Transcriptomic Analysis Reveals the High-Oleic Acid Feedback Regulating the Homologous Gene Expression of Stearoyl-ACP Desaturase 2 (SAD2) in Peanuts

 ,
,
Abstract
:1. Introduction
2. Results
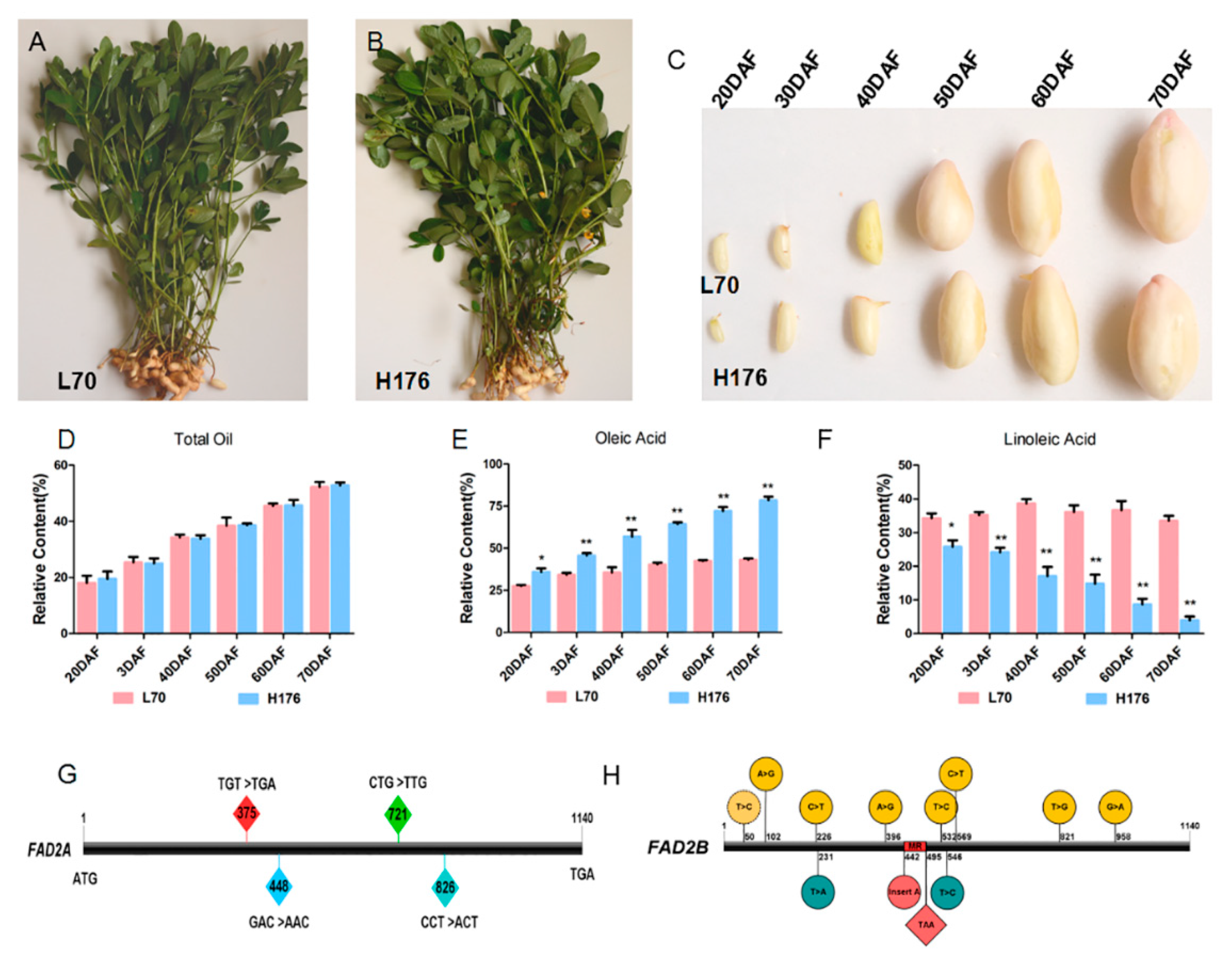
2.1. Changes of Oil Composition during the Developmental Stages of Peanut Seeds
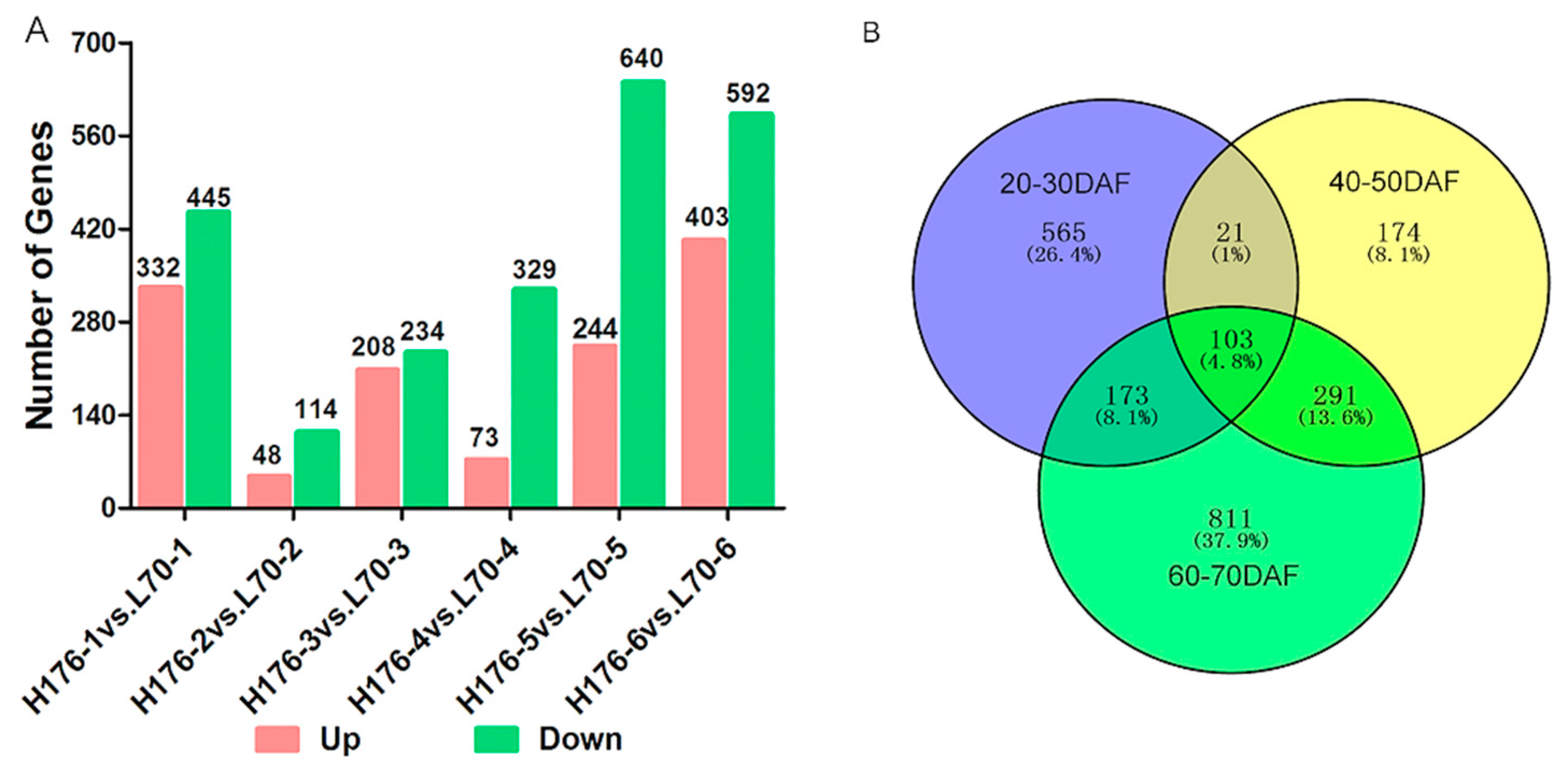
2.2. Transcriptome Assembly
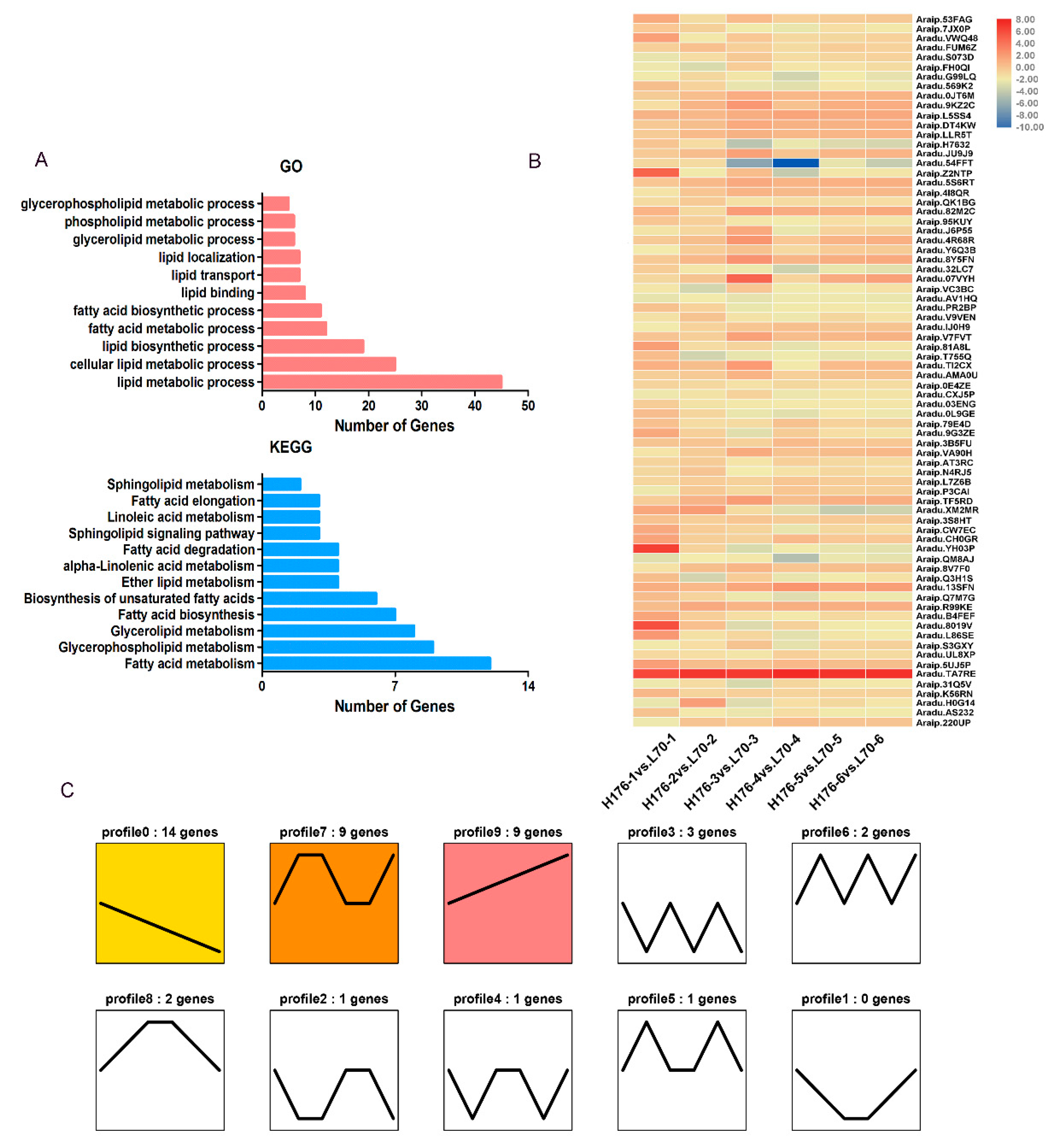
2.3. Analysis of Biological Processes in High-Oleic Peanuts
2.4. Expression Analysis of DEGs
2.5. Dynamic Changes of DEGs Related to Lipid Metabolism in High-Oleic Peanut
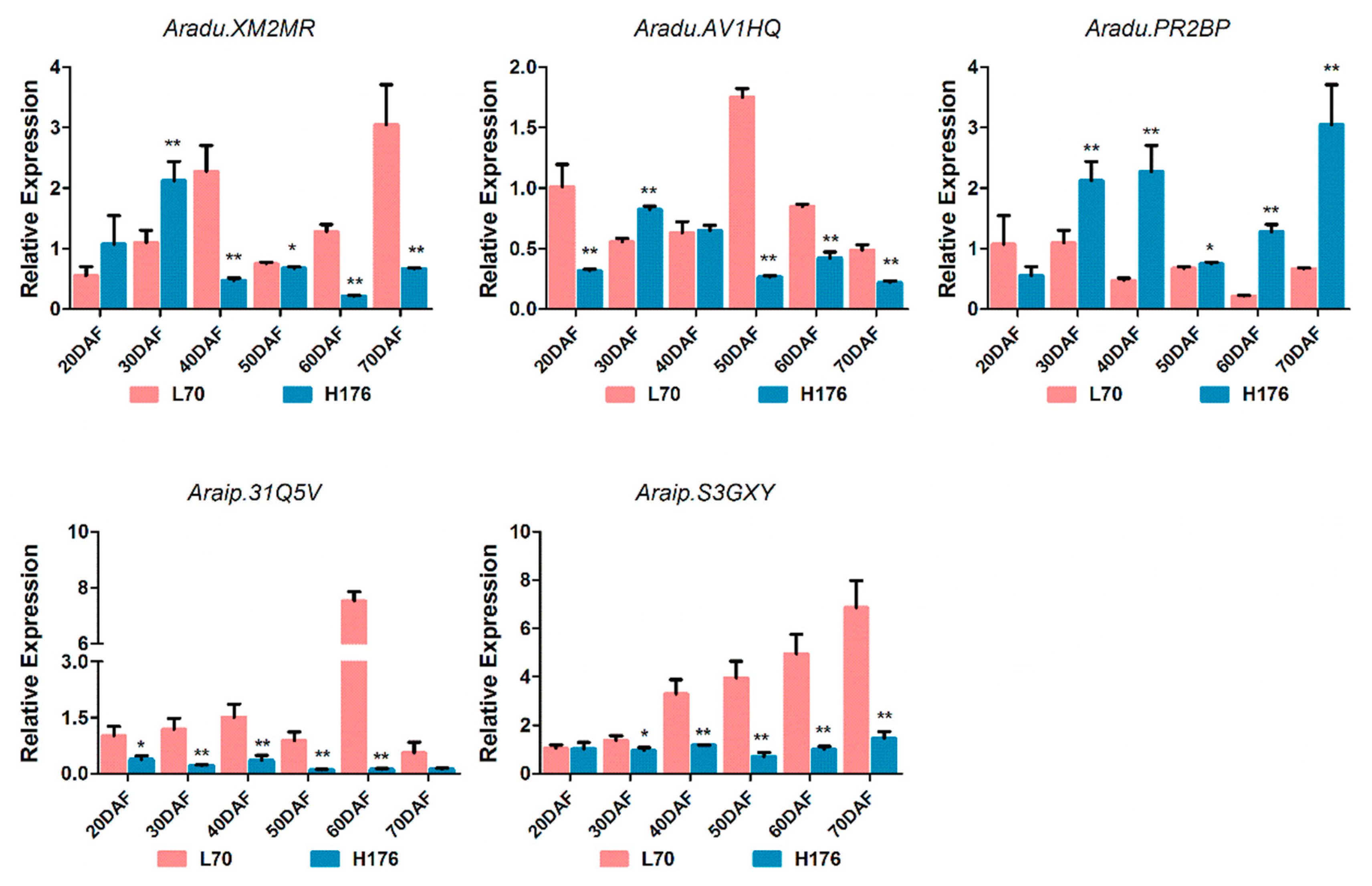
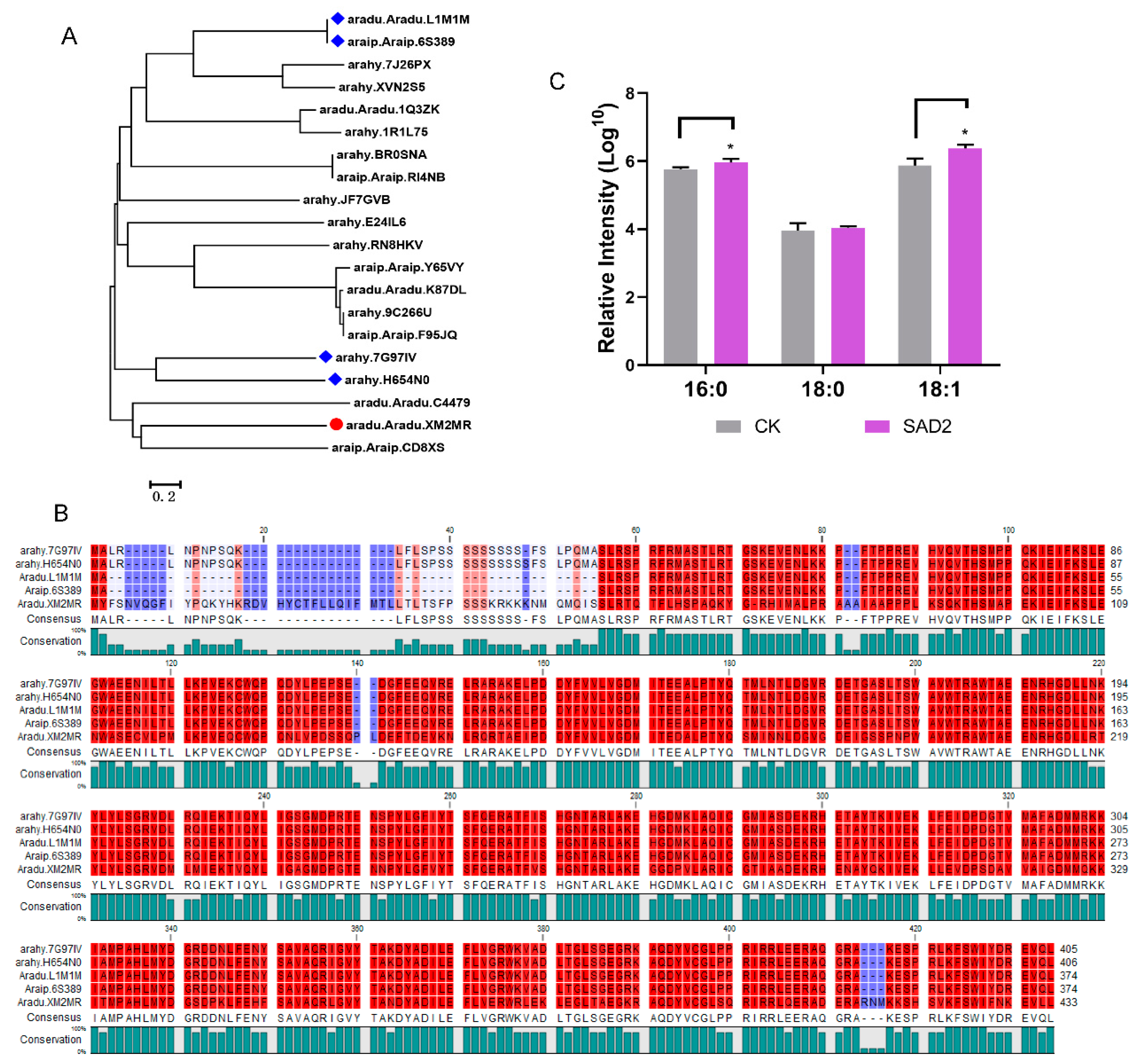
2.6. Analysis of the Expression of FADs at the mRNA level
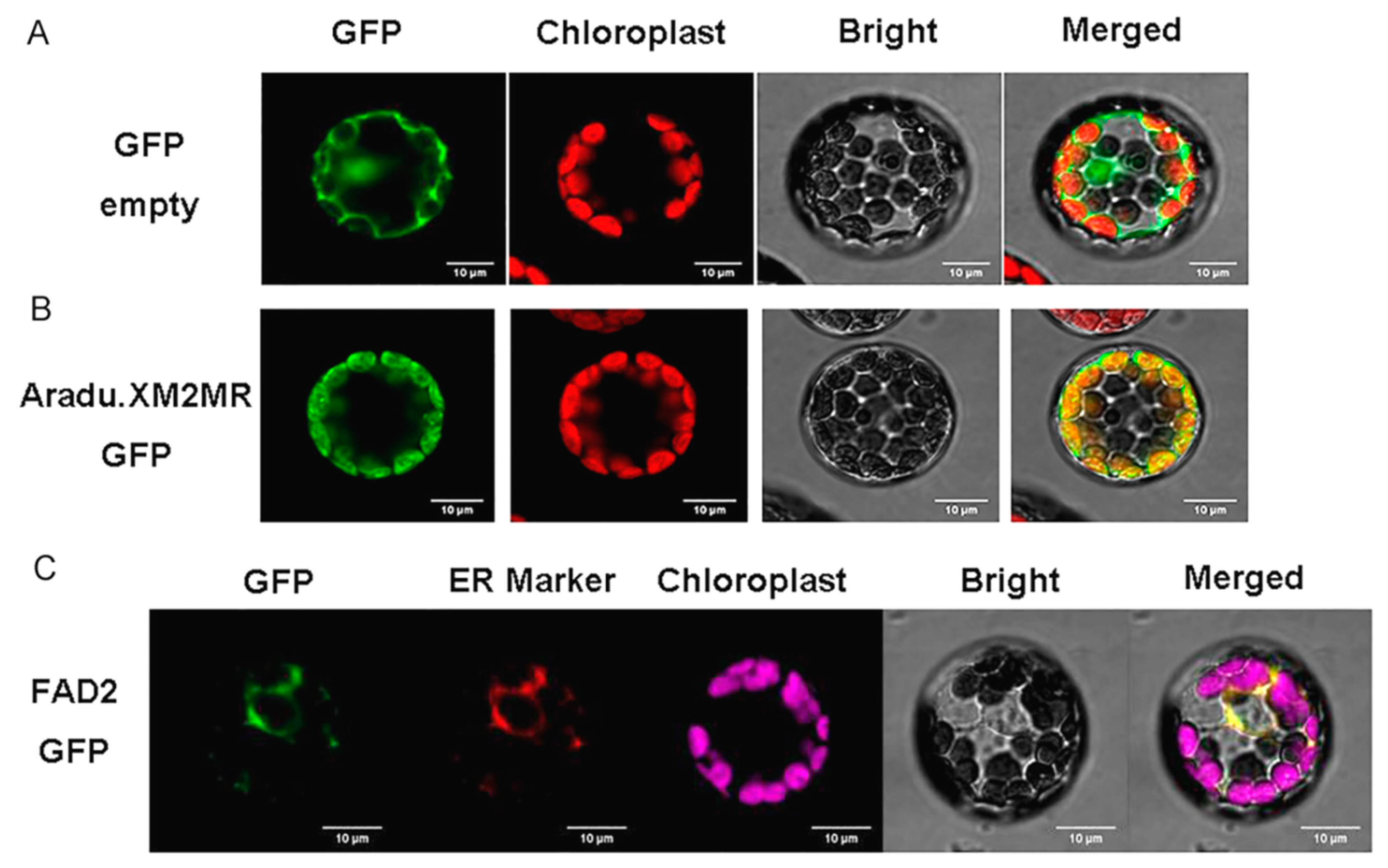
2.7. Subcellular Localization of FAD2 and SAD2 (Aradu.XM2MR)
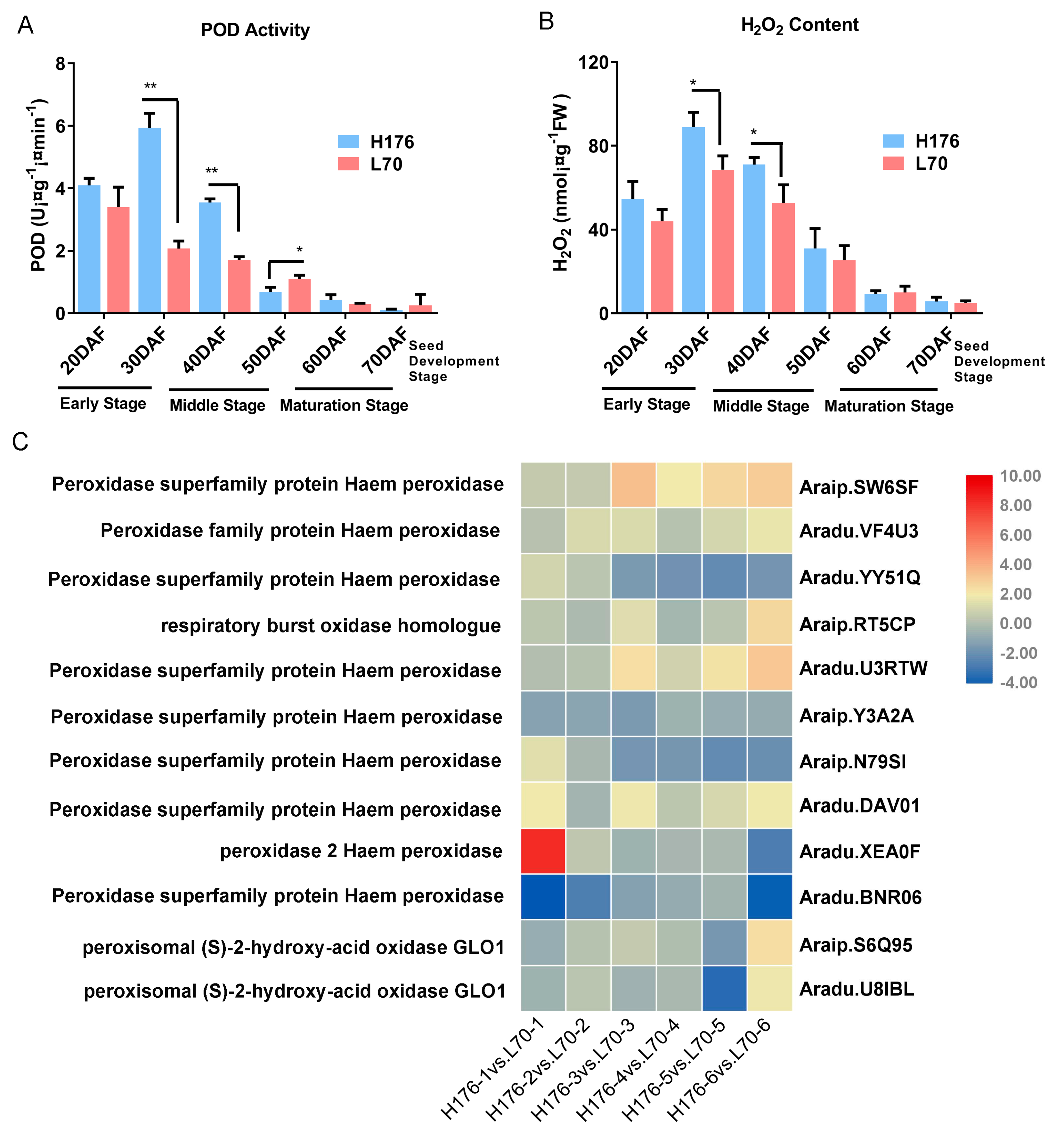
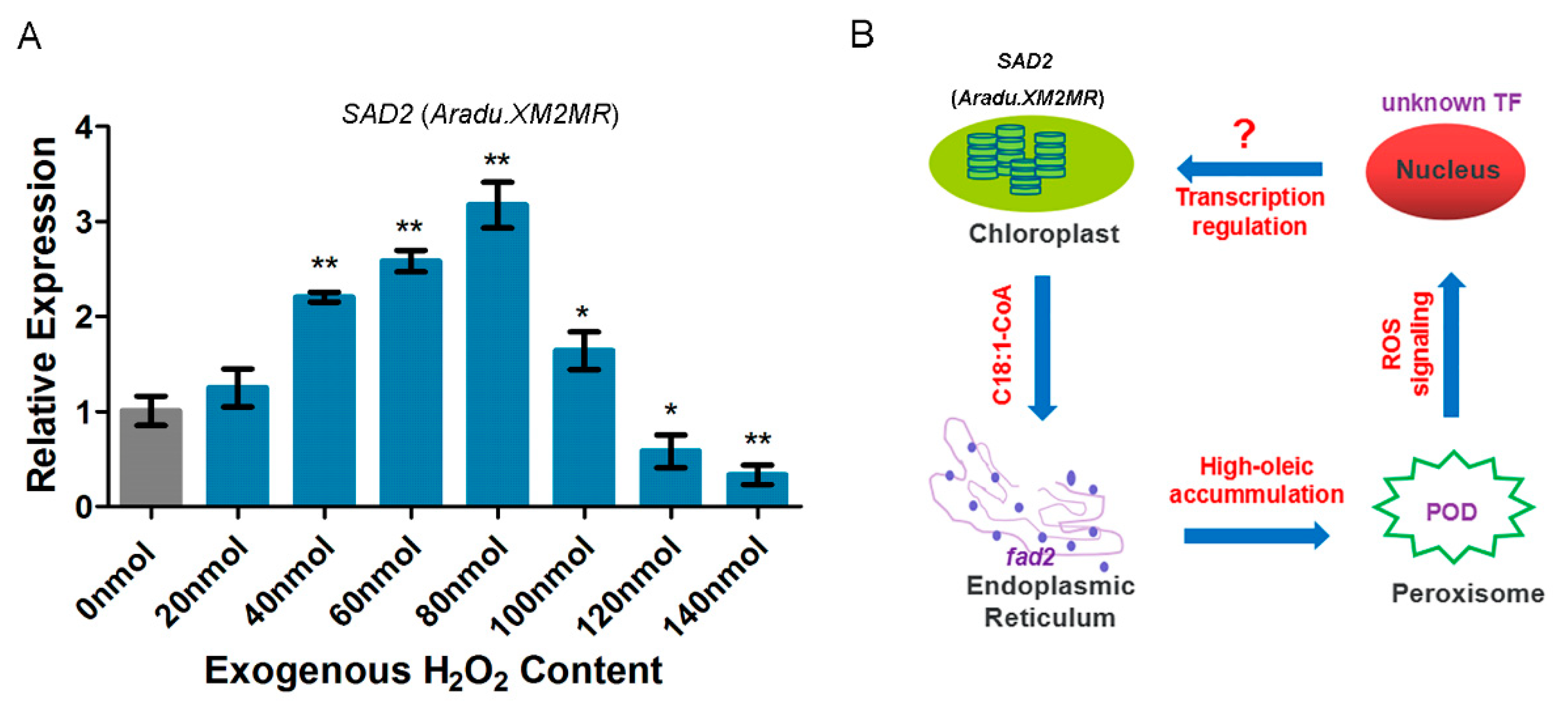
2.8. The Enhanced Activity of Peroxidase (POD) in High-Oleic H176
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Analysis of OA, LOA, and Oil Content
4.3. Total RNA Isolation, Library Construction, and Transcriptome Sequencing
4.4. Bioinformatic Analysis of Transcriptome Data
4.5. Real-Time PCR Analysis
4.6. Subcellular Localization Analysis
4.7. POD Activity Assay
4.8. Ectopic Expression of SAD2 (Aradu.XM2MR) in Yeast Cells and Fatty Acid Composition Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhao, J.; Huang, L.; Ren, X.; Pandey, M.K.; Wu, B.; Chen, Y.; Zhou, X.; Chen, W.; Xia, Y.; Li, Z.; et al. Genetic variation and association mapping of seed-related traits in cultivated peanut (Arachis hypogaea L.) using single-locus simple sequence repeat markers. Front. Plant Sci. 2017, 8, 2105. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Chen, C.Y.; Tonnis, B.; Pinnow, D.; Davis, J.; An, Y.C.; Dang, P. Changes of seed weight, fatty acid composition, and oil and protein contents from different peanut FAD2 genotypes at different seed developmental and maturation stages. J. Agric. Food Chem. 2018, 66, 3658–3665. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H.; Pandey, M.K.; Yang, Q.; Wang, X.; Garg, V.; Li, H.; Chi, X.; Doddamani, D.; Hong, Y.; et al. Draft genome of the peanut A-genome progenitor (Arachis duranensis) provides insights into geocarpy, oil biosynthesis, and allergens. Proc. Natl. Acad. Sci. USA 2016, 113, 6785–6790. [Google Scholar] [CrossRef] [PubMed]
- Isleib, T.G.; Pattee, H.E.; Sanders, T.H.; Hendrix, K.W.; Dean, L.O. Compositional and sensory comparisons between normal- and high-oleic peanuts. J. Agric. Food Chem. 2006, 54, 1759–1763. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.; Khalid, N.; Ahmed, I.; Shahzad, A.; Suleria, H.A. Physicochemical characteristics, functional properties, and nutritional benefits of peanut oil: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1562–1575. [Google Scholar] [CrossRef] [PubMed]
- Barkley, N.A.; Isleib, T.G.; Wang, M.L.; Pittman, R.N. Genotypic effect of ahFAD2 on fatty acid profiles in six segregating peanut (Arachis hypogaea L.) populations. BMC Genet. 2013, 14, 62. [Google Scholar] [CrossRef] [PubMed]
- Block, M.A.; Jouhet, J. Lipid trafficking at endoplasmic reticulum-chloroplast membrane contact sites. Curr. Opin. Cell Biol. 2015, 35, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Joyard, J.; Ferro, M.; Masselon, C.; Seigneurin-Berny, D.; Salvi, D.; Garin, J.; Rolland, N. Chloroplast proteomics highlights the subcellular compartmentation of lipid metabolism. Prog. Lipid. Res. 2010, 49, 128–158. [Google Scholar] [CrossRef]
- Dar, A.A.; Choudhury, A.R.; Kancharla, P.K.; Arumugam, N. The FAD2 gene in plants: Occurrence, regulation, and role. Front. Plant Sci. 2017, 8, 1789. [Google Scholar] [CrossRef]
- Patel, M.; Jung, S.; Moore, K.; Powell, G.; Ainsworth, C.; Abbott, A. High-oleate peanut mutants result from a MITE insertion into the FAD2 gene. Theor. Appl. Genet. 2004, 108, 1492–1502. [Google Scholar] [CrossRef]
- Jung, S.; Swift, D.; Sengoku, E.; Patel, M.; Teule, F.; Powell, G.; Moore, K.; Abbott, A. The high oleate trait in the cultivated peanut (Arachis hypogaea L.). I. Isolation and characterization of two genes encoding microsomal oleoyl-PC desaturases. Mol. Gen. Genet. 2000, 263, 796–805. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Zhao, Y.; Prakash, C.S.; He, G.; Yin, D. Insights into the novel members of the FAD2 gene family involved in high-oleate fluxes in peanut. Genome 2015, 58, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yang, X.; Tian, L.; Chen, L.; Yu, W. Identification of peanut (Arachis hypogaea) chromosomes using a fluorescence in situ hybridization system reveals multiple hybridization events during tetraploid peanut formation. New Phytol. 2016, 211, 1424–1439. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Chen, H.; Yang, M.; Wang, J.; Pandey, M.K.; Zhang, C.; Chang, W.C.; Zhang, L.; Zhang, X.; Tang, R.; et al. The genome of cultivated peanut provides insight into legume karyotypes, polyploid evolution and crop domestication. Nat. Genet. 2019, 51, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Bertioli, D.J.; Jenkins, J.; Clevenger, J.; Dudchenko, O.; Gao, D.; Seijo, G.; Leal-Bertioli, S.; Ren, L.; Farmer, A.D.; Pandey, M.K.; et al. The genome sequence of segmental allotetraploid peanut Arachis hypogaea. Nat. Genet. 2019, 51, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, Q.; Liu, H.; Zhang, J.; Hong, Y.; Lan, H.; Li, H.; Wang, J.; Liu, H.; Li, S.; et al. Sequencing of cultivated peanut, arachis hypogaea, yields insights into genome evolution and oil improvement. Mol. Plant 2019. [Google Scholar] [CrossRef] [PubMed]
- Clevenger, J.; Chu, Y.; Scheffler, B.; Ozias-Akins, P. A developmental transcriptome map for allotetraploid Arachis hypogaea. Front. Plant Sci. 2016, 7, 1446. [Google Scholar] [CrossRef]
- Bertioli, D.J.; Cannon, S.B.; Froenicke, L.; Huang, G.; Farmer, A.D.; Cannon, E.K.; Liu, X.; Gao, D.; Clevenger, J.; Dash, S.; et al. The genome sequences of Arachis duranensis and Arachis ipaensis, the diploid ancestors of cultivated peanut. Nat. Genet. 2016, 48, 438–446. [Google Scholar] [CrossRef]
- Chen, X.; Yang, Q.; Li, H.; Li, H.; Hong, Y.; Pan, L.; Chen, N.; Zhu, F.; Chi, X.; Zhu, W.; et al. Transcriptome-wide sequencing provides insights into geocarpy in peanut (Arachis hypogaea L.). Plant Biotechnol. J. 2016, 14, 1215–1224. [Google Scholar] [CrossRef]
- Yin, D.; Wang, Y.; Zhang, X.; Li, H.; Lu, X.; Zhang, J.; Zhang, W.; Chen, S. De novo assembly of the peanut (Arachis hypogaea L.) seed transcriptome revealed candidate unigenes for oil accumulation pathways. PLoS ONE 2013, 8, e73767. [Google Scholar] [CrossRef]
- Gupta, K.; Kayam, G.; Faigenboim-Doron, A.; Clevenger, J.; Ozias-Akins, P.; Hovav, R. Gene expression profiling during seed-filling process in peanut with emphasis on oil biosynthesis networks. Plant Sci. 2016, 248, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, X.; Yang, Q.; Pan, L.; Chen, M.; He, Y.; Yang, Z.; Yu, S. Isolation and characterization of fatty acid desaturase genes from peanut (Arachis hypogaea L.). Plant Cell Rep. 2011, 30, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Roman, A.; Hernandez, M.L.; Soria-Garcia, A.; Lopez-Gomollon, S.; Lagunas, B.; Picorel, R.; Martinez-Rivas, J.M.; Alfonso, M. Non-redundant contribution of the plastidial FAD8 omega-3 desaturase to glycerolipid unsaturation at different temperatures in Arabidopsis. Mol. Plant 2015, 8, 1599–1611. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Xu, C. Genetic analysis of Arabidopsis mutants impaired in plastid lipid import reveals a role of membrane lipids in chloroplast division. Plant Signal. Behav. 2011, 6, 458–460. [Google Scholar] [CrossRef]
- Zhang, J.T.; Zhu, J.Q.; Zhu, Q.; Liu, H.; Gao, X.S.; Zhang, H.X. Fatty acid desaturase−6 (Fad6) is required for salt tolerance in Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2009, 390, 469–474. [Google Scholar] [CrossRef] [PubMed]
- McConn, M.; Hugly, S.; Browse, J.; Somerville, C. A mutation at the fad8 locus of arabidopsis identifies a second chloroplast [omega]-3 desaturase. Plant Physiol. 1994, 106, 1609–1614. [Google Scholar] [CrossRef]
- Liu, H.; Li, H.; Gu, J.; Deng, L.; Ren, L.; Hong, Y.; Lu, Q.; Chen, X.; Liang, X. Identification of the candidate proteins related to oleic acid accumulation during peanut (Arachis hypogaea L.) seed development through comparative proteome analysis. Int. J. Mol. Sci. 2018, 19, 1235. [Google Scholar] [CrossRef]
- Yang, W.; Dong, R.; Liu, L.; Hu, Z.; Li, J.; Wang, Y.; Ding, X.; Chu, Z. A novel mutant allele of SSI2 confers a better balance between disease resistance and plant growth inhibition on Arabidopsis thaliana. BMC Plant Biol. 2016, 16, 208. [Google Scholar] [CrossRef]
- Mandal, M.K.; Chandra-Shekara, A.C.; Jeong, R.D.; Yu, K.; Zhu, S.; Chanda, B.; Navarre, D.; Kachroo, A.; Kachroo, P. Oleic acid-dependent modulation of NITRIC OXIDE ASSOCIATED1 protein levels regulates nitric oxide-mediated defense signaling in Arabidopsis. Plant Cell 2012, 24, 1654–1674. [Google Scholar] [CrossRef]
- Chen, G.; Peng, Z.Y.; Shan, L.; Xuan, N.; Tang, G.Y.; Zhang, Y.; Li, L.; He, Q.F.; Bi, Y.P. Cloning of acyl-ACP thioesterase FatA from Arachis hypogaea L. and its expression in Escherichia coli. J. Biomed. Biotechnol. 2012, 2012, 652579. [Google Scholar] [CrossRef] [PubMed]
- Lung, S.C.; Chye, M.L. Acyl-CoA-Binding Proteins (ACBPs) in plant development. Subcell. Biochem. 2016, 86, 363–404. [Google Scholar] [PubMed]
- Shimada, T.L.; Hayashi, M.; Hara-Nishimura, I. Membrane dynamics and multiple functions of oil bodies in seeds and leaves. Plant Physiol. 2018, 176, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Botella, C.; Sautron, E.; Boudiere, L.; Michaud, M.; Dubots, E.; Yamaryo-Botte, Y.; Albrieux, C.; Marechal, E.; Block, M.A.; Jouhet, J. ALA10, a phospholipid flippase, controls FAD2/FAD3 desaturation of phosphatidylcholine in the ER and affects chloroplast lipid composition in Arabidopsis thaliana. Plant Physiol. 2016, 170, 1300–1314. [Google Scholar] [PubMed]
- Van Lijsebettens, M.; Grasser, K.D. Transcript elongation factors: Shaping transcriptomes after transcript initiation. Trends Plant Sci. 2014, 19, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Baud, S. Seeds as oil factories. Plant Reprod. 2018, 31, 213–235. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.; Carrier, D.J.; Schaedler, T.; Waterham, H.R.; van Roermund, C.W.; Theodoulou, F.L. Peroxisomal ABC transporters: Functions and mechanism. Biochem. Soc. Trans. 2015, 43, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.T.; Gonzalez, K.L.; Bartel, B. Peroxisome function, biogenesis, and dynamics in plants. Plant Physiol. 2018, 176, 162–177. [Google Scholar] [CrossRef]
- Porta, H.; Rocha-Sosa, M. Plant lipoxygenases. physiological and molecular features. Plant Physiol. 2002, 130, 15–21. [Google Scholar] [CrossRef]
- Arent, S.; Pye, V.E.; Henriksen, A. Structure and function of plant acyl-CoA oxidases. Plant Physiol. Biochem. 2008, 46, 292–301. [Google Scholar] [CrossRef]
- Tognetti, V.B.; Bielach, A.; Hrtyan, M. Redox regulation at the site of primary growth: Auxin, cytokinin and ROS crosstalk. Plant Cell Environ. 2017, 40, 2586–2605. [Google Scholar] [CrossRef] [PubMed]
- Darnet, S.; Rahier, A. Plant sterol biosynthesis: Identification of two distinct families of sterol 4alpha-methyl oxidases. Biochem. J. 2004, 378, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.; Post-Beittenmiller, D.; Jaworski, J.G. KCS1 encodes a fatty acid elongase 3-ketoacyl-CoA synthase affecting wax biosynthesis in Arabidopsis thaliana. Plant J. 1999, 17, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Ekstrom, A.G.; Kelly, V.; Marles-Wright, J.; Cockroft, S.L.; Campopiano, D.J. Structural evidence for the covalent modification of FabH by 4,5-dichloro−1,2-dithiol−3-one (HR45). Org. Biomol. Chem. 2017, 15, 6310–6313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiao, J.; Xu, J.; Fu, T.; Cao, Z.; Zhu, L.; Chen, H.Z.; Shen, X.; Jiang, H.; Zhang, L. Crystal structure of FabZ-ACP complex reveals a dynamic seesaw-like catalytic mechanism of dehydratase in fatty acid biosynthesis. Cell Res. 2016, 26, 1330–1344. [Google Scholar] [CrossRef]
- Wen, S.; Liu, H.; Li, X.; Chen, X.; Hong, Y.; Li, H.; Lu, Q.; Liang, X. TALEN-mediated targeted mutagenesis of fatty acid desaturase 2 (FAD2) in peanut (Arachis hypogaea L.) promotes the accumulation of oleic acid. Plant Mol. Biol. 2018, 97, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Sui, N.; Wang, Y.; Liu, S.; Yang, Z.; Wang, F.; Wan, S. Transcriptomic and physiological evidence for the relationship between unsaturated fatty acid and salt stress in peanut. Front. Plant Sci. 2018, 9, 7. [Google Scholar] [CrossRef]
- Liu, H.; Dong, S.; Gu, F.; Liu, W.; Yang, G.; Huang, M.; Xiao, W.; Liu, Y.; Guo, T.; Wang, H.; et al. NBS-LRR protein Pik-H4 interacts with OsBIHD1 to balance rice blast resistance and growth by coordinating ethylene-brassinosteroid pathway. Front. Plant Sci. 2017, 8, 127. [Google Scholar] [CrossRef]
- Nelson, B.K.; Cai, X.; Nebenfuhr, A. A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 2007, 51, 1126–1136. [Google Scholar] [CrossRef]
- Liu, H.; Guo, Z.; Gu, F.; Ke, S.; Sun, D.; Dong, S.; Liu, W.; Huang, M.; Xiao, W.; Yang, G.; et al. 4-Coumarate-CoA ligase-like gene OsAAE3 negatively mediates the rice blast resistance, floret development and lignin biosynthesis. Front. Plant Sci. 2016, 7, 2041. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Average Raw Reads | Average Mapped Reads | Average Multiple Mapped Reads | Average Uniquely Mapped Reads | Average Mapped Ratio (%) |
|---|---|---|---|---|---|
| Reads mapped to Arachis duranensis (AA) genome | |||||
| H176 | 22,918,504.50 | 13,166,271.00 | 360,387.50 | 12,805,882.83 | 57.33 |
| L70 | 25,944,882.83 | 14,650,167.00 | 331,180.67 | 14,318,985.50 | 56.33 |
| Reads mapped to Arachis ipaensis (BB) genome | |||||
| H176 | 22,918,504.50 | 13,441,533.00 | 272,607.00 | 13,168,925.83 | 58.67 |
| L70 | 26,278,776.33 | 15,236,358.17 | 310,790.83 | 14,925,567.50 | 57.83 |
| Gene Name in Diploid Ancestor | Gene Name in Cultivated Peanut | Log2 Fold Change | Function Description | |||||
|---|---|---|---|---|---|---|---|---|
| Stage 1 H176-1 vs. L70-1 | Stage 2 H176-2 vs. L70-2 | Stage 3 H176-3 vs. L70-3 | Stage 4 H176-4 vs. L70-4 | Stage 5 H176-5 vs. L70-5 | Stage 6 H176-6 vs. L70-6 | |||
| Araip.53FAG | Arahy.6ZS5B3 | 1.39 | −1.1 | 0.47 | −0.56 | −0.43 | −0.23 | aldehyde dehydrogenase family 3 member F1-like |
| Araip.7JX0P | Arahy.7L16MH | −0.2 | −0.61 | −1.87 | −2.35 | −1.34 | −1.8 | triacylglycerol lipase |
| Aradu.VWQ48 | Arahy.94LY4L | 1.85 | −1.77 | −0.21 | −1.04 | −0.98 | −0.69 | lipid transfer protein |
| Aradu.FUM6Z | Arahy.R5P48D | −0.56 | 0.19 | −0.55 | −1.14 | −0.39 | −0.7 | UDP (uracil 5′-diphosphate) -glycosyltransferase superfamily protein |
| Aradu.S073D | Arahy.RJ9TTJ | −2.41 | −1.25 | −0.26 | −1.24 | −0.84 | −0.73 | senescence-associated carboxylesterase 101-like isoform X2 |
| Araip.FH0QI | Arahy.W0JK2Q | −2.16 | −3.06 | −0.45 | −1.74 | −1.45 | −1.12 | phosphatidylinositol-4-phosphate 5-kinase family protein |
| Aradu.G99LQ | Arahy.ZS7YSR | −1.86 | −0.99 | −1.64 | −3.45 | −1.98 | −2.53 | seed linoleate 9S-lipoxygenase |
| Aradu.569K2 | Arahy.LS6X36 | 0.31 | −0.71 | −2.37 | −2.68 | −1.73 | −2.22 | phosphoinositide phospholipase C 6-like |
| Aradu.0JT6M | Arahy.KBA53Y | −0.33 | 0.28 | 1.21 | 0.63 | 0.73 | 0.86 | biotin carboxyl carrier acetyl-CoA carboxylase |
| Aradu.9KZ2C | Arahy.UBEE9G | −1.37 | 0.78 | 2.29 | 0.15 | 1.13 | 1.21 | 3-oxoacyl-(acyl-carrier-protein) synthase 3 FabH |
| Araip.L5SS4 | Arahy.G623B4 | 0.93 | 0.65 | 1.24 | 1.66 | 1.34 | 1.42 | GDSL(Gly-Asp-Ser-Leu) -like lipase/acylhydrolase superfamily protein |
| Araip.DT4KW | Arahy.R4EZU5 | −0.27 | 0.31 | 1.39 | 0.96 | 0.98 | 1.13 | 1-acyl-sn-glycerol−3-phosphate acyltransferase-like protein |
| Araip.LLR5T | Arahy.H4YX61 | −0.36 | 0.51 | 0.99 | 0.66 | 0.73 | 0.8 | biotin carboxyl carrier acetyl-CoA carboxylase |
| Araip.H7632 | Arahy.Y66AIQ | 0.02 | −1.23 | −4.48 | −2.27 | −3.01 | −3.3 | 1-phosphatidylinositol phosphodiesterase-like protein |
| Aradu.JU9J9 | Arahy.4ZRU7U | −0.3 | 0.37 | 1.81 | −0.04 | 0.72 | 0.83 | trans-2-enoyl-CoA reductase, steroid 5-alpha-reductase |
| Aradu.54FFT | Arahy.DCBE83 | −0.99 | −1.1 | −6.54 | −11.5 | −2.45 | −3.88 | O-acyltransferase (WSD1-like) family protein |
| Araip.Z2NTP | Arahy.D8KQ2T | 4.91 | −1.86 | 0.36 | −4.05 | −1.73 | −1.61 | PATATIN-like protein 5 |
| Aradu.5S6RT | Arahy.UL6NUJ | −0.16 | 0.57 | 1.43 | 0.88 | 1.04 | 1.13 | 1-acyl-sn-glycerol−3-phosphate acyltransferase-like protein |
| Araip.4I8QR | Arahy.S0JXUX | −1.56 | −0.12 | 0.88 | 0.59 | 0.48 | 0.65 | biotin carboxyl carrier acetyl-CoA carboxylase |
| Araip.QK1BG | Arahy.Y1617F | −1.17 | −0.32 | −1.44 | −1.18 | −1.1 | −1.27 | phospholipase D P2 |
| Aradu.82M2C | Arahy.CC9CW3 | 0.82 | −1.03 | 2.01 | 1.27 | 1.1 | 1.45 | hydroxymethylglutaryl-CoA synthase-like |
| Araip.95KUY | Arahy.W7LA7X | −0.12 | −0.57 | −1.69 | −1.95 | −1.34 | −1.64 | patatin-like protein 6 |
| Aradu.J6P55 | Arahy.IFJ1V3 | −1.6 | −0.92 | 1.44 | −2.31 | −0.71 | −0.62 | 3-ketoacyl-CoA synthase 11 |
| Aradu.4R68R | Arahy.IBW8RK | −0.31 | 0.31 | 2.42 | −0.21 | 0.87 | 1.09 | sterol C4-methyl oxidase 1-2 |
| Aradu.Y6Q3B | Arahy.H1181I | −1.98 | −0.41 | 0.13 | −1.07 | −0.37 | −0.36 | fatty acid hydroxylase superfamily |
| Aradu.8Y5FN | Arahy.TAF7VU | −0.32 | 0.62 | 2.5 | 0.79 | 1.25 | 1.4 | niemann-pick C1-like protein |
| Aradu.32LC7 | Arahy.HNK90J | −0.38 | −1.75 | −2.19 | −3.58 | −2.33 | −2.64 | myo-inositol-1-phosphate synthase 3 |
| Aradu.07VYH | Arahy.JC4ZCG | −0.98 | 0.04 | 4.75 | −0.81 | 1.26 | 1.81 | 3-hydroxyacyl-[acyl-carrier-protein] dehydratase FabZ |
| Araip.VC3BC | Arahy.6JMM3R | −1.72 | −3.33 | −0.17 | −1.86 | −1.83 | −1.47 | lecithin:cholesterol acyltransferase family protein |
| Aradu.AV1HQ | Arahy.722ASC | −2.44 | −1.91 | −2.5 | −2.13 | −2.18 | −2.26 | fatty acid desaturase 8 |
| Aradu.PR2BP | Arahy.W5G5TD | 0.1 | −0.73 | −2.35 | −1.89 | −1.78 | −2.04 | fatty acid/sphingolipid desaturase, delta-6 desaturase |
| Aradu.V9VEN | Arahy.HXV4VT | −1.44 | 0.08 | −1.54 | −2.16 | −0.87 | −1.46 | phospholipid-transporting ATPase 9-like isoform X1 |
| Aradu.IJ0H9 | Arahy.D341M5 | −2 | −0.69 | −0.1 | 0.24 | −0.05 | 0.05 | probable LRR (leucine-rich repeat) receptor-like serine/threonine-protein kinase |
| Araip.V7FVT | Arahy.YVU30U | −0.02 | −0.52 | 1.8 | 0.36 | 0.63 | 0.95 | phosphatidylinositol−4-phosphate 5-kinase 1 |
| Araip.81A8L | Arahy.PKW5X9 | 1.95 | −1.24 | −0.76 | −2.52 | −1.48 | −1.65 | monogalactosyldiacylglycerol synthase 2 |
| Araip.T755Q | Arahy.NH00NZ | 0.36 | −3.44 | −2.36 | −2.09 | −2.43 | −2.31 | type I inositol 1,4,5-trisphosphate 5-phosphatase 11-like |
| Aradu.TI2CX | Arahy.6XGZ20 | 1.03 | 0.31 | 2.11 | −1.83 | 0.45 | 0.5 | alcohol dehydrogenase |
| Aradu.AMA0U | Arahy.IGY6G6 | −0.49 | −0.28 | 0.86 | −0.55 | 0.08 | 0.17 | LAG1 longevity assurance homolog 3, LAC1 |
| Araip.0E4ZE | Arahy.LFL6K6 | −0.83 | −1.04 | −1.5 | −0.94 | −1.21 | −1.24 | non-specific lipid-transfer protein |
| Aradu.CXJ5P | Arahy.XQ2AST | −2.21 | −1.55 | −0.69 | −2.23 | −1.56 | −1.56 | solanesyl diphosphate synthase 1 |
| Aradu.03ENG | Arahy.L0R0IL | −0.41 | −1.52 | −1.76 | −1.82 | −1.68 | −1.75 | non-specific lipid-transfer protein |
| Aradu.0L9GE | Arahy.BI8WMI | 0.35 | −0.85 | −2.34 | −2.68 | −1.74 | −2.2 | glycerol−3-phosphate acyltransferase |
| Araip.79E4D | Arahy.Z1TE7K | 0.21 | −1.23 | −1.17 | 0.18 | −0.7 | −0.63 | non-specific phospholipase C3 |
| Aradu.9G3ZE | Arahy.LLG5IK | 1.45 | −0.55 | −2.76 | −0.34 | −1.38 | −1.51 | alcohol dehydrogenase 1 |
| Araip.3B5FU | Arahy.R19W3K | −0.04 | 0.02 | −0.67 | 0.52 | −0.13 | −0.16 | lycopene cyclase, lycopene cyclase-type, FAD (flavin adenine dinucleotide) -binding |
| Araip.VA90H | Arahy.Z9XA8H | −1.65 | −0.34 | 1.35 | 0.15 | 0.4 | 0.64 | acyl carrier protein 4 |
| Araip.AT3RC | Arahy.68T27Q | −0.8 | −0.35 | −1.65 | −0.55 | −1.02 | −1.13 | Protein kinase superfamily protein |
| Araip.N4RJ5 | Arahy.68T27Q | −0.97 | −0.02 | −1.96 | −1.45 | −0.91 | −1.38 | putative phospholipid-transporting ATPase 9-like isoform X1 |
| Araip.L7Z6B | Arahy.PUH2BT | −0.91 | −0.39 | −0.66 | −0.1 | −0.38 | −0.38 | glycerophosphodiester phosphodiesterase GDE1 |
| Araip.P3CAI | Arahy.JGYC2J | −2.17 | −0.27 | −1 | −0.08 | −0.49 | −0.53 | GDSL-like lipase/acylhydrolase superfamily protein |
| Araip.TF5RD | Arahy.PY4INI | −0.42 | 0.93 | 2.05 | 0.31 | 1.09 | 1.12 | non-specific phospholipase C6 |
| Aradu.XM2MR | Arahy.E24IL6 | 1.53 | 2.06 | −1.42 | −1.1 | −3.71 | −2.54 | acyl-[acyl-carrier-protein] desaturase, SAD2 homologous gene |
| Araip.3S8HT | Arahy.05783X | 0.18 | −0.51 | −0.06 | −0.34 | −0.32 | −0.24 | 3-oxoacyl-[acyl-carrier-protein] synthase 3 FabH |
| Araip.CW7EC | Arahy.JEY2CC | 1.42 | −0.48 | −0.77 | −2.39 | −0.93 | −1.27 | polyketide cyclase/dehydrase and lipid transport superfamily protein |
| Aradu.CH0GR | Arahy.74RI4B | 1.71 | −0.51 | −0.68 | 0.56 | −0.22 | −0.2 | GDSL-like lipase/Acylhydrolase superfamily protein |
| Aradu.YH03P | Arahy.6FI8AN | 6.52 | −0.73 | −3.05 | −1.9 | −2.47 | −2.6 | alpha/beta-hydrolases superfamily protein |
| Araip.QM8AJ | Arahy.K79JM6 | −2.79 | −1.96 | −1.8 | −4.75 | −2.28 | −2.51 | glycerol−3-phosphate acyltransferase 2 |
| Araip.8V7F0 | Arahy.D341M5 | −1.45 | 0.25 | 0.77 | −0.12 | 0.27 | 0.28 | probable LRR receptor-like serine/threonine-protein kinase X3 |
| Araip.Q3H1S | Arahy.FH5Z1U | 0.31 | −3.42 | −0.43 | −2.38 | −1.94 | −1.47 | lipid transfer protein |
| Aradu.13SFN | Arahy.ZE5M4X | 1.18 | 0.57 | 1.49 | 2.47 | 1.8 | 1.91 | GDSL-like lipase/acylhydrolase superfamily protein |
| Araip.Q7M7G | Arahy.BI8WMI | 0.41 | −0.83 | −2.16 | −2.81 | −1.68 | −2.14 | glycerol−3-phosphate acyltransferase 2C chloroplastic-like isoform X2 |
| Araip.R99KE | Arahy.6H2079 | 0.39 | 1.42 | 0.87 | 1.18 | 1.13 | 1.05 | uncharacterized protein LOC100792273 isoform X2 |
| Aradu.B4FEF | Arahy.TZMQ1W | 1.54 | −0.54 | −1.37 | −1.84 | −1.04 | −1.37 | Polyketide cyclase/dehydrase and lipid transport superfamily protein |
| Aradu.8019V | Arahy.6WS8RX | 5.83 | 0.22 | −2.99 | −0.68 | −1.86 | −1.95 | GDSL-like lipase/acylhydrolase |
| Aradu.L86SE | Arahy.PKW5X9 | 2.29 | −1.4 | −0.92 | −2.62 | −1.64 | −1.81 | monogalactosyldiacylglycerol synthase 2 |
| Araip.S3GXY | Arahy.5913QL | −1.88 | −1.39 | 0.06 | −2.55 | −0.78 | −0.65 | fatty acid desaturase 2 |
| Aradu.UL8XP | Arahy.Q6L6L9 | −1.09 | −1.36 | −1.61 | −0.54 | −1.19 | −1.17 | 3-hydroxy−3-methylglutaryl-coenzyme A reductase-like protein |
| Araip.5UJ5P | Arahy.3WFA2L | 1.94 | 0.29 | 0.04 | 0.55 | 0.32 | 0.33 | long-chain acyl-CoA synthetase 2 |
| Aradu.TA7RE | Arahy.F8M8GF | 6.23 | 7.03 | 6.94 | 7.58 | 7.22 | 7.26 | Pleckstrin homology (PH) domain-containing protein |
| Araip.31Q5V | Arahy.GU7MJ6 | −2.11 | −1.23 | −3.16 | −0.87 | −1.68 | −1.81 | fatty acid desaturase 5, plant stearoyl-acyl-carrier protein |
| Araip.K56RN | Arahy.NDAY03 | 0.61 | −0.79 | −0.54 | −0.81 | −0.72 | −0.71 | seed linoleate 9S-lipoxygenase |
| Aradu.H0G14 | Arahy.5A3JDR | −2.32 | 1.93 | −2.92 | −1.34 | −0.98 | −1.66 | 12-oxophytodienoate reductase 1 |
| Aradu.AS232 | Arahy.7CJ2RM | 0.07 | −1.84 | −2.3 | −1.31 | −1.87 | −1.87 | lipoxygenase 3 |
| Araip.220UP | Arahy.K697CU | −2.13 | −0.02 | −0.4 | 0.42 | −0.02 | −0.02 | acyl-CoA-binding domain 3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Gu, J.; Lu, Q.; Li, H.; Hong, Y.; Chen, X.; Ren, L.; Deng, L.; Liang, X. Transcriptomic Analysis Reveals the High-Oleic Acid Feedback Regulating the Homologous Gene Expression of Stearoyl-ACP Desaturase 2 (SAD2) in Peanuts. Int. J. Mol. Sci. 2019, 20, 3091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20123091
Liu H, Gu J, Lu Q, Li H, Hong Y, Chen X, Ren L, Deng L, Liang X. Transcriptomic Analysis Reveals the High-Oleic Acid Feedback Regulating the Homologous Gene Expression of Stearoyl-ACP Desaturase 2 (SAD2) in Peanuts. International Journal of Molecular Sciences. 2019; 20(12):3091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20123091
Chicago/Turabian StyleLiu, Hao, Jianzhong Gu, Qing Lu, Haifen Li, Yanbin Hong, Xiaoping Chen, Li Ren, Li Deng, and Xuanqiang Liang. 2019. "Transcriptomic Analysis Reveals the High-Oleic Acid Feedback Regulating the Homologous Gene Expression of Stearoyl-ACP Desaturase 2 (SAD2) in Peanuts" International Journal of Molecular Sciences 20, no. 12: 3091. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20123091