H19-Dependent Transcriptional Regulation of β3 and β4 Integrins Upon Estrogen and Hypoxia Favors Metastatic Potential in Prostate Cancer

 ,
,  ,
,
Abstract
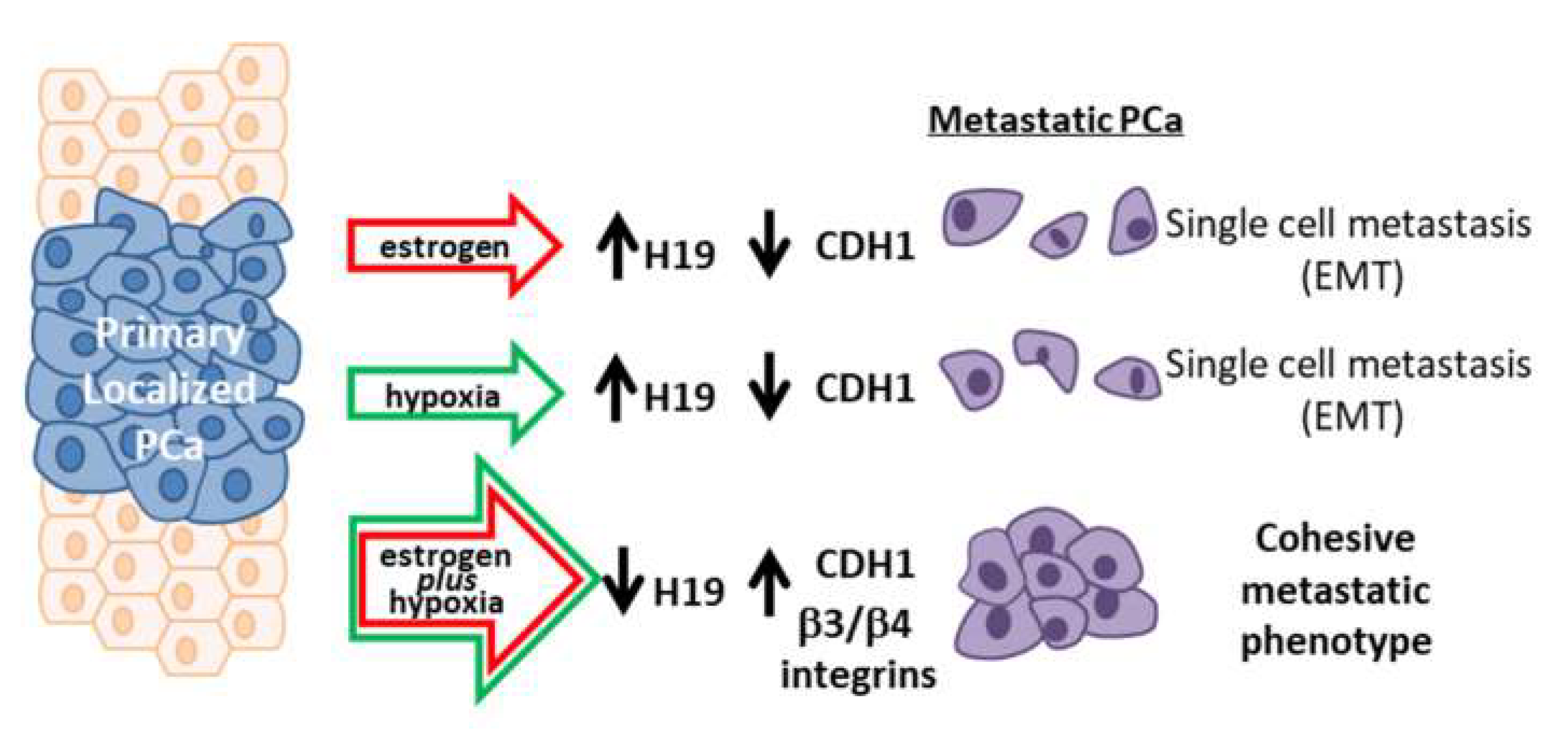
:1. Introduction
2. Results
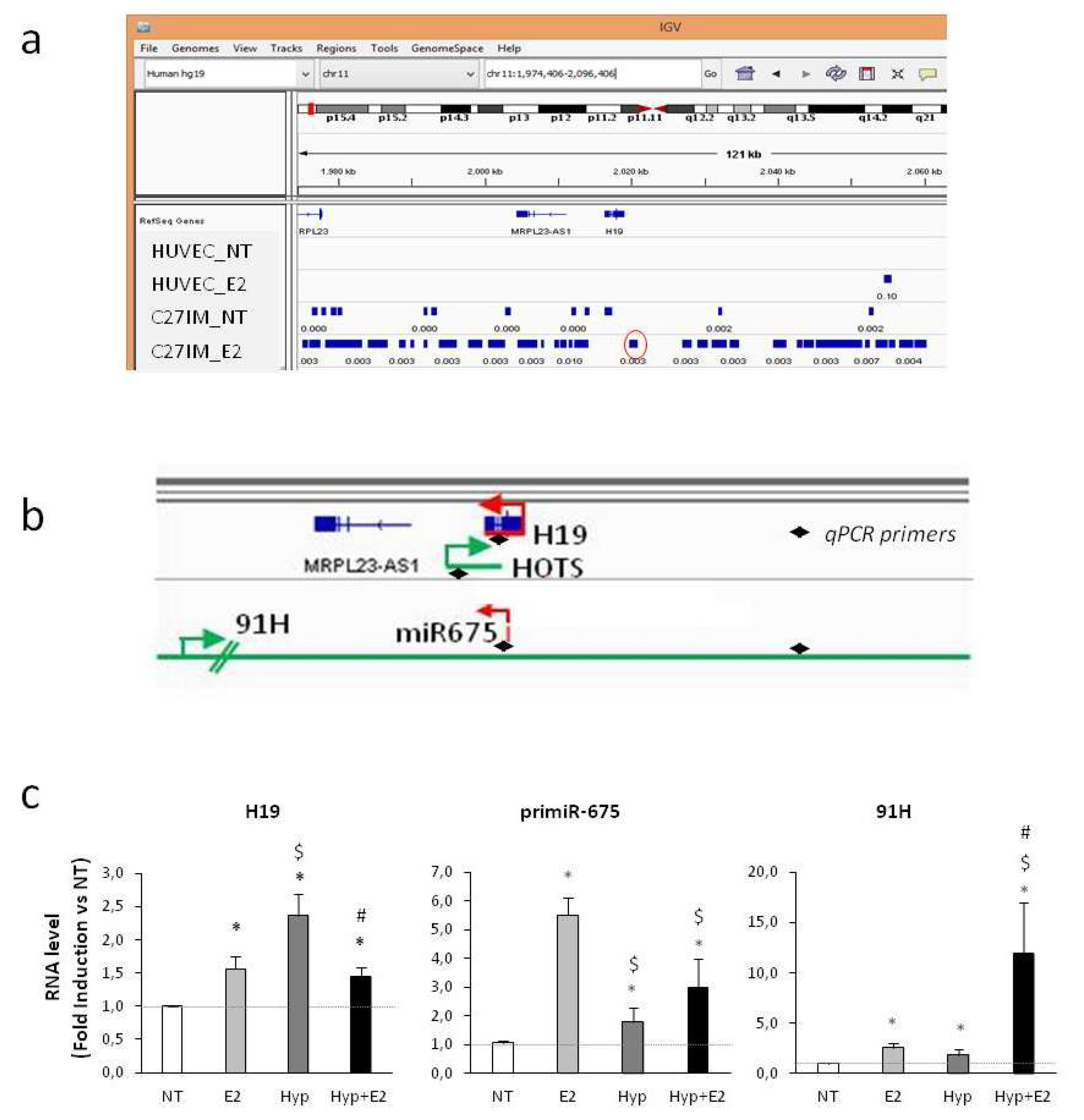
2.1. Response of H19 Transcripts to Combined Estrogen and Hypoxia
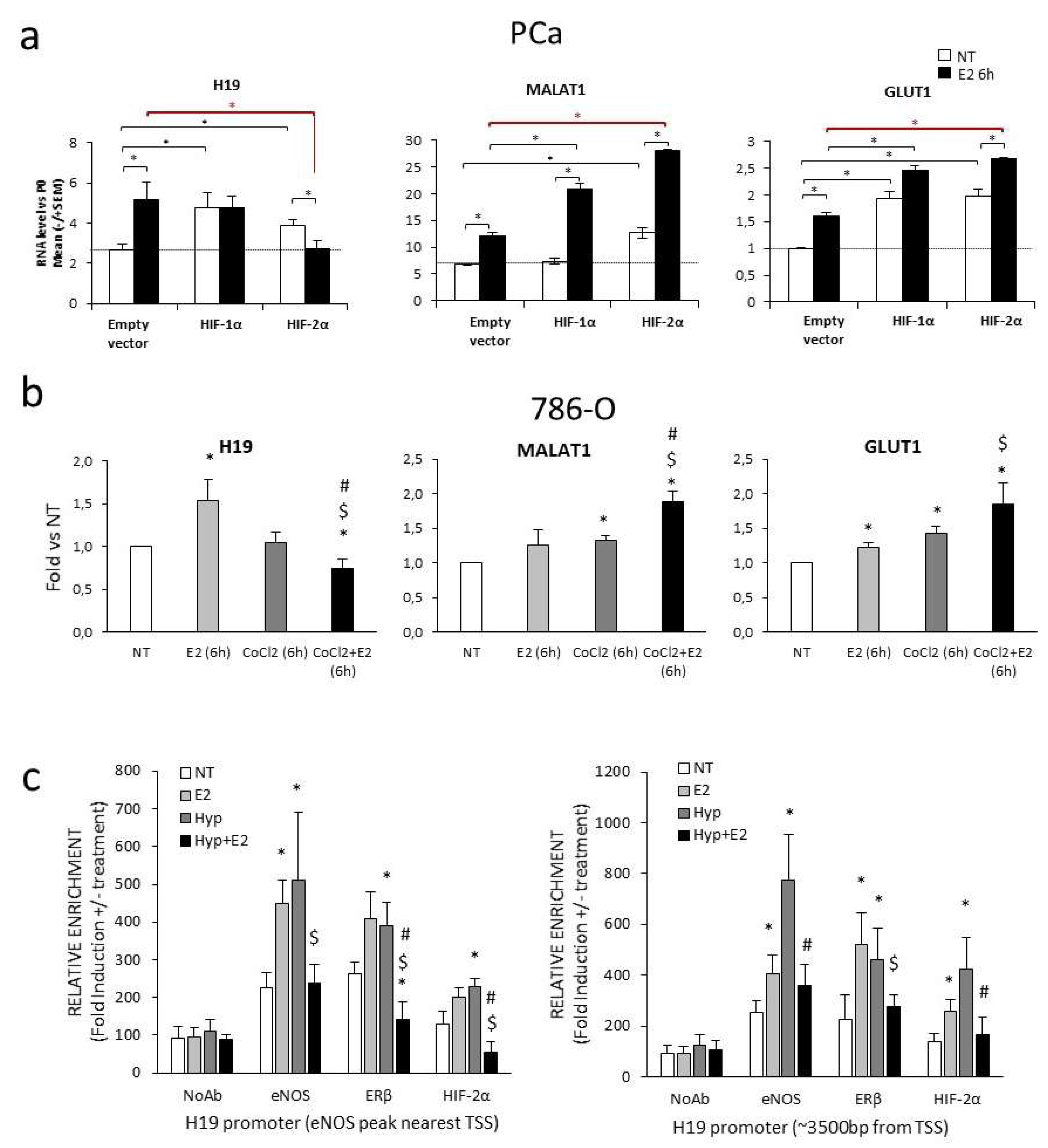
2.2. Transcriptional Regulation of H19 upon Combined Treatment
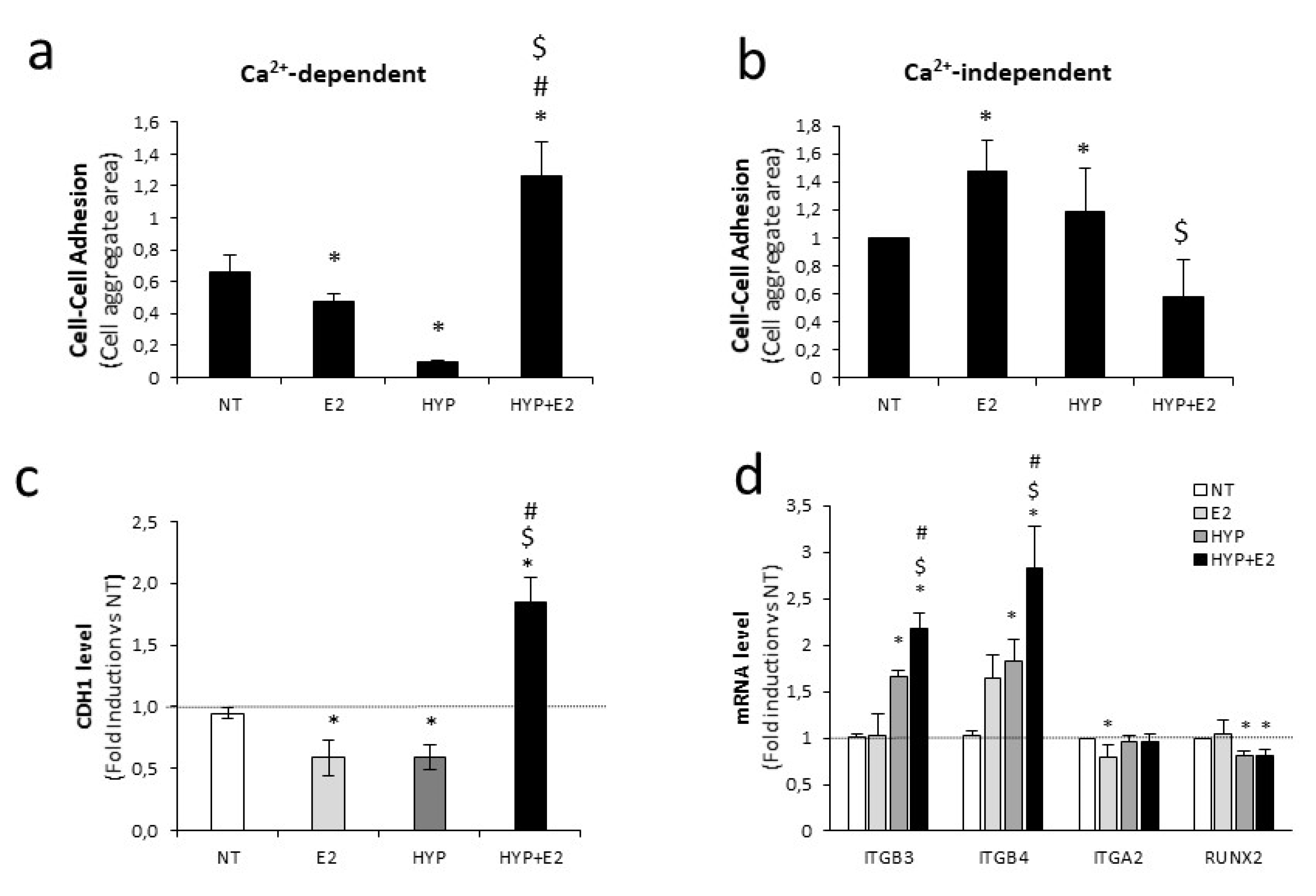
2.3. Cell Adhesion and Invasion Modulation Upon Combined Treatment
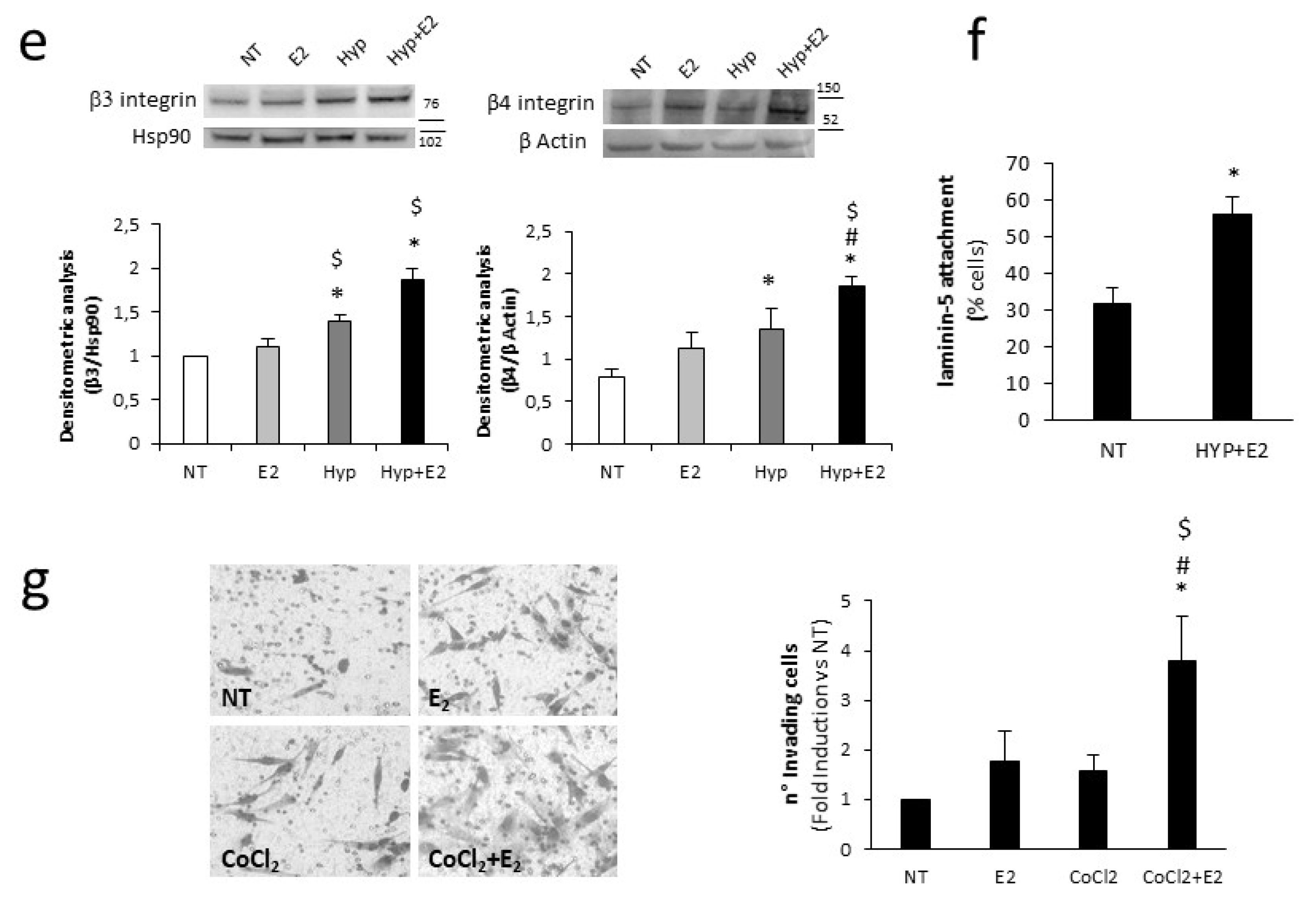
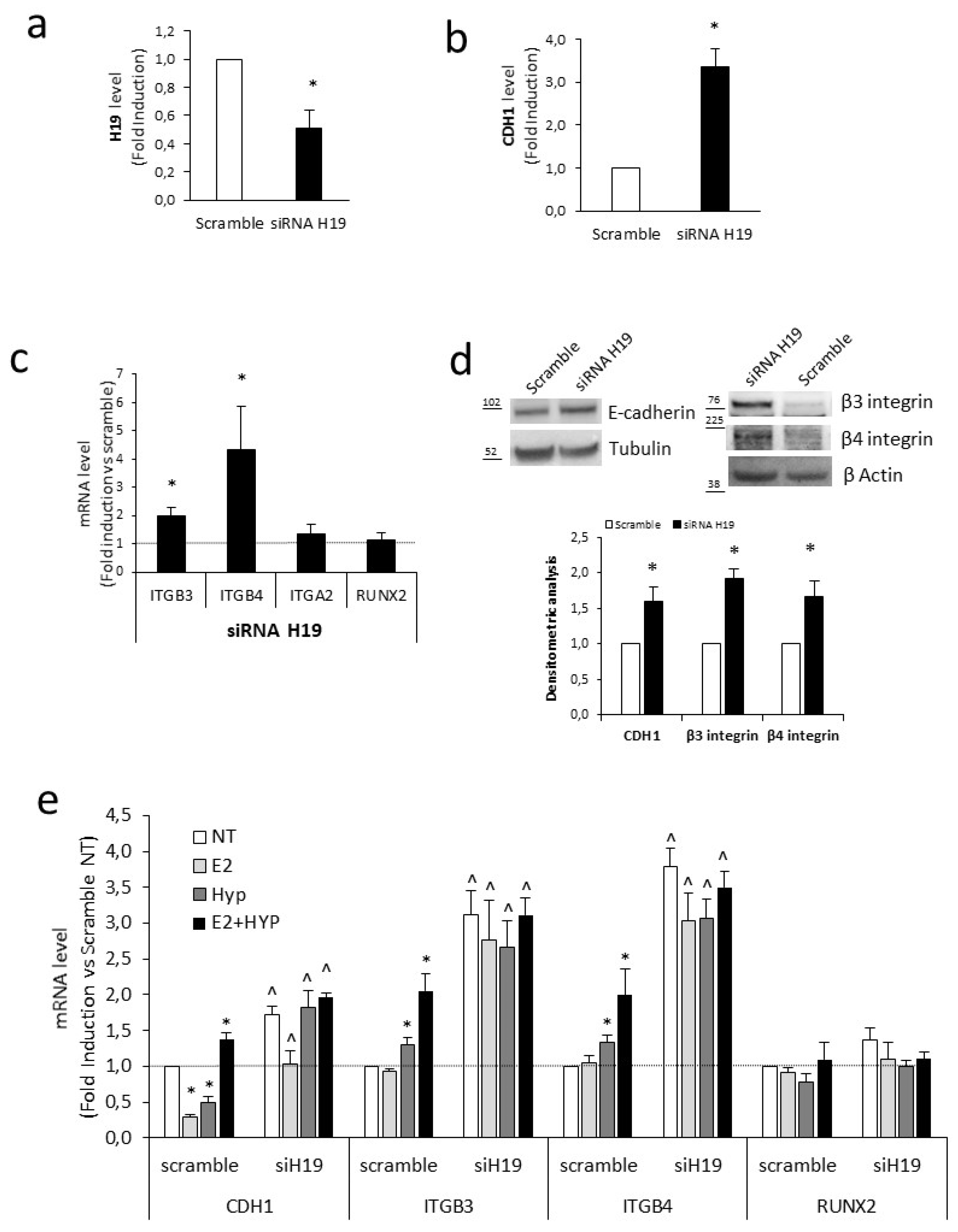
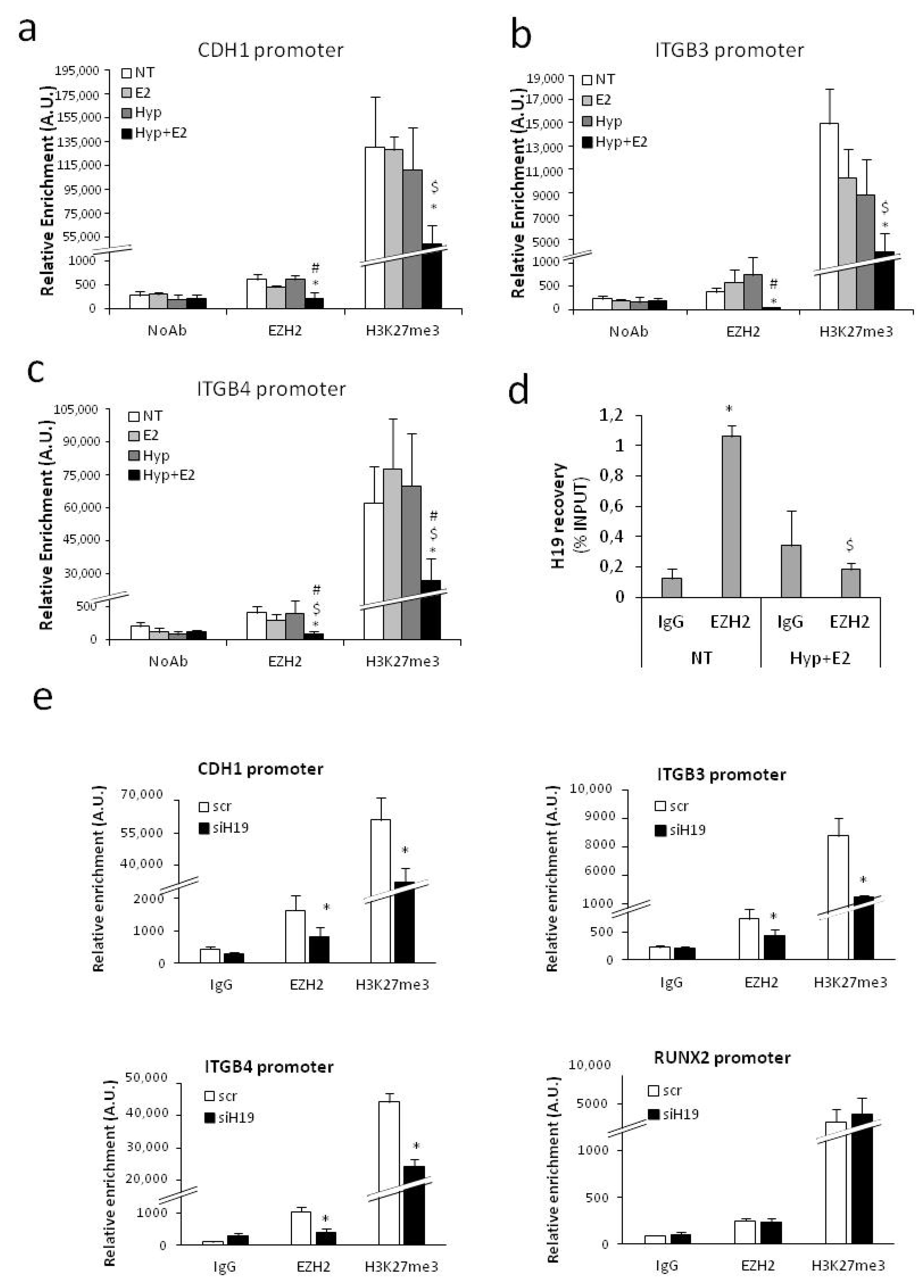
2.4. H19 Mediates Transcriptional Repression of Cell Adhesion Molecules
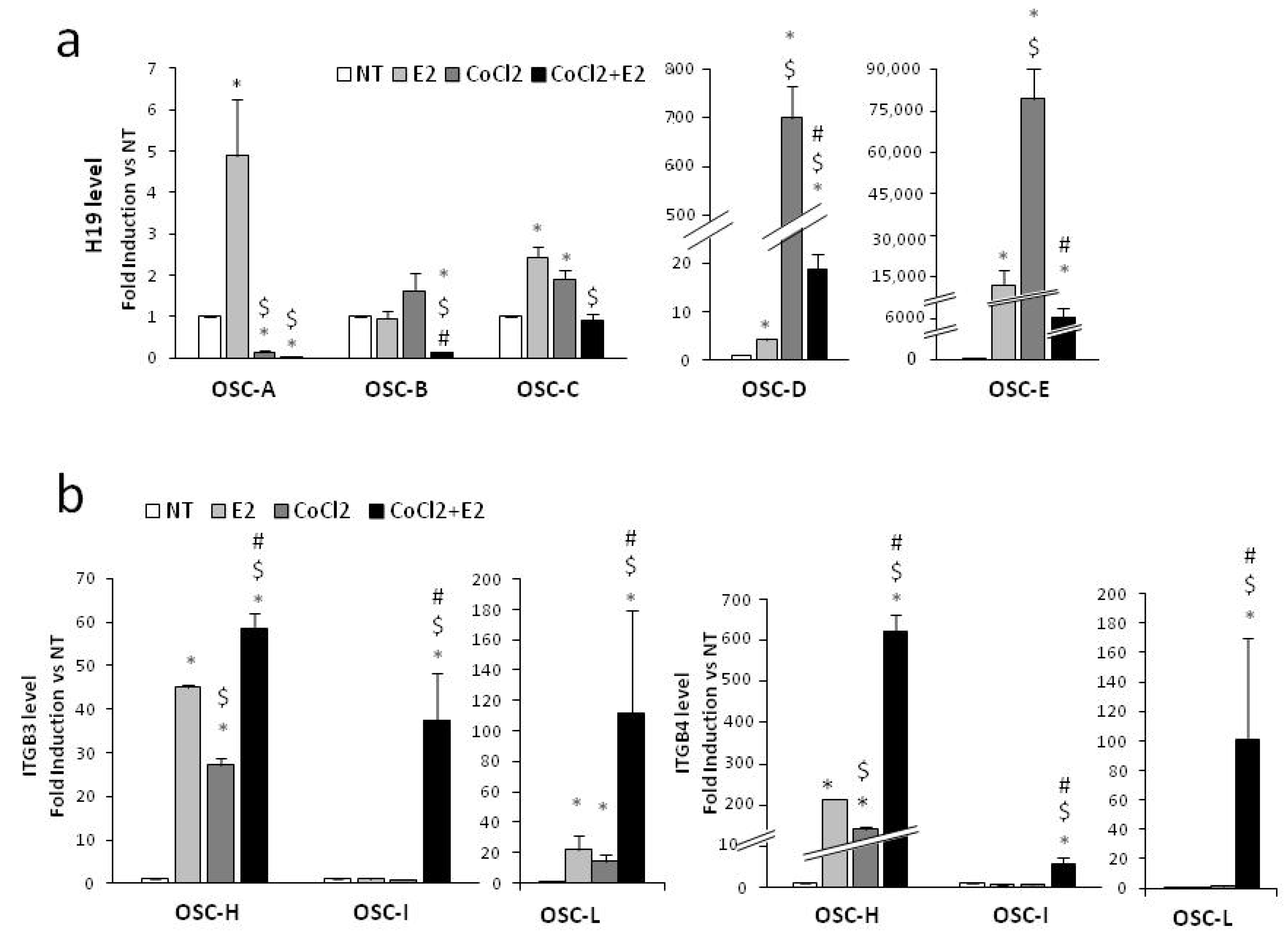
2.5. H19 and Integrin Modulation on Ex Vivo Organotypic Slice Cultures (OSCs) of Prostate Tumor upon Combined Treatment
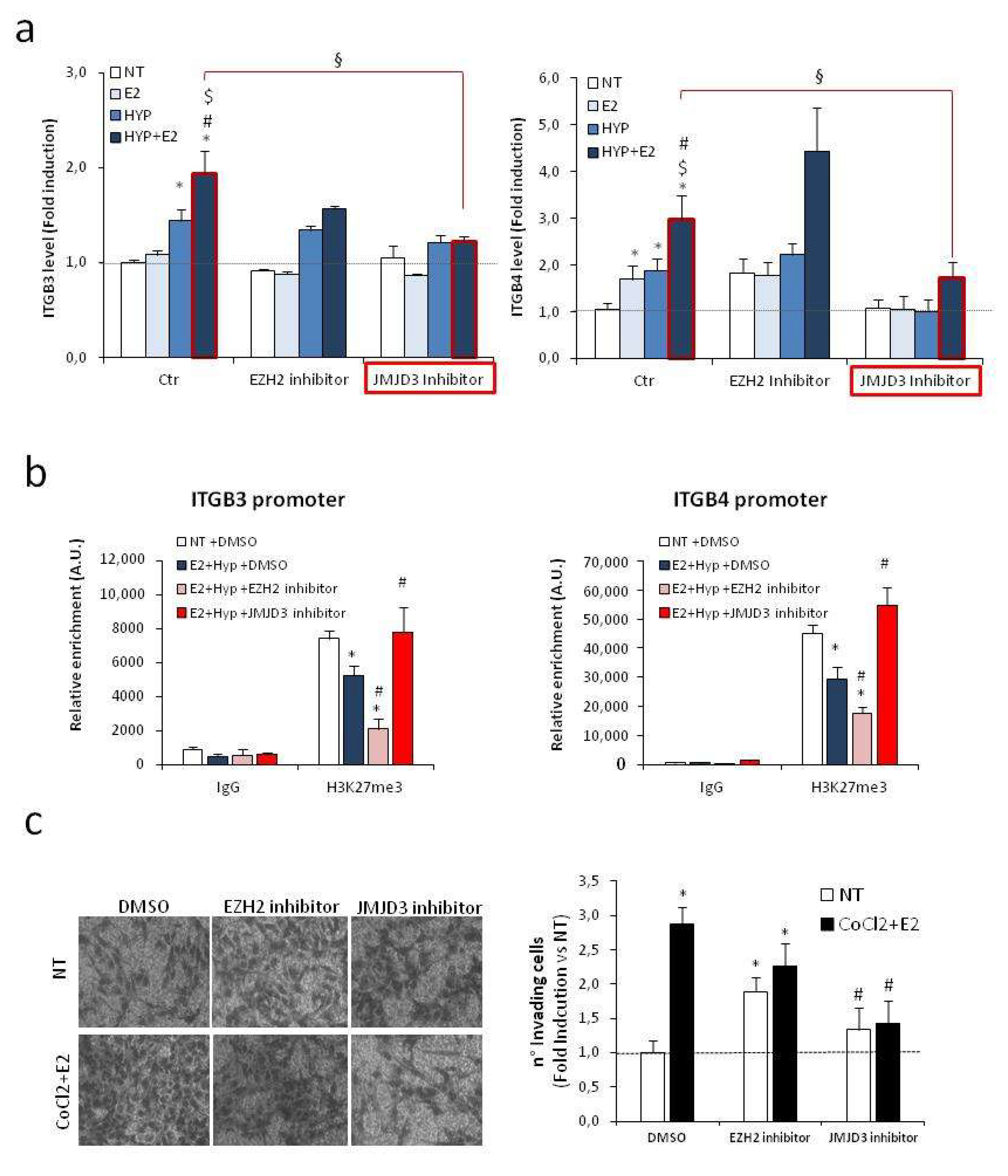
2.6. Effects of Epigenetic Drugs on H19/Integrin Pathway upon Combined Treatment
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cells and Treatments
4.3. RNA Extraction and Real Time PCR
4.4. Transient Transfection Assays
4.5. Chromatin Immunoprecipitation (ChIP)
4.6. RNA Interference
4.7. RNA-Chromatin Immunoprecipitation (RNA-ChIP)
4.8. Protein Extraction and Western Blot
4.9. Cell–Cell Adhesion Assay
4.10. Scratch Test
4.11. Laminin 5 Cell Motility Test
4.12. Boyden’s Chamber Assay
4.13. Organotypic Slice Cultures (OSCs) from Prostate Cancer Specimens
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PCa | prostate cancer |
| PSA | prostate-specific antigen |
| RP | radical prostatectomy |
| RT | radiation therapy |
| ECM | extracellular matrix |
| CDH1 | E-cadherin |
| EMT | epithelial to mesenchymal transition |
| lncRNAs | long non-coding RNAs |
| ER | estrogen receptor |
| eNOS | endothelial nitric oxide synthase |
| HIFs | hypoxia inducible factors |
| HOTS | H19 Opposite Tumor Suppressor |
| E2 | 17β-estradiol |
| HYP | hypoxia |
| CoCl2 | cobalt chloride |
| qPCR | quantitative polymerase chain reaction |
| KDR | vascular endothelial growth factor receptor 2 |
| EPO | erythropoietin |
| ChIP | chromatin immunoprecipitation |
| TSS | transcriptional start site |
| HDAC | histone deacetylases |
| VHL | von Hippel-Lindau tumor suppressor |
| Ca2+ | calcium |
| ITGB3 | β3 integrin subunit |
| ITGB4 | β4 integrin subunit |
| ITGA2 | α2 integrin subunit |
| RUNX2 | Runt-related transcription factor 2 |
| H3K27me3 | trimethylated lysine 27 of histone H3 |
| OSCs | organotypic slice cultures |
| CCND1 | Cyclin D1 |
| RNA-ChIP | RNA chromatin immunoprecipitation |
| DMSO | dimethyl sulfoxide |
| EDTA | ethylenediaminetetraacetic acid |
| PBS | phosphate-buffered saline |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: Globocan Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2015. CA Cancer J. Clin. 2015, 65, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Freedland, S.J.; Humphreys, E.B.; Mangold, L.A.; Eisenberger, M.; Dorey, F.J.; Walsh, P.C.; Partin, A.W. Risk of Prostate Cancer-Specific Mortality Following Biochemical Recurrence After Radical Prostatectomy. JAMA 2005, 294, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Kupelian, P.A.; Mahadevan, A.; Reddy, C.A.; Reuther, A.M.; Klein, E.A. Use of Different Definitions of Biochemical Failure After External Beam Radiotherapy Changes Conclusions about Relative Treatment Efficacy for Localized Prostate Cancer. Urology 2006, 68, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Boorjian, S.A.; Thompson, R.H.; Tollefson, M.K.; Rangel, L.J.; Bergstralh, E.J.; Blute, M.L.; Karnes, R.J. Long-Term Risk of Clinical Progression After Biochemical Recurrence Following Radical Prostatectomy: The Impact of Time from Surgery to Recurrence. Eur. Urol. 2011, 59, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-San Segundo, C.; Jove, J.; Zapatero, A.; Pastor-Peidro, J.; Vazquez, M.L.; Casana, M.; Mengual, J.L.; Gomez-Caamano, A.; Gomez-Iturriaga, A.; Vallejo, C.; et al. Survival After Biochemical Failure in Prostate Cancer Treated with Radiotherapy: Spanish Registry of Prostate Cancer (RECAP) Database Outcomes. Clin. Transl. Oncol. 2019, 21, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.R.; Saad, F.; Chowdhury, S.; Oudard, S.; Hadaschik, B.A.; Graff, J.N.; Olmos, D.; Mainwaring, P.N.; Lee, J.Y.; Uemura, H.; et al. Apalutamide Treatment and Metastasis-Free Survival in Prostate Cancer. N. Engl. J. Med. 2018, 378, 1408–1418. [Google Scholar] [CrossRef] [PubMed]
- Krieg, M.; Nass, R.; Tunn, S. Effect of Aging on Endogenous Level of 5 Alpha-Dihydrotestosterone, Testosterone, Estradiol, and Estrone in Epithelium and Stroma of Normal and Hyperplastic Human Prostate. J. Clin. Endocrinol. Metab. 1993, 77, 375–381. [Google Scholar]
- Di Zazzo, E.; Galasso, G.; Giovannelli, P.; di Donato, M.; Castoria, G. Estrogens and their Receptors in Prostate Cancer: Therapeutic Implications. Front. Oncol. 2018, 8, 2. [Google Scholar] [CrossRef]
- Bonkhoff, H. Estrogen Receptor Signaling in Prostate Cancer: Implications for Carcinogenesis and Tumor Progression. Prostate 2018, 78, 2–10. [Google Scholar] [CrossRef]
- Di Zazzo, E.; Galasso, G.; Giovannelli, P.; Di Donato, M.; Di Santi, A.; Cernera, G.; Rossi, V.; Abbondanza, C.; Moncharmont, B.; Sinisi, A.A.; et al. Prostate Cancer Stem Cells: The Role of Androgen and Estrogen Receptors. Oncotarget 2016, 7, 193–208. [Google Scholar] [CrossRef]
- Fixemer, T.; Remberger, K.; Bonkhoff, H. Differential Expression of the Estrogen Receptor Beta (ERbeta) in Human Prostate Tissue, Premalignant Changes, and in Primary, Metastatic, and Recurrent Prostatic Adenocarcinoma. Prostate 2003, 54, 79–87. [Google Scholar] [CrossRef]
- Muz, B.; de la Puente, P.; Azab, F.; Azab, A.K. The Role of Hypoxia in Cancer Progression, Angiogenesis, Metastasis, and Resistance to Therapy. Hypoxia 2015, 3, 83–92. [Google Scholar] [CrossRef]
- Stewart, G.D.; Ross, J.A.; McLaren, D.B.; Parker, C.C.; Habib, F.K.; Riddick, A.C. The Relevance of a Hypoxic Tumour Microenvironment in Prostate Cancer. BJU Int. 2010, 105, 8–13. [Google Scholar] [CrossRef]
- Hou, R.; Xu, Y.; Lu, Q.; Zhang, Y.; Hu, B. Effect of Low-Frequency Low-Intensity Ultrasound with Microbubbles on Prostate Cancer Hypoxia. Tumor Biol. 2017, 39, 1010428317719275. [Google Scholar] [CrossRef]
- Rankin, E.B.; Giaccia, A.J. The Role of Hypoxia-Inducible Factors in Tumorigenesis. Cell Death Differ. 2008, 15, 678–685. [Google Scholar] [CrossRef]
- Raval, R.R.; Lau, K.W.; Tran, M.G.; Sowter, H.M.; Mandriota, S.J.; Li, J.L.; Pugh, C.W.; Maxwell, P.H.; Harris, A.L.; Ratcliffe, P.J. Contrasting Properties of Hypoxia-Inducible Factor 1 (HIF-1) and HIF-2 in Von Hippel-Lindau-Associated Renal Cell Carcinoma. Mol. Cell. Biol. 2005, 25, 5675–5686. [Google Scholar] [CrossRef]
- Kim, W.Y.; Perera, S.; Zhou, B.; Carretero, J.; Yeh, J.J.; Heathcote, S.A.; Jackson, A.L.; Nikolinakos, P.; Ospina, B.; Naumov, G.; et al. HIF2alpha Cooperates with RAS to Promote Lung Tumorigenesis in Mice. J. Clin. Investig. 2009, 119, 2160–2170. [Google Scholar] [CrossRef]
- Najafi, M.; Farhood, B.; Mortezaee, K. Extracellular Matrix (ECM) Stiffness and Degradation as Cancer Drivers. J. Cell. Biochem. 2019, 120, 2782–2790. [Google Scholar] [CrossRef]
- Desgrosellier, J.S.; Cheresh, D.A. Integrins in Cancer: Biological Implications and Therapeutic Opportunities. Nat. Rev. Cancer 2010, 10, 9–22. [Google Scholar] [CrossRef]
- Slack-Davis, J.K.; Parsons, J.T. Emerging Views of Integrin Signaling: Implications for Prostate Cancer. J. Cell. Biochem. 2004, 91, 41–46. [Google Scholar] [CrossRef]
- Davies, G.; Jiang, W.G.; Mason, M.D. E-Cadherin and Associated Molecules in the Invasion and Progression of Prostate Cancer. Oncol. Rep. 1998, 5, 1567–1576. [Google Scholar] [CrossRef]
- Juan-Rivera, M.C.; Martinez-Ferrer, M. Integrin Inhibitors in Prostate Cancer. Cancers 2018, 10, 44. [Google Scholar] [CrossRef]
- Ramovs, V.; Te Molder, L.; Sonnenberg, A. The Opposing Roles of Laminin-Binding Integrins in Cancer. Matrix Biol. 2017, 57, 213–243. [Google Scholar] [CrossRef]
- Cooper, C.R.; Chay, C.H.; Pienta, K.J. The Role of Alpha(V)Beta(3) in Prostate Cancer Progression. Neoplasia 2002, 4, 191–194. [Google Scholar] [CrossRef]
- Van Roy, F. Beyond E-Cadherin: Roles of Other Cadherin Superfamily Members in Cancer. Nat. Rev. Cancer 2014, 14, 121–134. [Google Scholar] [CrossRef]
- Van Roy, F.; Berx, G. The Cell-Cell Adhesion Molecule E-Cadherin. Cell Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef]
- Prensner, J.R.; Chinnaiyan, A.M. The Emergence of lncRNAs in Cancer Biology. Cancer Discov. 2011, 1, 391–407. [Google Scholar] [CrossRef]
- Misawa, A.; Takayama, K.I.; Inoue, S. Long Non-Coding RNAs and Prostate Cancer. Cancer Sci. 2017, 108, 2107–2114. [Google Scholar] [CrossRef]
- Raveh, E.; Matouk, I.J.; Gilon, M.; Hochberg, A. The H19 Long Non-Coding RNA in Cancer Initiation, Progression and Metastasis a Proposed Unifying Theory. Mol. Cancer 2015, 14, 646. [Google Scholar] [CrossRef]
- Wu, W.; Hu, Q.; Nie, E.; Yu, T.; Wu, Y.; Zhi, T.; Jiang, K.; Shen, F.; Wang, Y.; Zhang, J.; et al. Hypoxia Induces H19 Expression through Direct and Indirect Hif-1alpha Activity, Promoting Oncogenic Effects in Glioblastoma. Sci. Rep. 2017, 7, 45029. [Google Scholar] [CrossRef]
- Lottin, S.; Adriaenssens, E.; Dupressoir, T.; Berteaux, N.; Montpellier, C.; Coll, J.; Dugimont, T.; Curgy, J.J. Overexpression of an Ectopic H19 Gene Enhances the Tumorigenic Properties of Breast Cancer Cells. Carcinogenesis 2002, 23, 1885–1895. [Google Scholar] [CrossRef]
- Manoharan, H.; Babcock, K.; Willi, J.; Pitot, H.C. Biallelic Expression of the H19 Gene during Spontaneous Hepatocarcinogenesis in the Albumin SV40 T Antigen Transgenic Rat. Mol. Carcinog. 2003, 38, 40–47. [Google Scholar] [CrossRef]
- Yoshimizu, T.; Miroglio, A.; Ripoche, M.A.; Gabory, A.; Vernucci, M.; Riccio, A.; Colnot, S.; Godard, C.; Terris, B.; Jammes, H.; et al. The H19 Locus Acts in Vivo as a Tumor Suppressor. Proc. Natl. Acad. Sci. USA 2008, 105, 12417–12422. [Google Scholar] [CrossRef]
- Hao, Y.; Crenshaw, T.; Moulton, T.; Newcomb, E.; Tycko, B. Tumour-Suppressor Activity of H19 RNA. Nature 1993, 365, 764–767. [Google Scholar] [CrossRef]
- Matouk, I.J.; DeGroot, N.; Mezan, S.; Ayesh, S.; Abu-lail, R.; Hochberg, A.; Galun, E. The H19 Non-Coding RNA is Essential for Human Tumor Growth. PLoS ONE 2007, 2, e845. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Lottin, S.; Dugimont, T.; Fauquette, W.; Coll, J.; Dupouy, J.P.; Boilly, B.; Curgy, J.J. Steroid Hormones Modulate H19 Gene Expression in both Mammary Gland and Uterus. Oncogene 1999, 18, 4460–4473. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, Q.; Liu, X.; Sun, Q.; Zhao, X.; Deng, R.; Wang, Y.; Huang, J.; Xu, M.; Yan, J.; et al. lncRNA H19/miR-675 Axis Represses Prostate Cancer Metastasis by Targeting TGFBI. FEBS J. 2014, 281, 3766–3775. [Google Scholar] [CrossRef]
- Nanni, S.; Benvenuti, V.; Grasselli, A.; Priolo, C.; Aiello, A.; Mattiussi, S.; Colussi, C.; Lirangi, V.; Illi, B.; D’Eletto, M.; et al. Endothelial NOS, Estrogen Receptor Beta, and HIFs Cooperate in the Activation of a Prognostic Transcriptional Pattern in Aggressive Human Prostate Cancer. J. Clin. Investig. 2009, 119, 1093–1108. [Google Scholar] [CrossRef]
- Aiello, A.; Bacci, L.; Re, A.; Ripoli, C.; Pierconti, F.; Pinto, F.; Masetti, R.; Grassi, C.; Gaetano, C.; Bassi, P.F.; et al. MALAT1 and HOTAIR Long Non-Coding RNAs Play Opposite Role in Estrogen-Mediated Transcriptional Regulation in Prostate Cancer Cells. Sci. Rep. 2016, 6, 38414. [Google Scholar] [CrossRef]
- Nanni, S.; Aiello, A.; Re, A.; Guffanti, A.; Benvenuti, V.; Colussi, C.; Castro-Vega, L.J.; Felsani, A.; Londono-Vallejo, A.; Capogrossi, M.C.; et al. Estrogen-Dependent Dynamic Profile of eNOS-DNA Associations in Prostate Cancer. PLoS ONE 2013, 8, e62522. [Google Scholar] [CrossRef]
- Nanni, S.; Priolo, C.; Grasselli, A.; D’Eletto, M.; Merola, R.; Moretti, F.; Gallucci, M.; De Carli, P.; Sentinelli, S.; Cianciulli, A.M.; et al. Epithelial-Restricted Gene Profile of Primary Cultures from Human Prostate Tumors: A Molecular Approach to Predict Clinical Behavior of Prostate Cancer. Mol. Cancer. Res. 2006, 4, 79–92. [Google Scholar] [CrossRef]
- Re, A.; Aiello, A.; Nanni, S.; Grasselli, A.; Benvenuti, V.; Pantisano, V.; Strigari, L.; Colussi, C.; Ciccone, S.; Mazzetti, A.P.; et al. Silencing of GSTP1, a Prostate Cancer Prognostic Gene, by the Estrogen Receptor-Beta and Endothelial Nitric Oxide Synthase Complex. Mol. Endocrinol. 2011, 25, 2003–2016. [Google Scholar] [CrossRef]
- Vennin, C.; Spruyt, N.; Robin, Y.M.; Chassat, T.; Le Bourhis, X.; Adriaenssens, E. The Long Non-Coding RNA 91H Increases Aggressive Phenotype of Breast Cancer Cells and Up-Regulates H19/IGF2 Expression through Epigenetic Modifications. Cancer Lett. 2017, 385, 198–206. [Google Scholar] [CrossRef]
- Lelli, A.; Nolan, K.A.; Santambrogio, S.; Goncalves, A.F.; Schonenberger, M.J.; Guinot, A.; Frew, I.J.; Marti, H.H.; Hoogewijs, D.; Wenger, R.H. Induction of Long Noncoding RNA MALAT1 in Hypoxic Mice. Hypoxia 2015, 3, 45–52. [Google Scholar]
- Maxwell, P.H.; Wiesener, M.S.; Chang, G.W.; Clifford, S.C.; Vaux, E.C.; Cockman, M.E.; Wykoff, C.C.; Pugh, C.W.; Maher, E.R.; Ratcliffe, P.J. The Tumour Suppressor Protein VHL Targets Hypoxia-Inducible Factors for Oxygen-Dependent Proteolysis. Nature 1999, 399, 271–275. [Google Scholar] [CrossRef]
- Shinojima, T.; Oya, M.; Takayanagi, A.; Mizuno, R.; Shimizu, N.; Murai, M. Renal Cancer Cells Lacking Hypoxia Inducible Factor (HIF)-1alpha Expression Maintain Vascular Endothelial Growth Factor Expression through HIF-2alpha. Carcinogenesis 2007, 28, 529–536. [Google Scholar] [CrossRef]
- Re, A.; Colussi, C.; Nanni, S.; Aiello, A.; Bacci, L.; Grassi, C.; Pontecorvi, A.; Farsetti, A. Nucleoporin 153 Regulates Estrogen-Dependent Nuclear Translocation of Endothelial Nitric Oxide Synthase and Estrogen Receptor Beta in Prostate Cancer. Oncotarget 2018, 9, 27985–27997. [Google Scholar] [CrossRef]
- Fournel, M.; Bonfils, C.; Hou, Y.; Yan, P.T.; Trachy-Bourget, M.C.; Kalita, A.; Liu, J.; Lu, A.H.; Zhou, N.Z.; Robert, M.F.; et al. MGCD0103, a Novel Isotype-Selective Histone Deacetylase Inhibitor, has Broad Spectrum Antitumor Activity in Vitro and in Vivo. Mol. Cancer Ther. 2008, 7, 759–768. [Google Scholar] [CrossRef]
- Rodriguez, M.; Aladowicz, E.; Lanfrancone, L.; Goding, C.R. Tbx3 Represses E-Cadherin Expression and Enhances Melanoma Invasiveness. Cancer Res. 2008, 68, 7872–7881. [Google Scholar] [CrossRef]
- Luo, M.; Li, Z.; Wang, W.; Zeng, Y.; Liu, Z.; Qiu, J. Long Non-Coding RNA H19 Increases Bladder Cancer Metastasis by Associating with EZH2 and Inhibiting E-Cadherin Expression. Cancer Lett. 2013, 333, 213–221. [Google Scholar] [CrossRef]
- Prieto-Garcia, E.; Diaz-Garcia, C.V.; Garcia-Ruiz, I.; Agullo-Ortuno, M.T. Epithelial-to-Mesenchymal Transition in Tumor Progression. Med. Oncol. 2017, 34, 122. [Google Scholar] [CrossRef]
- Weber, G.F.; Bjerke, M.A.; DeSimone, D.W. Integrins and Cadherins Join Forces to Form Adhesive Networks. J. Cell Sci. 2011, 124, 1183–1193. [Google Scholar] [CrossRef]
- Ayesh, S.; Matouk, I.; Schneider, T.; Ohana, P.; Laster, M.; Al-Sharef, W.; De-Groot, N.; Hochberg, A. Possible Physiological Role of H19 RNA. Mol. Carcinog. 2002, 35, 63–74. [Google Scholar] [CrossRef]
- Chen, Y.T.; Zhu, F.; Lin, W.R.; Ying, R.B.; Yang, Y.P.; Zeng, L.H. The Novel EZH2 Inhibitor, GSK126, Suppresses Cell Migration and Angiogenesis via Down-Regulating VEGF-A. Cancer Chemother. Pharmacol. 2016, 77, 757–765. [Google Scholar] [CrossRef]
- Hashizume, R.; Andor, N.; Ihara, Y.; Lerner, R.; Gan, H.; Chen, X.; Fang, D.; Huang, X.; Tom, M.W.; Ngo, V.; et al. Pharmacologic Inhibition of Histone Demethylation as a Therapy for Pediatric Brainstem Glioma. Nat. Med. 2014, 20, 1394–1396. [Google Scholar] [CrossRef]
- Tamgue, O.; Chai, C.S.; Hao, L.; Zambe, J.C.; Huang, W.W.; Zhang, B.; Lei, M.; Wei, Y.M. Triptolide Inhibits Histone Methyltransferase EZH2 and Modulates the Expression of its Target Genes in Prostate Cancer Cells. Asian Pac. J. Cancer Prev. 2013, 14, 5663–5669. [Google Scholar] [CrossRef]
- Qing, G.; Simon, M.C. Hypoxia Inducible Factor-2alpha: A Critical Mediator of Aggressive Tumor Phenotypes. Curr. Opin. Genet. Dev. 2009, 19, 60–66. [Google Scholar] [CrossRef]
- Yang, J.; Harris, A.L.; Davidoff, A.M. Hypoxia and Hormone-Mediated Pathways Converge at the Histone Demethylase KDM4B in Cancer. Int. J. Mol. Sci. 2018, 19, 240. [Google Scholar] [CrossRef]
- Shin, S.; Im, H.J.; Kwon, Y.J.; Ye, D.J.; Baek, H.S.; Kim, D.; Choi, H.K.; Chun, Y.J. Human Steroid Sulfatase Induces Wnt/Beta-Catenin Signaling and Epithelial-Mesenchymal Transition by Upregulating Twist1 and HIF-1alpha in Human Prostate and Cervical Cancer Cells. Oncotarget 2017, 8, 61604–61617. [Google Scholar] [CrossRef]
- Byrne, N.M.; Nesbitt, H.; Ming, L.; McKeown, S.R.; Worthington, J.; McKenna, D.J. Androgen Deprivation in LNCaP Prostate Tumour Xenografts Induces Vascular Changes and Hypoxic Stress, Resulting in Promotion of Epithelial-to-Mesenchymal Transition. Br. J. Cancer 2016, 114, 659–668. [Google Scholar] [CrossRef]
- Mak, P.; Leav, I.; Pursell, B.; Bae, D.; Yang, X.; Taglienti, C.A.; Gouvin, L.M.; Sharma, V.M.; Mercurio, A.M. ERbeta Impedes Prostate Cancer EMT by Destabilizing HIF-1alpha and Inhibiting VEGF-Mediated Snail Nuclear Localization: Implications for Gleason Grading. Cancer Cell 2010, 17, 319–332. [Google Scholar] [CrossRef]
- Yoshioka, T.; Otero, J.; Chen, Y.; Kim, Y.M.; Koutcher, J.A.; Satagopan, J.; Reuter, V.; Carver, B.; de Stanchina, E.; Enomoto, K.; et al. Beta4 Integrin Signaling Induces Expansion of Prostate Tumor Progenitors. J. Clin. Investig. 2013, 123, 682–699. [Google Scholar]
- Stewart, R.L.; O’Connor, K.L. Clinical Significance of the Integrin alpha6beta4 in Human Malignancies. Lab. Investig. 2015, 95, 976–986. [Google Scholar] [CrossRef]
- Harryman, W.L.; Hinton, J.P.; Rubenstein, C.P.; Singh, P.; Nagle, R.B.; Parker, S.J.; Knudsen, B.S.; Cress, A.E. The Cohesive Metastasis Phenotype in Human Prostate Cancer. Biochim. Biophys. Acta Rev. Cancer 2016, 1866, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Sese, M.; Fuentes, P.; Esteve-Codina, A.; Bejar, E.; McGrail, K.; Thomas, G.; Aasen, T.; Ramon, Y.; Cajal, S. Hypoxia-Mediated Translational Activation of ITGB3 in Breast Cancer Cells Enhances TGF-Beta Signaling and Malignant Features in Vitro and in Vivo. Oncotarget 2017, 8, 114856–114876. [Google Scholar] [CrossRef]
- Befani, C.; Liakos, P. Hypoxia Upregulates Integrin Gene Expression in Microvascular Endothelial Cells and Promotes their Migration and Capillary-Like Tube Formation. Cell Biol. Int. 2017, 41, 769–778. [Google Scholar] [CrossRef]
- Saintier, D.; Burde, M.A.; Rey, J.M.; Maudelonde, T.; de Vernejoul, M.C.; Cohen-Solal, M.E. 17beta-Estradiol Downregulates Beta3-Integrin Expression in Differentiating and Mature Human Osteoclasts. J. Cell. Physiol. 2004, 198, 269–276. [Google Scholar] [CrossRef]
- Reymond, M.J.; Nansel, D.D.; Burrows, G.H.; Neaves, W.B.; Porter, J.C. A New Clonal Strain of Rat Pituitary Tumour Cells: A Model for Non-Regulated Secretion of Prolactin. Eur. J. Endocrinol. 1984, 106, 459–470. [Google Scholar] [CrossRef]
- Ho, J.Y.; Chang, F.W.; Huang, F.S.; Liu, J.M.; Liu, Y.P.; Chen, S.P.; Liu, Y.L.; Cheng, K.C.; Yu, C.P.; Hsu, R.J. Estrogen Enhances the Cell Viability and Motility of Breast Cancer Cells through the ERalpha-DeltaNp63-Integrin Beta4 Signaling Pathway. PLoS ONE 2016, 11, e0148301. [Google Scholar] [CrossRef]
- Folgiero, V.; Bachelder, R.E.; Bon, G.; Sacchi, A.; Falcioni, R.; Mercurio, A.M. The alpha6beta4 Integrin can Regulate ErbB-3 Expression: Implications for alpha6beta4 Signaling and Function. Cancer Res. 2007, 67, 1645–1652. [Google Scholar] [CrossRef]
- Barbati, S.A.; Colussi, C.; Bacci, L.; Aiello, A.; Re, A.; Stigliano, E.; Isidori, A.M.; Grassi, C.; Pontecorvi, A.; Farsetti, A.; et al. Transcription Factor CREM Mediates High Glucose Response in Cardiomyocytes and in a Male Mouse Model of Prolonged Hyperglycemia. Endocrinology 2017, 158, 2391–2405. [Google Scholar] [CrossRef] [Green Version]
- Loria, R.; Laquintana, V.; Bon, G.; Trisciuoglio, D.; Frapolli, R.; Covello, R.; Amoreo, C.A.; Ferraresi, V.; Zoccali, C.; Novello, M.; et al. HMGA1/E2F1 Axis and NFkB Pathways Regulate LPS Progression and Trabectedin Resistance. Oncogene 2018, 37, 5926–5938. [Google Scholar] [CrossRef]
- Kopec, A.M.; Rivera, P.D.; Lacagnina, M.J.; Hanamsagar, R.; Bilbo, S.D. Optimized Solubilization of TRIzol-Precipitated Protein Permits Western Blotting Analysis to Maximize Data Available from Brain Tissue. J. Neurosci. Methods 2017, 280, 64–76. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCa Patients | Age | PSA (ng/mL) | Pathologic Gleason Score | Pathologic Stage |
|---|---|---|---|---|
| A | 61 | 8.5 | 7 (4 + 3) | pT2c pNx pMx |
| B | 63 | 5.36 | 7 (3 + 4) | pT2c pNx pMx |
| C | 70 | 18.04 | 7 (3 + 4) | pT2c pN0 pMx |
| D | 68 | 5.75 | 7 (4 + 3) | pT3b pN0 pMx |
| E | 55 | 6 | 7 (4 + 3) | pT3b pN0 pMx |
| H | 78 | 15 | 7 (4 + 3) | pT3a pNx pMx |
| I | 67 | 13.5 | 7 (4 + 3) | pT2c pN0 pMx |
| L | 66 | 8.5 | 7 (4 + 3) | pT3b pN0 pMx |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bacci, L.; Aiello, A.; Ripoli, C.; Loria, R.; Pugliese, D.; Pierconti, F.; Rotili, D.; Strigari, L.; Pinto, F.; Bassi, P.F.; et al. H19-Dependent Transcriptional Regulation of β3 and β4 Integrins Upon Estrogen and Hypoxia Favors Metastatic Potential in Prostate Cancer. Int. J. Mol. Sci. 2019, 20, 4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164012
Bacci L, Aiello A, Ripoli C, Loria R, Pugliese D, Pierconti F, Rotili D, Strigari L, Pinto F, Bassi PF, et al. H19-Dependent Transcriptional Regulation of β3 and β4 Integrins Upon Estrogen and Hypoxia Favors Metastatic Potential in Prostate Cancer. International Journal of Molecular Sciences. 2019; 20(16):4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164012
Chicago/Turabian StyleBacci, Lorenza, Aurora Aiello, Cristian Ripoli, Rossella Loria, Dario Pugliese, Francesco Pierconti, Dante Rotili, Lidia Strigari, Francesco Pinto, Pier Francesco Bassi, and et al. 2019. "H19-Dependent Transcriptional Regulation of β3 and β4 Integrins Upon Estrogen and Hypoxia Favors Metastatic Potential in Prostate Cancer" International Journal of Molecular Sciences 20, no. 16: 4012. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20164012