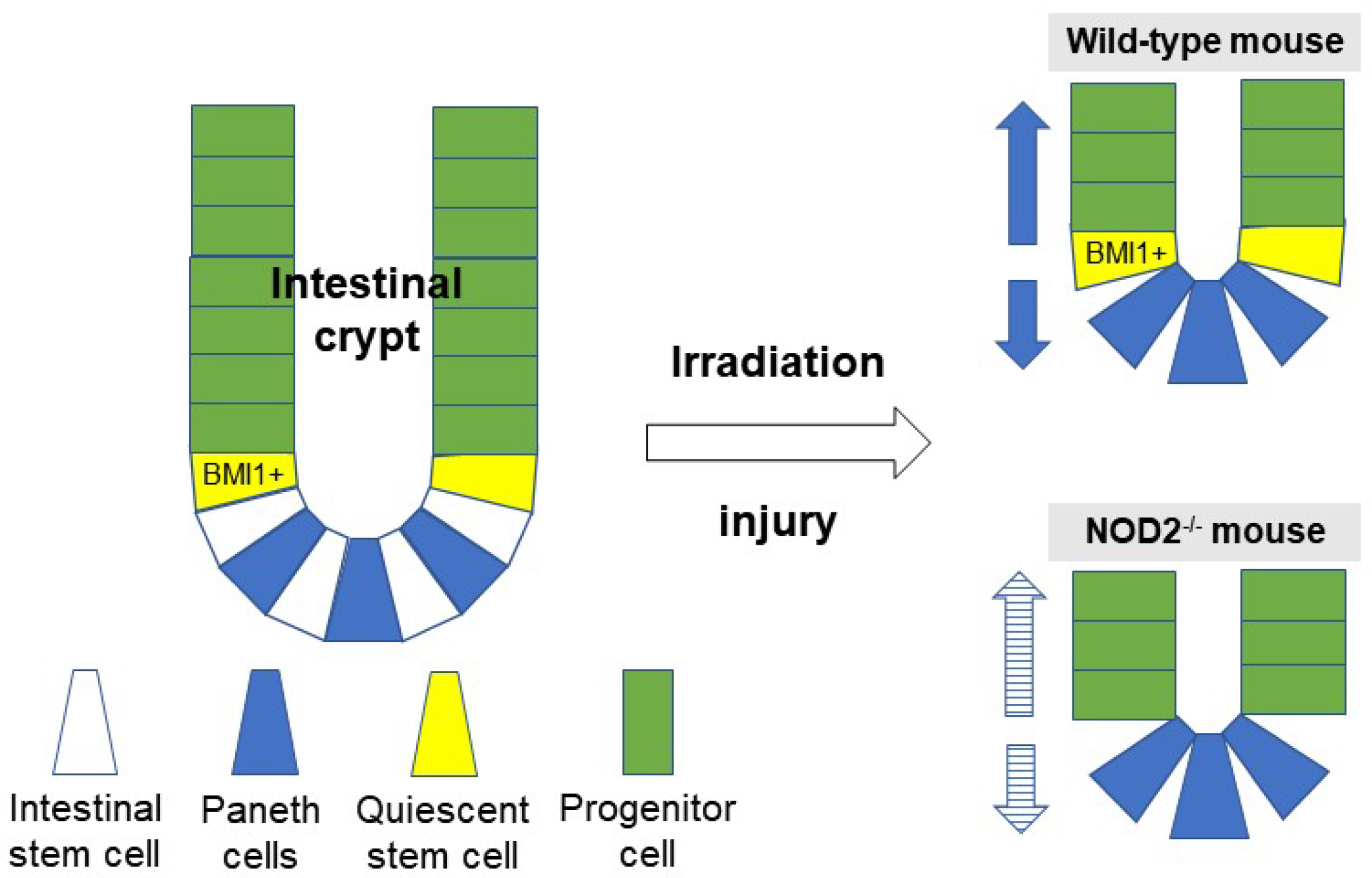
NOD2 Supports Crypt Survival and Epithelial Regeneration after Radiation-Induced Injury

 , , and
, , and 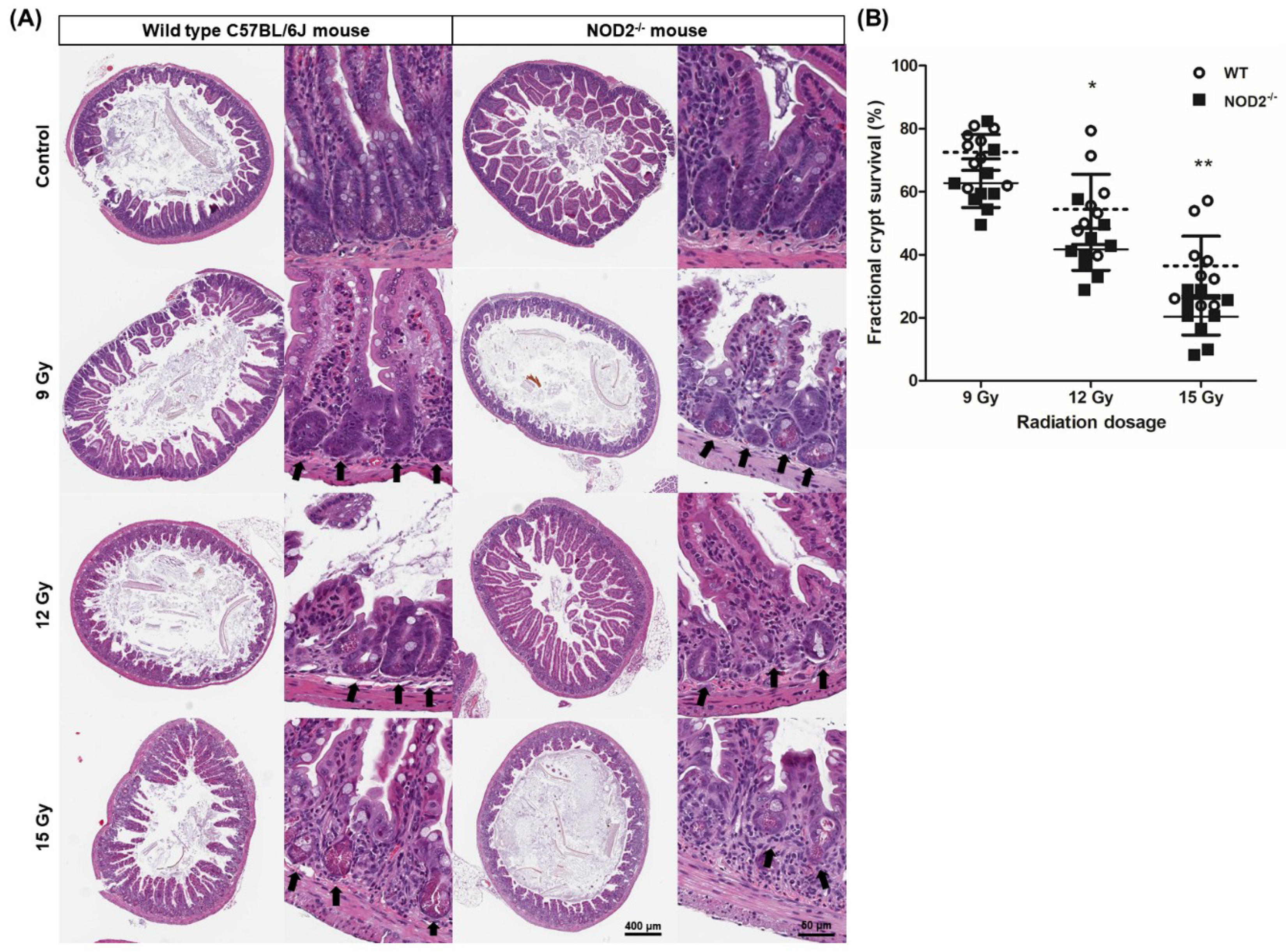{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Dose-Dependent Radiation-Induced Intestinal Damage of NOD2−/− Mice
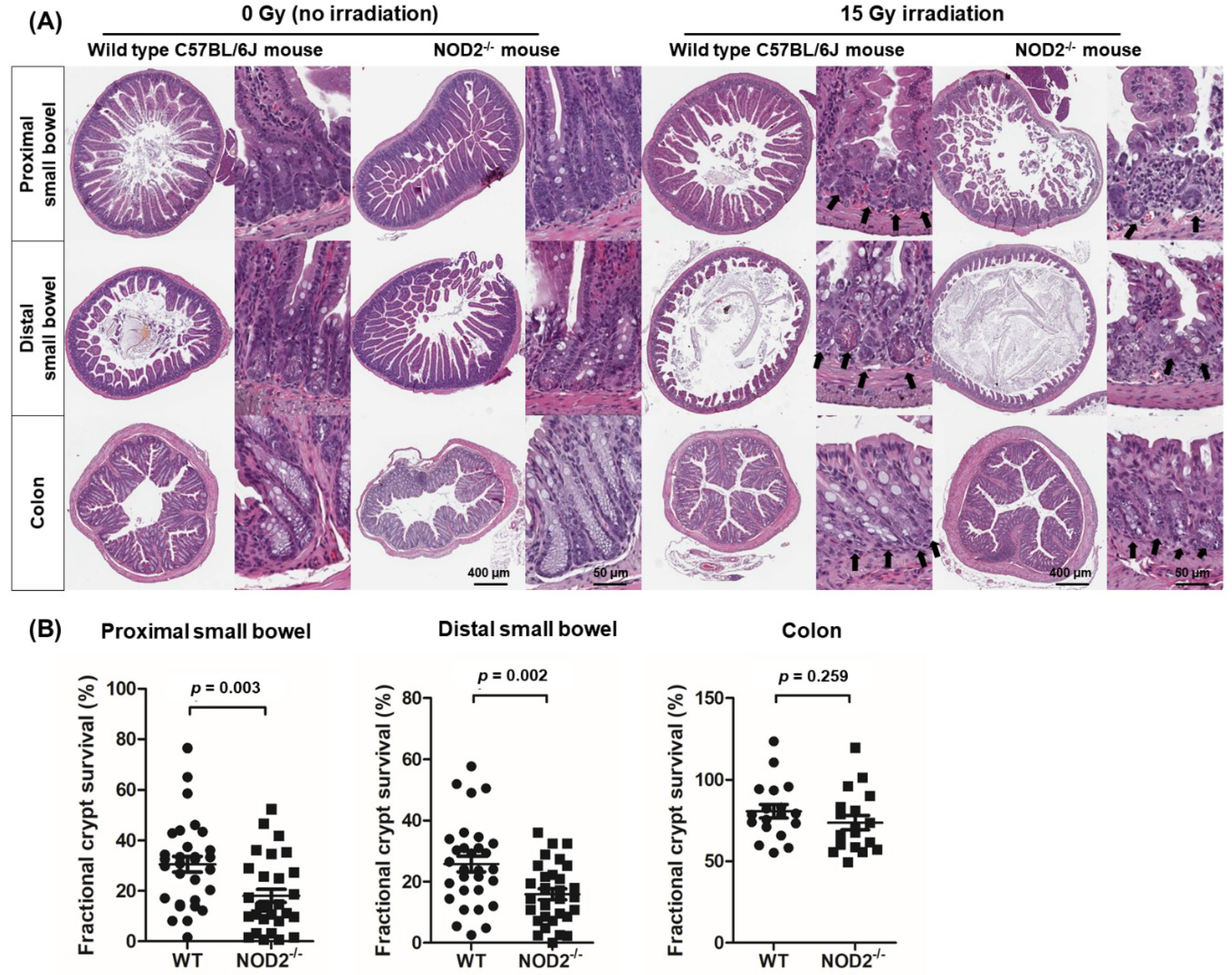
2.2. Location-Dependent Crypt Survival after Radiation Injury
2.3. EdU Assay of the Small Intestine after 15 Gy Irradiation
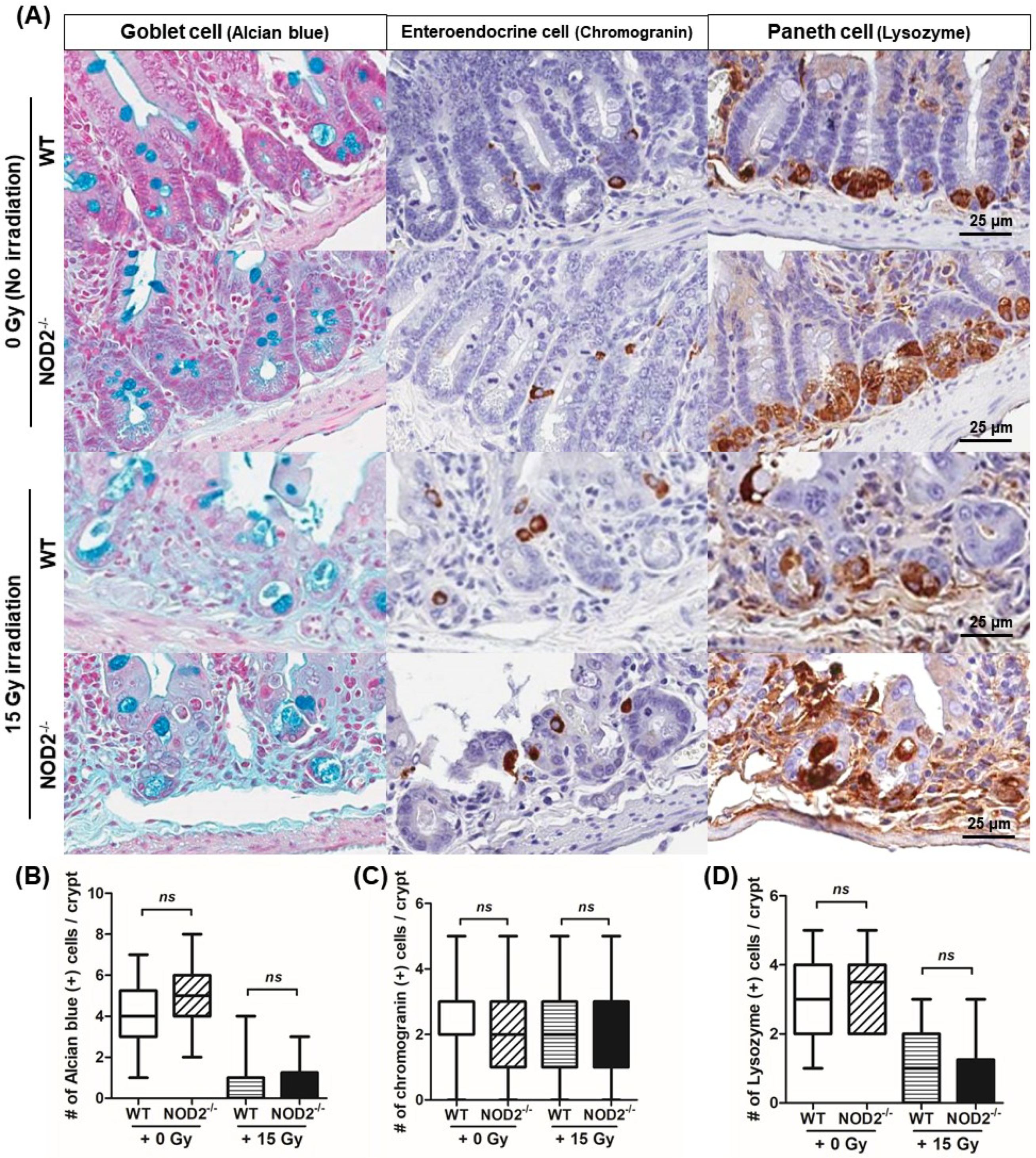
2.4. Differentiation, Regeneration, and Apoptosis of Epithelial Cells Following Irradiation
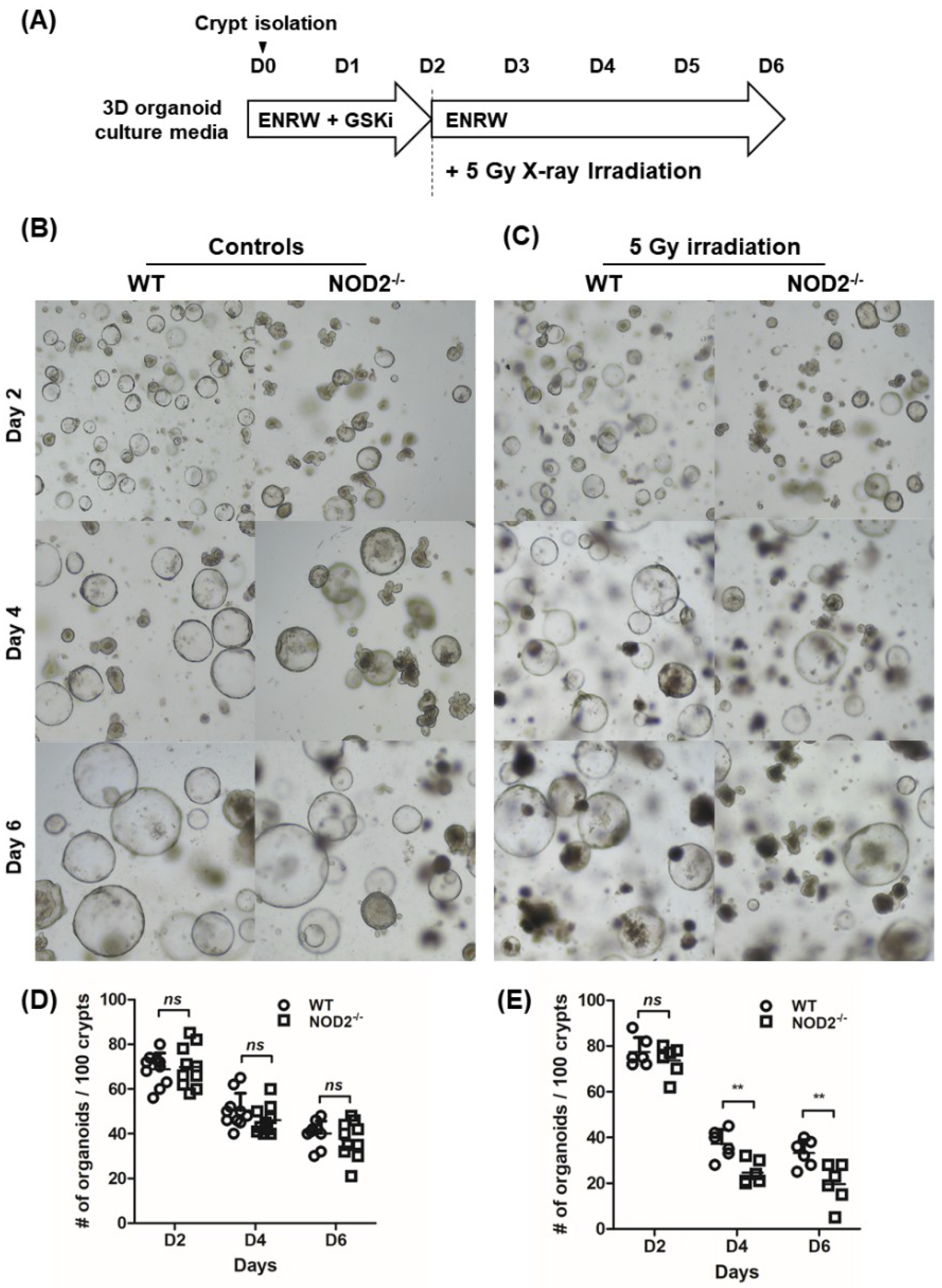
2.5. Organoid Reconstitution Assay
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Mice and Irradiations
4.3. Clonogenic Microcolony Assay
4.4. EdU Assay
4.5. Immunohistochemistry (IHC)
4.6. TUNEL Assay
4.7. Mouse Intestinal Crypt Isolation Culture
4.8. Organoid Reconstitution Assay
4.9. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Liu, J.Z.; van Sommeren, S.; Huang, H.; Ng, S.C.; Alberts, R.; Takahashi, A.; Ripke, S.; Lee, J.C.; Jostins, L.; Shah, T.; et al. Association analyses identify 38 susceptibility loci for inflammatory bowel disease and highlight shared genetic risk across populations. Nat. Genet. 2015, 47, 979–986. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, M.; Vermeire, S. NOD2 and bacterial recognition as therapeutic targets for Crohn’s disease. Expert Opin. Ther. Targets 2017, 21, 1123–1139. [Google Scholar] [CrossRef] [PubMed]
- Strober, W.; Asano, N.; Fuss, I.; Kitani, A.; Watanabe, T. Cellular and molecular mechanisms underlying NOD2 risk-associated polymorphisms in Crohn’s disease. Immunol. Rev. 2014, 260, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Philpott, D.J.; Girardin, S.E. Crohn’s disease-associated Nod2 mutants reduce IL10 transcription. Nat. Immunol. 2009, 10, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Uno, K.; Tanaka, K.; Yasuda, K. Current status of single-balloon enteroscopy: Insertability and clinical applications. World J. Gastrointest. Endosc. 2015, 7, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Bevins, C.L.; Stange, E.F.; Wehkamp, J. Decreased Paneth cell defensin expression in ileal Crohn’s disease is independent of inflammation, but linked to the NOD2 1007fs genotype. Gut 2009, 58, 882–883; discussion 883–884. [Google Scholar]
- Feerick, C.L.; McKernan, D.P. Understanding the regulation of pattern recognition receptors in inflammatory diseases—A ‘Nod’ in the right direction. Immunology 2017, 150, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Travassos, L.H.; Carneiro, L.A.; Ramjeet, M.; Hussey, S.; Kim, Y.G.; Magalhaes, J.G.; Yuan, L.; Soares, F.; Chea, E.; Le Bourhis, L.; et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nat. Immunol. 2010, 11, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Lala, S.; Ogura, Y.; Osborne, C.; Hor, S.Y.; Bromfield, A.; Davies, S.; Ogunbiyi, O.; Nunez, G.; Keshav, S. Crohn’s disease and the NOD2 gene: A role for paneth cells. Gastroenterology 2003, 125, 47–57. [Google Scholar] [CrossRef]
- Nigro, G.; Rossi, R.; Commere, P.H.; Jay, P.; Sansonetti, P.J. The cytosolic bacterial peptidoglycan sensor Nod2 affords stem cell protection and links microbes to gut epithelial regeneration. Cell Host Microbe 2014, 15, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.; Wang, C.; Pan, Y.; Zeng, Z.; Lee, S.G.; Martin, M.L.; Haimovitz-Friedman, A.; Fuks, Z.; Paty, P.B.; Kolesnick, R. Distinct Levels of Radioresistance in Lgr5(+) Colonic Epithelial Stem Cells versus Lgr5(+) Small Intestinal Stem Cells. Cancer Res. 2017, 77, 2124–2133. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Oudenaarden, A.; Clevers, H. Identifying the stem cell of the intestinal crypt: Strategies and pitfalls. Cell Stem Cell 2012, 11, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.S.; Chia, L.A.; Li, X.; Ootani, A.; Su, J.; Lee, J.Y.; Su, N.; Luo, Y.; Heilshorn, S.C.; Amieva, M.R.; et al. The intestinal stem cell markers Bmi1 and Lgr5 identify two functionally distinct populations. Proc. Natl. Acad. Sci. USA 2012, 109, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van de Wetering, M.; Clevers, H. The intestinal stem cell. Genes Dev. 2008, 22, 1856–1864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Arribillaga, E.; Rodilla, V.; Pellegrinet, L.; Guiu, J.; Iglesias, M.; Roman, A.C.; Gutarra, S.; Gonzalez, S.; Munoz-Canoves, P.; Fernandez-Salguero, P.; et al. Bmi1 regulates murine intestinal stem cell proliferation and self-renewal downstream of Notch. Development 2015, 142, 41–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, D.Y.; Lawrie, W.T.; Abrams, R.A.; Kafonek, D.R.; Bayless, T.M.; Welsh, J.S.; DeWeese, T.L. Acute and late radiotherapy toxicity in patients with inflammatory bowel disease. Int. J. Radiat. Oncol. Biol. Phys. 2001, 51, 455–459. [Google Scholar] [CrossRef]
- White, E.C.; Murphy, J.D.; Chang, D.T.; Koong, A.C. Low Toxicity in Inflammatory Bowel Disease Patients Treated With Abdominal and Pelvic Radiation Therapy. Am. J. Clin. Oncol. 2015, 38, 564–569. [Google Scholar] [CrossRef]
- Yilmaz, O.H.; Katajisto, P.; Lamming, D.W.; Gultekin, Y.; Bauer-Rowe, K.E.; Sengupta, S.; Birsoy, K.; Dursun, A.; Yilmaz, V.O.; Selig, M.; et al. mTORC1 in the Paneth cell niche couples intestinal stem-cell function to calorie intake. Nature 2012, 486, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Vidrich, A.; Buzan, J.M.; Barnes, S.; Reuter, B.K.; Skaar, K.; Ilo, C.; Cominelli, F.; Pizarro, T.; Cohn, S.M. Altered epithelial cell lineage allocation and global expansion of the crypt epithelial stem cell population are associated with ileitis in SAMP1/YitFc mice. Am. J. Pathol. 2005, 166, 1055–1067. [Google Scholar] [CrossRef]
- Lahar, N.; Lei, N.Y.; Wang, J.; Jabaji, Z.; Tung, S.C.; Joshi, V.; Lewis, M.; Stelzner, M.; Martin, M.G.; Dunn, J.C. Intestinal subepithelial myofibroblasts support in vitro and in vivo growth of human small intestinal epithelium. PLoS ONE 2011, 6, e26898. [Google Scholar] [CrossRef] [PubMed]
- Lei, N.Y.; Jabaji, Z.; Wang, J.; Joshi, V.S.; Brinkley, G.J.; Khalil, H.; Wang, F.; Jaroszewicz, A.; Pellegrini, M.; Li, L.; et al. Intestinal subepithelial myofibroblasts support the growth of intestinal epithelial stem cells. PLoS ONE 2014, 9, e84651. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C.; Choi, C.; Kang, H.S.; Shin, S.-W.; Kim, S.-Y.; Park, H.C.; Hong, S.N. NOD2 Supports Crypt Survival and Epithelial Regeneration after Radiation-Induced Injury. Int. J. Mol. Sci. 2019, 20, 4297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174297
Lee C, Choi C, Kang HS, Shin S-W, Kim S-Y, Park HC, Hong SN. NOD2 Supports Crypt Survival and Epithelial Regeneration after Radiation-Induced Injury. International Journal of Molecular Sciences. 2019; 20(17):4297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174297
Chicago/Turabian StyleLee, Chansu, Changhoon Choi, Ho Suk Kang, Sung-Won Shin, Shin-Yeong Kim, Hee Chul Park, and Sung Noh Hong. 2019. "NOD2 Supports Crypt Survival and Epithelial Regeneration after Radiation-Induced Injury" International Journal of Molecular Sciences 20, no. 17: 4297. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174297