14-3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease


Abstract
:1. Introduction
2. The Structure of 14-3-3 Proteins
3. 14-3-3 Proteins Have Consistent Expression Patterns in Aging and Cancer
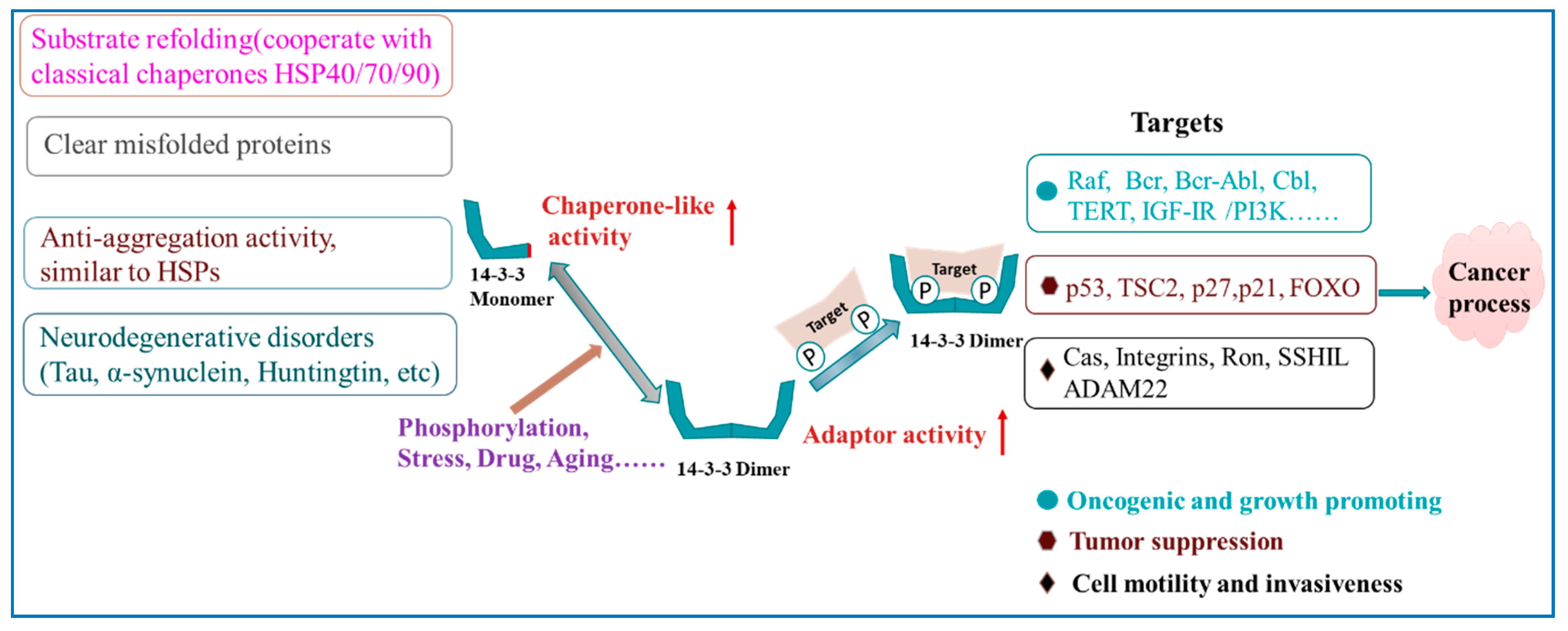
3.1. Cancer
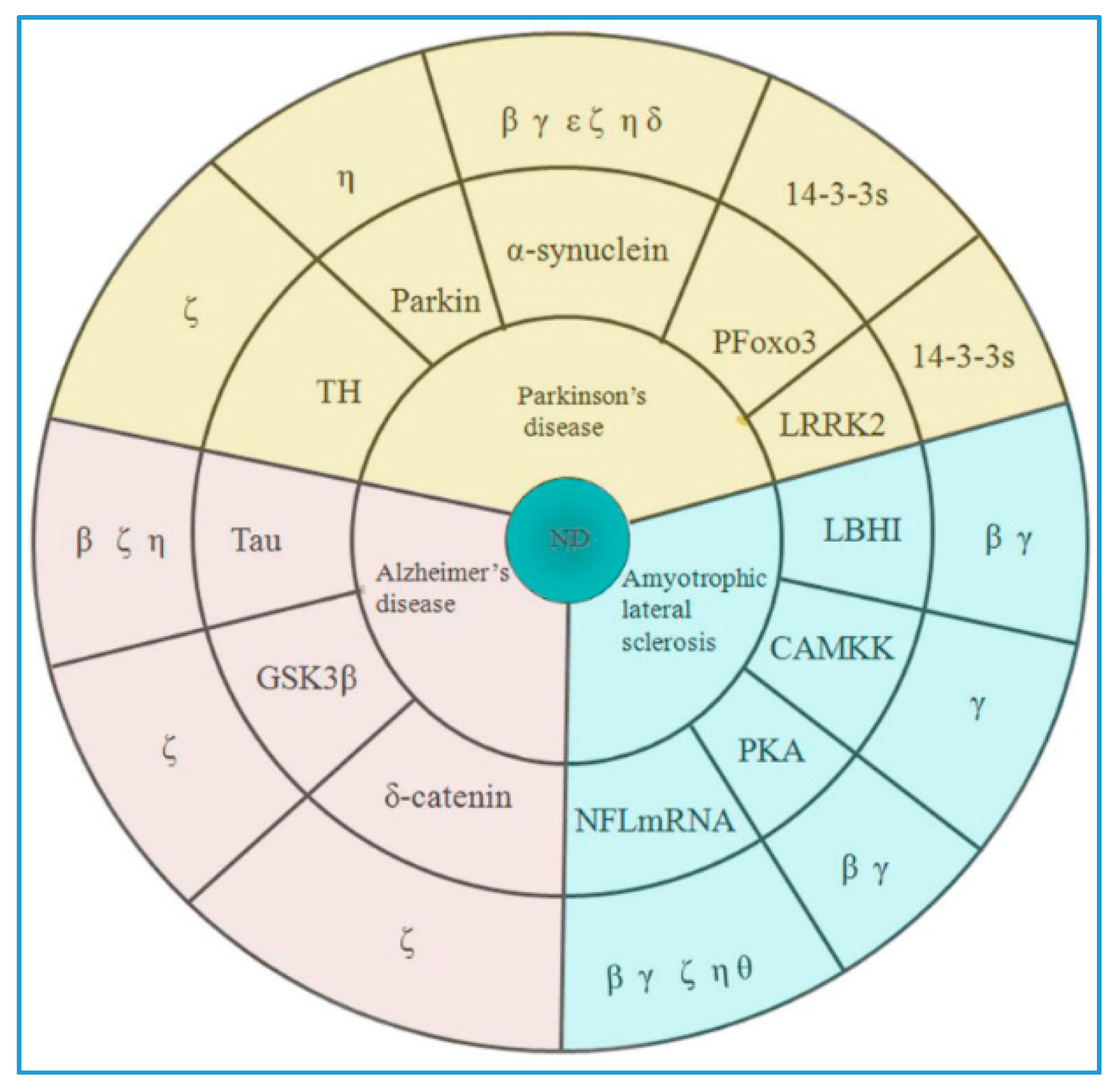
3.2. Age-Related Neurodegenerative Disease
3.2.1. Parkinson’s Disease
3.2.2. Alzheimer’s Disease
4. Aging Process
5. Conclusions and Challenges
Author Contributions
Funding
Conflicts of Interest
References
- Jackaman, C.; Tomay, F.; Duong, L.; Abdol Razak, N.B.; Pixley, F.J.; Metharom, P.; Nelson, D.J. Aging and cancer: The role of macrophages and neutrophils. Ageing Res. Rev. 2017, 36, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Niccoli, T.; Partridge, L. Ageing as a risk factor for disease. Curr. Biol. 2012, 22, R741–R752. [Google Scholar] [CrossRef] [PubMed]
- Bottazzi, B.; Riboli, E.; Mantovani, A. Aging, inflammation and cancer. Semin. Immunol. 2018, 40, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Aramillo, P.; Schauble, S.; Esser, D.; Groth, M.; Frahm, C.; Priebe, S.; Baumgart, M.; Hartmann, N.; Marthandan, S.; Menzel, U.; et al. Transcriptomic alterations during ageing reflect the shift from cancer to degenerative diseases in the elderly. Nat. Commun. 2018, 9, 327. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S. Cellular senescence, aging, and age-related disease: Special issue of BMB Reports in 2019. BMB Rep. 2019, 52, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Hartley, A.V.; Martin, M.; Lu, T. Aging: Cancer - an unlikely couple. Aging (Albany NY). 2017, 9, 1949–1950. [Google Scholar] [CrossRef]
- Meynet, O.; Ricci, J.E. Caloric restriction and cancer: Molecular mechanisms and clinical implications. Trends Mol. Med. 2014, 20, 419–427. [Google Scholar] [CrossRef]
- Zhu, Y.; Tchkonia, T.; Pirtskhalava, T.; Gower, A.C.; Ding, H.; Giorgadze, N.; Palmer, A.K.; Ikeno, Y.; Hubbard, G.B.; Lenburg, M.; et al. The Achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell. 2015, 14, 644–658. [Google Scholar] [CrossRef]
- Short, S.; Fielder, E.; Miwa, S.; von Zglinicki, T. Senolytics and senostatics as adjuvant tumour therapy. EBio Med. 2019, 41, 683–692. [Google Scholar] [Green Version]
- Aunan, J.R.; Cho, W.C.; Soreide, K. The Biology of Aging and Cancer: A Brief Overview of Shared and Divergent Molecular Hallmarks. Aging Dis. 2017, 8, 628–642. [Google Scholar] [CrossRef] [Green Version]
- van Heusden, G.P. 14-3-3 proteins: Regulators of numerous eukaryotic proteins. IUBMB Life. 2005, 57, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Pozuelo-Rubio, M. Proteomic and biochemical analysis of 14-3-3-binding proteins during C2-ceramide-induced apoptosis. FEBS J. 2010, 277, 3321–3342. [Google Scholar] [CrossRef] [PubMed]
- Pozuelo Rubio, M.; Geraghty, K.M.; Wong, B.H.; Wood, N.T.; Campbell, D.G.; Morrice, N.; Mackintosh, C. 14-3-3-affinity purification of over 200 human phosphoproteins reveals new links to regulation of cellular metabolism, proliferation and trafficking. Biochem. J. 2004, 379, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Cornell, B.; Toyo-Oka, K. 14-3-3 Proteins in Brain Development: Neurogenesis, Neuronal Migration and Neuromorphogenesis. Front Mol. Neurosci. 2017, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Skoulakis, E.C.; Acevedo, S.F. 14-3-3s are potential biomarkers for HIV-related neurodegeneration. J. Neurovirol. 2012, 18, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, A.; Ottmann, C.; Fournier, A.E. 14-3-3 adaptor protein-protein interactions as therapeutic targets for CNS diseases. Pharmacol. Res. 2017, 125, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.; Gusev, N.B. Moonlighting chaperone-like activity of the universal regulatory 14-3-3 proteins. FEBS J. 2017, 284, 1279–1295. [Google Scholar] [CrossRef] [PubMed]
- Aghazadeh, Y.; Papadopoulos, V. The role of the 14-3-3 protein family in health, disease, and drug development. Drug Discov. Today. 2016, 21, 278–287. [Google Scholar] [CrossRef]
- Morrison, D.K. The 14-3-3 proteins: Integrators of diverse signaling cues that impact cell fate and cancer development. Trends Cell Biol. 2009, 19, 16–23. [Google Scholar] [CrossRef]
- Tzivion, G.; Gupta, V.S.; Kaplun, L.; Balan, V. 14-3-3 proteins as potential oncogenes. Semin Cancer Biol. 2006, 16, 203–213. [Google Scholar] [CrossRef]
- Diallo, K.; Oppong, A.K.; Lim, G.E. Can 14-3-3 proteins serve as therapeutic targets for the treatment of metabolic diseases? Pharmacol Res. 2019, 139, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Fournier, A.E.; Yamagata, K. Neuroprotective function of 14-3-3 proteins in neurodegeneration. BioMed Res. Int. 2013, 2013, 564534. [Google Scholar] [CrossRef]
- Yacoubian, T.A.; Slone, S.R.; Harrington, A.J.; Hamamichi, S.; Schieltz, J.M.; Caldwell, K.A.; Caldwell, G.A.; Standaert, D.G. Differential neuroprotective effects of 14-3-3 proteins in models of Parkinson’s disease. Cell Death Dis. 2010, 1, e2. [Google Scholar] [CrossRef] [PubMed]
- Hermeking, H. 14-3-3 proteins and cancer biology. Semin Cancer Biol. 2006, 16, 161. [Google Scholar] [CrossRef]
- Muslin, A.J.; Xing, H. 14-3-3 proteins: Regulation of subcellular localization by molecular interference. Cell Signal. 2000, 12, 703–709. [Google Scholar] [CrossRef]
- Matta, A.; Siu, K.W.; Ralhan, R. 14-3-3 zeta as novel molecular target for cancer therapy. Expert Opin. Ther. Targets 2012, 16, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Macha, M.A.; Matta, A.; Chauhan, S.; Siu, K.M.; Ralhan, R. 14-3-3 zeta is a molecular target in guggulsterone induced apoptosis in head and neck cancer cells. BMC Cancer 2010, 10, 655. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, S.; Li, N.; Guo, W.; Shi, J.; Yu, H.; Zhang, L.; Wang, K.; Liu, S.; Cheng, S. 14-3-3ζ promotes hepatocellular carcinoma venous metastasis by modulating hypoxia-inducible factor-1α. Oncotarget 2016, 7, 15854–15867. [Google Scholar] [PubMed]
- Nishimura, Y.; Komatsu, S.; Ichikawa, D.; Nagata, H.; Hirajima, S.; Takeshita, H.; Kawaguchi, T.; Arita, T.; Konishi, H.; Kashimoto, K.; et al. Overexpression of YWHAZ relates to tumor cell proliferation and malignant outcome of gastric carcinoma. Br. J. Cancer 2013, 108, 1324–1331. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Wu, X.; Liu, X.; Yan, G.; Xu, C. LC-MS/MS analysis of ovarian cancer metastasis-related proteins using a nude mouse model: 14-3-3 zeta as a candidate biomarker. J. Proteome Res. 2010, 9, 6180–6190. [Google Scholar] [CrossRef]
- Janssen, J.J.; Klaver, S.M.; Waisfisz, Q.; Pasterkamp, G.; de Kleijn, D.P.; Schuurhuis, G.J.; Ossenkoppele, G.J. Identification of genes potentially involved in disease transformation of CML. Leukemia 2005, 19, 998–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Liu, J.Y.; Zhang, J.T. 14-3-3sigma, the double-edged sword of human cancers. Am. J. Transl. Res. 2009, 1, 326–340. [Google Scholar] [PubMed]
- Zhou, R.; Shao, Z.; Liu, J.; Zhan, W.; Gao, Q.; Pan, Z.; Wu, L.; Xu, L.; Ding, Y.; Zhao, L. COPS5 and LASP1 synergistically interact to downregulate 14-3-3sigma expression and promote colorectal cancer progression via activating PI3K/AKT pathway. Int. J. Cancer 2018, 142, 1853–1864. [Google Scholar] [CrossRef]
- Peng, C.; Jia, X.; Xiong, Y.; Yin, J.; Li, N.; Deng, Y.; Luo, K.; Zhang, Q.; Wang, C.; Zhang, Z.; et al. The 14-3-3 sigma/GSK3beta/beta-catenin/ZEB1 regulatory loop modulates chemo-sensitivity in human tongue cancer. Oncotarget 2015, 6, 20177–20189. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.J.; Wang, M.; Liu, R.M.; Wei, H.; Chao, W.X.; Zhang, T.; Lou, Q.; Li, X.M.; Ma, J.; Zhu, H.; et al. Downregulation of 14-3-3sigma correlates with multistage carcinogenesis and poor prognosis of esophageal squamous cell carcinoma. PLoS ONE 2014, 9, e95386. [Google Scholar]
- Lodygin, D.; Yazdi, A.S.; Sander, C.A.; Herzinger, T.; Hermeking, H. Analysis of 14-3-3sigma expression in hyperproliferative skin diseases reveals selective loss associated with CpG-methylation in basal cell carcinoma. Oncogene 2003, 22, 5519–5524. [Google Scholar] [CrossRef] [PubMed]
- Neupane, D.; Korc, M. 14-3-3sigma Modulates pancreatic cancer cell survival and invasiveness. Clin. Cancer Res. 2008, 14, 7614–7623. [Google Scholar] [CrossRef]
- Liu, C.C.; Jan, Y.J.; Ko, B.S.; Wu, Y.M.; Liang, S.M.; Chen, S.C.; Lee, Y.M.; Liu, T.A.; Chang, T.C.; Wang, J.; et al. 14-3-3sigma induces heat shock protein 70 expression in hepatocellular carcinoma. BMC Cancer 2014, 14, 425. [Google Scholar] [CrossRef]
- Raungrut, P.; Wongkotsila, A.; Lirdprapamongkol, K.; Svasti, J.; Geater, S.L.; Phukaoloun, M.; Suwiwat, S.; Thongsuksai, P. Prognostic significance of 14-3-3gamma overexpression in advanced non-small cell lung cancer. Asian Pac. J. Cancer Prev. 2014, 15, 3513–3518. [Google Scholar] [CrossRef]
- Yang, X.; Cao, W.; Lin, H.; Zhang, W.; Lin, W.; Cao, L.; Zhen, H.; Huo, J.; Zhang, X. Isoform-specific expression of 14-3-3 proteins in human astrocytoma. J. Neurol. Sci. 2009, 276, 54–59. [Google Scholar] [CrossRef]
- Liang, S.; Shen, G.; Liu, Q.; Xu, Y.; Zhou, L.; Xiao, S.; Xu, Z.; Gong, F.; You, C.; Wei, Y. Isoform-specific expression and characterization of 14-3-3 proteins in human glioma tissues discovered by stable isotope labeling with amino acids in cell culture-based proteomic analysis. Proteomics Clin. Appl. 2009, 3, 743–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Nesland, J.M.; Suo, Z.; Trope, C.G.; Holm, R. The prognostic value of 14-3-3 isoforms in vulvar squamous cell carcinoma cases: 14-3-3beta and epsilon are independent prognostic factors for these tumors. PLoS ONE 2011, 6, e24843. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Xu, Y.; Shen, G.; Liu, Q.; Zhao, X.; Xu, Z.; Xie, X.; Gong, F.; Li, R.; Wei, Y. Quantitative protein expression profiling of 14-3-3 isoforms in human renal carcinoma shows 14-3-3 epsilon is involved in limitedly increasing renal cell proliferation. Electrophoresis 2009, 30, 4152–4162. [Google Scholar] [CrossRef] [PubMed]
- Ko, B.S.; Chang, T.C.; Hsu, C.; Chen, Y.C.; Shen, T.L.; Chen, S.C.; Wang, J.; Wu, K.K.; Jan, Y.J.; Liou, J.Y. Overexpression of 14-3-3epsilon predicts tumour metastasis and poor survival in hepatocellular carcinoma. Histopathology 2011, 58, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Qi, W.; Liu, X.; Qiao, D.; Martinez, J.D. Isoform-specific expression of 14-3-3 proteins in human lung cancer tissues. Int. J. Cancer 2005, 113, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yan, L.; Gu, H.; Mu, Y.; Tong, G.; Zhang, G. 14-3-3ε functions as an oncogene in SGC7901 gastric cancer cells through involvement of cyclin E and p27kip1. Mol. Med. Rep. 2014, 10, 3145–3150. [Google Scholar] [CrossRef]
- Ko, B.S.; Lai, I.R.; Chang, T.C.; Liu, T.A.; Chen, S.C.; Wang, J.; Jan, Y.J.; Liou, J.Y. Involvement of 14-3-3γ overexpression in extrahepatic metastasis of hepatocellular carcinoma. Hum. Pathol. 2011, 42, 129–135. [Google Scholar] [CrossRef]
- Hiraoka, E.; Mimae, T.; Ito, M.; Kadoya, T.; Miyata, Y.; Ito, A.; Okada, M. Breast cancer cell motility is promoted by 14-3-3γ. Breast Cancer 2019. [Google Scholar] [CrossRef]
- Qi, W.; Liu, X.; Chen, W.; Li, Q.; Martinez, J.D. Overexpression of 14-3-3gamma causes polyploidization in H322 lung cancer cells. Mol. Carcinog. 2007, 46, 847–856. [Google Scholar] [CrossRef]
- Titus, M.A.; Tan, J.A.; Gregory, C.W.; Ford, O.H.; Subramanian, R.R.; Fu, H.; Wilson, E.M.; Mohler, J.L.; French, F.S. 14-3-3{eta} amplifies androgen receptor actions in prostate cancer. Clin. Cancer Res.: An Off. J. Am. Assoc. Cancer Res. 2009, 15, 7571–7581. [Google Scholar] [CrossRef]
- Li, X.H.; Noguchi, A.; Nishida, T.; Takahashi, H.; Zheng, Y.; Yang, X.H.; Masuda, S.; Kikuchi, K.; Takano, Y. Cytoplasmic expression of p33ING1b is correlated with tumorigenesis and progression of head and neck squamous cell carcinoma. Histol. Histopathol. 2011, 26, 597–607. [Google Scholar] [PubMed]
- Park, G.Y.; Han, J.Y.; Han, Y.K.; Kim, S.D.; Kim, J.S.; Jo, W.S.; Chun, S.H.; Jeong, D.H.; Lee, C.W.; Yang, K.; et al. 14-3-3 eta depletion sensitizes glioblastoma cells to irradiation due to enhanced mitotic cell death. Cancer Gene Ther. 2014, 21, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Lin, V.Y.; Ke, S.; Lin, G.E.; Lin, F.T.; Lin, W.C. 14-3-3tau promotes breast cancer invasion and metastasis by inhibiting RhoGDI alpha. Mol. Cell Biol. 2014, 34, 2635–2649. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.G.; Jeong, E.H.; Kim, S.Y.; Kim, H.R.; Kim, H.; Kim, C.H. Fhit, a tumor suppressor protein, induces autophagy via 14-3-3tau in non-small cell lung cancer cells. Oncotarget 2017, 8, 31923–31937. [Google Scholar] [PubMed]
- Yan, Y.; Xu, Y.; Gao, Y.Y.; Zong, Z.H.; Zhang, Q.; Li, C.; Wang, H.Q. Implication of 14-3-3epsilon and 14-3-3theta/tau in proteasome inhibition-induced apoptosis of glioma cells. Cancer Sci. 2013, 104, 55–61. [Google Scholar] [CrossRef]
- Obsilova, V.; Silhan, J.; Boura, E.; Teisinger, J.; Obsil, T. 14-3-3 proteins: A family of versatile molecular regulators. Physiol Res. 2008, 57, S11–S21. [Google Scholar]
- Bridges, D.; Moorhead, G.B. 14-3-3 proteins: A number of functions for a numbered protein. Sci. STKE 2005, 2005, re10. [Google Scholar] [CrossRef]
- Wilker, E.W.; Grant, R.A.; Artim, S.C.; Yaffe, M.B. A structural basis for 14-3-3sigma functional specificity. J. Biol. Chem. 2005, 280, 18891–18898. [Google Scholar] [CrossRef]
- Stevers, L.M.; Lam, C.V.; Leysen, S.F.; Meijer, F.A.; Scheppingen, D.S.; Vries, R.M.; Carlile, G.W.; Milroy, L.G.; Thomas, D.Y.; Brunsveld, L.; et al. Characterization and small-molecule stabilization of the multisite tandem binding between 14-3-3 and the R domain of CFTR. Proc. Natl. Acad. Sci. USA 2016, 113, 1152–1161. [Google Scholar] [CrossRef]
- Yang, X.; Lee, W.H.; Sobott, F.; Papagrigoriou, E.; Robinson, C.V.; Grossmann, J.G.; Sundstrom, M.; Doyle, D.A.; Elkins, J.M. Structural basis for protein-protein interactions in the 14-3-3 protein family. Proc. Natl. Acad. Sci. USA 2006, 103, 17237–17242. [Google Scholar] [CrossRef]
- Gardino, A.K.; Smerdon, S.J.; Yaffe, M.B. Structural determinants of 14-3-3 binding specificities and regulation of subcellular localization of 14-3-3-ligand complexes: A comparison of the X-ray crystal structures of all human 14-3-3 isoforms. Semin. Cancer Biol. 2006, 16, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Coblitz, B.; Wu, M.; Shikano, S.; Li, M. C-terminal binding: An expanded repertoire and function of 14-3-3 proteins. FEBS Lett. 2006, 580, 1531–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaffe, M.B.; Rittinger, K.; Volinia, S.; Caron, P.R.; Aitken, A.; Leffers, H.; Gamblin, S.J.; Smerdon, S.J.; Cantley, L.C. The structural basis for 14-3-3: phosphopeptide binding specificity. Cell 1997, 91, 961–971. [Google Scholar] [CrossRef]
- Alblova, M.; Smidova, A.; Docekal, V.; Vesely, J.; Herman, P.; Obsilova, V.; Obsil, T. Molecular basis of the 14-3-3 protein-dependent activation of yeast neutral trehalase Nth1. Proc. Natl. Acad. Sci. USA 2017, 114, E9811–E9820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganguly, S.; Weller, J.L.; Ho, A.; Chemineau, P.; Malpaux, B.; Klein, D.C. Melatonin synthesis: 14-3-3-dependent activation and inhibition of arylalkylamine N-acetyltransferase mediated by phosphoserine-205. Proc. Natl. Acad. Sci. USA 2005, 102, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Demmel, L.; Beck, M.; Klose, C.; Schlaitz, A.L.; Gloor, Y.; Hsu, P.P.; Havlis, J.; Shevchenko, A.; Krause, E.; Kalaidzidis, Y.; et al. Nucleocytoplasmic shuttling of the Golgi phosphatidylinositol 4-kinase Pik1 is regulated by 14-3-3 proteins and coordinates Golgi function with cell growth. Mol. Biol. Cell. 2008, 19, 1046–1061. [Google Scholar] [CrossRef]
- Yano, M.; Nakamuta, S.; Wu, X.; Okumura, Y.; Kido, H. A novel function of 14-3-3 protein: 14-3-3zeta is a heat-shock-related molecular chaperone that dissolves thermal-aggregated proteins. Mol. Biol. Cell. 2006, 17, 4769–4779. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.M.; Ecroyd, H.; Goodwin, K.L.; Dai, H.; Fu, H.; Woodcock, J.M.; Zhang, L.; Carver, J.A. NMR spectroscopy of 14-3-3zeta reveals a flexible C-terminal extension: Differentiation of the chaperone and phosphoserine-binding activities of 14-3-3zeta. Biochem. J. 2011, 437, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Sluchanko, N.N.; Roman, S.G.; Chebotareva, N.A.; Gusev, N.B. Chaperone-like activity of monomeric human 14-3-3zeta on different protein substrates. Arch. Biochem. Biophys. 2014, 549, 32–39. [Google Scholar] [CrossRef]
- Sluchanko, N.N.; Artemova, N.V.; Sudnitsyna, M.V.; Safenkova, I.V.; Antson, A.A.; Levitsky, D.I.; Gusev, N.B. Monomeric 14-3-3zeta has a chaperone-like activity and is stabilized by phosphorylated HspB6. Biochemistry 2012, 51, 6127–6138. [Google Scholar] [CrossRef]
- Lin, J.P.; Fan, Y.K.; Liu, H.M. The 14-3-3eta chaperone protein promotes antiviral innate immunity via facilitating MDA5 oligomerization and intracellular redistribution. PLoS Pathog. 2019, 15, e1007582. [Google Scholar] [CrossRef] [PubMed]
- Woodcock, J.M.; Goodwin, K.L.; Sandow, J.J.; Coolen, C.; Perugini, M.A.; Webb, A.I.; Pitson, S.M.; Lopez, A.F.; Carver, J.A. Role of salt bridges in the dimer interface of 14-3-3zeta in dimer dynamics, N-terminal alpha-helical order, and molecular chaperone activity. J. Biol. Chem. 2018, 293, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.L.; Yu, D. 14-3-3zeta as a prognostic marker and therapeutic target for cancer. Expert Opin. Ther. Targets 2010, 14, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.L.; Yao, J.; Yang, W.; Zhou, X.; Nguyen, N.T.; Lu, J.; Danes, C.G.; Guo, H.; Lan, K.H.; Ensor, J.; et al. 14-3-3zeta overexpression defines high risk for breast cancer recurrence and promotes cancer cell survival. Cancer Res. 2009, 69, 3425–3432. [Google Scholar] [CrossRef] [PubMed]
- Neal, C.L.; Xu, J.; Li, P.; Mori, S.; Yang, J.; Neal, N.N.; Zhou, X.; Wyszomierski, S.L.; Yu, D. Overexpression of 14-3-3ζ in cancer cells activates PI3K via binding the p85 regulatory subunit. Oncogene 2012, 31, 897–906. [Google Scholar] [CrossRef]
- Lee, J.J.; Lee, J.S.; Cui, M.N.; Yun, H.H.; Kim, H.Y.; Lee, S.H.; Lee, J.H. BIS targeting induces cellular senescence through the regulation of 14-3-3 zeta/STAT3/SKP2/p27 in glioblastoma cells. Cell Death Dis. 2014, 5, e1537. [Google Scholar] [CrossRef]
- Yu, J.; Chen, L.; Chen, Y.; Hasan, M.K.; Ghia, E.M.; Zhang, L.; Wu, R.; Rassenti, L.Z.; Widhopf, G.F.; Shen, Z.; et al. Wnt5a induces ROR1 to associate with 14-3-3ζ for enhanced chemotaxis and proliferation of chronic lymphocytic leukemia cells. Leukemia 2017, 31, 2608–2614. [Google Scholar] [CrossRef]
- Xu, J.; Acharya, S.; Sahin, O.; Zhang, Q.; Saito, Y.; Yao, J.; Wang, H.; Li, P.; Zhang, L.; Lowery, F.J.; et al. 14-3-3ζ turns TGF-β’s function from tumor suppressor to metastasis promoter in breast cancer by contextual changes of Smad partners from p53 to Gli2. Cancer Cell 2015, 27, 177–192. [Google Scholar] [CrossRef]
- Ling, C.; Su, V.M.; Zuo, D.; Muller, W.J. Loss of the 14-3-3σ tumor suppressor is a critical event in ErbB2-mediated tumor progression. Cancer Discov. 2012, 2, 68–81. [Google Scholar] [CrossRef]
- Umbricht, C.B.; Evron, E.; Gabrielson, E.; Ferguson, A.; Marks, J.; Sukumar, S. Hypermethylation of 14-3-3 sigma (stratifin) is an early event in breast cancer. Oncogene 2001, 20, 3348–3353. [Google Scholar] [CrossRef]
- Vercoutter-Edouart, A.S.; Lemoine, J.; Le, X.; Bourhis; Louis, H.; Boilly, B.; Nurcombe, V.; Révillion, F.; Peyrat, J.P.; Hondermarck, H. Proteomic analysis reveals that 14-3-3sigma is down-regulated in human breast cancer cells. Cancer Res. 2001, 61, 76–80. [Google Scholar] [PubMed]
- Radhakrishnan, V.M.; Jensen, T.J.; Cui, H.; Futscher, B.W.; Martinez, J.D. Hypomethylation of the 14-3-3σ promoter leads to increased expression in non-small cell lung cancer. Genes Chromosomes Cancer 2011, 50, 830–836. [Google Scholar] [CrossRef]
- Luo, J.; Feng, J.; Lu, J.; Wang, Y.; Tang, X.; Xie, F.; Li, W. Aberrant methylation profile of 14-3-3 sigma and its reduced transcription/expression levels in Chinese sporadic female breast carcinogenesis. Med. Oncol. 2010, 27, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Yi, B.; Tan, S.X.; Tang, C.E.; Huang, W.G.; Cheng, A.; Li, C.; Zhang, P.F.; Li, M.Y.; Li, J.L.; Yi, H.; et al. Inactivation of 14-3-3 sigma by promoter methylation correlates with metastasis in nasopharyngeal carcinoma. J. Cell. Biochem. 2009, 106, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Raungrut, P.; Petjaroen, P.; Geater, S.L.; Keeratichananont, W.; Phukaoloun, M.; Suwiwat, S.; Thongsuksai, P. Methylation of 14-3-3σ gene and prognostic significance of 14-3-3σ expression in non-small cell lung cancer. Oncology Lett. 2017, 14, 5257–5264. [Google Scholar] [CrossRef]
- Lodygin, D.; Hermeking, H. Epigenetic silencing of 14-3-3sigma in cancer. Semin. Cancer Biol. 2006, 16, 214–224. [Google Scholar] [CrossRef]
- Phan, L.; Chou, P.C.; Velazquez-Torres, G.; Samudio, I.; Parreno, K.; Huang, Y.; Tseng, C.; Vu, T.; Gully, C.; Su, C.H.; et al. The cell cycle regulator 14-3-3sigma opposes and reverses cancer metabolic reprogramming. Nat. Commun. 2015, 6, 7530. [Google Scholar] [CrossRef]
- Hong, H.Y.; Jeon, W.K.; Bae, E.J.; Kim, S.T.; Lee, H.J.; Kim, S.J.; Kim, B.C. 14-3-3 sigma and 14-3-3 zeta plays an opposite role in cell growth inhibition mediated by transforming growth factor-beta 1. Mol. Cells 2010, 29, 305–309. [Google Scholar] [CrossRef]
- Berg, D.; Holzmann, C.; Riess, O. 14-3-3 proteins in the nervous system. Nat. Rev. Neurosci. 2003, 4, 752–762. [Google Scholar] [CrossRef]
- Steinacker, P.; Aitken, A.; Otto, M. 14-3-3 proteins in neurodegeneration. Semin. Cell Dev. Biol. 2011, 22, 696–704. [Google Scholar] [CrossRef]
- Ge, W.W.; Volkening, K.; Leystra-Lantz, C.; Jaffe, H.; Strong, M.J. 14-3-3 protein binds to the low molecular weight neurofilament (NFL) mRNA 3’ UTR. Mol. Cell Neurosci. 2007, 34, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Cheah, P.S.; Ramshaw, H.S.; Thomas, P.Q.; Toyo-Oka, K.; Xu, X.; Martin, S.; Coyle, P.; Guthridge, M.A.; Stomski, F.; van den Buuse, M.; et al. Neurodevelopmental and neuropsychiatric behaviour defects arise from 14-3-3zeta deficiency. Mol. Psychiatry 2012, 17, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jaehne, E.J.; Greenberg, Z.; McCarthy, P.; Saleh, E.; Parish, C.L.; Camera, D.; Heng, J.; Haas, M.; Baune, B.T.; et al. 14-3-3zeta deficient mice in the BALB/c background display behavioural and anatomical defects associated with neurodevelopmental disorders. Sci. Rep. 2015, 5, 12434. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Hikita, T.; Taya, S.; Uraguchi-Asaki, J.; Toyo-oka, K.; Wynshaw-Boris, A.; Ujike, H.; Inada, T.; Takao, K.; Miyakawa, T.; et al. Identification of YWHAE, a gene encoding 14-3-3epsilon, as a possible susceptibility gene for schizophrenia. Hum. Mol. Genet. 2008, 17, 3212–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachi, T.; Cornell, B.; Toyo-Oka, K. Complete ablation of the 14-3-3epsilon protein results in multiple defects in neuropsychiatric behaviors. Behav. Brain Res. 2017, 319, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.E.; Cho, C.H.; Sim, K.M.; Kwon, O.; Hwang, E.M.; Kim, H.W.; Park, J.Y. 14-3-3gamma Haploinsufficient Mice Display Hyperactive and Stress-sensitive Behaviors. Exp. Neurobiol. 2019, 28, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Toyo-oka, K.; Wachi, T.; Hunt, R.F.; Baraban, S.C.; Taya, S.; Ramshaw, H.; Kaibuchi, K.; Schwarz, Q.P.; Lopez, A.F.; Wynshaw-Boris, A. 14-3-3epsilon and zeta regulate neurogenesis and differentiation of neuronal progenitor cells in the developing brain. J. Neurosci. 2014, 34, 12168–12181. [Google Scholar] [CrossRef] [PubMed]
- Cornell, B.; Toyo-oka, K. Deficiency of 14-3-3epsilon and 14-3-3zeta by the Wnt1 promoter-driven Cre recombinase results in pigmentation defects. BMC Res. Notes. 2016, 9, 180. [Google Scholar] [CrossRef]
- Foote, M.; Qiao, H.; Graham, K.; Wu, Y.; Zhou, Y. Inhibition of 14-3-3 Proteins Leads to Schizophrenia-Related Behavioral Phenotypes and Synaptic Defects in Mice. Biol. Psychiatry 2015, 78, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Graham, K.; Zhang, J.; Qiao, H.; Wu, Y.; Zhou, Y. Region-specific inhibition of 14-3-3 proteins induces psychomotor behaviors in mice. NPJ Schizophr. 2019, 5, 1. [Google Scholar] [CrossRef]
- Perez, R.G.; Waymire, J.C.; Lin, E.; Liu, J.J.; Guo, F.; Zigmond, M.J. A role for alpha-synuclein in the regulation of dopamine biosynthesis. J. Neurosci. 2002, 22, 3090–3099. [Google Scholar] [CrossRef] [PubMed]
- Ostrerova, N.; Petrucelli, L.; Farrer, M.; Mehta, N.; Choi, P.; Hardy, J.; Wolozin, B. alpha-Synuclein shares physical and functional homology with 14-3-3 proteins. J. Neurosci. 1999, 19, 5782–5791. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Kao, S.Y.; Lee, F.J.; Song, W.; Jin, L.W.; Yankner, B.A. Dopamine-dependent neurotoxicity of alpha-synuclein: A mechanism for selective neurodegeneration in Parkinson disease. Nat. Med. 2002, 8, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Plotegher, N.; Kumar, D.; Tessari, I.; Brucale, M.; Munari, F.; Tosatto, L.; Belluzzi, E.; Greggio, E.; Bisaglia, M.; Capaldi, S.; et al. The chaperone-like protein 14-3-3eta interacts with human alpha-synuclein aggregation intermediates rerouting the amyloidogenic pathway and reducing alpha-synuclein cellular toxicity. Hum. Mol. Genet. 2014, 23, 5615–5629. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lou, H.; Pedersen, C.J.; Smith, A.D.; Perez, R.G. 14-3-3zeta contributes to tyrosine hydroxylase activity in MN9D cells: Localization of dopamine regulatory proteins to mitochondria. J. Biol. Chem. 2009, 284, 14011–14019. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Chiba, T.; Sakata, E.; Kato, K.; Mizuno, Y.; Hattori, N.; Tanaka, K. 14-3-3eta is a novel regulator of parkin ubiquitin ligase. EMBO J. 2006, 25, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Paisan-Ruiz, C.; Jain, S.; Evans, E.W.; Gilks, W.P.; Simon, J.; van der Brug, M.; Lopez de Munain, A.; Aparicio, S.; Gil, A.M.; Khan, N.; et al. Cloning of the gene containing mutations that cause PARK8-linked Parkinson’s disease. Neuron 2004, 44, 595–600. [Google Scholar] [CrossRef]
- Su, B.; Liu, H.; Wang, X.; Chen, S.G.; Siedlak, S.L.; Kondo, E.; Choi, R.; Takeda, A.; Castellani, R.J.; Perry, G.; et al. Ectopic localization of FOXO3a protein in Lewy bodies in Lewy body dementia and Parkinson’s disease. Mol. Neurodegener 2009, 4, 32. [Google Scholar] [CrossRef]
- Ehrenberg, A.J.; Nguy, A.K.; Theofilas, P.; Dunlop, S.; Suemoto, C.K.; Di Lorenzo Alho, A.T.; Leite, R.P.; Diehl Rodriguez, R.; Mejia, M.B.; Rub, U.; et al. Quantifying the accretion of hyperphosphorylated tau in the locus coeruleus and dorsal raphe nucleus: The pathological building blocks of early Alzheimer’s disease. Neuropathol. Appl. Neurobiol. 2017, 43, 393–408. [Google Scholar] [CrossRef]
- Umahara, T.; Uchihara, T.; Tsuchiya, K.; Nakamura, A.; Iwamoto, T.; Ikeda, K.; Takasaki, M. 14-3-3 proteins and zeta isoform containing neurofibrillary tangles in patients with Alzheimer’s disease. Acta Neuropathol. 2004, 108, 279–286. [Google Scholar] [CrossRef]
- McFerrin, M.B.; Chi, X.; Cutter, G.; Yacoubian, T.A. Dysregulation of 14-3-3 proteins in neurodegenerative diseases with Lewy body or Alzheimer pathology. Ann. Clin. Transl. Neurol. 2017, 4, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Agarwal-Mawal, A.; Paudel, H.K. 14-3-3 binds to and mediates phosphorylation of microtubule-associated tau protein by Ser9-phosphorylated glycogen synthase kinase 3beta in the brain. J. Biol. Chem. 2004, 279, 26105–26114. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Han, J.R.; Chang, O.; Oh, M.; James, S.E.; Lu, Q.; Seo, Y.W.; Kim, H.; Kim, K. 14-3-3varepsilon/zeta Affects the stability of delta-catenin and regulates delta-catenin-induced dendrogenesis. FEBS Open Bio. 2013, 3, 16–21. [Google Scholar] [CrossRef]
- Zhou, J.; Liyanage, U.; Medina, M.; Ho, C.; Simmons, A.D.; Lovett, M.; Kosik, K.S. Presenilin 1 interaction in the brain with a novel member of the Armadillo family. Neuroreport 1997, 8, 2085–2090. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Underwood, R.; Lavalley, N.; Yacoubian, T.A. 14-3-3 inhibition promotes dopaminergic neuron loss and 14-3-3θ overexpression promotes recovery in the MPTP mouse model of Parkinson’s disease. Neuroscience 2015, 307, 73–82. [Google Scholar] [CrossRef]
- Betarbet, R.; Sherer, T.B.; MacKenzie, G.; Garcia-Osuna, M.; Panov, A.V.; Greenamyre, J.T. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 2000, 3, 1301–1306. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Wu, Y.; Zhou, Y. 14-3-3 and aggresome formation: Implications in neurodegenerative diseases. Prion 2014, 8, 173–177. [Google Scholar] [CrossRef]
- Xu, Y.; Ren, J.; He, X.; Chen, H.; Wei, T.; Feng, W. YWHA/14-3-3 proteins recognize phosphorylated TFEB by a noncanonical mode for controlling TFEB cytoplasmic localization. Autophagy 2019, 15, 1017–1030. [Google Scholar] [CrossRef]
- Wang, Y.; Oh, S.W.; Deplancke, B.; Luo, J.; Walhout, A.J.; Tissenbaum, H.A. C. elegans 14-3-3 proteins regulate life span and interact with SIR-2.1 and DAF-16/FOXO. Mech. Ageing Dev. 2006, 127, 741–747. [Google Scholar] [CrossRef]
- Berdichevsky, A.; Viswanathan, M.; Horvitz, H.R.; Guarente, L.C. elegans SIR-2.1 interacts with 14-3-3 proteins to activate DAF-16 and extend life span. Cell 2006, 125, 1165–1177. [Google Scholar] [CrossRef]
- Araiz, C.; Chateau, M.T.; Galas, S. 14-3-3 regulates life span by both DAF-16-dependent and -independent mechanisms in Caenorhabditis elegans. Exp. Gerontol. 2008, 43, 505–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.D.; Luo, X.; Biteau, B.; Syverson, K.; Jasper, H. 14-3-3 Epsilon antagonizes FoxO to control growth, apoptosis and longevity in Drosophila. Aging Cell. 2008, 7, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Le, T.P.; Vuong, L.T.; Kim, A.R.; Hsu, Y.C.; Choi, K.W. 14-3-3 proteins regulate Tctp-Rheb interaction for organ growth in Drosophila. Nat. Commun. 2016, 7, 11501. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.; Lithgow, G.J.; Link, W. Long live FOXO: Unraveling the role of FOXO proteins in aging and longevity. Aging cell. 2016, 15, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.S.; Kapahi, P.; Hsueh, W.C.; Kockel, L. TOR signaling never gets old: Aging, longevity and TORC1 activity. Ageing Res. Rev. 2011, 10, 225–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleppe, R.; Martinez, A.; Doskeland, S.O.; Haavik, J. The 14-3-3 proteins in regulation of cellular metabolism. Semin. Cell Dev. Biol. 2011, 22, 713–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozuelo, M.; Peggie, M.; Wong, B.H.; Morrice, N.; MacKintosh, C. 14-3-3s regulate fructose-2, 6-bisphosphate levels by binding to PKB-phosphorylated cardiac fructose-2,6-bisphosphate kinase/phosphatase. EMBO J. 2003, 22, 3514–3523. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yoo, S.; Kim, J.; An, H.T.; Kang, M.; Ko, J. 14-3-3beta and gamma differentially regulate peroxisome proliferator activated receptor gamma2 transactivation and hepatic lipid metabolism. Biochim. Biophys. Acta 2015, 1849, 1237–1247. [Google Scholar] [CrossRef]
- Insenser, M.; Montes-Nieto, R.; Vilarrasa, N.; Lecube, A.; Simo, R.; Vendrell, J.; Escobar-Morreale, H.F. A nontargeted proteomic approach to the study of visceral and subcutaneous adipose tissue in human obesity. Mol. Cell Endocrinol. 2012, 363, 10–19. [Google Scholar] [CrossRef]
- Lim, G.E.; Albrecht, T.; Piske, M.; Sarai, K.; Lee, T.C.; Ramshaw, H.S.; Sinha, S.; Guthridge, M.A.; Acker-Palmer, A.; Lopez, A.F.; et al. 14-3-3zeta coordinates adipogenesis of visceral fat. Nat. Commun. 2015, 6, 7671. [Google Scholar] [CrossRef]
- Taurino, F.; Stanca, E.; Vonghia, L.; Siculella, L.; Sardanelli, A.M.; Papa, S.; Zanotti, F.; Gnoni, A. Short-term type-1 diabetes differentially modulates 14-3-3 proteins in rat brain and liver. Eur. J. Clin. Invest. 2014, 44, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Skinner, C.; Easlon, E.; Lin, S.J. Deleting the 14-3-3 protein Bmh1 extends life span in Saccharomyces cerevisiae by increasing stress response. Genetics 2009, 183, 1373–1384. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.B.; Lee, J.J.; Yun, H.H.; Im, C.N.; Kim, Y.S.; Ko, J.H.; Lee, J.H. 14-3-3beta Depletion Drives a Senescence Program in Glioblastoma Cells Through the ERK/SKP2/p27 Pathway. Mol. Neurobiol. 2018, 55, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Sreedhar, R.; Giridharan, V.V.; Arumugam, S.; Karuppagounder, V.; Palaniyandi, S.S.; Krishnamurthy, P.; Quevedo, J.; Watanabe, K.; Konishi, T.; Thandavarayan, R.A. Role of MAPK-mediated endoplasmic reticulum stress signaling in the heart during aging in senescence-accelerated prone mice. BioFactors 2016, 42, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.Y.; Wang, P.W.; Huang, C.H.; Chen, M.H.; Wu, Y.R.; Pan, T.L. Skin aging caused by intrinsic or extrinsic processes characterized with functional proteomics. Proteomics 2016, 16, 2718–2731. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 14-3-3 Isoform | Cancer Type | Expression | Reference |
|---|---|---|---|
| 14-3-3ζ | Breast, lung, pancreas, esophageal, head and neck, oral, colon, chronic myeloid leukemia, ovarian | ↑ | [26,27,28,29,30,31] |
| 14-3-3σ | Lung, breast, esophageal, chronic myeloid leukemia, uterine, ovarian, skin | ↓ | [32,33,34,35,36] |
| Liver, pancreatic ductal | ↑ | [37,38] | |
| 14-3-3β | Lung, astrocytoma, glioma, colorectal, gastric squamous, liver | ↑ | [39,40,41,42] |
| 14-3-3ε | Renal, liver, squamous, breast, gastric | ↑ | [43,44,45,46] |
| 14-3-3γ | Liver, breast, lung | ↑ | [47,48,49] |
| 14-3-3η | Liver, prostate, squamous, glioma | ↑ | [49,50,51,52] |
| 14-3-3τ/θ | Breast, lung, glioma, prostate | ↑ | [53,54,55] |
| K/O Isoform | Phenotypes | References |
|---|---|---|
| 14-3-3ζ | Schizophrenia, autism spectrum disorder, and bipolar disorder; reduced learning, memory, and prepulse inhibition and locomotor hyperactivity | [92,93] |
| 14-3-3ε | Schizophrenic behavior; increased locomotor activity and sociability and decreased working memory | [94,95] |
| 14-3-3γ | Hyperactive and depressive-like behavior; sensitive responses to acute stress | [96] |
| 14-3-3ζ and 14-3-3ε | Neuronal migration and pigmentation defects and neural progenitor cells | [14,97,98] |
| 14-3-3 functional knock-out | Schizophrenic behavior; synaptic alterations | [99,100] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Cui, L.; Zeng, Y.; Song, W.; Gaur, U.; Yang, M. 14-3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease. Int. J. Mol. Sci. 2019, 20, 3518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143518
Fan X, Cui L, Zeng Y, Song W, Gaur U, Yang M. 14-3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease. International Journal of Molecular Sciences. 2019; 20(14):3518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143518
Chicago/Turabian StyleFan, Xiaolan, Lang Cui, Yao Zeng, Wenhao Song, Uma Gaur, and Mingyao Yang. 2019. "14-3-3 Proteins Are on the Crossroads of Cancer, Aging, and Age-Related Neurodegenerative Disease" International Journal of Molecular Sciences 20, no. 14: 3518. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143518