An Insight into the Proteome of Uveal Melanoma-Derived Ectosomes Reveals the Presence of Potentially Useful Biomarkers

 ,
,  and
and
Abstract
:1. Introduction
2. Results
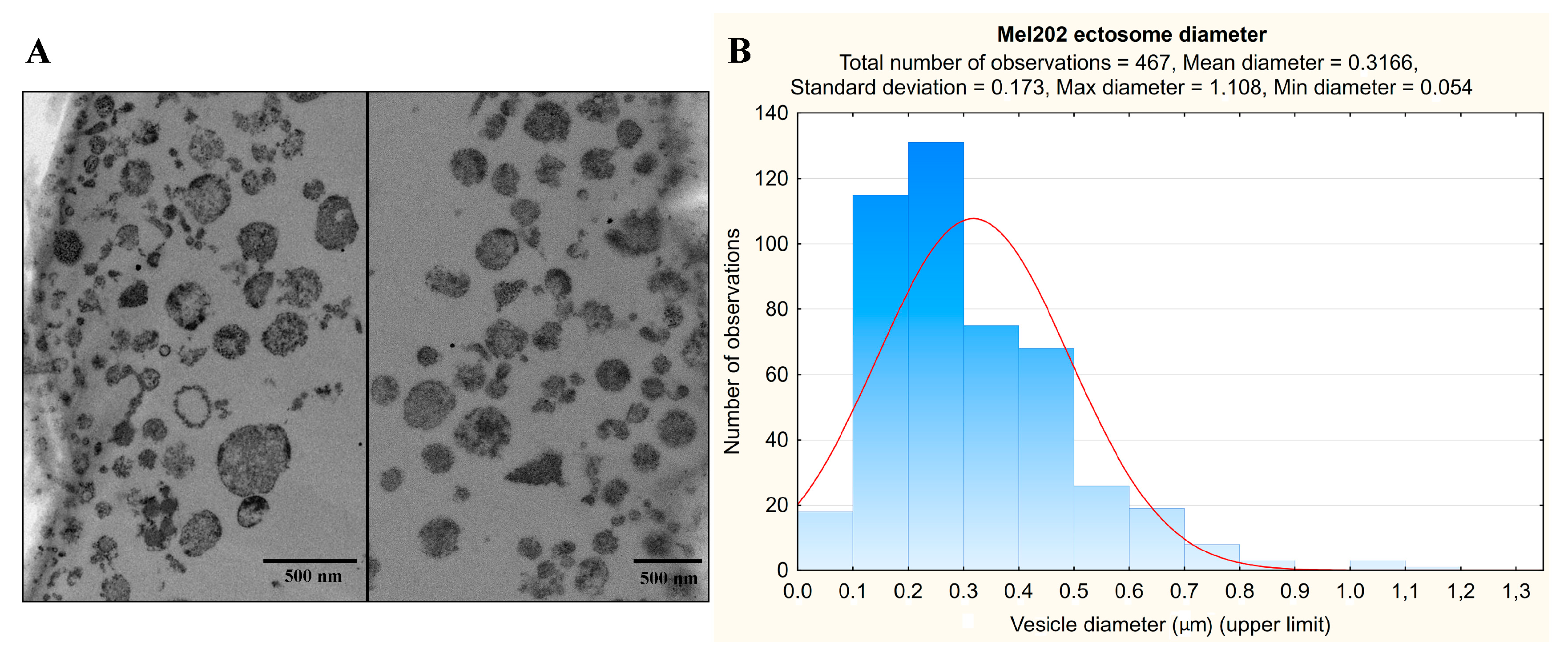
2.1. Transmission Electron Microscopy (TEM) of Mel202-Derived Ectosomes
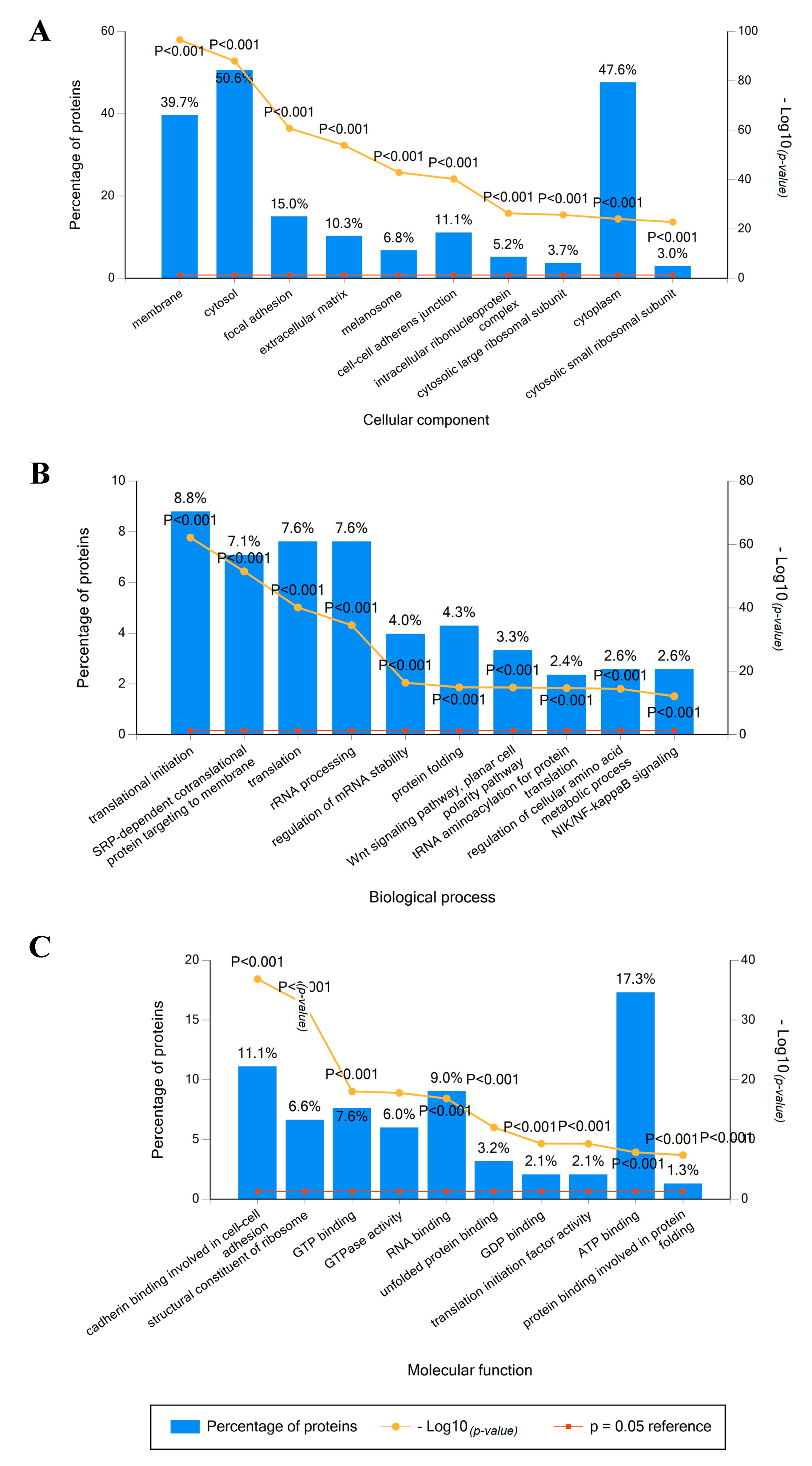
2.2. Identified Proteins in Mel202-Derived Ectosomes and Their Functional Classification
2.3. Cancer-Associated Proteins in Mel202-Derived Ectosomes
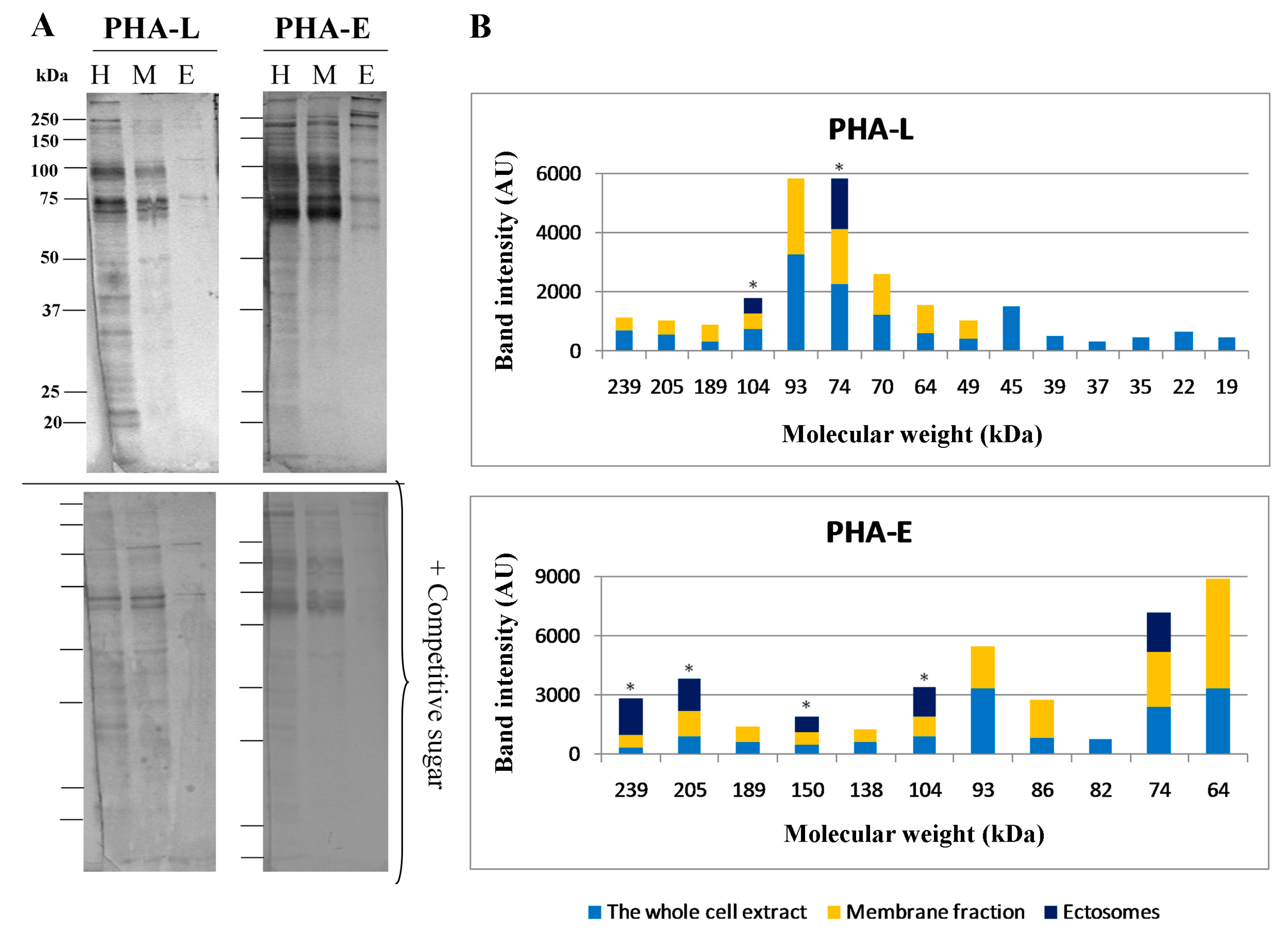
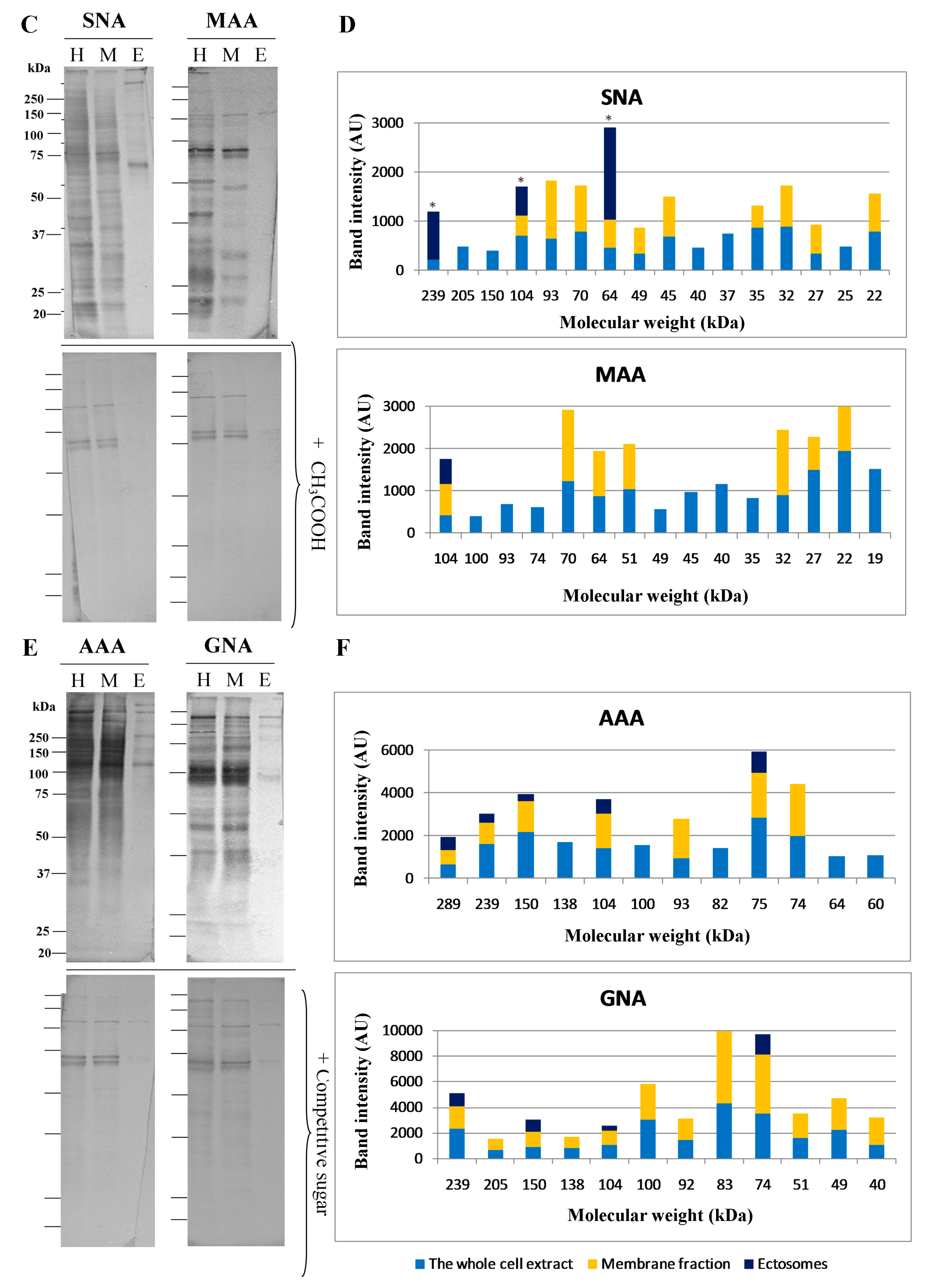
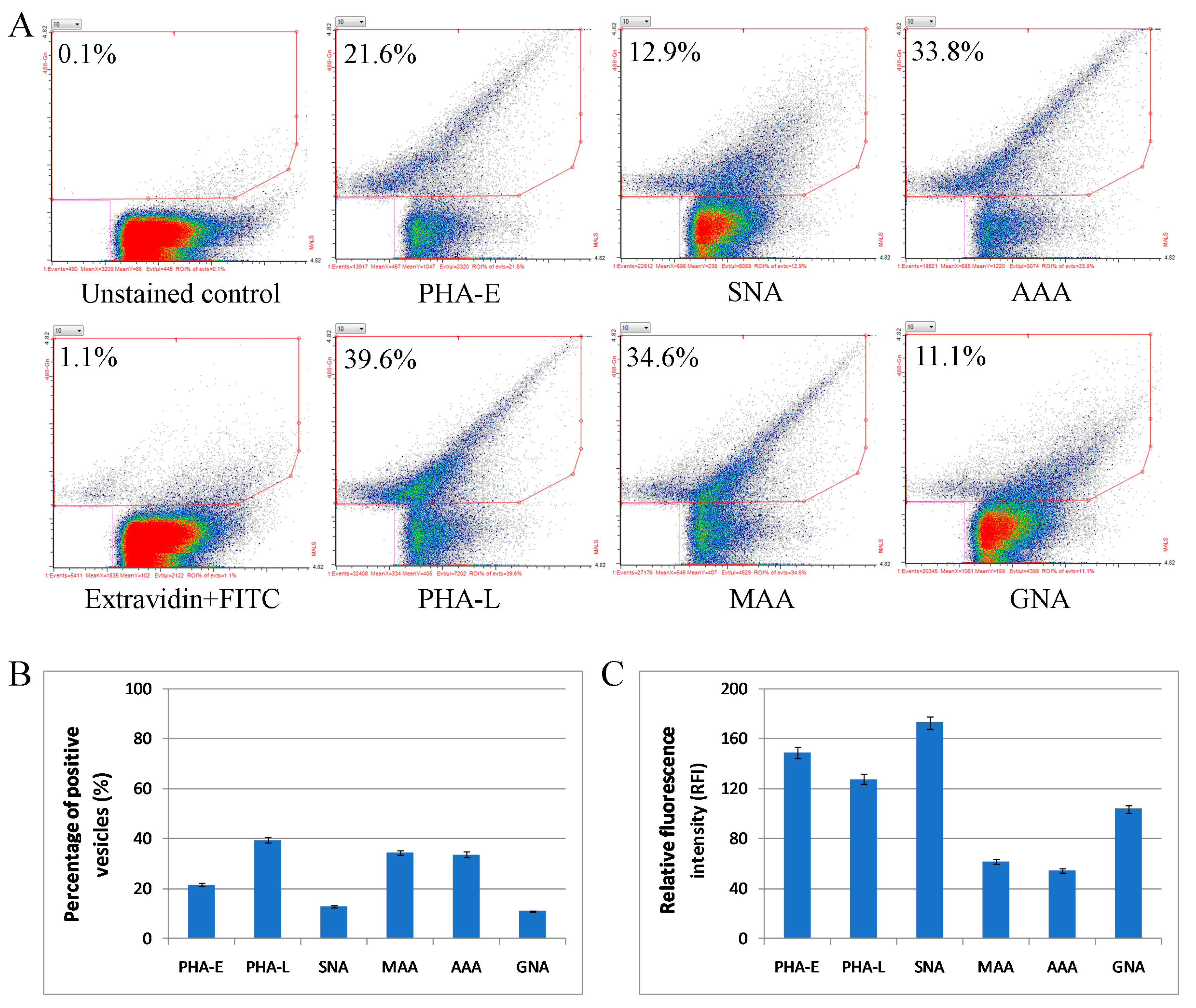
2.4. Glycosylation of Mel202-Derived Ectosomes and the Parental Cell Membranes
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture Conditions
4.2. Isolation of Ectosomes and Verification of the Purity of the Sample
4.3. Mass Spectrometry Analysis
4.3.1. Ectosome Lysis
4.3.2. Sample Preparation for Mass Spectrometric Analysis
4.3.3. Liquid Chromatography and Tandem Mass Spectrometry (LC-MS/MS)
4.3.4. Analysis of the Proteomic Data
4.3.5. Bioinformatic Analysis
4.4. Lectin-Based Analysis of Mel202 Cell Extracts, Cell Membranes, and Ectosomes
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AAA | Aleuria aurantia agglutinin |
| BCIP | 5-bromo-4-chloro-3-indolyl-phosphate |
| EV | extracellular vesicles |
| Gal-3 | galectin-3 |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| GNA | Galanthus nivalis agglutinin |
| HSP | heat shock protein |
| L1CAM | L1 cell adhesion molecule |
| LC-MS/MS | liquid chromatography coupled with mass spectrometry |
| LGALS3BP | galectin-3-binding protein |
| MAA | Maackia amurensis agglutinin |
| MDR | multidrug resistance |
| MMP | matrix metalloproteinase |
| NBT | 4-nitro blue tetrazolium chloride |
| PHA-E | Phaseolus vulgaris erythroagglutinin |
| PHA-L | Phaseolus vulgaris leukoagglutinin |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| SNA | Sambucus nigra agglutinin |
| TEM | transmission electron microscopy |
| UM | uveal melanoma |
References
- Blum, E.S.; Yang, J.; Komatsubara, K.M.; Carvajal, R.D. Clinical management of uveal and conjunctival melanoma. Oncology 2016, 30, 29–43. [Google Scholar] [PubMed]
- Goh, A.Y.; Layton, C.J. Evolving systemic targeted therapy strategies in uveal melanoma and implications for ophthalmic management: A review. Clin. Exp. Ophthalmol. 2016, 44, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; García, A.; Antrobus, R.; Blanco, M.J.; Dwek, R.A.; Zitzmann, N. Biomarker discovery from uveal melanoma secretomes: Identification of gp100 and cathepsin D in patient serum. J. Proteome Res. 2007, 6, 2802e2811. [Google Scholar] [CrossRef] [PubMed]
- Abildgaard, S.K.; Vorum, H. Proteomics of uveal melanoma: A minireview. J. Oncol. 2013, 2013, 820953. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, P.; Murphy, C.C.; Clynes, M.; Horgan, N.; Moriarty, P.; Tiernan, D.; Beatty, S.; Kennedy, S.; Meleady, P. Proteomics in uveal melanoma. Exp. Eye Res. 2014, 118, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Angi, M.; Kalirai, H.; Prendergast, S.; Simpson, D.; Hammond, D.E.; Madigan, M.C.; Beynon, R.J.; Coupland, S.E. In-depth proteomic profiling of the uveal melanoma secretome. Oncotarget 2016, 7, 49623–49635. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; García, A.; Thomas, B.; Piñeiro, A.; Akoulitchev, A.; Dwek, R.A.; Zitzmann, N. The characterization of the invasion phenotype of uveal melanoma tumour cells shows the presence of MUC18 and HMG-1 metastasis markers and leads to the identification of DJ-1 as a potential serum biomarker. Int. J. Cancer 2006, 119, 1014e1022. [Google Scholar] [CrossRef] [PubMed]
- Zuidervaart, W.; Hensbergen, P.J.; Wong, M.C.; Deelder, A.M.; Tensen, C.P.; Jager, M.J.; Gruis, N.A. Proteomic analysis of uveal melanoma reveals novel potential markers involved in tumor progression. Investig. Ophthalmol. Vis. Sci. 2006, 47, 786e793. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Bing, Z.; Zhang, Y.; Ao, B.; Zhang, S.; Ye, C.; He, J.; Ding, N.; Ye, W.; Xiong, J.; et al. Quantitative proteomic analysis for radiation-induced cell cycle suspension in 92-1 melanoma cell line. J. Radiat. Res. 2013, 54, 649e662. [Google Scholar] [CrossRef]
- Yan, L.B.; Shi, K.; Bing, Z.T.; Sun, Y.L.; Shen, Y. Proteomic analysis of energy metabolism and signal transduction in irradiated melanoma cells. Int. J. Ophthalmol. 2013, 6, 286e294. [Google Scholar]
- Crabb, J.W.; Hu, B.; Crabb, J.S.; Triozzi, P.; Saunthararajah, Y.; Tubbs, R.; Singh, A.D. iTRAQ quantitative proteomic comparison of metastatic and non-metastatic uveal melanoma tumors. PLoS ONE 2015, 10, e0135543. [Google Scholar] [CrossRef] [PubMed]
- Linge, A.; Kennedy, S.; O’Flynn, D.; Beatty, S.; Moriarty, P.; Henry, M.; Clynes, M.; Larkin, A.; Meleady, P. Differential expression of fourteen proteins between uveal melanoma from patients who subsequently developed distant metastases versus those who did not. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4634e4643. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular vesicles in cancer: Cell-to-cell mediators of metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Surman, M.; Stępień, E.; Hoja-Łukowicz, D.; Przybyło, M. Deciphering the role of ectosomes in cancer development and progression: Focus on the proteome. Clin. Exp. Metastasis 2017, 34, 273–289. [Google Scholar] [CrossRef] [PubMed]
- Surman, M.; Hoja-Łukowicz, D.; Szwed, S.; Drożdż, A.; Stępień, E.; Przybyło, M. Human melanoma-derived ectosomes are enriched with specific glycan epitopes. Life Sci. 2018, 207, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Vega, V.L.; Rodríguez-Silva, M.; Frey, T.; Gehrmann, M.; Diaz, J.C.; Steinem, C.; Multhoff, G.; Arispe, N.; De Maio, A. Hsp70 translocates into the plasma membrane after stress and is released into the extracellular environment in a membrane-associated form that activates macrophages. J. Immunol. 2008, 180, 4299–4307. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.; Barbagallo, C.; Statello, L.; Caltabiano, R.; Russo, A.; Puzzo, L.; Avitabile, T.; Longo, A.; Toro, M.D.; Barbagallo, D.; et al. miRNA profiling in vitreous humor, vitreal exosomes and serum from uveal melanoma patients: Pathological and diagnostic implications. Cancer Biol. Ther. 2015, 16, 1387–1396. [Google Scholar] [CrossRef]
- Eldh, M.; Olofsson Bagge, R.; Lässer, C.; Svanvik, J.; Sjöstrand, M.; Mattsson, J.; Lindnér, P.; Choi, D.S.; Gho, Y.S.; Lötvall, J.; et al. MicroRNA in exosomes isolated directly from the liver circulation in patients with metastatic uveal melanoma. BMC Cancer 2014, 14, 962. [Google Scholar] [CrossRef]
- Ang, E.Z.; Nguyen, H.T.; Sim, H.L.; Putti, T.C.; Lim, L.H. Annexin A1 regulates growth arrest induced by high levels of estrogen in MCF-7 breast cancer cells. Mol. Cancer Res. 2009, 7, 266–274. [Google Scholar] [CrossRef]
- Hendrix, M.J.; Seftor, E.A.; Seftor, R.E.; Gardner, L.M.; Boldt, H.C.; Meyer, M.; Pe’er, J.; Folberg, R. Biologic determinants of uveal melanoma metastatic phenotype: Role of intermediate filaments as predictive markers. Lab. Investig. 1998, 78, 153–163. [Google Scholar]
- Seftor, E.A.; Meltzer, P.S.; Kirschmann, D.A.; Pe’er, J.; Maniotis, A.J.; Trent, J.M.; Folberg, R.; Hendrix, M.J. Molecular determinants of human uveal melanoma invasion and metastasis. Clin. Exp. Metastasis 2002, 19, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Gangemi, R.; Mirisola, V.; Barisione, G.; Fabbi, M.; Brizzolara, A.; Lanza, F.; Mosci, C.; Salvi, S.; Gualco, M.; Truini, M.; et al. Mda-9/syntenin is expressed in uveal melanoma and correlates with metastatic progression. PLoS ONE 2012, 7, e29989.8. [Google Scholar] [CrossRef] [PubMed]
- Mourad-Zeidan, A.A.; Melnikova, V.O.; Wang, H.; Raz, A.; Bar-Eli, M. Expression profiling of Galectin-3-depleted melanoma cells reveals its major role in melanoma cell plasticity and vasculogenic mimicry. Am. J. Pathol. 2008, 173, 1839–1852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Kim, S.J.; Baek, J.H.; Lee, H.W.; Jeong, S.Y.; Chun, K.H. Galectin-3 increases the motility of mouse melanoma cells by regulating matrix metalloproteinase-1 expression. Exp. Mol. Med. 2012, 44, 387–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshaw, S.R.; Sisley, K.; Cross, N.; Murray, A.K.; MacNeil, S.M.; Wagner, M.; Nichols, C.E.; Rennie, I.G. A comparison of ocular melanocyte and uveal melanoma cell invasion and the implication of α1β1, α4β1 and α6β1 integrins. Br. J. Ophthalmol. 2001, 85, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Lüke, J.; Vukoja, V.; Brandenbusch, T.; Nassar, K.; Rohrbach, J.M.; Grisanti, S.; Lüke, M.; Tura, A. CD147 and matrix-metalloproteinase-2 expression in metastatic and non-metastatic uveal melanomas. BMC Ophthalmol. 2016, 16, 74. [Google Scholar] [CrossRef] [PubMed]
- Jorfi, S.; Ansa-Addo, E.A.; Kholia, S.; Stratton, D.; Valley, S.; Lange, S.; Inal, J. Inhibition of microvesiculation sensitizes prostate cancer cells to chemotherapy and reduces docetaxel dose required to limit tumor growth in vivo. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef]
- Walsh, N.; Kennedy, S.; Larkin, A.M.; Tryfonopoulos, D.; Eustace, A.J.; Mahgoub, T.; Conway, C.; Oglesby, I.; Collins, D.; Ballot, J.; et al. Membrane transport proteins in human melanoma: Associations with tumour aggressiveness and metastasis. Br. J. Cancer 2010, 102, 1157–1162. [Google Scholar] [CrossRef]
- Bande, M.F.; Santiago, M.; Blanco, M.J.; Mera, P.; Capeans, C.; Rodríguez-Alvarez, M.X.; Pardo, M.; Piñeiro, A. Serum DJ-1/PARK 7 is a potential biomarker of choroidal nevi transformation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 62–67. [Google Scholar] [CrossRef]
- Batista, B.S.; Eng, W.S.; Pilobello, K.T.; Hendricks-Munoz, K.D.; Mahal, L.K. Identification of a conserved glycan signature for microvesicles. J. Proteome Res. 2011, 10, 4624–4633. [Google Scholar] [CrossRef]
- Liang, Y.; Eng, W.S.; Colquhoun, D.R.; Dinglasan, R.R.; Graham, D.R.; Mahal, L.K. Complex N-linked glycans serve as a determinant for exosome/microvesicle cargo recruitment. J. Biol. Chem. 2014, 289, 32526–32537. [Google Scholar] [CrossRef] [PubMed]
- Escrevente, C.; Grammel, N.; Kandzia, S.; Zeiser, J.; Tranfield, E.M.; Conradt, H.S.; Costa, J. Sialoglycoproteins and N-glycans from secreted exosomes of ovariancarcinoma cells. PLoS ONE 2013, 8, e78631. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.; Gomes-Alves, P.; Carvalho, S.B.; Peixoto, C.; Alves, P.M.; Altevogt, P.; Costa, J. Extracellular vesicles from ovarian carcinoma cells display specific glycosignatures. Biomol. Ther. 2015, 5, 1741–1761. [Google Scholar] [CrossRef] [PubMed]
- Przybyło, M.; Pocheć, E.; Link-Lenczowski, P.; Lityńska, A. β1-6 branching of cell surface glycoproteins may contribute to uveal melanoma progression by up-regulating cell motility. Mol. Vis. 2008, 14, 625–636. [Google Scholar] [PubMed]
- Janik, M.E.; Lityńska, A.; Przybyło, M. Studies on primary uveal and cutaneous melanoma cell interaction with vitronectin. Cell Biol. Int. 2014, 38, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Pocheć, E.; Rydlewska, M.; Przybyło, M.; Lityńska, A. Diverse expression of N-acetylglucosaminyltransferase V and complex-type β1,6-branched N-glycans in uveal and cutaneous melanoma cells. Acta Biochim. Pol. 2015, 62, 323–328. [Google Scholar] [CrossRef] [PubMed]
- Costa, J. Glycoconjugates from extracellular vesicles: Structures, functions and emerging potential as cancer biomarkers. Biochim. Biophys. Acta Rev. Cancer 2017, 1968, 157–166. [Google Scholar] [CrossRef]
- Hughes, C.S.; Foehr, S.; Garfield, D.A.; Furlong, E.E.; Steinmetz, L.M.; Krijgsveld, J. Ultrasensitive proteome analysis using paramagnetic bead technology. Mol. Syst. Biol. 2014, 10, 757. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Csordas, A.; del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 2016, 44, D447–D456. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Ang, C.S.; Gangoda, L.; Quek, C.M.J.; Williamson, N.J.; Mouradov, D.; Sieber, O.M.; Simpson, R.J.; Salim, A.; et al. FunRich: A standalone tool for functional enrichment analysis. Proteomics 2015, 15, 2597–2601. [Google Scholar] [CrossRef]
- Pathan, M.; Keerthikumar, S.; Chisanga, D.; Alessandro, R.; Ang, C.S.; Askenase, P.; Batagov, A.O.; Benito-Martin, A.; Camussi, G.; Clayton, A.; et al. A novel community driven software for functional enrichment analysis of extracellular vesicles data. J. Extracell. Vesicles 2017, 1, 1321455. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Proliferation/Apoptosis | Cell Invasion and Metastasis | Cancer Cell Metabolism | Heat Shock Proteins and Chaperones | Drug Resistance | Others |
|---|---|---|---|---|---|
| Annexin A1 Annexin A2 Chloride intracellular channel protein 1 Heterogeneous nuclear ribonucleoprotein K Heterogeneous nuclear ribonucleoproteins C1/C2 Peptidyl-prolyl cis-trans isomerase A Peroxiredoxin-1 Peroxiredoxin-2 Peroxiredoxin-4 Peroxiredoxin-6 Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha isoform Stress-induced-phosphoprotein 1 T-complex protein 1 subunit beta | 14-3-3 protein zeta/delta Calreticulin Calumenin Cathepsin B Cathepsin D CD44 antigen Cell surface glycoprotein MUC18 Cofilin-1 Filamin-A Galectin-1 Galectin-3 Galectin-3-binding protein Histone H4 Integrin alpha-V Integrin beta-1 Keratin, type II cytoskeletal 5 Nucleoside diphosphate kinase A Phosphatidylethanolamine-binding protein 1 Profilin-1 Ras GTPase-activating protein-binding protein 1 Rho GDP-dissociation inhibitor 1 Serpin H1 Syntenin-1 Spectrin beta chain Superoxide dismutase [Cu-Zn] Vimentin WD repeat-containing protein 1 | Apolipoprotein E Fatty acid synthase Fructose-bisphosphate aldolase A Glyceraldehyde-3-phosphate dehydrogenase Isocitrate dehydrogenase (NADP) cytoplasmic Lactoylglutathione lyase L-lactate dehydrogenase A chain Nicotinamide phosphoribosyltransferase Phosphoglycerate kinase 1 Pyruvate kinase isozymes M1/M2 | 60 kDa heat shock protein, mitochondrial precursor Alpha-crystallin B chain Endoplasmin Heat shock 70 kDa protein 1A/1B Heat shock 70 kDa protein 4 Heat shock protein beta-1 Heat shock protein HSP 90-beta | Glutathione S-transferase omega-1 Multidrug resistance-associated protein 1 Proliferating cell nuclear antigen | Elongation factor 1-beta Epoxide hydrolase 1 Glutathione S-transferase P Heterogeneous nuclear ribonucleoproteins A2/B1 Nucleophosmin Transforming growth factor-beta-induced protein ig-h3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surman, M.; Hoja-Łukowicz, D.; Szwed, S.; Kędracka-Krok, S.; Jankowska, U.; Kurtyka, M.; Drożdż, A.; Lityńska, A.; Stępień, E.; Przybyło, M. An Insight into the Proteome of Uveal Melanoma-Derived Ectosomes Reveals the Presence of Potentially Useful Biomarkers. Int. J. Mol. Sci. 2019, 20, 3789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153789
Surman M, Hoja-Łukowicz D, Szwed S, Kędracka-Krok S, Jankowska U, Kurtyka M, Drożdż A, Lityńska A, Stępień E, Przybyło M. An Insight into the Proteome of Uveal Melanoma-Derived Ectosomes Reveals the Presence of Potentially Useful Biomarkers. International Journal of Molecular Sciences. 2019; 20(15):3789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153789
Chicago/Turabian StyleSurman, Magdalena, Dorota Hoja-Łukowicz, Sabina Szwed, Sylwia Kędracka-Krok, Urszula Jankowska, Magdalena Kurtyka, Anna Drożdż, Anna Lityńska, Ewa Stępień, and Małgorzata Przybyło. 2019. "An Insight into the Proteome of Uveal Melanoma-Derived Ectosomes Reveals the Presence of Potentially Useful Biomarkers" International Journal of Molecular Sciences 20, no. 15: 3789. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20153789