Protective Effects of Salidroside against Carbon Tetrachloride (CCl4)-Induced Liver Injury by Initiating Mitochondria to Resist Oxidative Stress in Mice


 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Salidroside Antagonized CCl4-Induced Hepatotoxicity in Mice
2.2. Salidroside Balanced Antioxidant System against Oxidative Stress
2.3. Salidroside Protected Mitochondria
2.4. Mitochondria Decreased ROS Generation with Salidroside
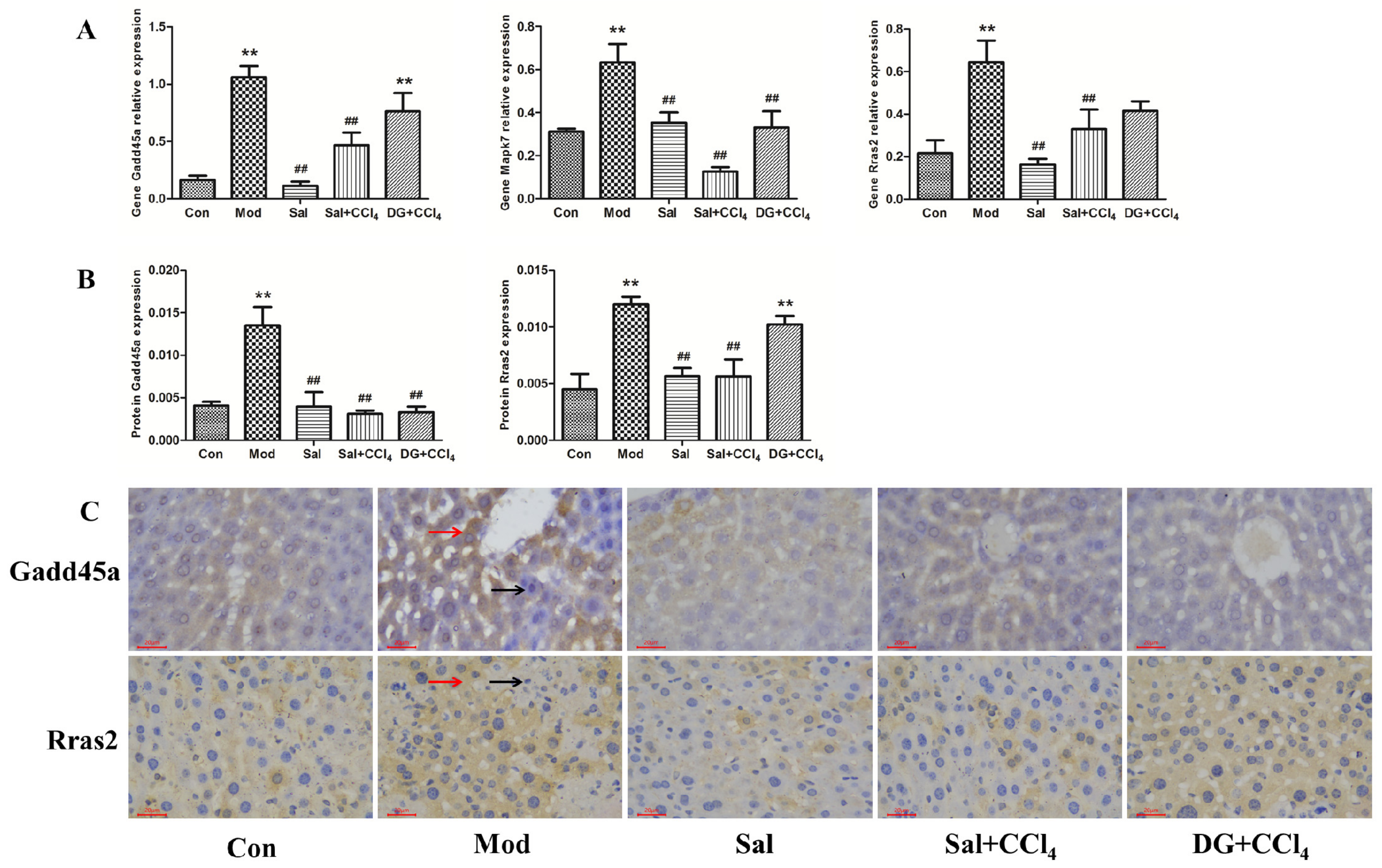
2.5. Oxidative Stress-Related Genes Gadd45a, Mapk7, and Rras2 Were Involved in Protective Effects of Salidroside against Liver Injury
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Chemicals
4.3. Experimental Procedure
4.4. Organ Index and Serum Biochemistry
4.5. Pathologic Histology Observation
4.6. Liver Antioxidant-related Biochemistry
4.7. ROS and MMP
4.8. Ultrastructure Observations
4.9. Fluorescence Real-Time Quantification PCR (QRT-PCR)
4.10. Immunohistochemistry
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lv, C.; Huang, Y.; Liu, Z.X.; Yu, D.; Bai, Z.M. Salidroside reduces renal cell carcinoma proliferation by inhibiting JAK2/STAT3 signaling. Cancer Biomark. 2016, 17, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.R.; Wang, H.F.; Zuo, T.C.; Guan, L.L.; Dai, N. Salidroside alleviates oxidative stress in the liver with non-alcoholic steatohepatitis in rats. BMC Pharmacol. Toxicol. 2016, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Dhar, P.; Bajpai, P.K.; Tayade, A.B.; Chaurasia, O.P.; Srivastava, R.B.; Singh, S.B. Chemical composition and antioxidant capacities of phytococktail extracts from trans-Himalayan cold desert. BMC Complement. Altern. Med. 2013, 13, 259. [Google Scholar] [CrossRef]
- Feng, J.; Niu, P.; Chen, K.; Wu, L.; Liu, T.; Xu, S.; Li, J.; Li, S.; Wang, W.; Lu, X. Salidroside mediates apoptosis and autophagy inhibition in concanavalin A-induced liver injury. Exp. Ther. Med. 2018, 15, 4599–4614. [Google Scholar] [CrossRef]
- Yuan, Y.; Wu, S.J.; Liu, X.; Zhang, L.L. Antioxidant effect of salidroside and its protective effect against furan-induced hepatocyte damage in mice. Food Funct. 2013, 4, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Hu, C.; Meehan, E.J.; Chen, L. X-ray crystal structure and antioxidant activity of salidroside, a phenylethanoid glycoside. Chem. Biodivers. 2007, 4, 508–513. [Google Scholar] [CrossRef]
- Zhang, X.R.; Fu, X.J.; Zhu, D.S.; Zhang, C.Z.; Hou, S.; Li, M.; Yang, X.H. Salidroside-regulated lipid metabolism with down-regulation of miR-370 in type 2 diabetic mice. Eur. J. Pharmacol. 2016, 779, 46–52. [Google Scholar] [CrossRef]
- Tao, Z.; Yang, X.Y.; Li, W.J.; Wang, Q.B.; Li, C.; Dan, W.; Fang, B.; Xing, S.S.; Si, J. Salidroside Attenuates High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease via AMPK-Dependent TXNIP/NLRP3 Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Piao, D.M.; Han, X.H.; Nan, J.X. Protective effects of salidroside against acetaminophen-induced toxicity in mice. Biol. Pharm. Bull. 2008, 31, 1523–1529. [Google Scholar] [CrossRef]
- Wu, Y.L.; Lian, L.H.; Jiang, Y.Z.; Nan, J.-X. Hepatoprotective effects of salidroside on fulminant hepatic failure induced by D-galactosamine and lipopolysaccharide in mice. J. Pharm. Pharmacol. 2009, 61, 1375–1382. [Google Scholar] [CrossRef]
- Slater, T.F.; Cheeseman, K.H.; Ingold, K.U.; Rice-Evans, C.; Sies, H. Carbon tetrachloride toxicity as a model for studying free-radical mediated liver injury. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1985, 311, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gong, X.; Jiang, R.; Wang, B.; Kuang, G.; Li, K.; Wan, J. Resolvin D1 attenuates CCl4-induced acute liver injury involving up-regulation of HO-1 in mice. Immunopharmacol. Immunotoxicol. 2015, 38, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Hafez, M.M.; Al-Shabanah, O.A.; Al-Harbi, N.O.; Al-Harbi, M.M.; Al-Rejaie, S.S.; Alsurayea, S.M.; Sayed-Ahmed, M.M. Association between paraoxonases gene expression and oxidative stress in hepatotoxicity induced by CCl4. Oxid. Med. Cell. Longev. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Šiviková, K.N.; Piešová, E.; Dianovský, J. The protection of Vitamin E and selenium against carbon tetrachloride-induced genotoxicity in ovine peripheral blood lymphocytes. Mutat. Res. 2001, 494, 135–142. [Google Scholar] [CrossRef]
- Shi, J.; Aisaki, K.; Ikawa, Y.; Wake, K. Evidence of Hepatocyte Apoptosis in Rat Liver after the Administration of Carbon Tetrachloride. Am. J. Pathol. 1998, 153, 515–525. [Google Scholar] [CrossRef] [Green Version]
- Tsai, W.-H.; Yang, C.-C.; Li, P.-C.; Chen, W.-C.; Chien, C.-T.; Wh, T.; Cc, Y.; Pc, L.; Wc, C.; Ct, C. Therapeutic Potential of Traditional Chinese Medicine on Inflammatory Diseases. J. Tradit. Complement. Med. 2013, 3, 142–151. [Google Scholar] [CrossRef] [Green Version]
- LaHair, M.M.; Franklin, R.A.; McCubrey, J.A.; McCubrey, J.A.; Mc Cubrey, J. Reactive oxygen species-induced activation of the MAP kinase signaling pathways. Antioxid. Redox Signal. 2006, 8, 1775–1789. [Google Scholar]
- Yang, J.; Zhou, L.; Wang, J.; Wang, G.; Davey, A.K. The disposition of diammonium glycyrrhizinate and glycyrrhetinic acid in the isolated perfused rat intestine and liver. Planta Med. 2008, 74, 1351–1356. [Google Scholar] [CrossRef]
- Wallace, D.C. A Mitochondrial Paradigm of Metabolic and Degenerative Diseases, Aging, and Cancer: A Dawn for Evolutionary Medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [Green Version]
- Hock, M.B.; Kralli, A. Transcriptional control of mitochondrial biogenesis and function. Annu. Rev. Physiol. 2009, 71, 177–203. [Google Scholar] [CrossRef]
- Greaves, L.C.; Reeve, A.K.; Taylor, R.W.; Turnbull, D.M. Mitochondrial DNA and disease. J. Pathol. 2011, 226, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xia, H.; Liu, Y.; Qiu, F.; Di, X. Simultaneous determination of three glucuronide conjugates of scutellarein in rat plasma by LC–MS/MS for pharmacokinetic study of breviscapine. J. Chromatogr. B 2014, 965, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y. Effects of salidroside on exhaustive exercise-induced oxidative stress in rats. Mol. Med. Rep. 2012, 6, 1195–1198. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C.; Lee, F.T.; Kuo, T.Y.; Yang, J.H.; Chien, C.T. Attenuation of long-term Rhodiola rosea supplementation on exhaustive swimming-evoked oxidative stress in the rat. Chin. J. Physiol. 2009, 52, 9. [Google Scholar] [CrossRef]
- Sookoian, S.; Pirola, C.J. Liver enzymes, metabolomics and genome-wide association studies: From systems biology to the personalized medicine. World J. Gastroenterol. 2015, 21, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Que, S.; Xu, J.; Peng, T. Alanine aminotransferase-old biomarker and new concept: A review. Int. J. Med. Sci. 2014, 11, 925. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Chen, K.; Xia, Y.; Wu, L.; Li, J.; Li, S.; Wang, W.; Lu, X.; Tong, L.; Guo, C. Salidroside ameliorates autophagy and activation of hepatic stellate cells in mice via NF-κB and TGF-β1/Smad3 pathways. Drug Des. Dev. Ther. 2018, 12, 1837–1853. [Google Scholar] [CrossRef]
- Slater, T.F. Necrogenic action of carbon tetrachloride in the rat: a speculative mechanism based on activation. Nature 1966, 209, 36–40. [Google Scholar] [CrossRef]
- Zorov, D.B.; Filburn, C.R.; Klotz, L.O.; Zweier, J.L.; Sollott, S.J. Reactive oxygen species (ROS)-induced ROS release: A new phenomenon accompanying induction of the mitochondrial permeability transition in cardiac myocytes. J. Exp. Med. 2000, 192, 1001–1014. [Google Scholar] [CrossRef]
- Lee, J.; Giordano, S.; Zhang, J. Autophagy, mitochondria and oxidative stress: cross-talk and redox signalling. Biochem. J. 2012, 441, 523–540. [Google Scholar] [CrossRef] [PubMed]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: the mitochondria-dependent and;mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef] [PubMed]
- Ulicná, O.; Greksák, M.; Vancová, O.; Zlatos, L.; Galbavý, S.; Bozek, P.; Nakano, M. Hepatoprotective effect of rooibos tea (Aspalathus linearis) on CCl4-induced liver damage in rats. Physiol. Res. 2003, 52, 461–466. [Google Scholar] [PubMed]
- Harrison, D.G.; Cai, H.; Landmesser, U.; Griendling, K.K. Interactions of angiotensin II with NAD(P)H oxidase, oxidant stress and cardiovascular disease. J. Renin Angiotensin Aldosterone Syst. 2003, 4, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Sauer, H.; Wartenberg, M.; Hescheler, J. Reactive oxygen species as intracellular messengers during cell growth and differentiation. Cell. Physiol. Biochem. 2001, 11, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.J.; Huang, Y.; Singleton, P.A.; Sammani, S.; Moitra, J.; Evenoski, C.L.; Husain, A.N.; Mitra, S.; Moreno-Vinasco, L.; Jacobson, J.R.; et al. GADD45a is a novel candidate gene in inflammatory lung injury via influences on Akt signaling. FASEB J. 2009, 23, 1325–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patterson, A.D.; Hildesheim, J.; Fornace, A.J., Jr.; Hollander, M.C. Neural tube development requires the cooperation of p53- and Gadd45a-associated pathways. Birth Defects Res. Part A Clin. Mol. Teratol. 2006, 76, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Na, Z.; Yan, S.; Lin, X.; Long, Y.; Luhong, S. Gadd45-alpha and Gadd45-gamma utilize p38 and JNK signaling pathways to induce cell cycle G2/M arrest in Hep-G2 hepatoma cells. Mol. Biol. Rep. 2009, 36, 2075. [Google Scholar]
- Sadiye Amcaoglu, R.; Prakash, N.; Mitzi, N. Multiple anti-inflammatory pathways triggered by resveratrol lead to amelioration of staphylococcal enterotoxin B-induced lung injury. Br. J. Pharmacol. 2012, 167, 1244–1258. [Google Scholar] [Green Version]
- Bulavin, D.V.; Demidov, O.N.; Shin’Ichi, S.; Paivikki, K.; Crissy, P.; Amundson, S.A.; Concetta, A.; Guido, S.; Nebreda, A.R.; Anderson, C.W. Amplification of PPM1D in human tumors abrogates p53 tumor-suppressor activity. Nat. Genet. 2002, 31, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Abe, J.; Kusuhara, M.; Ulevitch, R.J.; Berk, B.C.; Lee, J.D. Big mitogen-activated protein kinase 1 (BMK1) is a redox-sensitive kinase. J. Biol. Chem. 1996, 271, 16586–16590. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Vater, C.; Jacobi, A.; Liebers, C.; Zou, X.; Stiehler, M. Salidroside exerts angiogenic and cytoprotective effects on human bone marrow-derived endothelial progenitor cells via Akt/mTOR/p70S6K and MAPK signalling pathways. Br. J. Pharmacol. 2014, 171, 2440–2456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Y.; Sun, Z.N.; Wang, M.X.; Zhang, C. SIRT1 mediates salidroside-elicited protective effects against MPP(+) -induced apoptosis and oxidative stress in SH-SY5Y cells: Involvement in suppressing MAPK pathways. Cell Biology International 2017, 42, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Li, Y.; Li, W.; Guo, R. Salidroside protects against foam cell formation and apoptosis, possibly via the MAPK and AKT signaling pathways. Lipids Health Dis. 2017, 16, 198. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.L.; Li, Y.H.; Zhang, Q.Q. Salidroside protects rat liver against ischemia/reperfusion injury by regulating the GSK-3β/Nrf2-dependent antioxidant response and mitochondrial permeability transition. Eur. J. Pharmacol. 2017, 806, 32–42. [Google Scholar] [CrossRef]
- Feng, C.; Wang, H.X.; Yao, C.F.; Zhang, J.; Tian, Z.G. Diammonium glycyrrhizinate, a component of traditional Chinese medicine Gan-Cao, prevents murine T-cell-mediated fulminant hepatitis in IL-10- and IL-6-dependent manners. Int. Immunopharmacol. 2007, 7, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Xie, Y.M.; Yang, W.; Wang, Y.Y.; Yi, D.H.; Zhuang, Y. Analysis of characteristics of traditional Chinese medicine and western medicine clinical use in patients with viral hepatitis based on real world hospital information system data. China J. Chin. Mater. Med. 2014, 39, 3535–3540. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primers, 5′–3′ | Reverse Primers, 5′–3′ |
|---|---|---|
| β-Actin | GCCCTGAGGCTCTTTTCCA | GTTGGCATAGAGGTCTTTACGGAT |
| Gadd45a | GCAGAGCAGAAGACCGAAAG | TAACAGAACGCACGGATGAG |
| Mapk7 | TGTGACCAATGCCAAACGG | GCGGCTGTGAAGAGTGAATGA |
| Rras2 | CAGAGTAAAGGACCGTGATGAG | TGGTTCTGGTGAAGGAGGG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.-Y.; Xu, D.; Du, X.-X.; Ran, C.-L.; Xu, L.; Ren, S.-J.; Tang, Z.-T.; Yin, L.-Z.; He, C.-L.; Yuan, Z.-X.; et al. Protective Effects of Salidroside against Carbon Tetrachloride (CCl4)-Induced Liver Injury by Initiating Mitochondria to Resist Oxidative Stress in Mice. Int. J. Mol. Sci. 2019, 20, 3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133187
Lin S-Y, Xu D, Du X-X, Ran C-L, Xu L, Ren S-J, Tang Z-T, Yin L-Z, He C-L, Yuan Z-X, et al. Protective Effects of Salidroside against Carbon Tetrachloride (CCl4)-Induced Liver Injury by Initiating Mitochondria to Resist Oxidative Stress in Mice. International Journal of Molecular Sciences. 2019; 20(13):3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133187
Chicago/Turabian StyleLin, Shi-Yu, Dan Xu, Xia-Xia Du, Chong-Lin Ran, Lu Xu, Shao-Jun Ren, Zi-Ting Tang, Li-Zi Yin, Chang-Liang He, Zhi-Xiang Yuan, and et al. 2019. "Protective Effects of Salidroside against Carbon Tetrachloride (CCl4)-Induced Liver Injury by Initiating Mitochondria to Resist Oxidative Stress in Mice" International Journal of Molecular Sciences 20, no. 13: 3187. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20133187