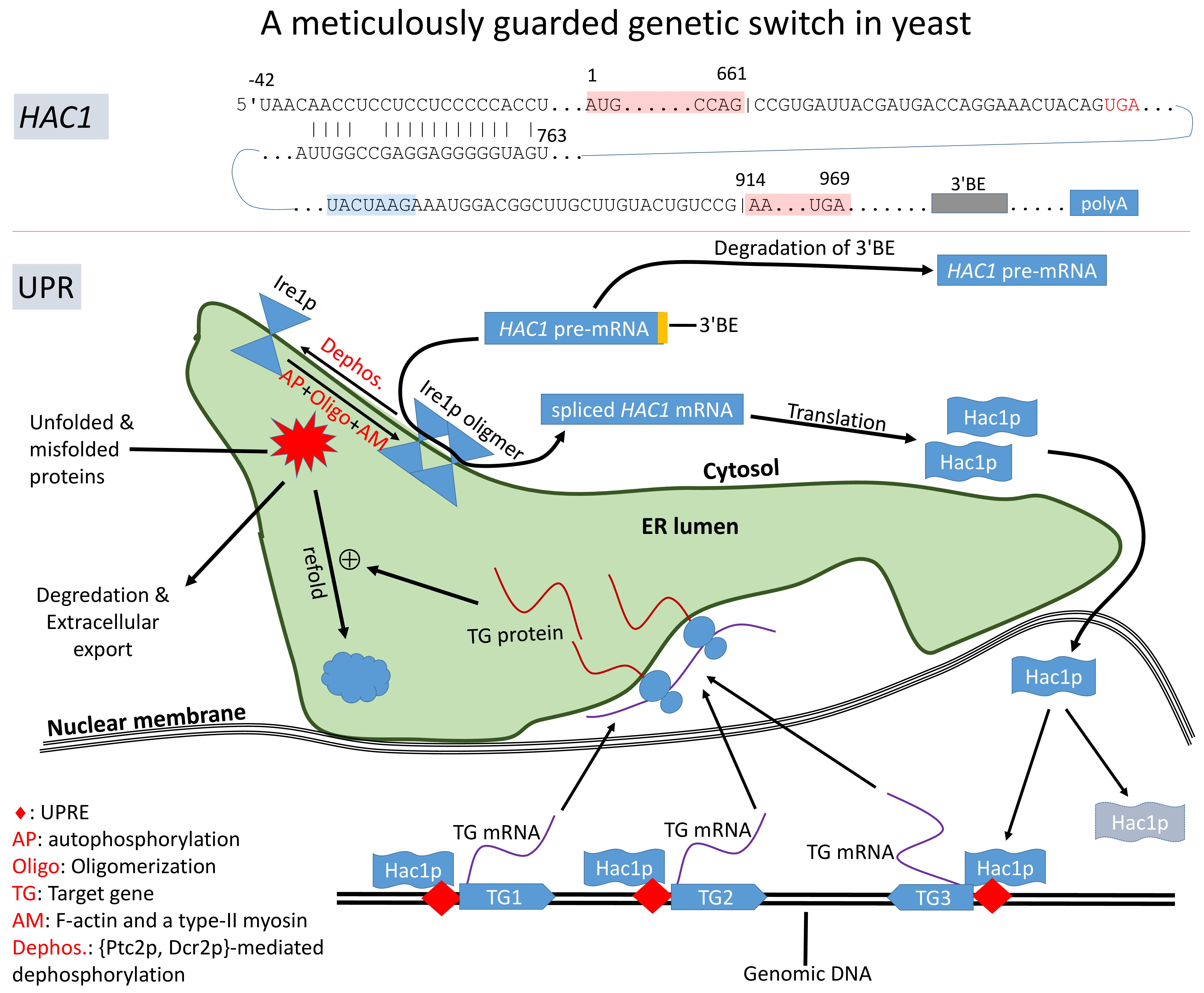
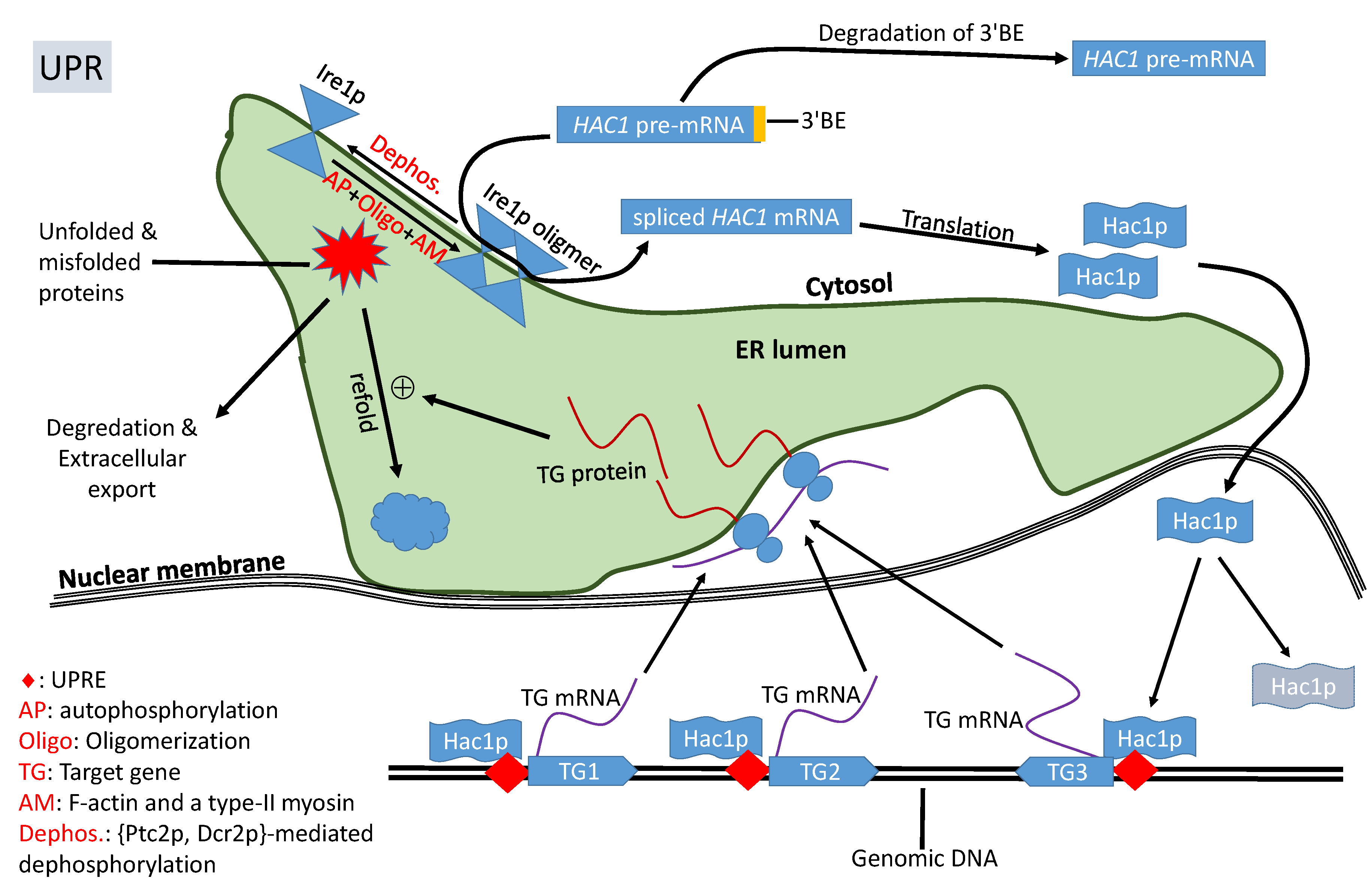
In response to accumulation of unfolded/misfolded proteins, the transmembrane Ire1p undergoes trans-autophosphorylation and congregates, in the presence of F-actin and a type-II myosin [41], into discrete foci of Ire1 oligomers within ER (Figure 2). The oligomerization is necessary for Ire1p-mediated splicing because the RNase activity of Ire1p is highly cooperative, so HAC1u cleavage is efficient only after Ire1p oligomerization [42,43]. It is at these Ire1 foci that HAC1 pre-mRNA are spliced [26,42,43,44,45]. HAC1 mRNA contains a 3’ cis-acting bipartite elements (3’BE, Figure 1) that signals to have HAC1 mRNA carried to Ire1p foci for splicing. This 3’BE functions in a modular way. Inserting this 3’BE and a translation block into another RNA is sufficient to direct the recipient RNA into Ire1p foci [26].
The 3’BE serves not only as a signal for HAC1 pre-mRNA to be carried to Ire1 foci for splicing, but also as a fail-safe mechanism to reduce accidental splicing of HAC1 pre-mRNA causing unneeded UPR. When the cell is not in ER stress, HAC1u undergoes rapid 3’ to 5’ degradation to remove its 3’BE [46] so that the resulting HAC1u will no longer be carried to the Ire1p foci for intron removal (Figure 2). UPR-inducing agents decreases this 3’BE degradation [46] so HAC1u can be transported to Ire1p foci for splicing. This fail-safe mechanism should increase the reliability of the signaling pathway to avoid accidental splicing of HAC1u to trigger unneeded UPR.
2.3. Hac1p Triggers UPR and Is Autoregulated
Spliced HAC1i, generated in response to ER stress, is translated efficiently to Hac1ip [24,25,36,37]. Being a transcription factor, Hac1p contains a nucleus localization sequence (RKRAKTK located at sites 29-35, shared by both forms of Hac1p) [47] which directs Hac1p into the nucleus. Hac1p binds to unfolded protein response elements (UPRE, Figure 2) of its regulated target genes such as those encoding protein chaperones [48,49]. Because a number of UPR-related chaperone genes do not share the same UPRE, and may not be activated directly by Hac1p, and coactivators such as Gcn4p have been proposed [50,51]. One particular coactivator complex involved in Hac1p-mediated UPR is Gcn5p/Ada which increases histone acetylation at chromatin encoding UPR-related genes [24,25]. Negatively charged DNA is typically wrapped tightly around positively charged histone complex, hiding promotor sequences and transcription factor binding sites. Histone acetylation reduces the basic charge in lysine residues in histone so that DNA will not be tightly wrapped around histones to facilitate transcription.
Both Hac1up and Hac1ip are rapidly degraded [17,34], with their half-life being about 1.5–2.0 min [17,34,47]. The short half-life of Hac1p indicates the importance or necessity of continuous production of Hac1p to sustain UPR. In this context, it is significant that the HAC1 gene is autoregulated, harboring a UPRE at its own promoter region [52]. This generates positive feedback to produce Hac1p to sustain UPR, i.e., the more Hac1p, the more active transcription of HAC1 and more Hac1p. This positive feedback is broken with dephosphorylation of Ire1p that reduces the Ire1p-mediated splicing of HAC1u [44,53] and rapid degradation of Hac1p.
3. Leaky Splicing of HAC1 mRNA in Non-UPR Yeast Cells
Leaky splicing of HAC1u in non-ER-stressed cells is undesirable because it could lead to the production and accumulation of Hac1ip and accidental induction of UPR. Two mechanisms help to avoid leaky splicing. First, weak signaling by residual Hac1ip may be handled by Kar2p which is a chaperone protein. When Kar2p is free from the task of protein-folding, it binds to Ire1p to hinder Ire1p clustering and oligomerization so that the endonuclease activity of the RNase domain of Ire1p is not activated [54,55,56]. This works because the RNase activity of Ire1p is highly cooperative, so HAC1u cleavage is efficient only after Ire1p oligomerization [42,43]. Thus, a residual amount of Hac1ip would upregulated Kar2p production which would 1) reduce unfolded proteins, and 2) reduce Ire1-mediated splicing by hindering Ire1p clustering and oligomerization. Second, even if dimers or oligomers of Ire1p occasionally form, the 3’BE of HAC1u is efficiently removed by 3’ to 5’ degradation [46] to reduce the traffic of HAC1u towards Ire1p oligomers.
In spite of the two fail-safe mechanisms mentioned above, leaky splicing of HAC1u may still occur. It is therefore interesting to characterize splicing efficiency of HAC1u in non-ER-stressed yeast cells. Empirical measurement of splicing efficiency (SE) of introns is based on the quantification of spliced (featuring exon-exon junctions) and unspliced (featuring exon-intron junctions) forms of individual genes or individual introns. An early attempt to characterize SE is by microarray, with exon-exon junction (EE) and exon-intron junctions at 5’ and 3’ sides of an intron (EI5 and EI3) as microarray probes to quantify spliced and unspliced form [57]. A simple but accurate method of quantifying SE by using RNA-seq data has been developed and applied to yeast introns [58]. The rationale is the same as the microarray approach. One obtains 1) the number of transcriptomic reads mapped to EE junctions (NEE) to quantify the spliced form, and 2) the number of reads mapped to EI5 and EI3 junctions of the intron (NEI5 and NEI3) to quantify the unspliced form. NEI5 is typically smaller than NEI3 because 1) step 1 splicing reaction occurs before step 2 splicing reaction so cleavage of EI5 junction occurs before EI3 junction, 2) enrichment of poly(A) mRNA by oligo-dT may lead to DNA replication reaching beyond EI3 but not EI5, and 3) 5’ degradation. Thus, the total mRNA (NTotal) is measured not as NTotal = NEE + (NEI5 + NEI3)/2, but as NTotal = NEE + p × NEI5 + (1-p) × NEI3, with p estimated from data [58]. SE is defined as SE = NEE/NTotal.
Most yeast genes, especially intron-containing ribosomal protein genes, are spliced efficiently (Table 1) with most mRNA in spliced form (large NEE values) and less in unspliced form (small NEI5 and NEI3 values). However, HAC1 mRNA is poorly spliced, with a small NEE value of 32 and large NEI5 and NEI3 values (844 and 704, respectively, last row in Table 1). The result also confirms that 1) HAC1 is transcribed constitutively in yeast [15,16], present in a significant amount in yeast cells not in UPR, and 2) HAC1 splicing does occur in non-UPR cells so that rapid degradation of Hac1p [17,33,34,47] is necessary as a fail-safe mechanism against accidental triggering of UPR.
There is indeed weak splicing of HAC1 in non-UPR yeast cells, with about 2% of HAC1 in spliced form in non-UPR cells. UPR induction increases the percentage to ~69% in yeast cells. Thus, given the leaking splicing of HAC1 mRNA, the few HAC1 mRNAs observed in polysomes in non-UPR cells [33,38] might in fact be in spliced form.
The leaky splicing implies the production of Hac1ip and the risk of inducing unwanted UPR in non-stressed cells. Yeast has two fail-safe mechanisms against accidental induction of UPR. First, Kar2p, known to raise the threshold of UPR induction by participating in protein-folding and by hindering Ire1 from oligomerization, is directly and strongly upregulated by Hac1ip and would consequently suppress the weak signaling of UPR by a small amount of Hac1ip [54,55,56,60,61,62]. The second and perhaps far more important mechanism is the rapid degradation of Hac1ip. The degradation of Hac1ip occurs by the ubiquitin-proteasome pathway [33,47] and requires a nuclear localization sequence (29)RKRAKTK(35) encoded in the first exon [47]. If the degradation signal is encoded in the first exon, then it would explain why Hac1up and Hac1ip are both highly unstable in the yeast [17,34], with the half-life being only about 1.5-2.0 min [17,34,47]. However, this turns out to be an incomplete story as is detailed in the next section.
4. Leaky Translation of HAC1u
Leaky translation of HAC1u was considered minimal given the well-documented intron-mediated translation inhibition (Figure 1), and the observation that Hac1up is undetectable in non-ER-stressed yeast cells. This, together with the observation that Hac1up is a much weaker transcription factor than Hac1ip when assessed by using β-gal as a reporter gene [17,28,34], does not suggest a risk of accidental UPR induced by Hac1up. Thus, there seems to be no need for the budding yeast to evolve any protein-degradation mechanism specifically targeting Hac1up. However, this reasoning turned out to be inadequate.
While leaky translation of Hac1u is indeed rare, there is substantial evidence that Hac1up could potentially accumulate without efficient degradation [33]. Thus, rapid degradation is needed in non-stressed cells, not only for Hac1ip produced through leaky splicing and subsequently translation, but also for Hac1up produced through leaky translation of Hac1u. In this context, a degron was discovered in the 10-aa encoded in the 5’ end of HAC1u intron [33]. In particular, this degron is recognized by Duh1p/Das1p, leading to a short half-life of Hac1up of less than a minute [33].
It was previously thought that the degradation signal is encoded in the first exon, and therefore shared with both Hac1ip and Hac1up, leading to both being equally highly unstable with half-life being 2 min or less [17,34,47]. However, if this were true, then there would be no need to have a degron in the last 10-aa of Hac1up. It is therefore possible that the degradation signal in Hac1ip and Hac1up are independently encoded, with a degron in the last 10-aa of Hac1up [33] and a separate degron in the last 18-aa of Hac1ip. One may check if the degron in Hac1up is conserved among different yeast lineages.
5. Ire1p Domain Structure and Its Splicing Activity
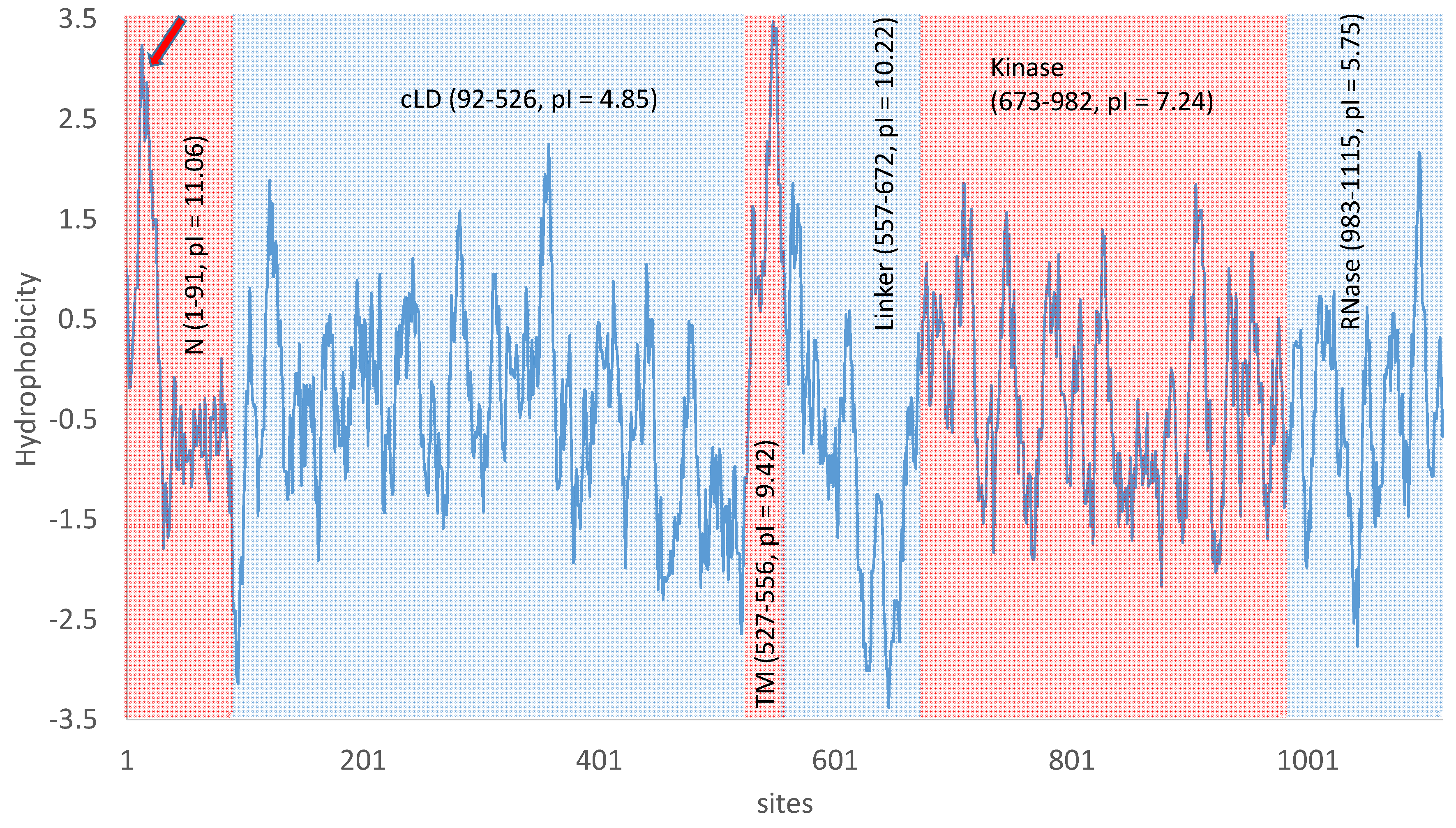
The luminal part of the transmembrane Ire1p can be roughly divided into two components [43,63,64]: the N-terminal domain with unsolved structure and the core luminal domain (N and cLD, respectively, in Figure 3). The N-terminal domain contains more positively charged residues than negatively charged ones (e.g., eight Arg and four Lys, but only three Asp and three Glu), leading to a high isoelectric point (pI) and positive charge under normal cytoplasmic pH range of 5.5–7.5 in yeast cells. However, the N-terminal domain also contains a consecutive stretch of amino acids (sites 8 to 30) that are highly hydrophobic (pointed by a red arrow in Figure 3). This hydrophobic stretch would need to be either buried deep into the folded protein or insert into a membrane. Given that the N-terminal appears structurally unresolvable [63], it does not seem likely that the hydrophobic stretch is buried deep inside a protein structure. Thus, the hydrophobic stretch might represent another transmembrane domain which would make Ire1p a double-pass instead of a single-pass transmembrane protein. However, this is a “backdoor” inference, i.e., one does not have direct evidence that the hydrophobic stretch is a transmembrane domain but makes the inference because an alternative is unlikely.
Multiple lines of empirical evidence supports the hypothesis that unfolded/misfolded proteins serve as direct binding ligands to the luminal domain of Ire1p to trigger Ire1p trans-autophosphorylation and oligomerization in both unicellular yeast [65] and multicellular eukaryotes [66]. The core luminal domain of the transmembrane Ire1p (cLD in Figure 3) is mainly responsible for oligomerization of Ire1p and formation of Ire1p foci to facilitate HAC1 splicing [26,42,43,44,45] because Ire1p monomers have little splicing activity [42]. Note that, at a cytosolic pH of 7, a protein domain with a pI of 4.85 would be negatively charged, so cLD would tend to repulse each other against oligomerization. This may help avoid accidental oligomerization to prevent unwanted HAC1 splicing and UPR, but would hinder Ire1p clustering at neutral pH. A cytosolic pH equal to 4.85 would reduce the cLD charge to 0, facilitating clustering and oligomerization. This suggests that UPR may be triggered more readily in an acidic environment. Acidosis does cause ER stress and induce UPR in human vascular endothelial cells [67]. S. cerevisiae can grow in a wide range of ambient pH, but the cytosolic pH is maintained in the range of about 5.5-7.5 [68,69]. Everything else being equal, one expects UPR to be induced more readily at low pH than at pH ≥ 7.
One negative regulator of the IRE1p-HAC1p pathway is the luminal Kar2p [72] which binds to Ire1p under non-ER-stress conditions but dissociates from Ire1p under ER stress [60,61,62]. Kar2p is not essential for UPR induction but may serve to desensitize Ire1p to avoid unnecessary UPR [54,55,56]. The theoretical pI of Kar2p is only 4.62, and is therefore even more negatively charged than the cLD domain of Ire1p at neutral pH. This seems to create a challenge for Kar2p to bind to the cLD domain of Ire1p. However, while the cLD domain of Ire1 is overall negatively charged under neutral cytosolic pH, it has a stretch of positively charged amino acids, (91)RRANKKGRR(99), which may electrostatically interact favorably with the negatively charged Kar2p. Alternatively, the negative Kar2p could either bind to the positively charged N-terminal domain (Figure 3), or bind to cLM when the cellular cytoplasmic pH is near the pI of cLM and Kar2p so that they will not carry net negative charge. Given that KAR2 is strongly upregulated by Hac1p [73] and that overexpression of Kar2p attenuates UPR [74], one may infer an alternative function of Kar2p as helping the cell exit UPR once ER homeostasis is restored. That the three regulatory motifs in KAR2 that regulate KAR2 expression independently [72] suggests additional functions of KAR2.
The cytosolic components of Ire1p include a linker domain that facilitates the docking of HAC1u to Ire1p, a kinase domain, and an RNase domain that cleaves the HAC1u intron [42]. The RNase activity is highly cooperative, so the cleavage of HAC1u is more efficient after oligomerization [42,43]. This series of Ire1p oligomerization, HAC1u docking and splicing should reduce the chance of accidental splicing of HAC1u by Ire1p [42].
The 30-aa transmembrane domain of Ire1p is highly hydrophobic (TM, Figure 3) and structurally unresolved as is typical of transmembrane protein domains. Its relatively high pI is mainly due to two residues that tend to be positively charged (one Arg and one Lys) and only one Glu that tends to be negatively charged. Its last 16 aa carry neither positive nor negative charges.
The cytoplasmic linker domain (Figure 3) has a high theoretical pI (= 10.27) and is therefore strongly positively charged at neutral pH. This positive charge would facilitate the binding of the linker domain to negatively charged backbone of HAC1 substrate. It is indeed crucial for the docking of HAC1 pre-mRNA to the linker domain before cleavage by the RNase domain of Ire1p [42]. The linker domain also appears necessary for oligomerization in vitro [43].
The kinase domain of Ire1p features a stretch of negatively charged amino acids (624)DNDDADEDDE(633) almost immediately followed by a stretch of positively charged amino acids (641)KKKRKRGSRGGKKGRKSR(659). However, this segment of Ire1p is missing in the many structural studies on Ire1p. The kinase domain contains several phosphorylation sites (S840, S841, T844, and S850), so the empirical pI is expected to be lower than the theoretical pI of 7.24 (Figure 3). These phosphorylation sites serve as a genetic switch in the UPR circuit. When S840, S841 and T844 are mutated to Asp (which would be equivalent to permanent phosphorylation in terms of electric charge), yeast cells are unable to exit UPR after its induction [75].
6. How Is Ire1p Activity Regulated in Response to Unfolded/Misfolded Proteins in Yeast?
Many factors contribute to protein unfolding and misfolding. These include overexpression of an exogenous protein, e.g., a mouse protein in yeast cells [76], disruption of O-mannosylation in yeast [77], imbalance of ion concentrations such as calcium [78] and cadmium [79], excess of acetic acid, propionic acid and sorbic acid [80], antifungal agents that perturb protein folding such as monoterpene carvacrol [81,82], tunicamycin [18], dithiothreitol [83,84,85], hypoxia [86], or acidosis [67].
Ire1p senses ER stress caused by the accumulation of unfolded/misfolded proteins, and helps transmitting the signal of UPR from ER to nucleus by splicing of HAC1 pre-mRNA (Figure 2). Ire1p activity of HAC1 splicing can potentially be regulated through three mechanisms: (1) transcription, (2) translation, and (3) posttranslational modification. Although transcription and translation do not directly affect enzymatic activity of Ire1p, these two processes will increase Ire1p abundance on ER membrane and consequently higher probability of colliding with each other to oligomerize and activate the HAC1-splicing function. While transcriptomic characterization before and after UPR induction has recently been carried out for S. cerevisiae [73] and Candida parapsilosis [84], most studies focused on regulating Ire1p’s splicing activity by posttranslational modification, especially on autophosphorylation/dephosphorylation [87], which modulates HAC1-splicing activity of Ire1p and oligomerization, which is required for efficient Ire1p-mediated HAC1-spilcing [26,41,43,44,45].
There are multiple lines of evidence suggesting the HAC1-splicing activity of Ire1p is regulated by posttranslational modification. First, Ire1p-containing complexes do associate with RNAs in normal growing cells without UPR (UPR−), but these RNA species are longer than those found in Ire1p-containing complexes in cells under UPR (UPR+) [87]. This indicates (1) that splicing activity is absent in UPR− cells or at least weaker in UPR− Cells than in UPR+ cells and (2) that Ire1p-containing complexes have changed during the switch between UPR− and UPR+ states. Second, Ire1p has a cluster of phosphorylation sites: S840, S841, T844, S850, as well as a persistently negatively charged D836. Phosphorylation adds a negative charge to these sites and would potentially affect protein structure and its electrostatic interactions with its RNA substrates. Mutation at these sites in Ire1p, or replacing the negatively charged D836 by A836, reduces HAC1 pre-mRNA splicing [88], although Ire1p clustering is not affected [42]. Third, protein phosphatase Dcr2p in yeast targets Ire1p specifically, physically interact with Ire1p’s phosphorylation sites S840 and S841, dephosphorylate Ire1p in vitro and downregulate UPR [53]. Another protein phosphatase Ptc2p also dephosphorylates Ire1p and negatively regulates UPR [44].
Exactly how cells exit UPR is not well elucidated, but three complementary mechanisms may operate for the purpose. The first is dephosphorylation of Ire1p [75] by either Ptc2p [44] or Dcr2p [53] or both. The three phosphorylation sites in the kinase domain (Figure 3), S840, S841, and T844, are particularly important for exiting UPR in yeast [75]. Phosphomimetic mutations of these sites to Asp results in failure for yeast cells to exit UPR after UPR induction [75]. The second is rapid degradation of Hac1p (Figure 2) which has a half-life of only about 1.5-2.0 min [17,34,47]. Hac1up has a half-life even shorter because of the presence of a degron in its C-terminus [33]. The degradation of Hac1p is mediated by the ubiquitin-proteasome pathway [33,47] and requires a nuclear localization sequence (29)RKRAKTK(35) [47]. The third is the upregulation of Kar2p by Hac1p during UPR induction. Overexpressed Kar2p binds to Ire1p to attenuate UPR [74]. The time taken for yeast cells to exit UPR after its induction is in the order of hours [75].
7. Conservation and Diversification of Ire1p-Mediated UPR Signaling
There are three classes of potential cellular responses to the accumulation of unfolded/misfolded proteins: (1) reducing protein-folding demand in ER, (2) increasing protein-folding capacity in ER, and (3) apoptosis. Reducing protein-folding demand can be achieved by (1) selective mRNA degradation known as regulated Ire1p-dependent decay (RIDD) documented in both Sch. pombe [89,90] and mammalian cells [91,92], (2) reduced ribosomal proteins documented in yeast [93], (3) increased ER-associated degradation of unfolded/misfolded proteins known as ERAD [76,94,95], and (4) export unfolded/misfolded proteins to extracellular matrix [94]. Increasing protein-folding capacity in ER can be achieved by (1) the Ire1p-Hac1p signaling pathway to increase the production of chaperone proteins [48,49] or (2) direct processing and stabilization of mRNA encoding chaperone proteins, such as BIP1 mRNA in Schizosaccharomyces pombe [89]. These responses are often referred to as adaptive because they all contribute to restoring ER homeostasis. When all these fail to restore ER homeostasis after prolonged UPR, apoptosis is triggered [79,85,96,97,98].
These cellular responses are not independent of each other. For example, RIDD is involved in (1) restoring ER homeostasis when it degrades mRNA encoding ER-translocating proteins to reduce protein-folding demand, and (2) apoptosis when RIDD eventually begins to degrade anti-apoptosis miRNAs after prolonged UPR failing to restore ER homeostasis. Two characteristic events occur during the transition of RIDD from contributing to ER homeostasis to apoptosis. First, it degrades KAR2 mRNA and consequently increases ER stress [97]. Second, it cleaves anti-apoptosis microRNAs such as miR-17, miR-34a, miR-96 and miR-125b allowing apoptosis gene products to accumulate [97,99]. There might be a switch between adaptive response and apoptosis [79,85,96,97,98].
While Ire1p-mediated UPR contributes to all these responses, individual species may possess only a subset of responses. For example, RIDD is a dominant response in UPR to restore ER homeostasis in Sch. pombe which has Ire1p but no Hac1p homologue [89,90]. RIDD is also well developed in many multicellular species that do have Hac1p/Xbp1 homologues, including insects, mammals and plants [91,92,100]. However, RIDD is missing in S. cerevisiae as Hac1 mRNA is the only detected substrate of Ire1p cleavage [100]. Thus, while many species feature both RIDD and Ire1p-Hac1p/Xbp1 pathways, Sch. pombe has lost the Ire1p-Hac1p/Xbp1 pathway, and S. cerevisiae has lost the RIDD pathway.
7.1. Conservation and Diversification of the Ire1p-Hac1p Pathway
The key genes involved in UPR signaling in the yeast are IRE1 and HAC1, together with genes such as TRL1 that helps ligate cleaved HAC1 exons into a processed and efficiently translatable mRNA (Figure 2). The evolutionary conservation of this UPR signaling pathway is best exemplified by IRE1 which has homologues (e.g., ERN1 in mammals) in many studied eukaryotes [101,102] including mammals [103] with universally conserved function of participating in the unconventional splicing of pre-mRNA of HAC1 or its functional homologues such as Xbp1 [104,105] in metazoans and bZIP60 in plants [101,106].
The IRE1+HAC1 combination in yeast and Ire1+Xbp1 in metazoans work analogously. For example, in both Caenorhabditis elegans and mice, Xbp1 splicing occurs during UPR, with proteins from spliced Xbp1 mRNA accumulating during UPR, but not proteins from unprocessed Xbp1 mRNA [107]. There is, however, little sequence homology between yeast HAC1 and mammalian Xbp1, either at the nucleotide or amino acid level, except for a short stretch in the protein sequence that is rich in positively charged amino acids (arginine and lysine) corresponds to sites 29-53 in the yeast Hac1p (with sites 29-35 being the nuclear localization sequence of Hac1p). Surprisingly, both Hac1up and Hac1ip, when expressed in mammalian cells, can induce UPR [108].
Mammalian species have evolved PERK and ATF6 pathway as two UPR branches in addition to the Ire1p/Xbp1 pathway [109,110,111]. In contrast, Sch. pombe has lost the Hac1p homologue and consequently the Ire1p-Hac1p pathway [89,90]. Nature has offered different options to different evolutionary lineages to respond to ER stress. A detailed evolutionary reconstruction would shed light on the loss/gain of signal transduction pathways.
7.2. Conservation and Diversity of Ire1p-Mediated Splicing
HAC1 and Xbp1 mRNA form hairpins recognized by Ire1p, and this hairpin structure is conserved among diverse evolutionary lineages from yeast to mammals [112,113] so that HAC1 or Xbp1 mRNA can sometimes be properly spliced by Ire1p from other species. What is particularly remarkable is that Sch. pombe, which no longer has a Hac1p/Xbp1 homologue, can still splice an engineered intron based on the secondary structure template shared among diverse organisms [114]. This is somewhat unexpected because Ire1p in Sch. pombe participates actively in RIDD [89,90] and cleaves a variety of different RNA species, with no clearly defined cleavage sites. Thus, one would expect Ire1p in Sch. pombe to be promiscuous in RNA substrate selection. More studies are needed to understand how Sch. pombe Ire1p can specifically cleave the intron at specific sites yet seem to act less discriminately against other RNA species.
S. cerevisiae has only one tRNA ligase (Trl1p) which is also used to ligate cleaved HAC1 exons. In amphioxus, three proteins (RLN, Clp1, PNK/CPDase) are involved in tRNA ligation, but only RNL can ligate cleaved Xbp1 exons [115]. Similarly, among mammalian tRNA ligases, only RtcB is for ligating cleaved Xbp1 exons [116,117]. Arabidopsis thaliana tRNA ligase (AtRlg1p) can substitute for yeast Trl1p in tRNA splicing as well as ligate the cleaved HAC1 exons [118]. However, the resulting HAC1 mRNA is not translated efficiently because the HAC1 intron is circularized and likely still capable of forming the base-pair interaction with the 5’ UTR of HAC1 mRNA to block translation initiation.
The mammalian tRNA ligase RtcB can be knocked down or even eliminated without affecting cell survival [116,117]. This raises the possibility of regulating Xbp1 at the ligating stage. If Ire1p cleaves Xbp1u, but there is no RtcB to ligate the cleaved exon, then no mature Xbp1s would form. Is RtcB lowly expressed in non-ER-stressed cells but highly expressed under ER stress? Such a possible regulation mediated by tRNA ligase is not possible in the budding yeast. The yeast Trl1p, in contrast to mammalian RtcB, needs to be continually produced and functional for tRNA maturation, and its ligase activity consequently cannot be blocked. Another possibility that has not been examined is that cleavage sites of Ire1p in some species are degenerate but tRNA ligase picks the right exon-containing segment to ligate.
7.3. Diversity in Translation Control
The HAC1 intron is conserved in various fungal species only around the splice sites that contribute to the formation of the hairpin structure recognized by Ire1p for splicing [113], but not the segment forming base-pair interaction with 5’UTR (Figure 1). This means that the mechanism of blocking translation initiation in HAC1 is not shared beyond close relatives of the yeast. Introns in HAC1 homologues vary dramatically in intron length. For example, Candida parapsilosis has an HAC1 intron of 626 nt [84], whereas the intron is 29 nt long in Yarrowia lipolytica [119] and only 20 nt in Aspergillus niger [120]. What is particularly significant is that the short 20-nt intron in A. niger forms a stable stem-loop structure so it can no longer have base-pair interaction with the 5’UTR of hacA (homologue of HAC1 in A. niger). This implies that a new translation-blocking mechanism is needed in A. niger, and indeed there is one through differential transcription start site. When cells are not in UPR, the transcribed hacA has a long 5’UTR with a GC-rich inverted repeat (18 base-pairs) that would interfere with the cap-dependent ribosome scanning to find the initiation codon. When UPR is triggered, hacA transcripts with a short 5’UTR missing the left half of the inverted repeat are produced. This short hacA mRNA can be efficiently translated to activate UPR [120]. Translation regulation mediated by different transcription start sites is also observed in filamentous fungi A. nidulans and Trichoderma reesei [121]. These two species, upon UPR induction, generate HAC1 transcripts without a short upstream ORF. These HAC1 messages are more efficiently translated than those with the short uORF included. However, the unconventional splicing of a 20-nt intron in these two species also contributes to increased protein production, demonstrated by different gene constructs [121].
Several Candida-related fungal species do not have an intron in their HAC1 homologue, and would seem to require new translation-blocking mechanisms that have not yet been identified. Sch. pombe has Ire1p, but does not have a HAC1/Xbp1 orthologue. However, its UPR signaling pathway does involve IRE1 and BIP1 which encode a major ER-chaperone [89]. The dominant hypothesis for UPR in Sch. pombe [89,90] is that Sch. pombe has recently lost its Hac1p/Xbp1 homologue, and that its Ire1p 1) participates in RIDD to reduce protein production and consequently folding load within ER, and 2) cleaves and stabilizes BIP1 mRNA leading to increased production of Bip1p which, being a major ER-chaperone, helps to alleviate ER stress. Thus, ER chaperone proteins are increased (and UPR induced) in S. pombe without involving Hac1p serving as a transcription factor activating the transcription of genes encoding ER chaperones. The empirical support for a recent loss of Hac1p/Xbp1 homologue is that Sch. pombe Ire1p still retains the function of splicing an engineered intron with secondary structure similar to that in Xbp1 [114]. This secondary structure is highly conserved in vertebrate IRE1 intron [112]. However, it is also possible that another protein, very different in sequence from Hac1p, serves as a functional analogue of Hac1p in Sch. pombe.
Regulation of Hac1p in yeast is different from regulation of Xbp1 in metazoans. Unspliced Hac1u is hardly translated, but mammalian unspliced Xbp1 mRNA is constitutively translated but rapidly degraded [122]. The yeast’s way of regulating the abundance of Hac1p seems more economical than the mammalian way of limiting the abundance of Xbp1.
Given the high sequence divergence, it is not surprising that yeast HAC1 expressed in Hela and HEK293 human cell lines is generally not processed, but is translated efficiently [123]. However, other studies show that yeast HAC1 (yHAC1) is correctly spliced in mammalian cells upon UPR inductions [124]. Interestingly, when yHAC1 is expressed in mammalian cells, proteins from both spliced and unspliced yHAC1 are active in inducing the production of chaperone proteins [108].
8. Conclusions
(Ire1p+Hac1p)-mediated UPR signaling represents a beautiful translation control mechanism created by nature. However, there are still many questions unanswered. What recognizes 3’BE of HAC1 pre-mRNA and carries HAC1 pre-mRNA to Ire1p foci to be spliced? How is translation regulated in HAC1 homologues that do not have introns in fungal species? Is Ire1p oligomerization a consequence of Ire1p phosphorylation or the opposite? How does Ire1p dephosphorylation impact Ire1p oligomerization and HAC1 splicing activity? How do cells exit UPR after its induction? How does this signaling system evolve and diverge in different lineages? I hope that this review will serve as an anchor for addressing all these questions in coming years.
Funding
This research was funded by a Discovery Grant from the Natural Science and Engineering Research Council (NSERC, RGPIN/2018-03878) of Canada.
Acknowledgments
I thank J. Silke and Y. Wei for discussion and comments. Three reviewers provided excellent feedback.
Conflicts of Interest
I declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Zid, B.M.; Rogers, A.N.; Katewa, S.D.; Vargas, M.A.; Kolipinski, M.C.; Lu, T.A.; Benzer, S.; Kapahi, P. 4e-bp extends lifespan upon dietary restriction by enhancing mitochondrial activity in Drosophila. Cell 2009, 139, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W.V.; Zhou, K.; Butler, T.K.; Doudna, J.A. Cap-independent translation is required for starvation-induced differentiation in yeast. Science 2007, 317, 1224–1227. [Google Scholar] [CrossRef] [PubMed]
- Fauchon, M.; Lagniel, G.; Aude, J.C.; Lombardia, L.; Soularue, P.; Petat, C.; Marguerie, G.; Sentenac, A.; Werner, M.; Labarre, J. Sulfur sparing in the yeast proteome in response to sulfur demand. Mol. Cell 2002, 9, 713–723. [Google Scholar] [CrossRef]
- Gebauer, F.; Preiss, T.; Hentze, M.W. From cis-regulatory elements to complex RNPs and back. Cold Spring Harb. Perspect. Biol. 2012, 4, a012245. [Google Scholar] [CrossRef] [PubMed]
- Roux, P.P.; Topisirovic, I. Regulation of mRNA translation by signaling pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a012252. [Google Scholar] [CrossRef] [PubMed]
- De Melo Neto, O.P.; Standart, N.; Martins de Sa, C. Autoregulation of poly(a)-binding protein synthesis in vitro. Nucleic. Acids Res. 1995, 23, 2198–2205. [Google Scholar] [CrossRef]
- Wu, J.; Bag, J. Negative control of the poly(a)-binding protein mRNA translation is mediated by the adenine-rich region of its 5’-untranslated region. J. Biol. Chem. 1998, 273, 34535–34542. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; MacKay, V.; Yao, X.; Wu, J.; Miura, F.; Ito, T.; Morris, D.R. Translation initiation: A regulatory role for poly(a) tracts in front of the AUG codon in Saccharomyces cerevisiae. Genetics 2011, 189, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Craigen, W.J.; Caskey, C.T. Expression of peptide chain release factor 2 requires high-efficiency frameshift. Nature 1986, 322, 273–275. [Google Scholar] [CrossRef]
- Craigen, W.J.; Cook, R.G.; Tate, W.P.; Caskey, C.T. Bacterial peptide chain release factors: Conserved primary structure and possible frameshift regulation of release factor 2. Proc. Natl. Acad. Sci. USA 1985, 82, 3616–3620. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. Bioinformatics and translation termination in bacteria. In Bioinformatics and the Cell: Modern Computational Approaches in Genomics, Proteomics and Transcriptomics; Springer: Cham, Switzerland, 2018; pp. 239–254. [Google Scholar]
- Wei, Y.; Wang, J.; Xia, X. Coevolution between stop codon usage and release factors in bacterial species. Mol. Biol. Evol. 2016, 33, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Xia, X. The role of +4u as an extended translation termination signal in bacteria. Genetics 2017, 205, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Nojima, H.; Leem, S.H.; Araki, H.; Sakai, A.; Nakashima, N.; Kanaoka, Y.; Ono, Y. Hac1: A novel yeast bZIP protein binding to the CRE motif is a multicopy suppressor for cdc10 mutant of Schizosaccharomyces pombe. Nucleic. Acids Res. 1994, 22, 5279–5288. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Kawahara, T.; Yoshida, H.; Yanagi, H.; Yura, T. Signalling from endoplasmic reticulum to nucleus: Transcription factor with a basic-leucine zipper motif is required for the unfolded protein-response pathway. Genes Cells 1996, 1, 803–817. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.S.; Walter, P. A novel mechanism for regulating activity of a transcription factor that controls the unfolded protein response. Cell 1996, 87, 391–404. [Google Scholar] [CrossRef]
- Chapman, R.E.; Walter, P. Translational attenuation mediated by an mRNA intron. Curr. Biol. 1997, 7, 850–859. [Google Scholar] [CrossRef] [Green Version]
- Nikawa, J.; Akiyoshi, M.; Hirata, S.; Fukuda, T. Saccharomyces cerevisiae IRE2/HAC1 is involved in IRE1-mediated KAR2 expression. Nucleic. Acids Res. 1996, 24, 4222–4226. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Ogawa, N.; Kawahara, T.; Yanagi, H.; Yura, T. Palindrome with spacer of one nucleotide is characteristic of the cis-acting unfolded protein response element in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 9912–9920. [Google Scholar] [CrossRef] [PubMed]
- Sidrauski, C.; Cox, J.S.; Walter, P. tRNA ligase is required for regulated mRNA splicing in the unfolded protein response. Cell 1996, 87, 405–413. [Google Scholar] [CrossRef]
- Sidrauski, C.; Walter, P. The transmembrane kinase Ire1p is a site-specific endonuclease that initiates mRNA splicing in the unfolded protein response. Cell 1997, 90, 1031–1039. [Google Scholar] [CrossRef]
- Gonzalez, T.N.; Sidrauski, C.; Dorfler, S.; Walter, P. Mechanism of non-spliceosomal mRNA splicing in the unfolded protein response pathway. Embo. J. 1999, 18, 3119–3132. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J. Stress signaling from the lumen of the endoplasmic reticulum: Coordination of gene transcriptional and translational controls. Genes Dev. 1999, 13, 1211–1233. [Google Scholar] [CrossRef] [PubMed]
- Welihinda, A.A.; Tirasophon, W.; Kaufman, R.J. The cellular response to protein misfolding in the endoplasmic reticulum. Gene. Expr. 1999, 7, 293–300. [Google Scholar] [PubMed]
- Welihinda, A.A.; Tirasophon, W.; Kaufman, R.J. The transcriptional co-activator ada5 is required for HAC1 mRNA processing in vivo. J. Biol. Chem. 2000, 275, 3377–3381. [Google Scholar] [CrossRef]
- Aragon, T.; van Anken, E.; Pincus, D.; Serafimova, I.M.; Korennykh, A.V.; Rubio, C.A.; Walter, P. Messenger RNA targeting to endoplasmic reticulum stress signalling sites. Nature 2009, 457, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Yanagi, H.; Yura, T.; Mori, K. Unconventional splicing of HAC1/ern4 mRNA required for the unfolded protein response. Sequence-specific and non-sequential cleavage of the splice sites. J. Biol. Chem. 1998, 273, 1802–1807. [Google Scholar] [CrossRef]
- Mori, K.; Ogawa, N.; Kawahara, T.; Yanagi, H.; Yura, T. MRNA splicing-mediated c-terminal replacement of transcription factor Hac1p is required for efficient activation of the unfolded protein response. Proc. Natl. Acad. Sci. USA 2000, 97, 4660–4665. [Google Scholar] [CrossRef]
- Schroder, M.; Clark, R.; Kaufman, R.J. Ire1- and HAC1-independent transcriptional regulation in the unfolded protein response of yeast. Mol. Microbiol. 2003, 49, 591–606. [Google Scholar] [CrossRef]
- Sidrauski, C.; Chapman, R.; Walter, P. The unfolded protein response: An intracellular signalling pathway with many surprising features. Trends Cell Biol. 1998, 8, 245–249. [Google Scholar] [CrossRef]
- Anshu, A.; Mannan, M.A.; Chakraborty, A.; Chakrabarti, S.; Dey, M. A novel role for protein kinase kin2 in regulating HAC1 mRNA translocation, splicing, and translation. Mol. Cell Biol. 2015, 35, 199–210. [Google Scholar] [CrossRef]
- Ruegsegger, U.; Leber, J.H.; Walter, P. Block of HAC1 mRNA translation by long-range base pairing is released by cytoplasmic splicing upon induction of the unfolded protein response. Cell 2001, 107, 103–114. [Google Scholar] [CrossRef]
- Di Santo, R.; Aboulhouda, S.; Weinberg, D.E. The fail-safe mechanism of post-transcriptional silencing of unspliced HAC1 mRNA. eLife 2016, 5, e20069. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, T.; Yanagi, H.; Yura, T.; Mori, K. Endoplasmic reticulum stress-induced mRNA splicing permits synthesis of transcription factor Hac1p/ern4p that activates the unfolded protein response. Mol. Biol. Cell 1997, 8, 1845–1862. [Google Scholar] [CrossRef] [PubMed]
- Sathe, L.; Bolinger, C.; Mannan, M.A.; Dever, T.E.; Dey, M. Evidence that base-pairing interaction between intron and mRNA leader sequences inhibits initiation of HAC1 mRNA translation in yeast. J. Biol. Chem. 2015, 290, 21821–21832. [Google Scholar] [CrossRef] [PubMed]
- Payne, T.; Hanfrey, C.; Bishop, A.L.; Michael, A.J.; Avery, S.V.; Archer, D.B. Transcript-specific translational regulation in the unfolded protein response of Saccharomyces cerevisiae. FEBS Lett. 2008, 582, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, K.M.; DeRisi, J.L.; Brown, P.O.; Sarnow, P. Global and specific translational regulation in the genomic response of Saccharomyces cerevisiae to a rapid transfer from a fermentable to a nonfermentable carbon source. Mol. Cell Biol. 2001, 21, 916–927. [Google Scholar] [CrossRef]
- Arava, Y.; Wang, Y.; Storey, J.D.; Liu, C.L.; Brown, P.O.; Herschlag, D. Genome-wide analysis of mRNA translation profiles in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2003, 100, 3889–3894. [Google Scholar] [CrossRef]
- Cox, J.S.; Shamu, C.E.; Walter, P. Transcriptional induction of genes encoding endoplasmic reticulum resident proteins requires a transmembrane protein kinase. Cell 1993, 73, 1197–1206. [Google Scholar] [CrossRef]
- Mori, K.; Ma, W.; Gething, M.J.; Sambrook, J. A transmembrane protein with a cdc2+/cdc28-related kinase activity is required for signaling from the ER to the nucleus. Cell 1993, 74, 743–756. [Google Scholar]
- Ishiwata-Kimata, Y.; Yamamoto, Y.H.; Takizawa, K.; Kohno, K.; Kimata, Y. F-actin and a type-ii myosin are required for efficient clustering of the ER stress sensor IRE1. Cell Struct. Funct. 2013, 38, 135–143. [Google Scholar] [CrossRef]
- Van Anken, E.; Pincus, D.; Coyle, S.; Aragon, T.; Osman, C.; Lari, F.; Gomez Puerta, S.; Korennykh, A.V.; Walter, P. Specificity in endoplasmic reticulum-stress signaling in yeast entails a step-wise engagement of HAC1 mRNA to clusters of the stress sensor IRE1. eLife 2014, 3, e05031. [Google Scholar] [CrossRef] [PubMed]
- Korennykh, A.V.; Egea, P.F.; Korostelev, A.A.; Finer-Moore, J.; Zhang, C.; Shokat, K.M.; Stroud, R.M.; Walter, P. The unfolded protein response signals through high-order assembly of IRE1. Nature 2009, 457, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Welihinda, A.A.; Tirasophon, W.; Green, S.R.; Kaufman, R.J. Protein serine/threonine phosphatase ptc2p negatively regulates the unfolded-protein response by dephosphorylating Ire1p kinase. Mol. Cell Biol. 1998, 18, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Korennykh, A.V.; Korostelev, A.A.; Egea, P.F.; Finer-Moore, J.; Stroud, R.M.; Zhang, C.; Shokat, K.M.; Walter, P. Structural and functional basis for RNA cleavage by IRE1. BMC Biol. 2011, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Paira, S.; Das, B. Nuclear mRNA degradation tunes the gain of the unfolded protein response in Saccharomyces cerevisiae. Nucleic. Acids Res. 2018, 46, 1139–1156. [Google Scholar] [CrossRef]
- Pal, B.; Chan, N.C.; Helfenbaum, L.; Tan, K.; Tansey, W.P.; Gething, M.J. Scfcdc4-mediated degradation of the Hac1p transcription factor regulates the unfolded protein response in Saccharomyces cerevisiae. Mol. Biol. Cell 2007, 18, 426–440. [Google Scholar] [CrossRef]
- Mori, K. The unfolded protein response: The dawn of a new field. Proc. Jpn. Acad. Ser. B Physic. Biol. Sci. 2015, 91, 469–480. [Google Scholar] [CrossRef]
- Walter, P. Walking along the serendipitous path of discovery. Mol. Biol. Cell 2010, 21, 15–17. [Google Scholar] [CrossRef]
- Patil, C.K.; Li, H.; Walter, P. Gcn4p and novel upstream activating sequences regulate targets of the unfolded protein response. PLoS Biol. 2004, 2, E246. [Google Scholar] [CrossRef]
- Herzog, B.; Popova, B.; Jakobshagen, A.; Shahpasandzadeh, H.; Braus, G.H. Mutual cross talk between the regulators HAC1 of the unfolded protein response and gcn4 of the general amino acid control of Saccharomyces cerevisiae. Eukaryot. Cell. 2013, 12, 1142–1154. [Google Scholar] [CrossRef]
- Ogawa, N.; Mori, K. Autoregulation of the HAC1 gene is required for sustained activation of the yeast unfolded protein response. Genes Cells 2004, 9, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Polymenis, M. Dcr2 targets IRE1 and downregulates the unfolded protein response in Saccharomyces cerevisiae. EMBO Rep. 2006, 7, 1124–1127. [Google Scholar] [CrossRef] [PubMed]
- Kimata, Y.; Oikawa, D.; Shimizu, Y.; Ishiwata-Kimata, Y.; Kohno, K. A role for bip as an adjustor for the endoplasmic reticulum stress-sensing protein IRE1. J. Cell Biol. 2004, 167, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Pincus, D.; Chevalier, M.W.; Aragon, T.; van Anken, E.; Vidal, S.E.; El-Samad, H.; Walter, P. Bip binding to the ER-stress sensor IRE1 tunes the homeostatic behavior of the unfolded protein response. PLoS Biol. 2010, 8, e1000415. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, D.; Kimata, Y.; Kohno, K. Self-association and bip dissociation are not sufficient for activation of the ER stress sensor IRE1. J. Cell Sci. 2007, 120, 1681–1688. [Google Scholar] [CrossRef] [PubMed]
- Pleiss, J.A.; Whitworth, G.B.; Bergkessel, M.; Guthrie, C. Rapid, transcript-specific changes in splicing in response to environmental stress. Mol. Cell 2007, 27, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Xia, X. Rna-seq approach for accurate characterization of splicing efficiency of yeast introns. Methods 2019. [Google Scholar] [CrossRef]
- Xia, X. Arsda: A new approach for storing, transmitting and analyzing transcriptomic data. G3 Genes Genom. Genet. 2017, 7, 3839–3848. [Google Scholar] [CrossRef]
- Kimata, Y.; Kohno, K. Endoplasmic reticulum stress-sensing mechanisms in yeast and mammalian cells. Curr. Opin. Cell. Biol. 2011, 23, 135–142. [Google Scholar] [CrossRef]
- Bertolotti, A.; Zhang, Y.; Hendershot, L.M.; Harding, H.P.; Ron, D. Dynamic interaction of bip and ER stress transducers in the unfolded-protein response. Nat. Cell Biol. 2000, 2, 326–332. [Google Scholar] [CrossRef]
- Okamura, K.; Kimata, Y.; Higashio, H.; Tsuru, A.; Kohno, K. Dissociation of Kar2p/bip from an ER sensory molecule, Ire1p, triggers the unfolded protein response in yeast. Biochem. Biophys. Res. Commun. 2000, 279, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Credle, J.J.; Finer-Moore, J.S.; Papa, F.R.; Stroud, R.M.; Walter, P. On the mechanism of sensing unfolded protein in the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2005, 102, 18773–18784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, C.; Pincus, D.; Korennykh, A.; Schuck, S.; El-Samad, H.; Walter, P. Homeostatic adaptation to endoplasmic reticulum stress depends on IRE1 kinase activity. J. Cell Biol. 2011, 193, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Gardner, B.M.; Walter, P. Unfolded proteins are IRE1-activating ligands that directly induce the unfolded protein response. Science 2011, 333, 1891–1894. [Google Scholar] [CrossRef] [PubMed]
- Karagoz, G.E.; Acosta-Alvear, D.; Nguyen, H.T.; Lee, C.P.; Chu, F.; Walter, P. An unfolded protein-induced conformational switch activates mammalian IRE1. eLife 2017, 6, e30700. [Google Scholar] [CrossRef]
- Dong, L.; Krewson, E.A.; Yang, L.V. Acidosis activates endoplasmic reticulum stress pathways through gpr4 in human vascular endothelial cells. Int. J. Mol. Sci. 2017, 18, 278. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Nino, M.; Marquina, M.; Swinnen, S.; Rodriguez-Porrata, B.; Nevoigt, E.; Arino, J. The cytosolic ph of individual Saccharomyces cerevisiae cells is a key factor in acetic acid tolerance. Appl. Environ. Microbiol. 2015, 81, 7813–7821. [Google Scholar] [CrossRef] [PubMed]
- Valli, M.; Sauer, M.; Branduardi, P.; Borth, N.; Porro, D.; Mattanovich, D. Intracellular ph distribution in Saccharomyces cerevisiae cell populations, analyzed by flow cytometry. Appl. Environ. Microbiol. 2005, 71, 1515–1521. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Xia, X. Dambe7: New and improved tools for data analysis in molecular biology and evolution. Mol. Biol. Evol. 2018, 35, 1550–1552. [Google Scholar] [CrossRef]
- Mori, K.; Sant, A.; Kohno, K.; Normington, K.; Gething, M.J.; Sambrook, J.F. A 22 bp cis-acting element is necessary and sufficient for the induction of the yeast KAR2 (bip) gene by unfolded proteins. Embo J. 1992, 11, 2583–2593. [Google Scholar] [CrossRef] [PubMed]
- Van Dalfsen, K.M.; Hodapp, S.; Keskin, A.; Otto, G.M.; Berdan, C.A.; Higdon, A.; Cheunkarndee, T.; Nomura, D.K.; Jovanovic, M.; Brar, G.A. Global proteome remodeling during ER stress involves HAC1-driven expression of long undecoded transcript isoforms. Dev. Cell 2018, 46, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Kohno, K.; Normington, K.; Sambrook, J.; Gething, M.J.; Mori, K. The promoter region of the yeast KAR2 (bip) gene contains a regulatory domain that responds to the presence of unfolded proteins in the endoplasmic reticulum. Mol. Cell Biol. 1993, 13, 877–890. [Google Scholar] [CrossRef] [PubMed]
- Chawla, A.; Chakrabarti, S.; Ghosh, G.; Niwa, M. Attenuation of yeast UPR is essential for survival and is mediated by IRE1 kinase. J. Cell Biol. 2011, 193, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casagrande, R.; Stern, P.; Diehn, M.; Shamu, C.; Osario, M.; Zuniga, M.; Brown, P.O.; Ploegh, H. Degradation of proteins from the ER of s. Cerevisiae requires an intact unfolded protein response pathway. Mol. Cell 2000, 5, 729–735. [Google Scholar] [CrossRef]
- Arroyo, J.; Hutzler, J.; Bermejo, C.; Ragni, E.; Garcia-Cantalejo, J.; Botias, P.; Piberger, H.; Schott, A.; Sanz, A.B.; Strahl, S. Functional and genomic analyses of blocked protein o-mannosylation in baker’s yeast. Mol. Microbiol. 2011, 79, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, M.; Nastase, K.K.; Cunningham, K.W. Essential role of calcineurin in response to endoplasmic reticulum stress. Embo J. 2002, 21, 2343–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardarin, A.; Chedin, S.; Lagniel, G.; Aude, J.C.; Godat, E.; Catty, P.; Labarre, J. Endoplasmic reticulum is a major target of cadmium toxicity in yeast. Mol. Microbiol. 2010, 76, 1034–1048. [Google Scholar] [CrossRef] [PubMed]
- Kawazoe, N.; Kimata, Y.; Izawa, S. Acetic acid causes endoplasmic reticulum stress and induces the unfolded protein response in Saccharomyces cerevisiae. Front. Microbiol. 2017, 8, 1192. [Google Scholar] [CrossRef]
- Chaillot, J.; Tebbji, F.; Remmal, A.; Boone, C.; Brown, G.W.; Bellaoui, M.; Sellam, A. The monoterpene carvacrol generates endoplasmic reticulum stress in the pathogenic fungus Candida albicans. Antimicrob. Agents Chemother 2015, 59, 4584–4592. [Google Scholar] [CrossRef]
- Burt, S.A.; van der Zee, R.; Koets, A.P.; de Graaff, A.M.; van Knapen, F.; Gaastra, W.; Haagsman, H.P.; Veldhuizen, E.J.A. Carvacrol Induces Heat Shock Protein 60 and Inhibits Synthesis of Flagellin in Escherichia coli O157:H7. Appl. Environ. Microbiol. 2007, 73, 4484. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.; Gasser, B.; Dragosits, M.; Sauer, M.; Leparc, G.G.; Tuchler, T.; Kreil, D.P.; Mattanovich, D. Novel insights into the unfolded protein response using Pichia pastoris specific DNA microarrays. BMC Genom. 2008, 9, 390. [Google Scholar] [CrossRef]
- Iracane, E.; Donovan, P.D.; Ola, M.; Butler, G.; Holland, L.M. Identification of an exceptionally long intron in the HAC1 gene of Candida parapsilosis. mSphere 2018, 3, e00532-18. [Google Scholar] [CrossRef] [PubMed]
- Shechtman, C.F.; Henneberry, A.L.; Seimon, T.A.; Tinkelenberg, A.H.; Wilcox, L.J.; Lee, E.; Fazlollahi, M.; Munkacsi, A.B.; Bussemaker, H.J.; Tabas, I.; et al. Loss of subcellular lipid transport due to arv1 deficiency disrupts organelle homeostasis and activates the unfolded protein response. J. Biol. Chem. 2011, 286, 11951–11959. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Feldman, D.E.; Deng, C.; Brown, J.A.; De Giacomo, A.F.; Gaw, A.F.; Shi, G.; Le, Q.T.; Brown, J.M.; Koong, A.C. Identification of mitogen-activated protein kinase signaling pathways that confer resistance to endoplasmic reticulum stress in Saccharomyces cerevisiae. Mol. Cancer Res. 2005, 3, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Bertolotti, A.; Ron, D. Alterations in an IRE1-RNA complex in the mammalian unfolded protein response. J. Cell Sci. 2001, 114, 3207–3212. [Google Scholar] [PubMed]
- Armstrong, M.C.; Sestak, S.; Ali, A.A.; Sagini, H.A.M.; Brown, M.; Baty, K.; Treumann, A.; Schroder, M. Bypass of activation loop phosphorylation by aspartate 836 in activation of the endoribonuclease activity of IRE1. Mol. Cell Biol. 2017, 37. [Google Scholar] [CrossRef]
- Kimmig, P.; Diaz, M.; Zheng, J.; Williams, C.C.; Lang, A.; Aragon, T.; Li, H.; Walter, P. The unfolded protein response in fission yeast modulates stability of select mRNAs to maintain protein homeostasis. eLife 2012, 1, e00048. [Google Scholar] [CrossRef] [PubMed]
- Guydosh, N.R.; Kimmig, P.; Walter, P.; Green, R. Regulated IRE1-dependent mRNA decay requires no-go mRNA degradation to maintain endoplasmic reticulum homeostasis in S. pombe. eLife 2017, 6, e29216. [Google Scholar] [CrossRef] [PubMed]
- Hollien, J.; Lin, J.H.; Li, H.; Stevens, N.; Walter, P.; Weissman, J.S. Regulated IRE1-dependent decay of messenger RNAs in mammalian cells. J. Cell Biol. 2009, 186, 323–331. [Google Scholar] [CrossRef]
- Hollien, J.; Weissman, J.S. Decay of endoplasmic reticulum-localized mRNAs during the unfolded protein response. Science 2006, 313, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Steffen, K.K.; McCormick, M.A.; Pham, K.M.; MacKay, V.L.; Delaney, J.R.; Murakami, C.J.; Kaeberlein, M.; Kennedy, B.K. Ribosome deficiency protects against ER stress in Saccharomyces cerevisiae. Genetics 2012, 191, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.A.; DiDone, L.; Krysan, D.J. Extracellular secretion of overexpressed glycosylphosphatidylinositol-linked cell wall protein utr2/crh2p as a novel protein quality control mechanism in Saccharomyces cerevisiae. Eukaryot. Cell 2010, 9, 1669–1679. [Google Scholar] [CrossRef] [PubMed]
- Travers, K.J.; Patil, C.K.; Wodicka, L.; Lockhart, D.J.; Weissman, J.S.; Walter, P. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell 2000, 101, 249–258. [Google Scholar] [CrossRef]
- Cakir, B. Bax induces activation of the unfolded protein response by inducing HAC1 mRNA splicing in Saccharomyces cerevisiae. Yeast 2012, 29, 395–406. [Google Scholar] [CrossRef]
- Han, D.; Lerner, A.G.; Vande Walle, L.; Upton, J.P.; Xu, W.; Hagen, A.; Backes, B.J.; Oakes, S.A.; Papa, F.R. Ire1alpha kinase activation modes control Alternate endoribonuclease outputs to determine divergent cell fates. Cell 2009, 138, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Brandizzi, F. Ire1: Er stress sensor and cell fate executor. Trends Cell Biol. 2013, 23, 547–555. [Google Scholar] [CrossRef]
- Upton, J.P.; Wang, L.; Han, D.; Wang, E.S.; Huskey, N.E.; Lim, L.; Truitt, M.; McManus, M.T.; Ruggero, D.; Goga, A.; et al. Ire1alpha cleaves select microRNAs during ER stress to derepress translation of proapoptotic caspase-2. Science 2012, 338, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Patil, C.K.; DeRisi, J.; Walter, P. Genome-scale approaches for discovering novel nonconventional splicing substrates of the IRE1 nuclease. Genom. Biol. 2005, 6, R3. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, H.; Brandizzi, F.; Verchot, J.; Wang, A. The UPR branch IRE1-bZIP60 in plants plays an essential role in viral infection and is complementary to the only UPR pathway in yeast. PLoS Genet. 2015, 11, e1005164. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, C.; Wang, A. Divergence and conservation of the major UPR branch IRE1-bZIP signaling pathway across eukaryotes. Sci. Rep. 2016, 6, 27362. [Google Scholar] [CrossRef] [PubMed]
- Tirasophon, W.; Welihinda, A.A.; Kaufman, R.J. A stress response pathway from the endoplasmic reticulum to the nucleus requires a novel bifunctional protein kinase/endoribonuclease (Ire1p) in mammalian cells. Genes Dev. 1998, 12, 1812–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. Xbp1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef]
- Iwakoshi, N.N.; Lee, A.H.; Glimcher, L.H. The x-box binding protein-1 transcription factor is required for plasma cell differentiation and the unfolded protein response. Immunol. Rev. 2003, 194, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Wakasa, Y.; Takahashi, H.; Kawakatsu, T.; Takaiwa, F. Signal transduction by IRE1-mediated splicing of bzip50 and other stress sensors in the endoplasmic reticulum stress response of rice. Plant J. 2012, 69, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Calfon, M.; Zeng, H.; Urano, F.; Till, J.H.; Hubbard, S.R.; Harding, H.P.; Clark, S.G.; Ron, D. Ire1 couples endoplasmic reticulum load to secretory capacity by processing the xbp-1 mRNA. Nature 2002, 415, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Foti, D.M.; Welihinda, A.; Kaufman, R.J.; Lee, A.S. Conservation and divergence of the yeast and mammalian unfolded protein response. Activation of specific mammalian endoplasmic reticulum stress element of the grp78/bip promoter by yeast HAC1. J. Biol. Chem. 1999, 274, 30402–30409. [Google Scholar] [CrossRef]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.J.; Strudwick, N.; Ali, A.A.; Paton, A.W.; Paton, J.C.; Schroder, M. Measuring signaling by the unfolded protein response. Methods Enzymol. 2011, 491, 261–292. [Google Scholar]
- Hernandez-Elvira, M.; Torres-Quiroz, F.; Escamilla-Ayala, A.; Dominguez-Martin, E.; Escalante, R.; Kawasaki, L.; Ongay-Larios, L.; Coria, R. The unfolded protein response pathway in the yeast kluyveromyces lactis. A comparative view among yeast species. Cells 2018, 7, 106. [Google Scholar] [CrossRef]
- Peschek, J.; Acosta-Alvear, D.; Mendez, A.S.; Walter, P. A conformational RNA zipper promotes intron ejection during non-conventional XBP1 mRNA splicing. EMBO Rep. 2015, 16, 1688–1698. [Google Scholar] [CrossRef] [PubMed]
- Hooks, K.B.; Griffiths-Jones, S. Conserved RNA structures in the non-canonical HAC1/XBP1 intron. RNA Biol. 2011, 8, 552–556. [Google Scholar] [CrossRef]
- Li, W.; Okreglak, V.; Peschek, J.; Kimmig, P.; Zubradt, M.; Weissman, J.S.; Walter, P. Engineering ER-stress dependent non-conventional mRNA splicing. eLife 2018, 7, e35388. [Google Scholar] [CrossRef] [PubMed]
- Iwawaki, T.; Tokuda, M. Function of yeast and amphioxus tRNA ligase in IRE1alpha-dependent XBP1 mRNA splicing. Biochem. Biophys. Res. Commun. 2011, 413, 527–531. [Google Scholar] [CrossRef]
- Jurkin, J.; Henkel, T.; Nielsen, A.F.; Minnich, M.; Popow, J.; Kaufmann, T.; Heindl, K.; Hoffmann, T.; Busslinger, M.; Martinez, J. The mammalian tRNA ligase complex mediates splicing of XBP1 mRNA and controls antibody secretion in plasma cells. Embo J. 2014, 33, 2922–2936. [Google Scholar] [CrossRef]
- Lu, Y.; Liang, F.X.; Wang, X. A synthetic biology approach identifies the mammalian UPR RNA ligase rtcb. Mol. Cell 2014, 55, 758–770. [Google Scholar] [CrossRef]
- Mori, T.; Ogasawara, C.; Inada, T.; Englert, M.; Beier, H.; Takezawa, M.; Endo, T.; Yoshihisa, T. Dual functions of yeast tRNA ligase in the unfolded protein response: Unconventional cytoplasmic splicing of HAC1 pre-mRNA is not sufficient to release translational attenuation. Mol. Biol. Cell 2010, 21, 3722–3734. [Google Scholar] [CrossRef]
- Oh, M.H.; Cheon, S.A.; Kang, H.A.; Kim, J.Y. Functional characterization of the unconventional splicing of yarrowia lipolytica HAC1 mRNA induced by unfolded protein response. Yeast 2010, 27, 443–452. [Google Scholar] [CrossRef]
- Mulder, H.J.; Nikolaev, I. Haca-dependent transcriptional switch releases haca mRNA from a translational block upon endoplasmic reticulum stress. Eukaryot. Cell 2009, 8, 665–675. [Google Scholar] [CrossRef] [PubMed]
- Saloheimo, M.; Valkonen, M.; Penttila, M. Activation mechanisms of the HAC1-mediated unfolded protein response in filamentous fungi. Mol. Microbiol. 2003, 47, 1149–1161. [Google Scholar] [CrossRef]
- Tirosh, B.; Iwakoshi, N.N.; Glimcher, L.H.; Ploegh, H.L. Rapid turnover of unspliced xbp-1 as a factor that modulates the unfolded protein response. J. Biol. Chem. 2006, 281, 5852–5860. [Google Scholar] [CrossRef] [PubMed]
- Bowring, C.E.; Llewellyn, D.H. Differences in HAC1 mRNA processing and translation between yeast and mammalian cells indicate divergence of the eukaryotic ER stress response. Biochem. Biophys. Res. Commun. 2001, 287, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Sidrauski, C.; Kaufman, R.J.; Walter, P. A role for presenilin-1 in nuclear accumulation of IRE1 fragments and induction of the mammalian unfolded protein response. Cell 1999, 99, 691–702. [Google Scholar] [CrossRef]
Figure 1.
HAC1 gene in Baker’s yeast with a 252-nt intron flanked by two exons (whose coding sequences are pink-shaded), with part of the intron forming base-pair interaction with 5’UTR to inhibit translation initiation. 3’BE is a cis-acting bipartite element signaling for HAC1 pre-mRNA to be carried to discrete foci of Ire1p oligomers to be spliced [26]. The red ‘UGA’ within the intron stops occasional translation of the unspliced mRNA. The blue-shaded UACUAAG resembles a branchpoint site (BPS, consensus UACUAAC), but its deletion does not affect HAC1 pre-mRNA splicing [27].
Figure 1.
HAC1 gene in Baker’s yeast with a 252-nt intron flanked by two exons (whose coding sequences are pink-shaded), with part of the intron forming base-pair interaction with 5’UTR to inhibit translation initiation. 3’BE is a cis-acting bipartite element signaling for HAC1 pre-mRNA to be carried to discrete foci of Ire1p oligomers to be spliced [26]. The red ‘UGA’ within the intron stops occasional translation of the unspliced mRNA. The blue-shaded UACUAAG resembles a branchpoint site (BPS, consensus UACUAAC), but its deletion does not affect HAC1 pre-mRNA splicing [27].

Figure 2.
A simplified view of IRE1+HAC1 UPR signaling pathway in yeast. Accumulation of unfolded/misfolded proteins in the endoplasmic reticulum (ER) lumen triggers the autophosphorylation and oligomerization of Ire1p which, together with Trl1p, removes the intron to release translation inhibition of HAC1 mRNA. Hac1p enters the nucleus and transactivates target genes with a UPR element (UPRE) whose encoded proteins alleviate ER stress.
Figure 2.
A simplified view of IRE1+HAC1 UPR signaling pathway in yeast. Accumulation of unfolded/misfolded proteins in the endoplasmic reticulum (ER) lumen triggers the autophosphorylation and oligomerization of Ire1p which, together with Trl1p, removes the intron to release translation inhibition of HAC1 mRNA. Hac1p enters the nucleus and transactivates target genes with a UPR element (UPRE) whose encoded proteins alleviate ER stress.

Figure 3.
Ire1p domains, domain-specific theoretical isoelectric point (pI), and hydrophobicity [70] along the protein (sliding window of 10 amino acids). N: N-terminal domain; cLD: core luminal domain; TM: transmembrane domain; Linker: positively charged linker domain; Kinase: Kinase domain; RNase: RNase domain. The red arrow points to a highly hydrophobic stretch near the N-terminus that needs to be either buried deep into the folded Ire1p or insert into a membrane. Results generated from DAMBE [71].
Figure 3.
Ire1p domains, domain-specific theoretical isoelectric point (pI), and hydrophobicity [70] along the protein (sliding window of 10 amino acids). N: N-terminal domain; cLD: core luminal domain; TM: transmembrane domain; Linker: positively charged linker domain; Kinase: Kinase domain; RNase: RNase domain. The red arrow points to a highly hydrophobic stretch near the N-terminus that needs to be either buried deep into the folded Ire1p or insert into a membrane. Results generated from DAMBE [71].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Contrast in splicing efficiency between some efficiently spliced yeast genes and HAC1, quantified by software ARSDA [59] and available in Supplemental file in Xia [58].
| Gene | SystName | NEE | NEI5 | NEI3 |
|---|---|---|---|---|
| RPL36B | YPL249C-A | 3603 | 10 | 16 |
| RPL18A | YOL120C | 2789 | 10 | 11 |
| RPL39 | YJL189W | 7590 | 11 | 40 |
| RIM1 | YCR028C-A | 146 | 0 | 1 |
| RPL43A | YPR043W | 4083 | 14 | 15 |
| RPL40A | YIL148W | 3928 | 5 | 19 |
| RPL23B | YER117W | 3192 | 19 | 7 |
| RPL2B | YIL018W | 2974 | 4 | 14 |
| RPS21B | YJL136C | 4192 | 10 | 14 |
| RPL31B | YLR406C | 919 | 4 | 2 |
| RPL25 | YOL127W | 6536 | 13 | 22 |
| RPL31A | YDL075W | 3032 | 10 | 6 |
| RPL6A | YML073C | 3009 | 5 | 10 |
| RPS4B | YHR203C | 3411 | 13 | 6 |
| RPS21A | YKR057W | 2205 | 5 | 4 |
| RPS4A | YJR145C | 4206 | 8 | 10 |
| RPL33A | YPL143W | 4880 | 11 | 10 |
| SAC6 | YDR129C | 242 | 1 | 0 |
| RPS7A | YOR096W | 5093 | 4 | 13 |
| HAC1 | YFL031W | 32 | 844 | 704 |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xia, X. Translation Control of HAC1 by Regulation of Splicing in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2019, 20, 2860. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122860
AMA Style
Xia X. Translation Control of HAC1 by Regulation of Splicing in Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2019; 20(12):2860. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122860
Chicago/Turabian StyleXia, Xuhua. 2019. "Translation Control of HAC1 by Regulation of Splicing in Saccharomyces cerevisiae" International Journal of Molecular Sciences 20, no. 12: 2860. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20122860
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.