Drought Resistance in Rice from Conventional to Molecular Breeding: A Review

 , , , ,
, , , ,
Abstract
:1. Introduction
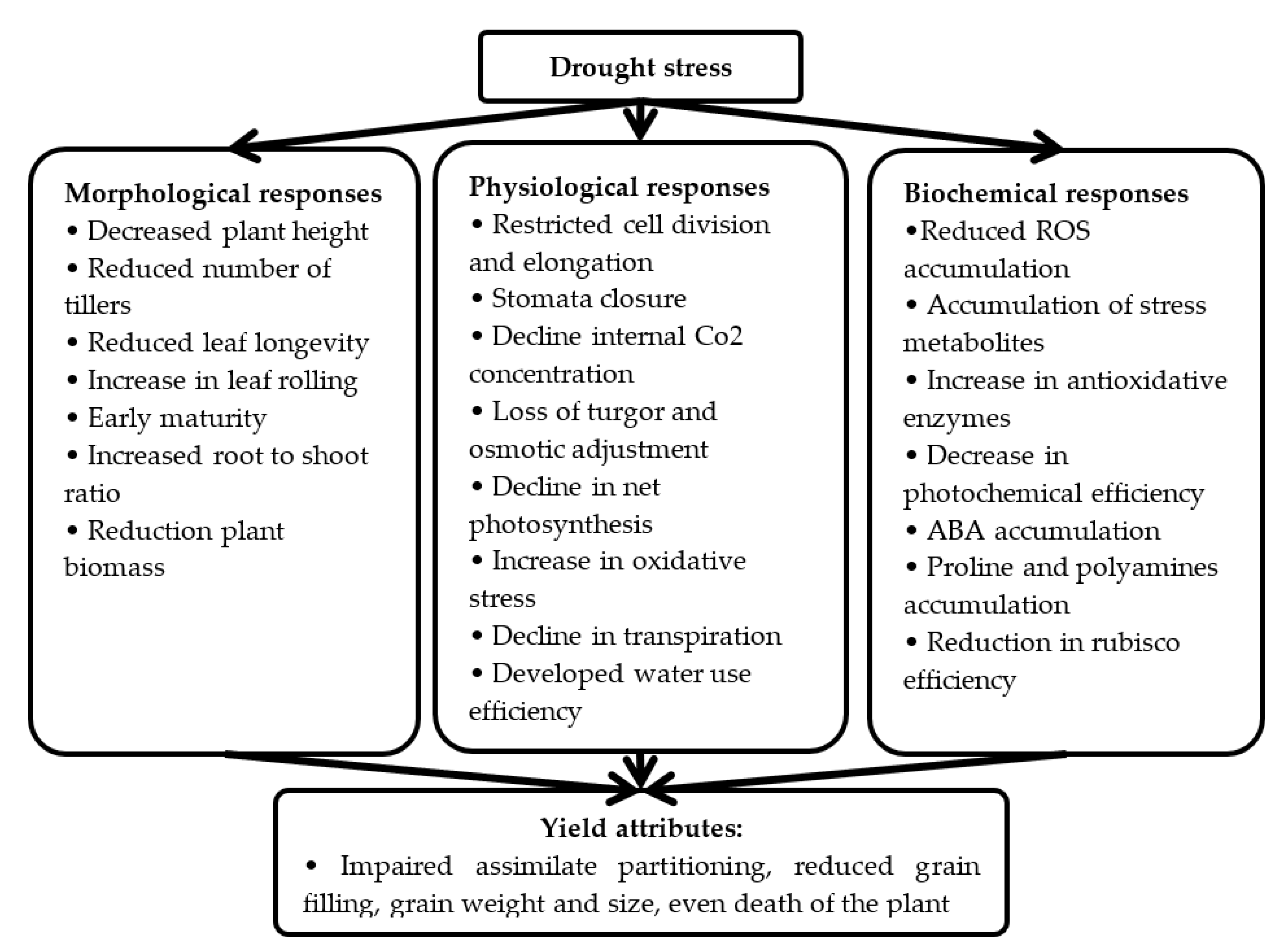
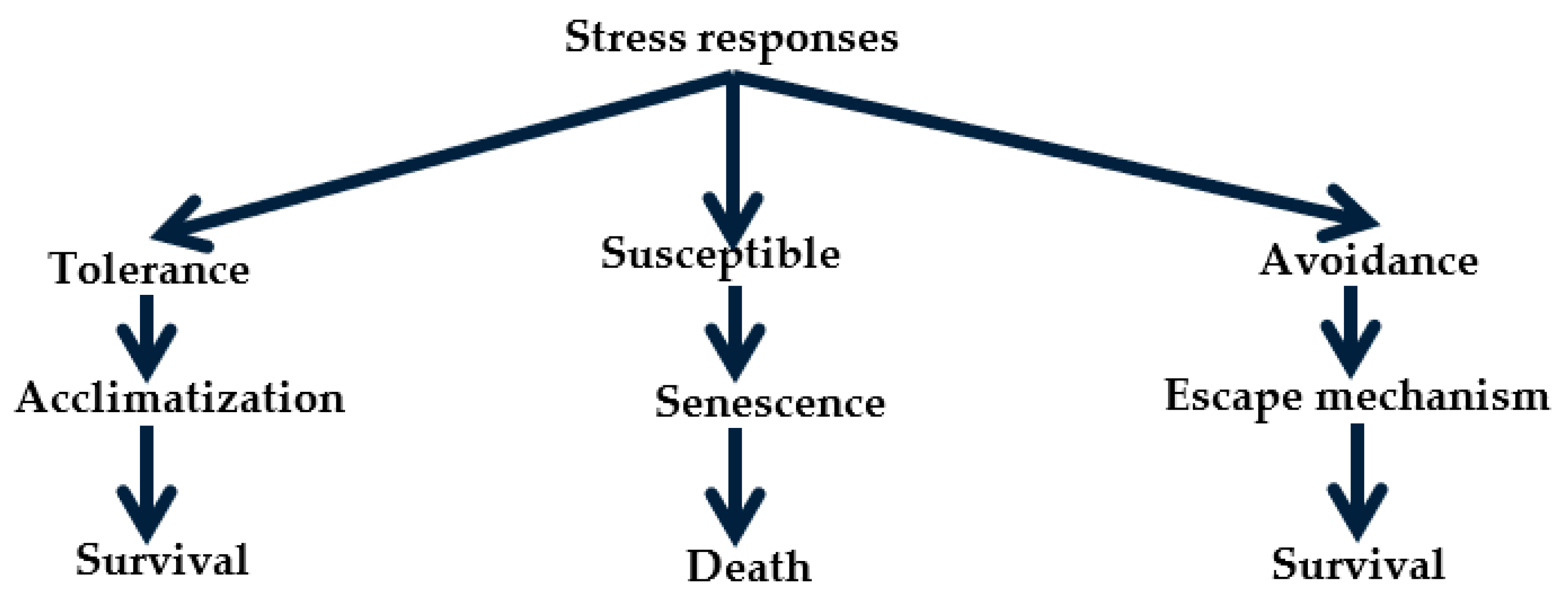
2. Drought Stress: Perception, Biochemical Responses and Mechanism
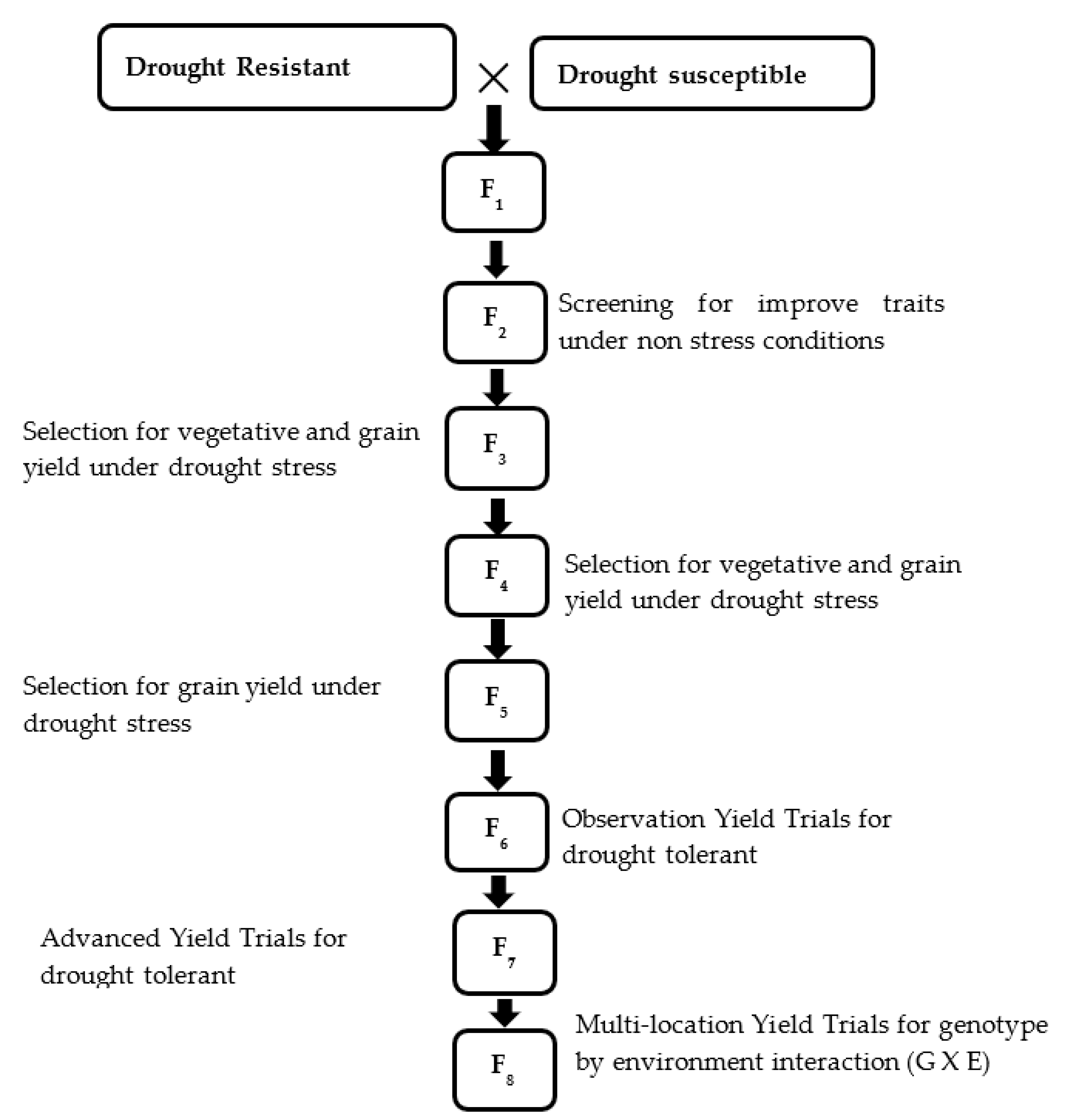
3. Conventional Breeding
4. Biotechnological and Molecular Approaches for Drought Tolerance
4.1. QTL for Drought (List of QTL Genes for Drought)
4.2. Genetic Engineering for Drought
4.3. Marker-Assisted Selection (MAS) for Drought Tolerance
5. Limitations and Future Prospects
- It is highly questionable that only one experiment in the laboratory determines the ability of a certain gene to confer drought resistance that is beneficial in breeding.
- Traits of interest should consist of aerial and root organs, or phenology, particularly those influencing plant–water relations. This is expected to help in the setup of succeeding stress trials and drawing a conclusion on the transgene.
- Multiple stress trials should be conducted to determine early whether transgenic plants have the ability to avoid and/or tolerate dehydration.
- In general, the relative water content is preferred to leaf water potential as a measure of leaf water status since the relative water content also signifies the capacity for osmotic adjustment. For the same leaf water potential, a dehydration-avoidant genotype will express higher relative water content.
- In the case where dehydration avoidance is detected in the transgenic plant, the trait should be evaluated to confirm whether it results from sensitive stomatal closure, turgor preservation by osmotic adjustment or preservation of turgor due to enhanced ability of the root to take up soil moisture.
- Measurement of physiological functions should be associated with growth and photosynthesis. Antioxidant accumulation and function are also common measurements despite their relationship with whole-plant growth are not as well established, unlike photosynthesis.
- Due to the effect of dehydration avoidance or tolerance of meristems on plant survival, survival or recovery from stress should, among others, be determined by relative water content at peak stress prior recovery.
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sahebi, M.; Hanafi, M.M.; Rafii, M.Y.; Mahmud, T.M.M.; Azizi, P.; Osman, M.; Miah, G. Improvement of drought tolerance in rice (Oryza sativa L.): Genetics, genomic tools, and the WRKY gene family. BioMed Res. Int. 2018, 2018, 3158474. [Google Scholar] [CrossRef] [PubMed]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Hasan, M.M.; Oladosu, Y.A.; Olalekan, K.K. Bacterial leaf blight resistance in rice: A review of conventional breeding to molecular approach. Mol. Biol. Rep. 2019, 46, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Puteh, A.B.; Rahim, H.A.; Asfaliza, R.; Latif, M.A. Blast resistance in rice: A review of conventional breeding to molecular approaches. Mol. Biol. Rep. 2013, 40, 2369–2388. [Google Scholar] [CrossRef] [PubMed]
- Singhal, P.; Jan, A.T.; Azam, M.; Haq, Q.M.R. Plant abiotic stress: A prospective strategy of exploiting promoters as alternative to overcome the escalating burden. Front. Life Sci. 2016, 9, 52–63. [Google Scholar] [CrossRef]
- Gosal, S.S.; Wani, S.H.; Kang, M.S. Biotechnology and drought tolerance. J. Crop Improv. 2009, 23, 19–54. [Google Scholar] [CrossRef]
- Myers, S.S.; Smith, M.R.; Sarah, G.; Golden, C.D.; Bapu, V.; Mueller, N.D.; Dangour, A.D.; Peter, H. Climate change and global food systems: Potential impacts on food security and undernutrition. Annu. Rev. Publ. Health 2017, 38, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Ozga, J.A.; Kaur, H.; Savada, R.P.; Reinecke, D.M. Hormonal regulation of reproductive growth under normal and heat-stress conditions in legume and other model crop species. J. Exp. Bot. 2016, 68, 1885–1894. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Magaji, U.; Miah, G.; Hussin, G.; Ramli, A. Genotype× Environment interaction and stability analyses of yield and yield components of established and mutant rice genotypes tested in multiple locations in Malaysia. Acta. Agr. Scand. 2017, 67, 590–606. [Google Scholar] [CrossRef]
- Polania, J.; Rao, I.M.; Cajiao, C.; Grajales, M.; Rivera, M.; Velasquez, F.; Beebe, S.E. Shoot and root traits contribute to drought resistance in recombinant inbred lines of MD 23–24× SEA 5 of common bean. Front. Plant Sci. 2017, 8, 296. [Google Scholar] [CrossRef]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Zohaib, A.; Tanveer, M.; Naeem, M.; Ali, I.; Nazir, U. Growth and development responses of crop plants under drought stress: A review. Zemdirbyste 2017, 104, 267–276. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Ihsan, M.Z. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Fahramand, M.; Mahmoody, M.; Keykha, A.; Noori, M.; Rigi, K. Influence of abiotic stress on proline, photosynthetic enzymes and growth. Int. Res. J. Appl. Basic Sci. 2014, 8, 257–265. [Google Scholar]
- Lum, M.S.; Hanafi, M.M.; Rafii, Y.M.; Akmar, A.S.N. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Capell, T.; Bassie, L.; Christou, P. Modulation of the polyamine biosynthetic pathway in transgenic rice confers tolerance to drought stress. Proc. Natl. Acad. Sci. USA 2004, 101, 9909–9914. [Google Scholar] [CrossRef] [Green Version]
- Raghavendra, A.S.; Gonugunta, V.K.; Christmann, A.; Grill, E. ABA perception and signalling. Trends Plant Sci. 2010, 15, 395–401. [Google Scholar] [CrossRef]
- Li, J.; Wu, Y.; Xie, Q.; Gong, Z. Hormone Metabolism and Signaling in Plants; Academic Press: Cambridge, MA, USA, 2017; pp. 161–202. [Google Scholar]
- Nezhadahmadi, A.; Prodhan, Z.H.; Faruq, G. Drought tolerance in wheat. Sci. World J. 2013. [Google Scholar] [CrossRef]
- Chourasia, K.N. Resistance/Tolerance mechanism under water deficit (Drought) condition in plants. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 66–78. [Google Scholar]
- Luo, L.J. Breeding for water-saving and drought-resistance rice (WDR) in China. J. Exp. Bot. 2010, 61, 3509–3517. [Google Scholar] [CrossRef] [Green Version]
- Bolaños, J.; Edmeades, G.O. Eight cycles of selection for drought tolerance in lowland tropical maize. II. Responses in reproductive behaviour. Field Crops Res. 1993, 31, 253–268. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef] [PubMed]
- Dixit, S.; Singh, A.; Kumar, A. Rice breeding for high grain yield under drought: A strategic solution to a complex problem. Int. J. Agron. 2014. [Google Scholar] [CrossRef]
- Khush, G.S. IRRI breeding program and its worldwide impact on increasing rice production. In Gene manipulation in plant improvement; Springer: Boston, MA, USA, 1984; pp. 61–94. [Google Scholar]
- Pang, Y.; Chen, K.; Wang, X.; Xu, J.; Ali, J.; Li, Z. Recurrent selection breeding by dominant male sterility for multiple abiotic stresses tolerant rice cultivars. Euphytica 2017, 213, 268. [Google Scholar] [CrossRef]
- Posadas, L.G.; Eskridge, K.M.; Specht, J.E.; Graef, G.L. Elite performance for grain yield from unadapted exotic soybean germplasm in three cycles of a recurrent selection experiment. Crop Sci. 2014, 54, 2536–2546. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Condon, A.G.; Richards, R.A.; Farquhar, G.D. Selection for reduced carbon isotope discrimination increases aerial biomass and grain yield of rainfed bread wheat. Crop Sci. 2002, 42, 739–745. [Google Scholar] [CrossRef]
- Wiersma, J.J.; Busch, R.H.; Fulcher, G.G.; Hareland, G.A. Recurrent selection for kernel weight in spring wheat. Crop Sci. 2001, 41, 999–1005. [Google Scholar] [CrossRef]
- Delzer, B.W.; Hareland, G.A. Recurrent selection for grain protein in hard red spring wheat. Crop Sci. 1995, 35, 730–735. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Li, Z.K.; Vijayakumar, C.H.M.; Gao, Y.M.; Shi, Y.; Xu, J.L.; Maghirang, R. Improvement of rice drought tolerance through backcross breeding: Evaluation of donors and selection in drought nurseries. Field Crop Res. 2006, 97, 77–86. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; AbdulMalek, M.; Rahim, H.A.; Hussin, G.; Kareem, I. Genetic variability and selection criteria in rice mutant lines as revealed by quantitative traits. Sci. World J. 2014. [Google Scholar] [CrossRef]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; AbdulMalek, M.; Rahim, H.A.; Hussin, G.; Kareem, I. Genetic variability and diversity of mutant rice revealed by quantitative traits and molecular markers. Agrociencia 2015, 49, 249–266. [Google Scholar]
- Oladosu, Y.; Rafii, M.Y.; Abdullah, N.; Hussin, G.; Ramli, A.; Rahim, H.A.; Usman, M. Principle and application of plant mutagenesis in crop improvement: A review. Biotechnol. Biotechnol. Equip. 2016, 30, 1–16. [Google Scholar] [CrossRef]
- Soe, H.M.; Myat, M.; Khaing, Z.L.; Nyo, N.M.; Phyu, P.T. Development of Drought Tolerant Mutant from Rice var. Manawthukha through Mutation Breeding Technique using 60Co Gamma Source. Int. J. Innov. Res. Sci. Eng. Technol. 2016, 4, 11205–11212. [Google Scholar]
- Hallajian, M.T.; Ebadi, A.A.; Mohammadi, M.; Muminjanov, H.; Jamali, S.S.; Aghamirzaei, M. Integration of Mutation and Conventional Breeding Approaches to Develop New Superior Drought-tolerant Plants in Rice (Oryza sativa). Annu. Res. Rev. Biol. 2014, 4, 1173–1186. [Google Scholar] [CrossRef]
- Efendi, B.; Sabaruddin, Z.; Lukman, H. Mutation with gamma raysirradiation to assemble green super rice tolerant to drought stress and high yield rice (oryza sativa l.). Int. J. Adv. Sci. Eng. Tech. 2017, 5, 1–5. [Google Scholar]
- Rahim, H.A.; Zarifth, S.K.; Bhuiyan, M.A.R.; Narimah, M.K.; Wickneswari, R.; Abdullah, M.Z. Evaluation and characterization of advanced rice mutant line of rice (Oryza sativa), MR219-4 and MR219-9 under drought condition. Res. Dev. Semin. 2012, 44, 26–28. [Google Scholar]
- Fleury, D.; Jefferies, S.; Kuchel, H.; Langridge, P. Genetic and genomic tools to improve drought tolerance in wheat. J. Exp. Bot. 2010, 61, 3211–3222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinclair, T.R. Challenges in breeding for yield increase for drought. Trends Plant Sci. 2011, 16, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Reif, J.C.; Ranc, N.; Della, P.G.; Würschum, T. Comparison of biometrical approaches for QTL detection in multiple segregating families. Theor. Appl. Genet. 2012, 125, 987–998. [Google Scholar] [CrossRef]
- Rafalski, J.A. Association genetics in crop improvement. Curr. Opin. Plant Biol. 2010, 13, 174–180. [Google Scholar] [CrossRef]
- Zheng, B.S.; Yang, L.; Mao, C.Z.; Huang, Y.J.; Wu, P. Mapping QTLs for morphological traits under two water supply conditions at the young seedling stage in rice. Plant Sci. 2008, 175, 767–776. [Google Scholar] [CrossRef]
- Tripathy, J.N.; Zhang, J.; Robin, S.; Nguyen, T.T.; Nguyen, H.T. QTLs for cell-membrane stability mapped in rice (Oryza sativa L.) under drought stress. Theor. Appl. Genet. 2000, 100, 1197–1202. [Google Scholar] [CrossRef]
- Khowaja, F.S.; Price, A.H. QTL mapping rolling, stomata conductance and dimension traits of excised leaves in the Bala×Azucena recombinant inbred population of rice. Field crop res. 2008, 106, 248–257. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Q.; Zheng, T.; Cui, Y.; Zhang, W.; Xu, J.; Li, Z. Drought-tolerance QTLs commonly detected in two sets of reciprocal introgression lines in rice. Crop Pasture Sci. 2014, 65, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.L.; Pathan, M.S.; Zhang, J.; Bai, G.; Sarkarung, S.; Nguyen, H.T. Mapping QTLs for root traits in a recombinant inbred population from two indica ecotypes in rice. Theor. Appl. Genet. 2000, 101, 756–766. [Google Scholar] [CrossRef]
- Yadav, R.; Courtois, B.; Huang, N.; McLaren, G. Mapping genes controlling root morphology and root distribution in a doubled-haploid population of rice. Theor. Appl. Genet. 1997, 94, 619–632. [Google Scholar] [CrossRef]
- Zheng, B.S.; Yang, L.; Zhang, W.P.; Mao, C.Z.; Wu, Y.R.; Yi, K.K.; Wu, P. Mapping QTLs and candidate genes for rice root traits under different water-supply conditions and comparative analysis across three populations. Theor. Appl. Genet. 2003, 107, 1505–1515. [Google Scholar] [CrossRef]
- Lou, Q.; Chen, L.; Mei, H.; Wei, H.; Feng, F.; Wang, P.; Luo, L. Quantitative trait locus mapping of deep rooting by linkage and association analysis in rice. J. Exp. Bot. 2015, 66, 4749–4757. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.D.; Yu, L.; McCouch, S.R.; Champoux, M.C.; Wang, G.; Nguyen, H.T. Mapping quantitative trait loci associated with root penetration ability in rice (Oryza sativa L.). Theor. Appl. Genet. 1996, 92, 627–636. [Google Scholar] [CrossRef]
- Price, A.H.; Steele, K.A.; Moore, B.J.; Barraclough, P.P.; Clark, L.J. A combined RFLP and AFLP linkage map of upland rice (Oryza sativa L.) used to identify QTLs for root-penetration ability. Theor. Appl. Genet. 2000, 100, 49–56. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.M.; Dixit, S.; Ahmed, H.; Cruz, M.S.; Singh, A.K.; Kumar, A. Bulk segregant analysis:“An effective approach for mapping consistent-effect drought grain yield QTLs in rice”. Field Crop Res. 2012, 134, 185–192. [Google Scholar] [CrossRef]
- Venuprasad, R.; Bool, M.E.; Quiatchon, L.; Atlin, G.N. A QTL for rice grain yield in aerobic environments with large effects in three genetic backgrounds. Theor. Appl. Genet. 2012, 124, 323–332. [Google Scholar] [CrossRef]
- Trijatmiko, K.R.; Prasetiyono, J.; Thomson, M.J.; Cruz, C.M.V.; Moeljopawiro, S.; Pereira, A. Meta-analysis of quantitative trait loci for grain yield and component traits under reproductive-stage drought stress in an upland rice population. Mol. Breed. 2014, 34, 283–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikumar, S.; Gouda, P.K.; Saiharini, A.; Varma, C.M.K.; Vineesha, O.; Padmavathi, G.; Shenoy, V.V. Major QTL for enhancing rice grain yield under lowland reproductive drought stress identified using an O. sativa/O. glaberrima introgression line. Field Crop Res. 2014, 163, 119–131. [Google Scholar] [CrossRef]
- Kumar, R.; Venuprasad, R.; Atlin, G.N. Genetic analysis of rainfed lowland rice drought tolerance under naturally-occurring stress in eastern India: Heritability and QTL effects. Field Crop Res. 2007, 103, 42–52. [Google Scholar] [CrossRef]
- Yadav, R.B.; Dixit, S.; Raman, A.; Mishra, K.K.; Vikram, P.; Swamy, B.M.; Kumar, A. A QTL for high grain yield under lowland drought in the background of popular rice variety Sabitri from Nepal. Field Crop Res. 2013, 144, 281–287. [Google Scholar]
- Venuprasad, R.; Dalid, C.O.; Del Valle, M.; Zhao, D.; Espiritu, M.; Cruz, M.S.; Atlin, G.N. Identification and characterization of large-effect quantitative trait loci for grain yield under lowland drought stress in rice using bulk-segregant analysis. Theor. Appl. Genet. 2009, 120, 177–190. [Google Scholar] [CrossRef]
- Palanog, A.D.; Swamy, B.M.; Shamsudin, N.A.A.; Dixit, S.; Hernandez, J.E.; Boromeo, T.H.; Kumar, A. Grain yield QTLs with consistent-effect under reproductive-stage drought stress in rice. Field Crop Res. 2014, 161, 46–54. [Google Scholar] [CrossRef]
- Hemamalini, G.S.; Shashidhar, H.E.; Hittalmani, S. Molecular marker assisted tagging of morphological and physiological traits under two contrasting moisture regimes at peak vegetative stage in rice (Oryza sativa L.). Euphytica 2000, 112, 69–78. [Google Scholar] [CrossRef]
- Price, A.H.; Townend, J.; Jones, M.P.; Audebert, A.; Courtois, B. Mapping QTLs associated with drought avoidance in upland rice grown in the Philippines and West Africa. Plant Mol. Biol. 2002, 48, 683–695. [Google Scholar] [CrossRef]
- Lilley, J.M.; Ludlow, M.M.; McCouch, S.R.; O’toole, J.C. Locating QTL for osmotic adjustment and dehydration tolerance in rice. J. Exp. Bot. 1996, 47, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Zheng, H.G.; Aarti, A.; Pantuwan, G.; Nguyen, T.T.; Tripathy, J.N.; Sarkarung, S. Locating genomic regions associated with components of drought resistance in rice: Comparative mapping within and across species. Theor. Appl. Genet. 2001, 103, 19–29. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Molecular analysis of Hsp70 mechanisms in plants and their function in response to stress. Biotechnol. Genet. Eng. Rev. 2017, 33, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Chao, D.Y.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomata aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.Q.; Huang, J.; Guo, S.Q.; Yang, X.; Bao, Y.M.; Tang, H.J.; Zhang, H.S. Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice (Oryza sativa L.). FEBS Lett. 2008, 582, 1037–1043. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.; Baek, W.; Jung, J.; Kim, J.H.; Lee, S. Function of ABA in stomatal defense against biotic and drought stresses. Int. J. Mol. Sci. 2015, 16, 15251–15270. [Google Scholar] [CrossRef]
- Yadira, O.-C.; Reyes, J.L.; Alejandra, A.C. Late embryogenesis abundant proteins. Plant Signal Behav. 2011, 6, 586–589. [Google Scholar] [Green Version]
- Duan, J.; Cai, W. OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance. PLoS ONE 2012, 7, e45117. [Google Scholar] [CrossRef]
- Xiao, B.; Huang, Y.; Tang, N.; Xiong, L. Over-expression of a LEA gene in rice improves drought resistance under the field conditions. Theor. Appl. Genet. 2007, 115, 35–46. [Google Scholar] [CrossRef]
- Sivamani, E.; Bahieldin, A.; Wraith, J.M.; Al-Niemi, T.; Dyer, W.E.; Ho, T.H.D.; Qu, R. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 2000, 155, 1–9. [Google Scholar] [CrossRef]
- Babu, R.C.; Zhang, J.; Blum, A.; Ho, T.H.D.; Wu, R.; Nguyen, H.T. HVA1, a LEA gene from barley confers dehydration tolerance in transgenic rice (Oryza sativa L.) via cell membrane protection. Plant Sci. 2004, 166, 855–862. [Google Scholar] [CrossRef]
- Ahmed, C.B.; Rouina, B.B.; Sensoy, S.; Boukhris, M.; Abdallah, F.B. Changes in gas exchange, proline accumulation and antioxidative enzyme activities in three olive cultivars under contrasting water availability regimes. Environ Exper. Bot. 2009, 67, 345–352. [Google Scholar] [CrossRef]
- Koskeroglu, S.; Tuna, AL. The investigation on accumulation levels of proline and stress parameters of the maize (Zea mays L.) plants under salt and water stress. Acta. Physiol. Plant 2010, 32, 541–549. [Google Scholar] [CrossRef]
- Zhu, B.; Su, J.; Chang, M.; Verma, D.P.S.; Fan, Y.L.; Wu, R. Overexpression of a Δ1-pyrroline-5-carboxylate synthetase gene and analysis of tolerance to water-and salt-stress in transgenic rice. Plant Sci. 1998, 139, 41–48. [Google Scholar] [CrossRef]
- You, J.; Hu, H.; Xiong, L. An ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 2012, 197, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.C.; Oh, S.J.; Seo, J.S.; Choi, W.B.; Song, S.I.; Kim, C.H.; Kim, JK. Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth. Plant physiol. 2003, 131, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Capell, T.; Escobar, C.; Liu, H.; Burtin, D.; Lepri, O.; Christou, P. Over-expression of the oat arginine decarboxylase cDNA in transgenic rice (Oryza sativa L.) affects normal development patterns in vitro and results in putrescine accumulation in transgenic plants. Theor. Appl. Genet. 1998, 97, 246–254. [Google Scholar] [CrossRef]
- Du, H.; Wang, N.; Cui, F.; Li, X.; Xiao, J.; Xiong, L. Characterization of a β-carotene hydroxylase gene DSM2 conferring drought and oxidative stress resistance by increasing xanthophylls and ABA synthesis in rice. Plant Physiol. 2010, 154, 1304–1318. [Google Scholar] [CrossRef]
- You, J.; Zong, W.; Li, X.; Ning, J.; Hu, H.; Li, X.; Xiong, L. The SNAC1-targeted gene OsSRO1c modulates stomata closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J. Exp. Bot. 2013, 64, 569–583. [Google Scholar] [CrossRef]
- Phung, T.; Jung, H.I.; Park, J.; Kim, J.G.; Back, K.; Jung, S. Porphyrin biosynthesis control under water stress: Sustained porphyrin status correlates with drought tolerance in transgenic rice. Plant Physiol. 2011, 157, 1746–1764. [Google Scholar] [CrossRef]
- Li, H.W.; Zang, B.S.; Deng, X.W.; Wang, X.P. Overexpression of the trehalose-6-phosphate synthase gene OsTPS1 enhances abiotic stress tolerance in rice. Planta 2011, 234, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Wu, R. Stress-inducible synthesis of proline in transgenic rice confers faster growth under stress conditions than that with constitutive synthesis. Plant Sci. 2004, 166, 941–948. [Google Scholar] [CrossRef]
- Rohila, J.S.; Jain, R.K.; Wu, R. Genetic improvement of Basmati rice for salt and drought tolerance by regulated expression of a barley HVA1 cDNA. Plant Sci. 2002, 163, 525–532. [Google Scholar] [CrossRef]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.H.D.; Wu, R. Expression of a late embryogenesis abundant protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol. 1996, 110, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Song, S.I.; Kim, Y.S.; Jang, H.J.; Kim, S.Y.; Kim, M.; Kim, J.K. Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol. 2005, 138, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kim, Y.S.; Kwon, C.W.; Park, H.K.; Jeong, J.S.; Kim, J.K. Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Tang, N.; Du, H.; Ye, H.; Xiong, L. Characterization of OsbZIP23 as a key player of the basic leucine zipper transcription factor family for conferring abscisic acid sensitivity and salinity and drought tolerance in rice. Plant Physiol. 2008, 148, 1938–1952. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Gao, C.; Zheng, X.; Han, B. Identification of OsbZIP72 as a positive regulator of ABA response and drought tolerance in rice. Planta 2009, 229, 605–615. [Google Scholar] [CrossRef]
- Ito, Y.; Katsura, K.; Maruyama, K.; Taji, T.; Kobayashi, M.; Seki, M.; Yamaguchi-Shinozaki, K. Functional analysis of rice DREB1/CBF-type transcription factors involved in cold-responsive gene expression in transgenic rice. Plant Cell Physiol. 2006, 47, 141–153. [Google Scholar] [CrossRef]
- Bihani, P.; Char, B.; Bhargava, S. Transgenic expression of sorghum DREB2 in rice improves tolerance and yield under water limitation. J. Agr. Sci. 2011, 149, 95–101. [Google Scholar] [CrossRef]
- Oh, S.J.; Kwon, C.W.; Choi, D.W.; Song, S.I.; Kim, J.K. Expression of barley HvCBF4 enhances tolerance to abiotic stress in transgenic rice. Plant Biotechnol. J. 2007, 5, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiao, S.; Li, W.; Feng, W.; Li, J.; Wu, Z.; Shao, M. Overexpression of a Harpin-encoding gene hrf1 in rice enhances drought tolerance. J. Exp. Bot. 2011, 62, 4229–4238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, F.; Li, D.; Zhang, H.; Huang, R. Expression of ethylene response factor JERF1 in rice improves tolerance to drought. Planta 2010, 232, 765–774. [Google Scholar] [CrossRef]
- Quan, R.; Hu, S.; Zhang, Z.; Zhang, H.; Zhang, Z.; Huang, R. Overexpression of an ERF transcription factor TSRF1 improves rice drought tolerance. Plant Biotechnol. J. 2010, 8, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Wang, L.; Xu, Y.; Chen, N.; Ma, Q.; Li, F.; Chong, K. Overexpression of OsCOIN, a putative cold inducible zinc finger protein, increased tolerance to chilling, salt and drought, and enhanced proline level in rice. Planta 2007, 226, 1007–1016. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, V.; Gupta, A.K. Overexpression of OsiSAP8, a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol. Biol. 2008, 66, 445–462. [Google Scholar] [CrossRef]
- Bae, H.; Kim, S.K.; Cho, S.K.; Kang, B.G.; Kim, W.T. Overexpression of OsRDCP1, a rice RING domain-containing E3 ubiquitin ligase, increased tolerance to drought stress in rice (Oryza sativa L.). Plant Sci. 2011, 180, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Wu, Y.; Zhang, Y.; Liu, L.; Ning, Y.; Wang, D.; Xie, Q. OsSDIR1 overexpression greatly improves drought tolerance in transgenic rice. Plant Mol. Biol. 2011, 76, 145–156. [Google Scholar] [CrossRef]
- Miah, G.; Rafii, M.Y.; Ismail, M.R.; Sahebi, M.; Hashemi, F.S.G.; Yusuff, O.; Usman, M.G. Blast disease intimidation towards rice cultivation: A review of pathogen and strategies to control. J Anim. Plant Sci. 2017, 27, 1058–1066. [Google Scholar]
- Usman, M.G.; Rafii, M.Y.; Martini, M.Y.; Yusuff, O.A.; Ismail, M.R.; Miah, G. Introgression of heat shock protein (Hsp70 and sHsp) genes into the Malaysian elite chilli variety Kulai (Capsicum annuum L.) through the application of marker-assisted backcrossing (MAB). Cell Stress Chaperon 2018, 23, 223–234. [Google Scholar] [CrossRef]
- Chukwu, S.C.; Rafii, M.Y.; Ramlee, S.I.; Ismail, S.I.; Oladosu, Y.; Okporie, E.; Jalloh, M. Marker-assisted selection and gene pyramiding for resistance to bacterial leaf blight disease of rice (Oryza sativa L.). Biotechnol. Biotechnol. Equip. 2019. [Google Scholar] [CrossRef]
- Knapp, S.J. Marker-assisted selection as a strategy for increasing the probability of selecting superior genotypes. Crop Sci. 1998, 38, 1164–1174. [Google Scholar] [CrossRef]
- Lande, R.; Thompson, R. Efficiency of marker-assisted selection in the improvement of quantitative traits. Genetics 1990, 124, 743–756. [Google Scholar] [PubMed]
- Francia, E.; Tacconi, G.; Crosatti, C.; Barabaschi, D.; Bulgarelli, D.; Dall’Aglio, E.; Valè, G. Marker assisted selection in crop plants. Plant Cell Tissue Organ Cult. 2005, 82, 317–342. [Google Scholar] [CrossRef]
- Yang, W.; Duan, L.; Chen, G.; Xiong, L.; Liu, Q. Plant phenomics and high-throughput phenotyping: Accelerating rice functional genomics using multidisciplinary technologies. Curr Opin Plant Biol. 2013, 16, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Großkinsky, D.K.; Svensgaard, J.; Christensen, S.; Roitsch, T. Plant phenomics and the need for physiological phenotyping across scales to narrow the genotype-to-phenotype knowledge gap. J. Exp. Bot. 2015, 66, 5429–5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hairmansis, A.; Berger, B.; Tester, M.; Roy, S.J. Image-based phenotyping for non-destructive screening of different salinity tolerance traits in rice. Rice 2014, 7, 16. [Google Scholar] [CrossRef]
- Honsdorf, N.; March, T.J.; Berger, B.; Tester, M.; Pillen, K. High-throughput phenotyping to detect drought tolerance QTL in wild barley introgression lines. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Neilson, E.H.; Edwards, A.M.; Blomstedt, C.K.; Berger, B.; Moller, B.L.; Gleadow, R.M. Utilization of a high-throughput shoot imaging system to examine the dynamic phenotypic responses of a C4 cereal crop plant to nitrogen and water deficiency over time. J. Exp. Bot. 2015, 66, 1817–1832. [Google Scholar] [CrossRef]
- Miglani, G.S. Genome editing in crop improvement: Present scenario and future prospects. J. Crop Improv. 2017, 31, 453–559. [Google Scholar] [CrossRef]
- Bortesi, L.; Fischer, R. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol. Adv. 2015, 33, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Baltes, N.J.; Gil-Humanes, J.; Cermak, T.; Atkins, P.A.; Voytas, D.F. DNA replicons for plant genome engineering. Plant Cell 2014, 26, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Weeks, D.P.; Spalding, M.H.; Yang, B. Use of designer nucleases for targeted gene and genome editing in plants. Plant Biotechnol. J. 2016, 14, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Yang, Y.; Qin, R.; Li, H.; Qiu, C.; Li, L. Rapid improvement of grain weight via highly efficient CRISPR/Cas9-mediated multiplex genome editing in rice. J. Genet. Genomics 2016, 43, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Shim, J.S.; Oh, N.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Kim, J.K. Overexpression of OsNAC14 improves drought tolerance in rice. Front. Plant Sci. 2018, 9, 310. [Google Scholar] [CrossRef] [PubMed]
- Volaire, F.; Barkaoui, K.; Norton, M. Designing resilient and sustainable grasslands for a drier future: Adaptive strategies, functional traits and biotic interactions. Eur. J. Agron. 2014, 52, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Blum, A. Genomics for drought resistance–getting down to earth. Funct. Plant Biol. 2014, 41, 1191–1198. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Population | Marker | Type | QTL | References |
|---|---|---|---|---|---|
| Seedling drought resistance | Indica × Azucena | RFLP, AFLP & SSR | Recombinant inbred line | 7 | [43] |
| Cellular membrane stability | IR62266 × CT9993 | RFLP, AFLP & SSR | Doubled haploid line | 9 | [44] |
| Leaf water relations and rolling | Azucena × Bala | RFLP, AFLP & SSR | Recombinant inbred line | 13 | [45] |
| Seed fertility, spikelet per panicle and grain yield | Teqing × Lemont | SNP | Introgression lines | 5 | [46] |
| Root number, thickness, length, and penetration index | IR58821 × IR52561 | AFLP & RFLP | Recombinant inbred line | 28 | [47] |
| Root architecture and distribution | IR64 × Azucena | RFLP | Doubled haploid line | 39 | [48] |
| Root traits | IR1552 × Azucena | SSR | Recombinant inbred line | 23 | [49] |
| Deep roots | 3 populations | SSR, SNP | Recombinant inbred line | 6 | [50] |
| Root penetration, root number, and tiller number | CO39 × Moroberekan | RFLP | Recombinant inbred line | 39 | [51] |
| Root-penetration | Azucena × Bala | RFLP & AFLP | Recombinant inbred line | 18 | [52] |
| Grain yield under drought | Two population | SSR | Bulk-segregant and Selective genotyping | - | [53] |
| Grain yield in aerobic environments | Three populations | SSR | Bulk-segregant | 1 | [54] |
| Yield and yield traits at the reproductive stage | IR64 × Cabacu | SNP | Recombinant inbred line | 1 | [55] |
| Yield under reproductive stage stress over seasons | Swarna × WAB | SSR | Backcross inbred line | 1 | [56] |
| Heritability for grain yield | CT9993 × IR62266 | AFLP | Doubled haploid lines | 1 | [57] |
| Grain yield under severe lowland drought over environments | R77298 × Sabitri, | SSR | BC1 derived | 1 | [58] |
| Grain yield over years and location | Apo/2 × Swarna | SSR | Recombinant inbred lines | 1 | [59] |
| Yield at reproductive stage over environments | Two populations | SSR | Bulk-segregant analysis | 2 | [60] |
| Morphological and physiological traits | IR64 × Azucena | RFLP | Doubled haploid Lines | 15 | [61] |
| Dehydration avoidance | Bala × Azucena | RFLP, AFLP & SSR | Recombinant inbred lines | 17 | [62] |
| Osmotic adjustment and Dehydration tolerance | CO39 × Moroberekan | RFLP | Recombinant inbred line | 1 | [63] |
| Osmotic adjustment | CT9993 × IR62266 | RFLP, AFLP & SSR | Doubled haploid line | 5 | [64] |
| Gene Action | Gene | Promoter | Transformation | Phenotype | References |
|---|---|---|---|---|---|
| Genes Encoding Enzymes That Synthesize Osmotic and Other Protectants | |||||
| Arginine decarboxylase | ADC | CaMV35S | Biolistic | Reduction in chlorophyll loss under water deficiency | [79] |
| Polyamine synthesis | ADC | Ubi-1 | Agrobacterium | Improved drought tolerance by producing higher levels of putrescine and spermine synthesis. | [16] |
| abscisic acid Metabolism | CaMV35SP | DSM2 | Agrobacterium | Oxidative and drought stress resistance and increase of the xanthophylls and non-photochemical quenching. | [80] |
| Amino acid metabolism | OsOAT | Ubi1 | Agrobacterium | Improve drought tolerance and increase seed setting | [77] |
| Reactive oxygen species scavenging | OsSRO1c | Ubi1 | Agrobacterium | Oxidative stress tolerance and stomata closure regulation | [81] |
| Protoporphyrinogen oxidase | PPO | - | Agrobacterium | Less oxidative damage, and drought tolerance | [82] |
| Trehalose synthesis | OsTPS1 | Actin1 | Agrobacterium | Tolerance of rice seedling to drought, cold, and high salinity | [83] |
| Trehalose synthesis | TPSP | Ubi1 | Agrobacterium | Cold, salt and drought tolerance expressed by chlorophyll fluorescence | [78] |
| Proline synthesis | P5CS | Act1 | Agrobacterium | Resistance to water and salinity stress | [84] |
| Proline synthesis | P5CS | AIPC | Biolistic | Increased biomass production under salinity and drought stresses | [76] |
| Late Embryogenesis Abundant (LEA) Related Genes | |||||
| LEA protein gene | HVA1 | Actin1 | Agrobacterium | Cell membrane stability, higher leaf relative water content (RWC) and increase in growth under drought stress. | [73] |
| HVA1 | Actin1 | Agrobacterium | Drought and salinity tolerance | [85] | |
| HVA1 | Actin 1 | Biolistic | Increased drought tolerance and salinity stress. | [86] | |
| OsLEA3-1 | rice LEA3-1 | Agrobacterium | Drought resistance for grain yield under field conditions | [71] | |
| OsLEA3-2 | CaMV35S | Agrobacterium | Drought resistance and increase grain per panicle | [70] | |
| Various Regulatory Genes | |||||
| Transcription factor | ABF3 | Ubi1 | Agrobacterium | Improved salinity and drought tolerance | [87] |
| AP37 | OsCc1 | Agrobacterium | Improve growth performance under drought stress | [88] | |
| OsbZIP23 | Ubi1 | Agrobacterium | Wide spectrum to salt and drought tolerance and improvement in yield. | [89] | |
| OsbZIP72 | CaMV35S | Agrobacterium | Drought resistance and ABA sensitivity | [90] | |
| DREB1 or OsDREB1 | CaMV 35S | Agrobacterium | Tolerance to water deficient, low-temperature and high-salt stresses | [91] | |
| DREB2 | rd29A | Agrobacterium | Improve grain yield under drought stress | [92] | |
| HvCBF4 | Ubi1 | Agrobacterium | Tolerance to drought, high-salinity, and low-temperature. | [93] | |
| Harpin protein | Hrf1 | CaMV 35S | Agrobacterium | Drought resistance through ABA signalling and antioxidants, and stomata closure regulation | [94] |
| Jasmonate and ethylene-responsive factor 1 | JERF1 | CaMV35S | Agrobacterium | Drought resistance | [95] |
| Ethylene-responsive factor 1 | TSRF1 | - | Agrobacterium | Enhances the osmotic and drought tolerance | [96] |
| RING finger protein | OsCOIN | CaMV35S | Agrobacterium | Cold, salt and drought tolerance | [97] |
| Stress/zinc finger protein | OsiSAP8 | - | Agrobacterium | Tolerance to salt, drought and cold stress | [98] |
| Protein degradation (E3 ubiquitin ligase) | OsRDCP1 | CaMV35S | Agrobacterium | Improved tolerance to drought stress | [99] |
| OsSDIR1 | Ubi1 | Agrobacterium | Stomata regulation under drought stress | [100] | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oladosu, Y.; Rafii, M.Y.; Samuel, C.; Fatai, A.; Magaji, U.; Kareem, I.; Kamarudin, Z.S.; Muhammad, I.; Kolapo, K. Drought Resistance in Rice from Conventional to Molecular Breeding: A Review. Int. J. Mol. Sci. 2019, 20, 3519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143519
Oladosu Y, Rafii MY, Samuel C, Fatai A, Magaji U, Kareem I, Kamarudin ZS, Muhammad I, Kolapo K. Drought Resistance in Rice from Conventional to Molecular Breeding: A Review. International Journal of Molecular Sciences. 2019; 20(14):3519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143519
Chicago/Turabian StyleOladosu, Yusuff, Mohd Y. Rafii, Chukwu Samuel, Arolu Fatai, Usman Magaji, Isiaka Kareem, Zarifth Shafika Kamarudin, Isma’ila Muhammad, and Kazeem Kolapo. 2019. "Drought Resistance in Rice from Conventional to Molecular Breeding: A Review" International Journal of Molecular Sciences 20, no. 14: 3519. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20143519