Insights into the Relationships Between Herbicide Activities, Molecular Structure and Membrane Interaction of Cinnamon and Citronella Essential Oils Components

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
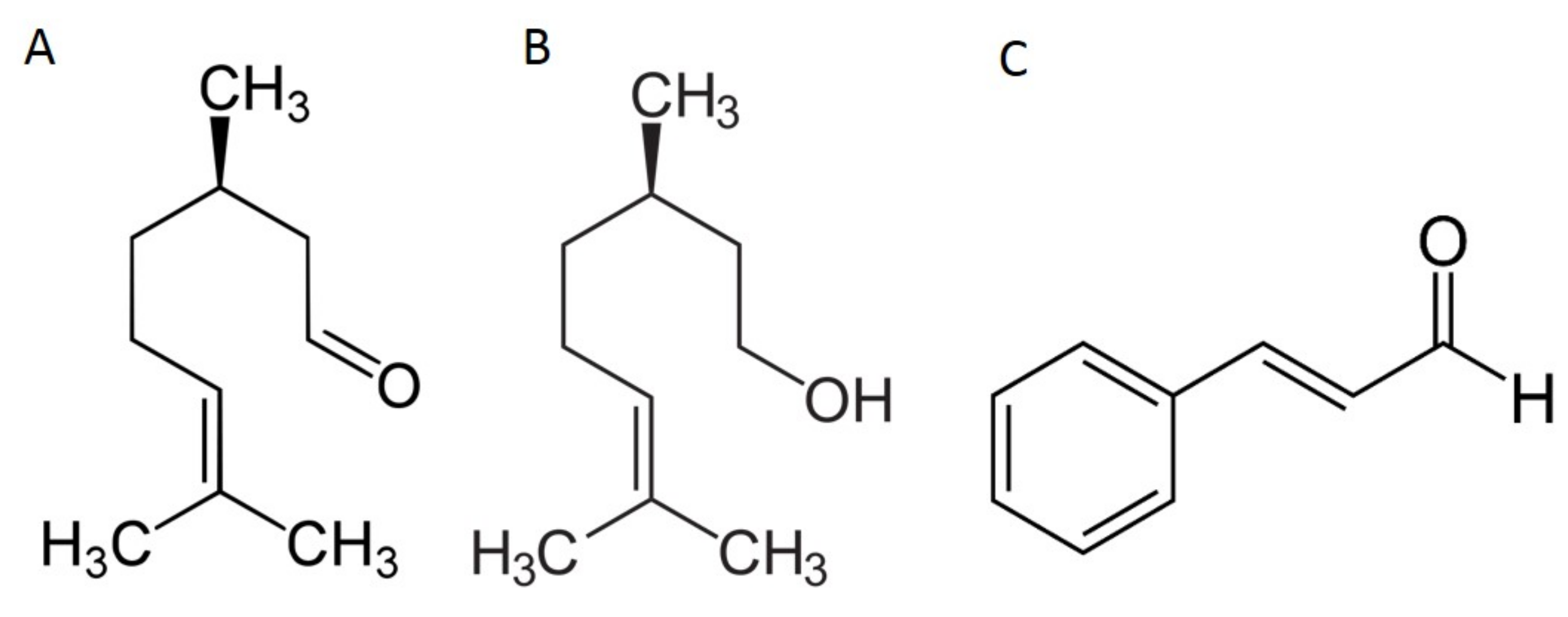
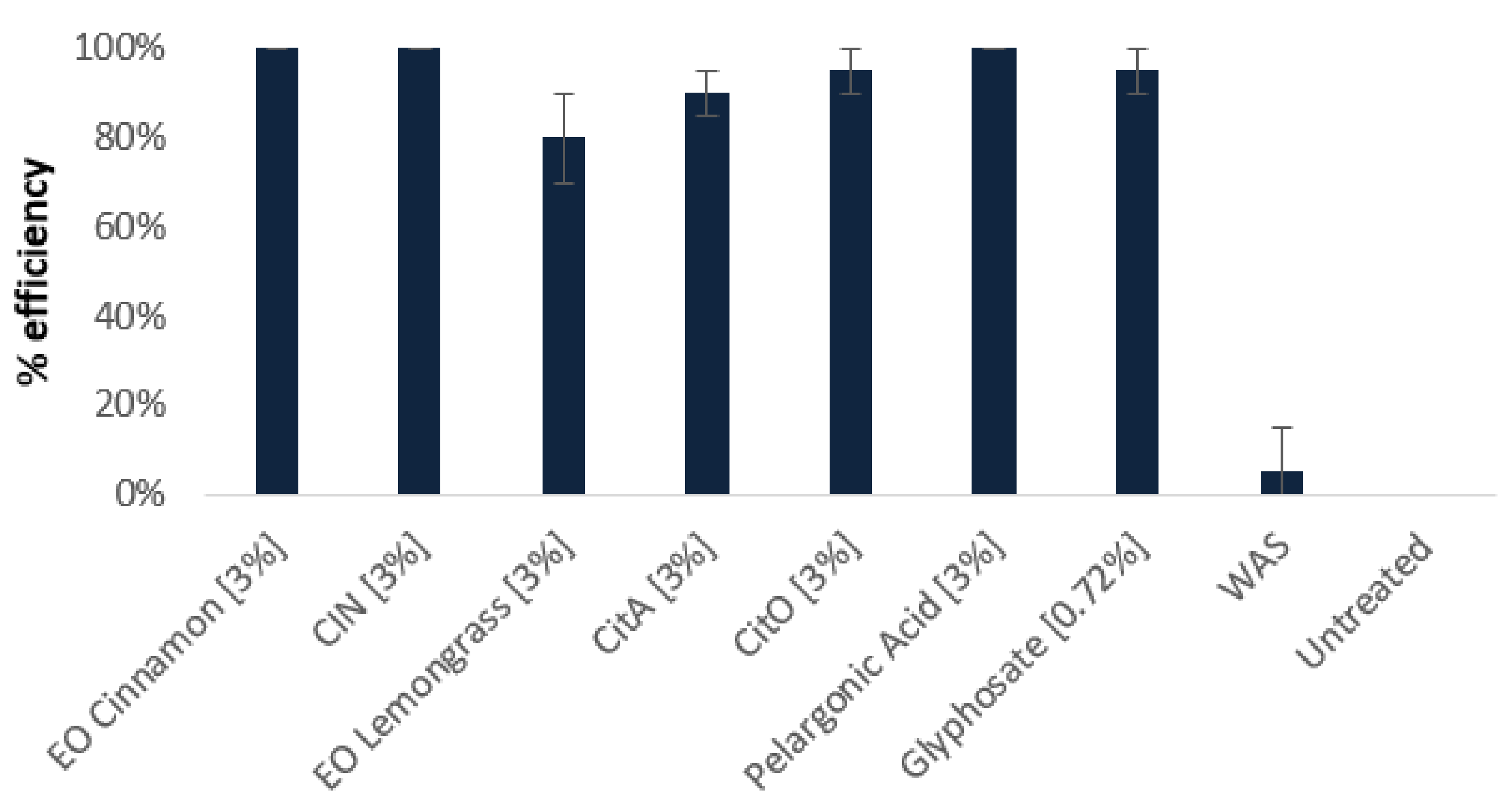
2.1. Herbicidal Effects of Cinnamon and Java Citronella Oils and Their Main Components
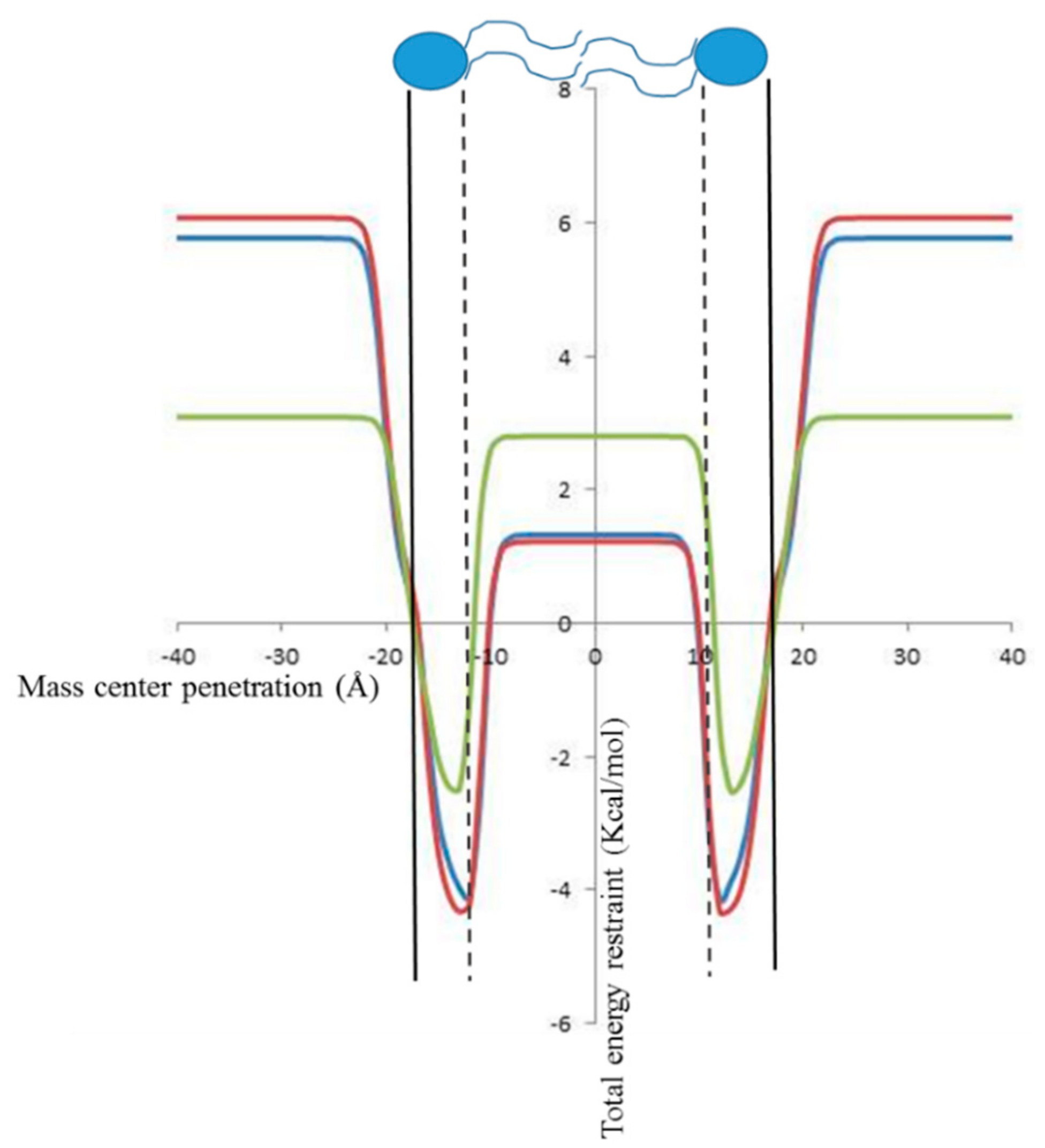
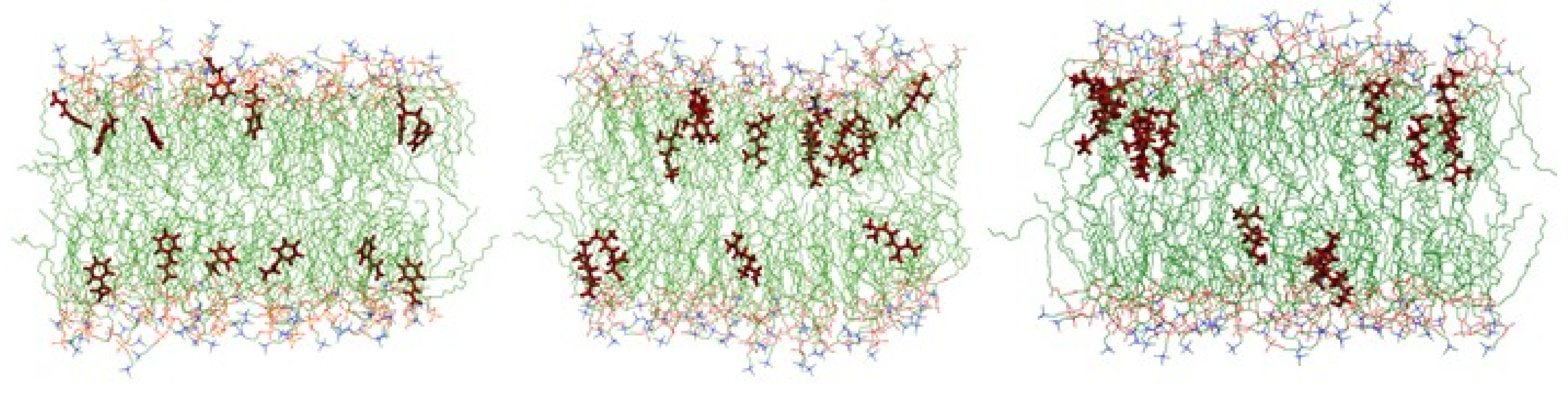
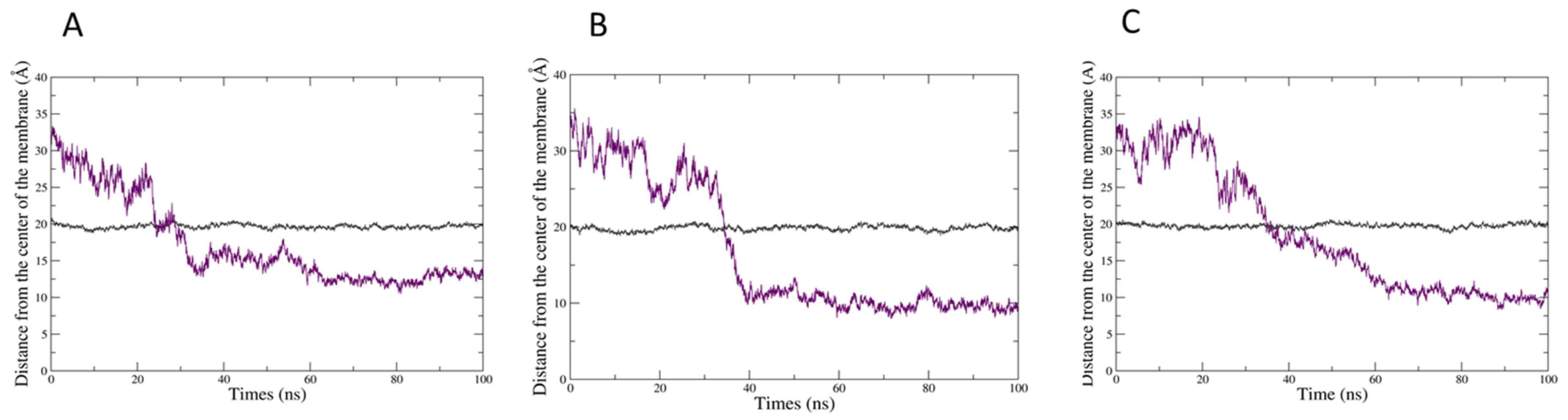
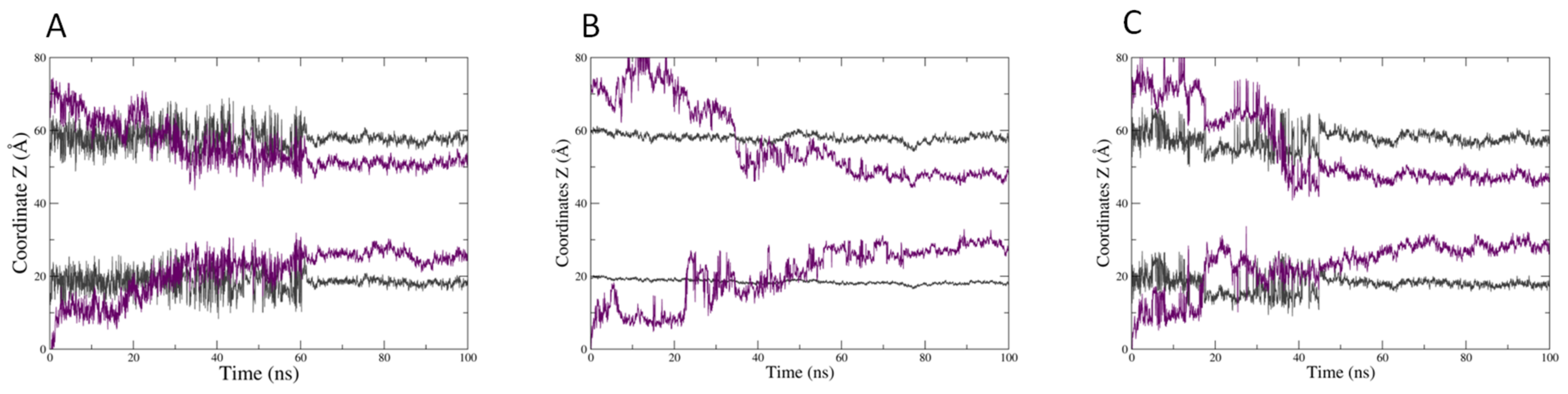
2.2. In Silico Prediction of The Insertion of CIN, CitO and CitA in Model Membranes
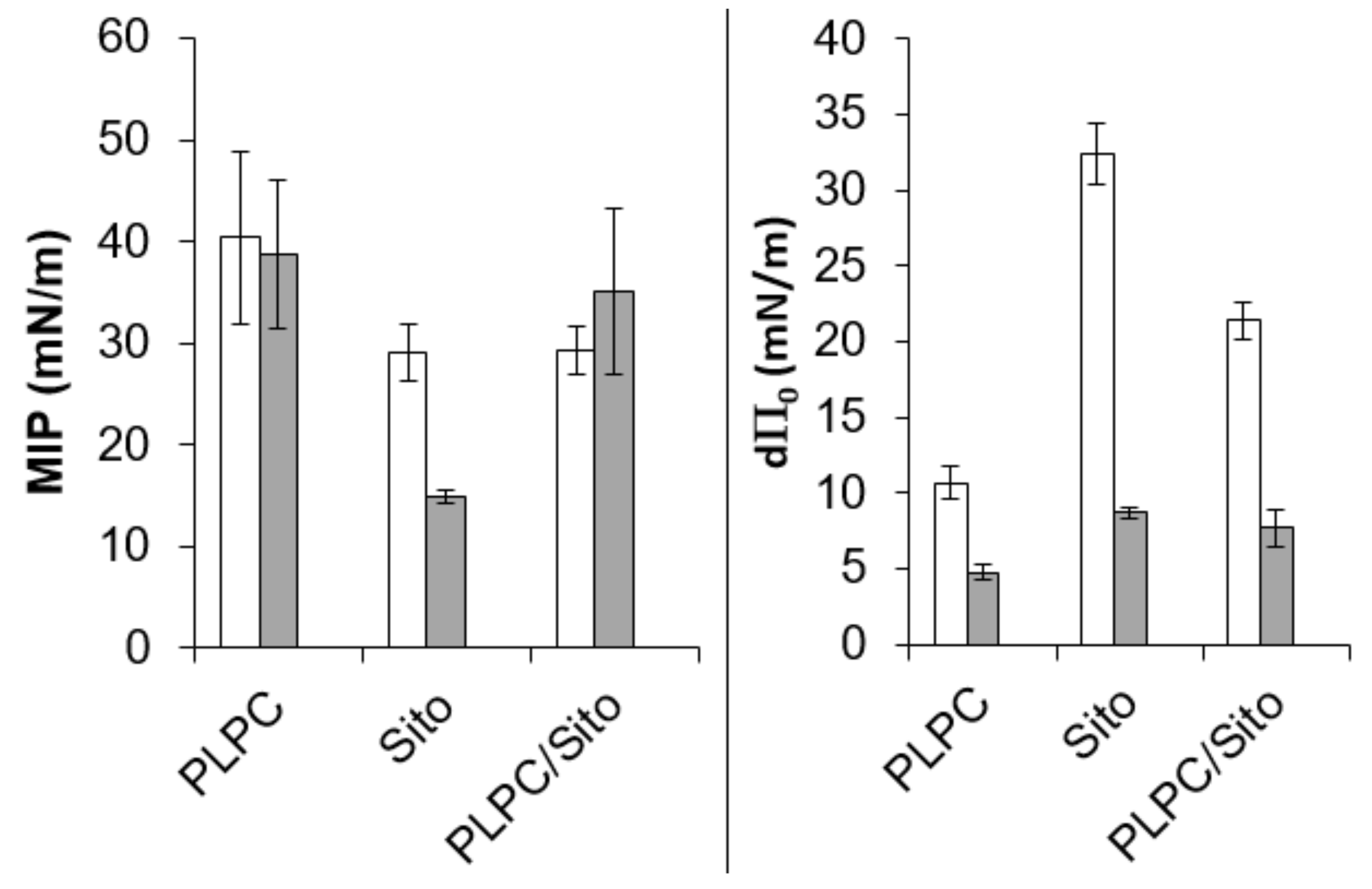
2.3. In Vitro Biophysical Assays
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Chromatographic Analysis of Cinnamon and Java Citronella Essential Oils
4.3. Penetration and Stability of Essential Oils and Pure Compounds Tests
4.4. Herbicide Tests on A. Thaliana
4.5. In Silico Approaches to Study the Interaction of the EOs Main Components with Lipids
4.6. Molecular Dynamics Simulations
4.7. In Vitro Biophysical Assays to Study the Interaction of CIN, CitO and CitA with Model PPM
4.7.1. Liposome Preparation
4.7.2. Isothermal Titration Calorimetry
4.7.3. Adsorption Experiments into a Lipid Monolayer
4.7.4. Permeability assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dayan, F.E.; Duke, S.O. Natural Compounds as Next-Generation Herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudai, N.; Poljakoff-Mayber, A.; Mayer, A.M.; Putievsky, E.; Lerner, H.R. Essential Oils as Allelochemicals and Their Potential Use as Bioherbicides. J. Chem. Ecol. 1999, 25, 1079–1089. [Google Scholar] [CrossRef]
- Tworkoski, T. Herbicide effects of essential oils. Weed Sci. 2002, 50, 425–431. [Google Scholar] [CrossRef]
- Amri, I.; Hamrouni, L.; Hanana, M.; Jamoussi, B. Reviews on phytotoxic effects of essential oils and their individual components: news approach for weeds management. Int. J. Appl. Biol. Pharm. Technol. 2013, 4, 96–114. [Google Scholar]
- Maffei, M.; Camusso, W.; Sacco, S. Effect of Mentha x piperita essential oil and monoterpenes on cucumber root membrane potential. Phytochemistry 2001, 58, 703–707. [Google Scholar] [CrossRef]
- Zunino, M.P.; Zygadlo, J.A. Effect of monoterpenes on lipid oxidation in maize. Planta 2004, 219, 303–309. [Google Scholar] [PubMed]
- Baser, K.H.C.; Buchbauer, G. Handbook of Essential Oils: Science, Technology, and Applications, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Harikumar, K.G.; Puri, V.; Singh, R.D.; Hanada, K.; Pagano, R.E.; Miller, L.J. Differential Effects of Modification of Membrane Cholesterol and Sphingolipids on the Conformation, Function, and Trafficking of the G Protein-coupled Cholecystokinin Receptor. J. Biol. Chem. 2005, 280, 2176–2185. [Google Scholar] [CrossRef] [Green Version]
- Maxfield, F.R.; Tabas, I. Role of cholesterol and lipid organization in disease. Nature 2005, 438, 612–621. [Google Scholar] [CrossRef]
- Khandelia, H.; Ipsen, J.H.; Mouritsen, O.G. The impact of peptides on lipid membranes. Biochim. Biophys. Acta Biomembr. 2008, 1778, 1528–1536. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.G. How lipids affect the activities of integral membrane proteins. Biochim. Biophys. Acta. 2004, 1666, 62–87. [Google Scholar] [CrossRef] [Green Version]
- Ruysschaert, J.-M.; Lonez, C. Role of lipid microdomains in TLR-mediated signalling ☆. BBA Biomembr. 2015, 1848, 1860–1867. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Altshuler, O.; Abu-Abied, M.; Chaimovitsh, D.; Shechter, A.; Frucht, H.; Dudai, N.; Sadot, E. Enantioselective Effects of (+)- and (−)-Citronellal on Animal and Plant Microtubules. J. Nat. Prod. 2013, 76, 1598–1604. [Google Scholar] [CrossRef] [PubMed]
- Chaimovitsh, D.; Shachter, A.; Abu-Abied, M.; Rubin, B.; Sadot, E.; Dudai, N. Herbicidal Activity of Monoterpenes Is Associated with Disruption of Microtubule Functionality and Membrane Integrity. Weed Sci. 2017, 65, 19–30. [Google Scholar] [CrossRef]
- Cavalieri, A.; Fischer, R.; Larkov, O.; Dudai, N. Enantioselectivity of the Bioconversion of Chiral Citronellal during the Inhibition of Wheat Seeds Germination. Chem. Biodivers. 2014, 11, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.C.; Lopes, P.M.; De Azevedo, M.M.; Costa, D.C.; Alviano, C.S.; Alviano, D.S. Biological Activities of a-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef] [PubMed]
- Tao, N.; Jia, L.; Zhou, H. Anti-fungal activity of Citrus reticulata Blanco essential oil against Penicillium italicum and Penicillium digitatum. Food Chem. 2014, 153, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Sharma, Y.; Rastogi, S.K.; Perwez, A.; Rizvi, M.A.; Manzoor, N. β-citronellol alters cell surface properties of Candida albicans to influence pathogenicity related traits. Med. Mycol. 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Pereira, F.; Mendes, J.M.; Lima, I.O.; Mota, K.S.; Oliveira, W.A.; Lima Ede, O. Antifungal activity of geraniol and citronellol, two monoterpenes alcohols, against Trichophyton rubrum involves inhibition of ergosterol biosynthesis. Pharm. Biol. 2005, 53, 228–234. [Google Scholar] [CrossRef]
- Lim, S.; Shin, S. Effects of citronellol and thymol on cell membrane composition of Candida albicans. Korean J. Pharmacogn. 2009, 40, 357–364. [Google Scholar]
- Lange, Y.; Ye, J.; Duban, M.-E.; Steck, T.L. Activation of Membrane Cholesterol by 63 Amphipaths. Biochemistry 2009, 48, 8505–8515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Rana, S.; Pal Singh, H.; Batish, D.; Kohli, R. Citronellol Disrupts Membrane Integrity by Inducing Free Radical Generation. Z. Naturforsch. C. 2011, 66, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; De Feo, V. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Zore, G.B.; Thakre, A.D.; Jadhav, S.; Karuppayil, S.M.; Candida, A. Terpenoids inhibit Candida albicans growth by affecting membrane integrity and arrest of cell cycle. Eur. J. Integr. Med. 2011, 18, 1181–1190. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Fatima, Z.; Hameed, S. Citronellal-induced disruption of membrane homeostasis in Candida albicans and attenuation of its virulence attributes. Rev. Soc. Bras. Med. Trop. 2016, 49, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudai, N.; Larkov, O.; Putievsky, E.; Lerner, H.R.; Ravid, U.; Lewinsohn, E.; Mayer, A.M. Biotransformation of constituents of essential oils by germinating wheat seed. Phytochemistry 2000, 55, 375–382. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Kohli, R.K.; Arora, K. Phytotoxicity of the volatile monoterpene citronellal against some weeds. Zeitschrift fur Naturforsch. Sect. C J. Biosci. 2006, 61, 334–340. [Google Scholar] [CrossRef]
- Ootani, M.A.; Dos Reis, M.R.; Cangussu, A.S.R.; Capone, A.; Fidelis, R.R.; Oliverira, W.; Barros, H.B.; Portella, A.C.F.; De Souza Aguiar, R.W.; Dos Santos, W.F. Phytotoxic effects of essential oils in controlling weed species Digitaria horizontalis and Cenchrus echinatus. Biocatal. Agric. Biotechnol. 2017, 12, 59–65. [Google Scholar] [CrossRef]
- Gill, A.O.; Holley, R.A. Disruption of Escherichia coli, Listeria monocytogenes and Lactobacillus sakei cellular membranes by plant oil aromatics. Int. J. Food Microbiol. 2006, 108, 1–9. [Google Scholar] [CrossRef]
- Shen, S.; Zhang, T.H.; Yuan, Y.; Lin, S.Y.; Xu, J.Y.; Ye, H.Y. Effects of cinnamaldehyde on Escherichia coli and Staphylococcus aureus membrane. Food Control 2015, 47, 196–202. [Google Scholar] [CrossRef]
- He, T.-F.; Zhang, Z.-H.; Zeng, X.-A.; Wang, L.-H.; Brennan, C.S. Determination of membrane disruption and genomic DNA binding of cinnamaldehyde to Escherichia coli by use of microbiological and spectroscopic techniques. J. Photochem. Photobiol. B Biol. 2018, 178, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Di Pasqua, R.; Hoskins, N.; Betts, G.; Mauriello, G. Changes in membrane fatty acids composition of microbial cells induced by addiction of thymol, carvacrol, limonene, cinnamaldehyde, and eugenol in the growing media. J. Agric. Food Chem. 2006, 54, 2745–2749. [Google Scholar] [CrossRef] [PubMed]
- Nowotarska, S.; Nowotarski, K.; Friedman, M.; Situ, C. Effect of Structure on the Interactions between Five Natural Antimicrobial Compounds and Phospholipids of Bacterial Cell Membrane on Model Monolayers. Molecules 2014, 19, 7497–7515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shreaz, S.; Sheikh, R.A.; Rimple, B.; Hashmi, A.A.; Nikhat, M.; Khan, L.A. Anticandidal activity of cinnamaldehyde, its ligand and Ni(II) complex: Effect of increase in ring and side chain. Microb. Pathog. 2010, 49, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Kong, J.; Ju, J.; Zhang, Y.; Guo, Y.; Cheng, Y.; Qian, H.; Xie, Y.; Yao, W. Membrane damage mechanism contributes to inhibition of trans-cinnamaldehyde on Penicillium italicum using Surface-Enhanced Raman Spectroscopy (SERS). Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chotsaeng, N.; Laosinwattana, C.; Charoenying, P. Inhibitory Effects of a Variety of Aldehydes on Amaranthus tricolor L. and Echinochloa crus-galli (L.) Beauv. Molecules 2018, 23, 471. [Google Scholar] [CrossRef] [PubMed]
- Genva, M.; Lins, L.; Fauconnier, M.L. Penetration of the cinnamon and citronella essential oils components into A. thaliana leaves. 2019; unpublished work. [Google Scholar]
- Furt, F.; Simon-Plas, F.; Mongrand, S. Lipids of the Plant Plasma Membrane; Springer International Publishing: Berlin/Heidelberg, Germany, 2011; pp. 3–30. [Google Scholar] [CrossRef]
- Ducarme, P.; Rahman, M.; Brasseur, R. IMPALA: a simple restraint field to simulate the biological membrane in molecular structure studies. Proteins 1998, 30, 357–371. [Google Scholar] [CrossRef]
- Marsh, D. Lateral pressure in membranes. Biochim. Biophys. Acta Biomembr. 1996, 1286, 183–223. [Google Scholar] [CrossRef]
- Cristani, M.; D’Arrigo, M.; Mandalari, G.; Castelli, F.; Sarpietro, M.G.; Micieli, D.; Venuti, V.; Bisignano, G.; Saija, A.; Trombetta, D. Interaction of four monoterpenes contained in essential oils with model membranes: implications for their antibacterial activity. J. Agric. Food Chem. 2007, 55, 6300–6308. [Google Scholar] [CrossRef]
- Henry, G.; Deleu, M.; Jourdan, E.; Thonart, P.; Ongena, M. The bacterial lipopeptide surfactin targets the lipid fraction of the plant plasma membrane to trigger immune-related defence responses. Cell. Microbiol. 2011, 13, 1824–1837. [Google Scholar] [CrossRef] [PubMed]
- Tsagareli, M.G.; Tsiklauri, N.; Zanotto, K.L.; Carstens, M.I.; Klein, A.H.; Sawyer, C.M.; Gurtskaia, G.; Abzianidze, E.; Carstens, E. Behavioral evidence of thermal hyperalgesia and mechanical allodynia induced by intradermal cinnamaldehyde in rats. Neurosci. Lett. 2010, 473, 233–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, G.; Im, S.T.; Kim, Y.H.; Jung, S.J.; Rhyu, M.R.; Oh, S.B. Activation of transient receptor potential ankyrin 1 by eugenol. Neuroscience 2014, 261, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Namer, B.; Seifert, F.; Handwerker, H.O.; Maihofner, C. TRPA1 and TRPM8 activation in humans: effects of cinnamaldehyde and menthol. Neuroreport 2005, 16, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Tanoh, E.A.; Nea, F.; Kemene, T.K.; Genva, M.; Saive, M.; Tonzibo, F.Z.; Fauconnier, M.L. Antioxidant and Lipoxygenase Inhibitory Activities of Essential Oils from Endemic Plants of Côte d’Ivoire: Zanthoxylum mezoneurispinosum Ake Assi and Zanthoxylum psammophilum Ake Assi. Molecules 2019, 24, 2445. [Google Scholar] [CrossRef] [PubMed]
- Benini, C.; Ringuet, M.; Wathelet, J.P.; Lognay, G.; Du Jardin, P.; Fauconnier, M.L. Variations in the essential oils from ylang-ylang (Cananga odorata [Lam.] Hook f. & Thomson forma genuina) in the Western Indian Ocean islands. Flavour Fragr. J. 2012, 27, 356–366. [Google Scholar]
- Harhour, A.; Brada, M.; Fauconnier, M.-L.; Lognay, G. Chemical Composition and Antioxidant Activity of Algerian Juniperus Phoenicea Essential Oil. Nat. Prod. Sci. 2018, 24, 125. [Google Scholar] [CrossRef]
- Lichiheb, N.; Bedos, C.; Personne, E.; Benoit, P.; Bergheaud, V.; Fanucci, O.; Bouhlel, J.; Barriuso, E. Measuring Leaf Penetration and Volatilization of Chlorothalonil and Epoxiconazole Applied on Wheat Leaves in a Laboratory-Scale Experiment. J. Environ. Qual. 2015, 44, 1782–1790. [Google Scholar] [CrossRef] [Green Version]
- Lins, L.; Brasseur, R.; Malaisse, W.J.; Biesemans, M.; Verheyden, P.; Willem, R. Importance of the hydrophobic energy: Structural determination of a hypoglycemic drug of the meglitinide family by nuclear magnetic resonance and molecular modeling. Biochem. Pharmacol. 1996, 52, 1155–1168. [Google Scholar] [CrossRef]
- Hess, B.; Bekker, H.; Berendsen, M.H.J.C.; Fraaije, J.G.E. LINCS: A Linear Constraint Solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Malde, A.K.; Zuo, L.; Breeze, M.; Stroet, M.; Poger, D.; Nair, P.C.; Oostenbrink, C.; Mark, A.E. An Automated force field Topology Builder (ATB) and repository: Version 1.0. J. Chem. Theory Comput. 2011, 7, 4026–4037. [Google Scholar] [CrossRef] [PubMed]
- Parrinello, M.; Rahman, A. Polymorphic transitions in single crystals: A new molecular dynamics method. J. Appl. Phys. 1981, 7182. [Google Scholar] [CrossRef]
- Berger, O.; Edholm, O.; Jähnig, F. Molecular dynamics simulations of a fluid bilayer of dipalmitoylphosphatidylcholine at full hydration, constant pressure, and constant temperature. Biophys. J. 1997, 72, 2002–2013. [Google Scholar] [CrossRef] [Green Version]
- Bachar, M.; Brunelle, P.; Tieleman, D.P.; Rauk, A. Molecular dynamics simulation of a polyunsaturated lipid bilayer susceptible to lipid peroxidation. J. Phys. Chem. B 2004, 108, 7170–7179. [Google Scholar] [CrossRef]
- Knight, C.J.; Hub, J.S. MemGen: A general web server for the setup of lipid membrane simulation systems. Bioinformatics 2015, 31, 2897–2899. [Google Scholar] [CrossRef] [PubMed]
- Hermans, J.; Berendsen, M.H.J.C.; Van Gunsteren, W.F.; Postma, J.P.M. A consistent empirical potential for water–protein interactions. Biopolymers 1984, 23, 1513–1518. [Google Scholar] [CrossRef]
- Hoover, W.G. Canonical dynamics: Equilibrium phase-space distributions. Phys. Rev. A 1985, 31, 1685. [Google Scholar] [CrossRef] [PubMed]
- Nosé, S. A unified formulation of the constant temperature molecular dynamics methods. J. Chem. Phys. 1984, 81, 511. [Google Scholar] [CrossRef]
- Essmann, U.; Perera, L.; Berkowitz, M.L. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Razafindralambo, H.; Dufour, S.; Paquot, M.; Deleu, M. Thermodynamic studies of the binding interactions of surfactin analogues to lipid vesicles. J. Therm. Anal. Calorim. 2009, 95, 817–821. [Google Scholar] [CrossRef]
- Calvez, P.; Bussieres, S.; Eric, D.; Salesse, C. Parameters modulating the maximum insertion pressure of proteins and peptides in lipid monolayers. Biochimie 2009, 91, 718–733. [Google Scholar] [CrossRef] [PubMed]
- Van Bambeke, F.; Kerkhofs, A.; Schanck, A.; Remacle, C.; Sonveaux, E.; Tulkens, P.M.; Mingeot-Leclercq, M.P. Biophysical studies and intracellular destabilization of pH-sensitive liposomes. Lipids 2000, 35, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Cacas, J.-L.; Buré, C.; Grosjean, K.; Gerbeau-Pissot, P.; Lherminier, J.; Rombouts, Y.; Maes, E.; Bossard, C.; Gronnier, J.; Furt, F.; et al. Revisiting plant plasma membrane lipids in tobacco: A focus on sphingolipids. Plant Physiol. 2016, 170, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Bouquillon, S.; Fauconnier, M.L. Encapsulation of Essential Oils for the Development of Biosourced Pesticides with Controlled Release: A Review. Molecules 2019, 24, 2539. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| K (mM−1) | ∆HDw →b (kJ·mol−1) | T∆SDw →b (kJ·mol−1) | ∆GDw →b (kJ·mol−1) | |
|---|---|---|---|---|
| CitA | 0.03 ± 0.00 | 5.1 ± 0.6 | 21.6 ± 0.3 | −16.5 ± 0.9 |
| CitO | 0.01 ± 0.01 | 5.1 ± 0.5 | 23.3 ± 0.5 | −18.5 ± 0.5 |
| CIN | Not quantifiable | Not quantifiable | Not quantifiable | Not quantifiable |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lins, L.; Dal Maso, S.; Foncoux, B.; Kamili, A.; Laurin, Y.; Genva, M.; Jijakli, M.H.; De Clerck, C.; Fauconnier, M.L.; Deleu, M. Insights into the Relationships Between Herbicide Activities, Molecular Structure and Membrane Interaction of Cinnamon and Citronella Essential Oils Components. Int. J. Mol. Sci. 2019, 20, 4007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164007
Lins L, Dal Maso S, Foncoux B, Kamili A, Laurin Y, Genva M, Jijakli MH, De Clerck C, Fauconnier ML, Deleu M. Insights into the Relationships Between Herbicide Activities, Molecular Structure and Membrane Interaction of Cinnamon and Citronella Essential Oils Components. International Journal of Molecular Sciences. 2019; 20(16):4007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164007
Chicago/Turabian StyleLins, Laurence, Simon Dal Maso, Berenice Foncoux, Anouar Kamili, Yoann Laurin, Manon Genva, M. Haissam Jijakli, Caroline De Clerck, Marie Laure Fauconnier, and Magali Deleu. 2019. "Insights into the Relationships Between Herbicide Activities, Molecular Structure and Membrane Interaction of Cinnamon and Citronella Essential Oils Components" International Journal of Molecular Sciences 20, no. 16: 4007. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164007