The ZT Biopolymer: A Self-Assembling Protein Scaffold for Stem Cell Applications

 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
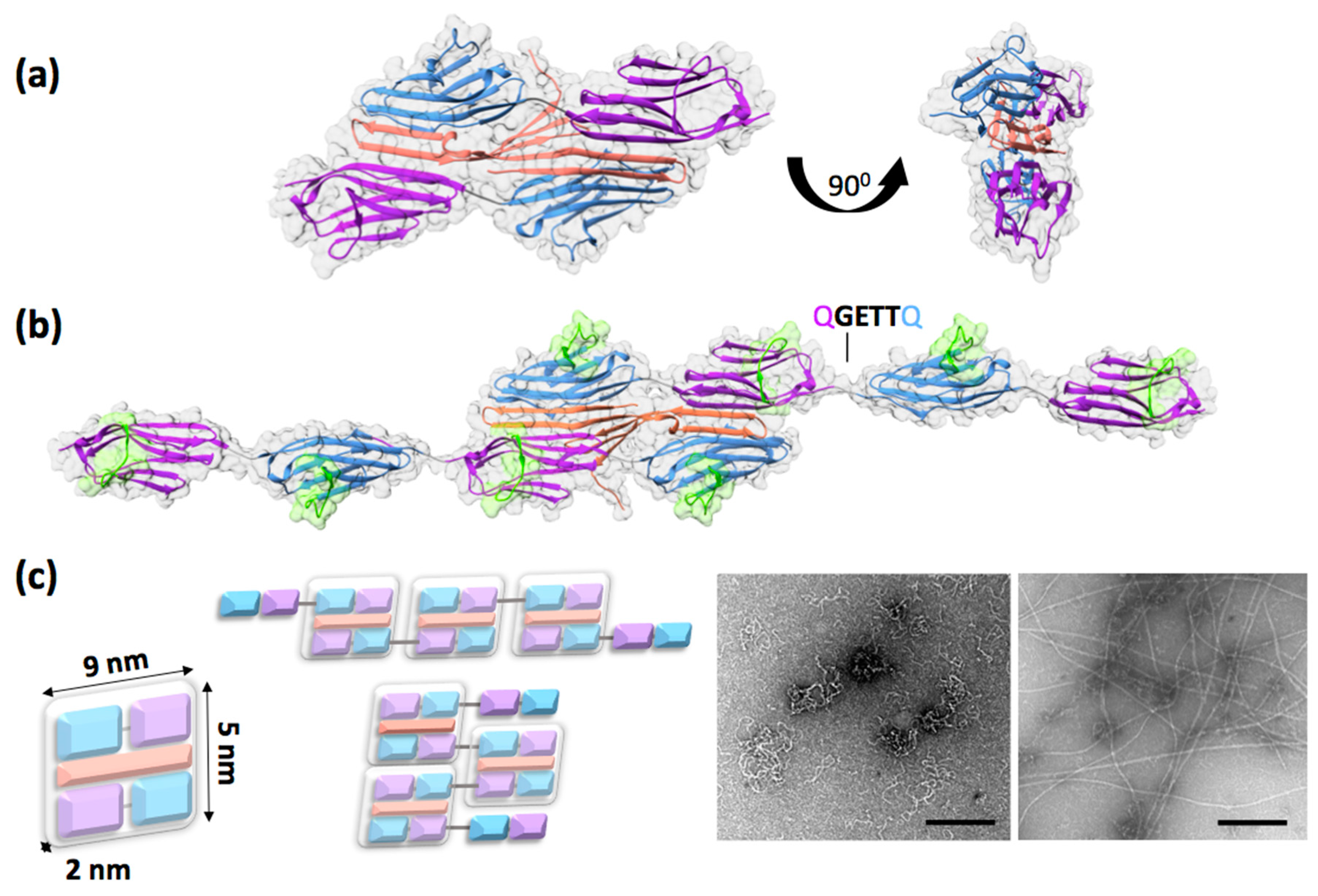
2. The ZT Building Block: A Unique Palindromic Assembly of Titin Z1Z2 with its Binding Partner Telethonin
3. Design of the Self-Assembling ZT Polymer
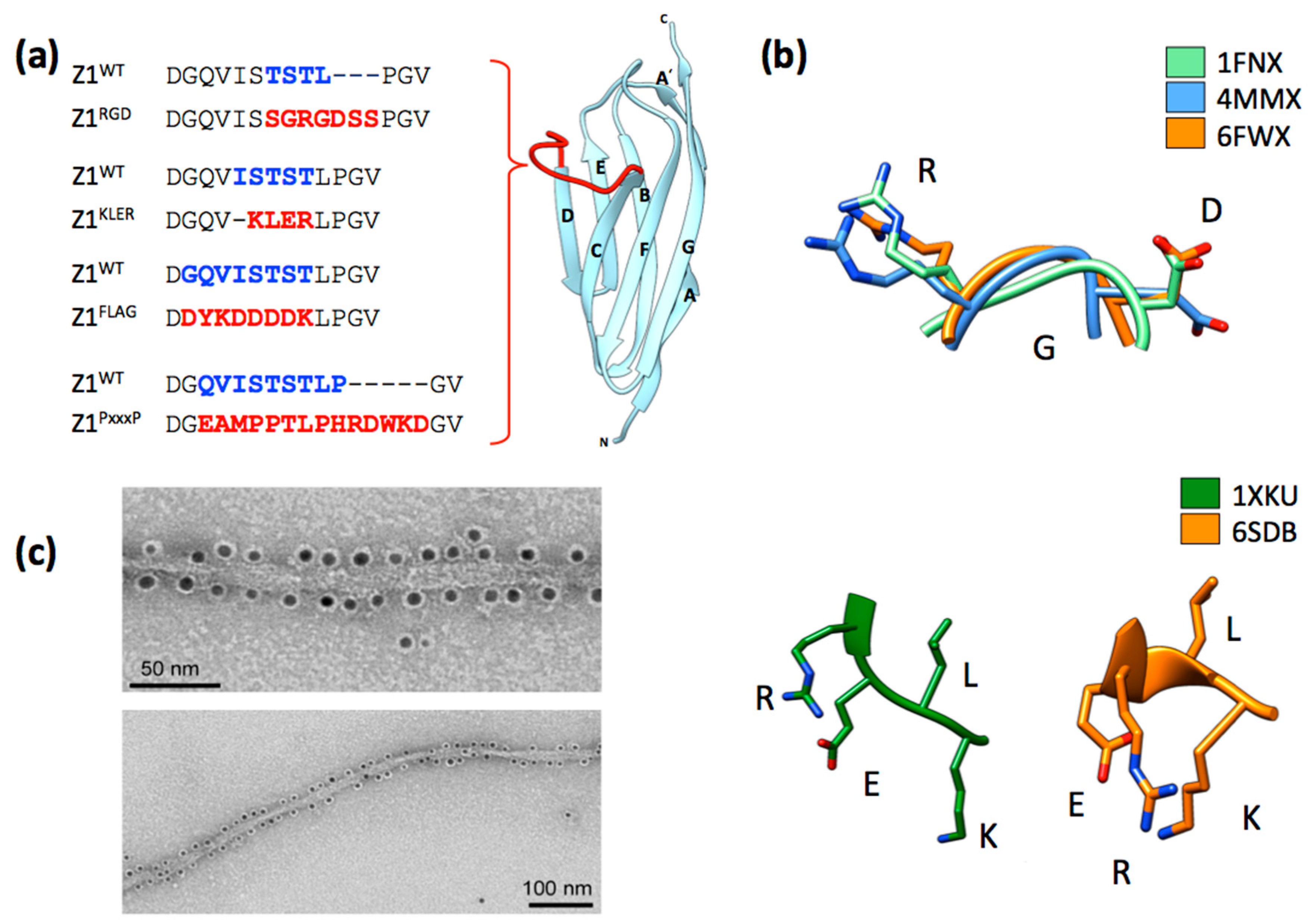
4. Functionalization of the ZT Polymer Using Genetic Engineering: Display of Exogenous Peptide Sequences
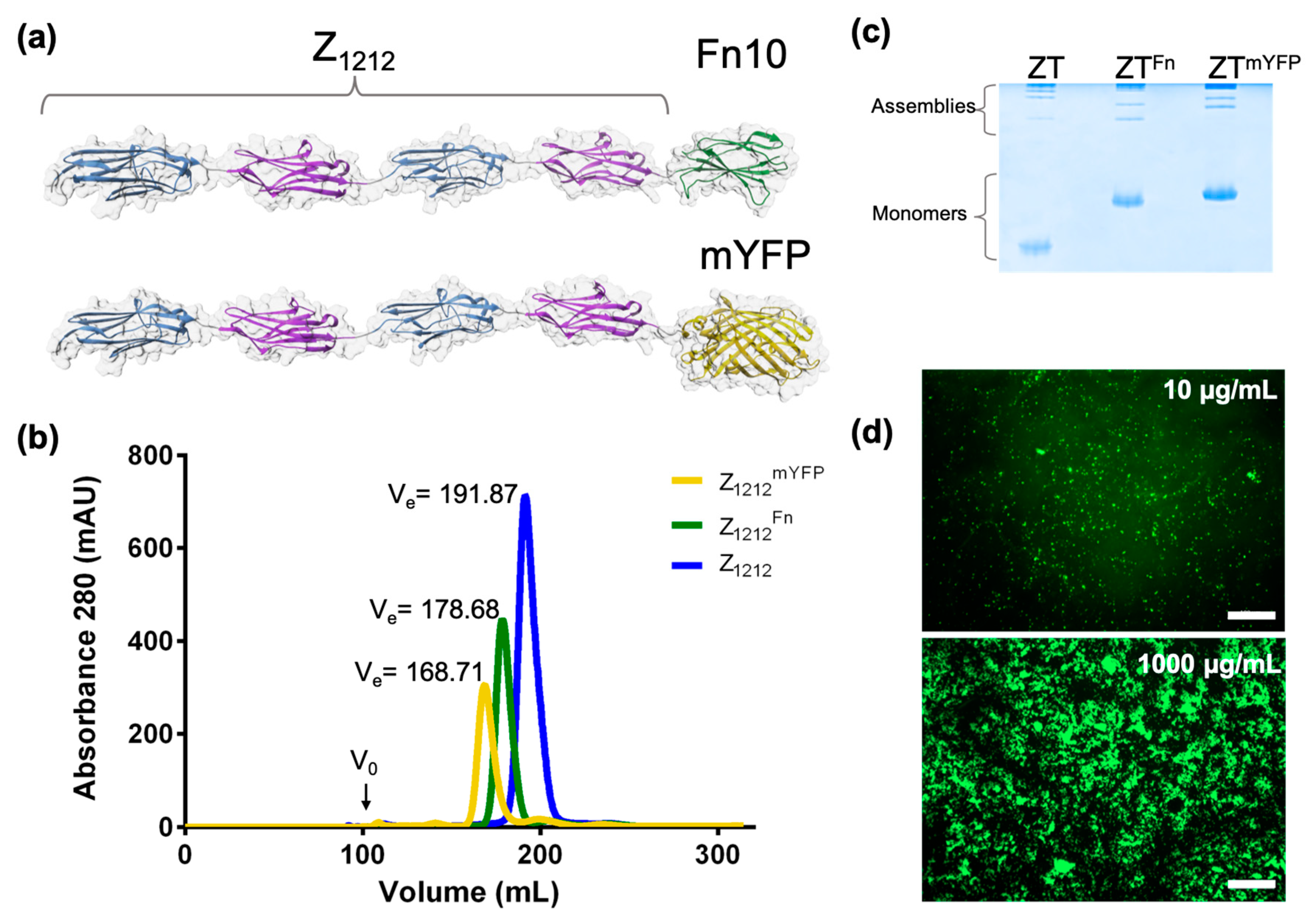
5. Functionalization Through Domain Fusion: Protein Chimeras
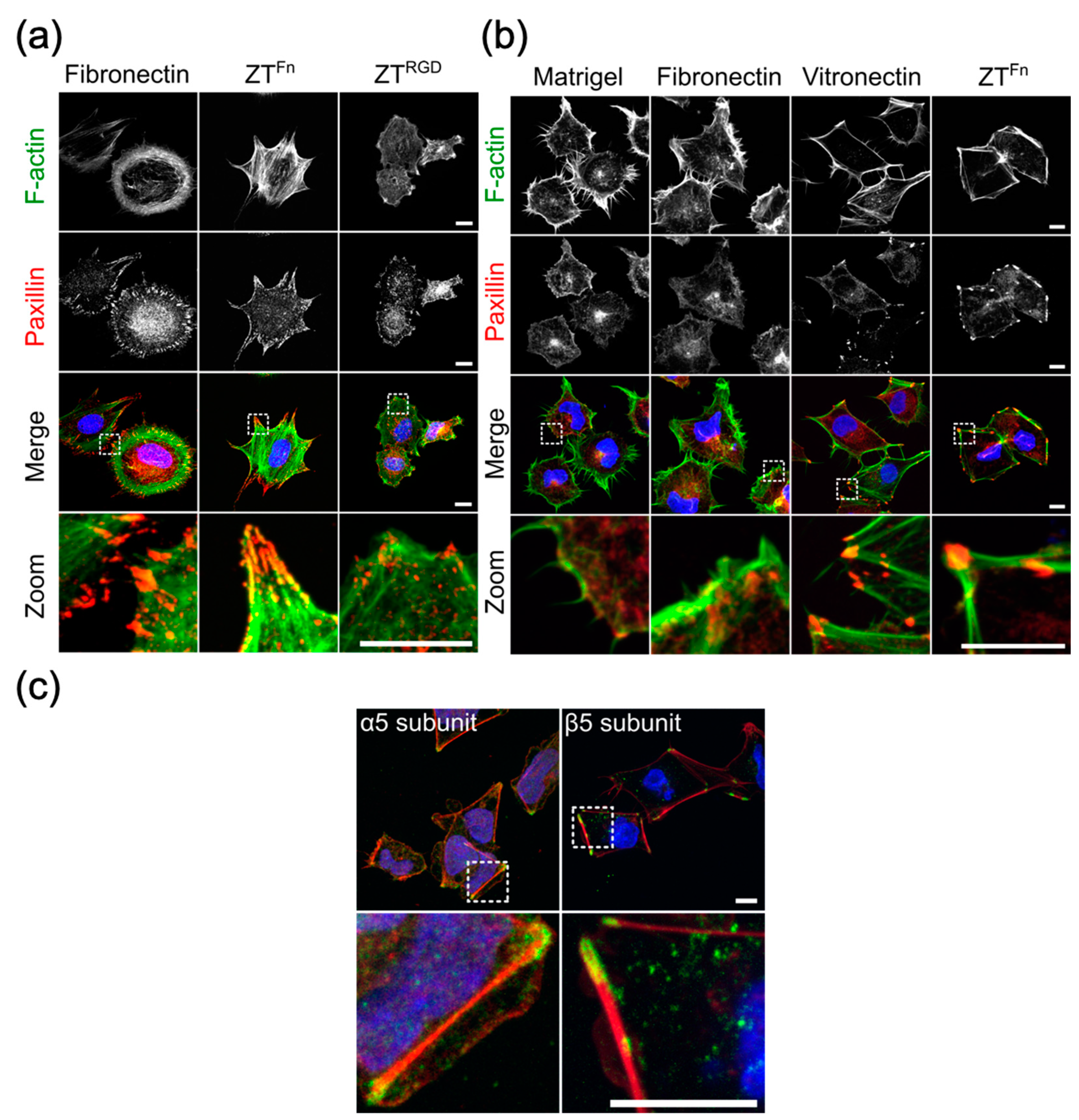
6. The Functionalized ZT Polymer is an Effective Substratum for Stem Cell Culturing
7. Conclusions
Funding
Conflicts of Interest
Substrate Availability
Abbreviations
| AuNPs | Gold nanoparticles |
| CTPR | Consensus tetratricopeptide repeat protein |
| ECM | Extracellular matrix |
| hiPSC | Human induced pluripotent stem cells |
| hESC | Human embryonic stem cells |
| Ig | Immunoglobulin |
| mMSC | Murine mesenchymal stromal cells |
| NMR | Nuclear magnetic resonance |
| PAGE | Polyacrylamide gel electrophoresis |
| PEG | Polyethylene glycol |
| RGD | Arg-Gln-Asp |
| SEC | Size-exclusion chromatography |
| Tel | Telethonin |
| TEM | Transmission electron microscopy |
References
- Kleinman, H.K.; Martin, G.R. Matrigel: Basement membrane matrix with biological activity. Semin. Cancer Biol. 2005, 15, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Hughes, C.S.; Postovit, L.M.; Lajoie, G.A. Matrigel: A complex protein mixture required for optimal growth of cell culture. PROTEOMICS 2010, 10, 1886–1890. [Google Scholar] [CrossRef] [PubMed]
- Benton, G.; Arnaoutova, I.; George, J.; Kleinman, H.K.; Koblinski, J. Matrigel: From discovery and ECM mimicry to assays and models for cancer research. Adv. Drug Deliv. Rev. 2014, 79–80, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Shafiq, M.; Jung, Y.; Kim, S.H. Insight on stem cell preconditioning and instructive biomaterials to enhance cell adhesion, retention, and engraftment for tissue repair. Biomaterials 2016, 90, 85–115. [Google Scholar] [CrossRef] [PubMed]
- Rodin, S.; Domogatskaya, A.; Ström, S.; Hansson, E.M.; Chien, K.R.; Inzunza, J.; Hovatta, O.; Tryggvason, K. Long-term self-renewal of human pluripotent stem cells on human recombinant laminin-511. Nat. Biotechnol. 2010, 28, 611–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prowse, A.B.J.; Doran, M.R.; Cooper-White, J.J.; Chong, F.; Munro, T.P.; Fitzpatrick, J.; Chung, T.-L.; Haylock, D.N.; Gray, P.P.; Wolvetang, E.J. Long term culture of human embryonic stem cells on recombinant vitronectin in ascorbate free media. Biomaterials 2010, 31, 8281–8288. [Google Scholar] [CrossRef]
- Kalaskar, D.M.; Downes, J.E.; Murray, P.; Edgar, D.H.; Williams, R.L. Characterization of the interface between adsorbed fibronectin and human embryonic stem cells. J. R. Soc. Interface 2013, 10. [Google Scholar] [CrossRef]
- Ross, A.M.; Nandivada, H.; Ryan, A.L.; Lahann, J. Synthetic substrates for long-term stem cell culture. Polymer 2012, 53, 2533–2539. [Google Scholar] [CrossRef] [Green Version]
- Hellmund, K.S.; Koksch, B. Self-Assembling Peptides as Extracellular Matrix Mimics to Influence Stem Cell’s Fate. Front. Chem. 2019, 7. [Google Scholar] [CrossRef]
- Melkoumian, Z.; Weber, J.L.; Weber, D.M.; Fadeev, A.G.; Zhou, Y.; Dolley-Sonneville, P.; Yang, J.; Qiu, L.; Priest, C.A.; Shogbon, C.; et al. Synthetic peptide-acrylate surfaces for long-term self-renewal and cardiomyocyte differentiation of human embryonic stem cells. Nat. Biotechnol. 2010, 28, 606–610. [Google Scholar] [CrossRef]
- Higuchi, A.; Kao, S.-H.; Ling, Q.-D.; Chen, Y.-M.; Li, H.-F.; Alarfaj, A.A.; Munusamy, M.A.; Murugan, K.; Chang, S.-C.; Lee, H.-C.; et al. Long-term xeno-free culture of human pluripotent stem cells on hydrogels with optimal elasticity. Sci. Rep. 2015, 5, 18136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambshead, J.W.; Meagher, L.; Goodwin, J.; Labonne, T.; Ng, E.; Elefanty, A.; Stanley, E.; O’Brien, C.M.; Laslett, A.L. Long-Term Maintenance of Human Pluripotent Stem Cells on cRGDfK-Presenting Synthetic Surfaces. Sci. Rep. 2018, 8, 701. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.; Goktas, M.; Tekinay, A.B.; Guler, M.O.; Hauser, C.A.E.; Mitraki, A. Self-Assembled Proteins and Peptides as Scaffolds for Tissue Regeneration. Adv. Healthc. Mater. 2015, 4, 2557–2586. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Discovery and design of self-assembling peptides. Interface Focus 2017, 7, 20170028. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Collier, J.H. α-Helical Coiled Coil Peptide Materials for Biomedical Applications. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9. [Google Scholar] [CrossRef]
- Seroski, D.T.; Hudalla, G.A. Self-Assembled Peptide and Protein Nanofibers for Biomedical Applications. Biomed. Appl. Funct. Nanomater. Elsevier 2018, 569–598. [Google Scholar]
- Beesley, J.L.; Woolfson, D.N. The de novo design of α-helical peptides for supramolecular self-assembly. Curr. Opin. Biotechnol. 2019, 58, 175–182. [Google Scholar] [CrossRef]
- Cai, L.; Heilshorn, S.C. Designing ECM-mimetic materials using protein engineering. Acta Biomater. 2014, 10, 1751–1760. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.J.; Appel, E.A.; Meijer, E.W.; Langer, R. Supramolecular biomaterials. Nat. Mater. 2016, 15, 13–26. [Google Scholar] [CrossRef]
- Wang, Y.; Katyal, P.; Montclare, J.K. Protein-Engineered Functional Materials. Adv. Healthc. Mater. 2019, 8, e1801374. [Google Scholar] [CrossRef]
- Mejias, S.H.; Aires, A.; Couleaud, P.; Cortajarena, A.L. Designed Repeat Proteins as Building Blocks for Nanofabrication. Adv. Exp. Med. Biol. 2016, 940, 61–81. [Google Scholar] [PubMed] [Green Version]
- Cannon, K.A.; Ochoa, J.M.; Yeates, T.O. High-symmetry protein assemblies: Patterns and emerging applications. Curr. Opin. Struct. Biol. 2019, 55, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Protein Assemblies: Nature-Inspired and Designed Nanostructures. Biomacromolecules 2019, 20, 1829–1848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, P.; Pinotsis, N.; Lange, S.; Song, Y.-H.; Popov, A.; Mavridis, I.; Mayans, O.M.; Gautel, M.; Wilmanns, M. Palindromic assembly of the giant muscle protein titin in the sarcomeric Z-disk. Nature 2006, 439, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Remaut, H.; Waksman, G. Protein-protein interaction through beta-strand addition. Trends Biochem. Sci. 2006, 31, 436–444. [Google Scholar] [CrossRef]
- Benian, G.M.; Mayans, O. Titin and Obscurin: Giants Holding Hands and Discovery of a New Ig Domain Subset. J. Mol. Biol. 2015, 427, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Zou, P.; Gautel, M.; Geerlof, A.; Wilmanns, M.; Koch, M.H.J.; Svergun, D.I. Solution Scattering Suggests Cross-linking Function of Telethonin in the Complex with Titin. J. Biol. Chem. 2003, 278, 2636–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, H.; Su, W.; Wang, P.; Zhang, W.; Sattler, M.; Zou, P. Expression and purification of a difficult sarcomeric protein: Telethonin. Protein Expr. Purif. 2017, 140, 74–80. [Google Scholar] [CrossRef]
- Bertz, M.; Wilmanns, M.; Rief, M. The titin-telethonin complex is a directed, superstable molecular bond in the muscle Z-disk. Proc. Natl. Acad. Sci. USA 2009, 106, 13307–133310. [Google Scholar] [CrossRef] [Green Version]
- Bruning, M.; Kreplak, L.; Leopoldseder, S.; Müller, S.A.; Ringler, P.; Duchesne, L.; Fernig, D.G.; Engel, A.; Ucurum-Fotiadis, Z.; Mayans, O. Bipartite design of a self-fibrillating protein copolymer with nanopatterned peptide display capabilities. Nano Lett. 2010, 10, 4533–4537. [Google Scholar] [CrossRef]
- Politou, A.S.; Thomas, D.J.; Pastore, A. The folding and stability of titin immunoglobulin-like modules, with implications for the mechanism of elasticity. Biophys. J. 1995, 69, 2601–2610. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Svergun, D.I.; Kreplak, L.; Konarev, P.V.; Maco, B.; Labeit, D.; Mayans, O. Poly-Ig tandems from I-band titin share extended domain arrangements irrespective of the distinct features of their modular constituents. J. Muscle Res. Cell Motil. 2005, 26, 355–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Castelmur, E.; Marino, M.; Svergun, D.I.; Kreplak, L.; Ucurum-Fotiadis, Z.; Konarev, P.V.; Urzhumtsev, A.; Labeit, D.; Labeit, S.; Mayans, O. A regular pattern of Ig super-motifs defines segmental flexibility as the elastic mechanism of the titin chain. Proc. Natl. Acad. Sci. USA 2008, 105, 1186–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, M.; Zou, P.; Svergun, D.; Garcia, P.; Edlich, C.; Simon, B.; Wilmanns, M.; Muhle-Goll, C.; Mayans, O. The Ig doublet Z1Z2: A model system for the hybrid analysis of conformational dynamics in Ig tandems from titin. Struct. Lond. Engl. 1993 2006, 14, 1437–1447. [Google Scholar] [CrossRef]
- Lee, E.H.; Hsin, J.; Mayans, O.; Schulten, K. Secondary and tertiary structure elasticity of titin Z1Z2 and a titin chain model. Biophys. J. 2007, 93, 1719–1735. [Google Scholar] [CrossRef]
- Lee, E.H.; Hsin, J.; von Castelmur, E.; Mayans, O.; Schulten, K. Tertiary and secondary structure elasticity of a six-Ig titin chain. Biophys. J. 2010, 98, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Zacharchenko, T.; von Castelmur, E.; Rigden, D.J.; Mayans, O. Structural advances on titin: Towards an atomic understanding of multi-domain functions in myofilament mechanics and scaffolding. Biochem. Soc. Trans. 2015, 43, 850–855. [Google Scholar] [CrossRef]
- Iadanza, M.G.; Jackson, M.P.; Hewitt, E.W.; Ranson, N.A.; Radford, S.E. A new era for understanding amyloid structures and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 755–773. [Google Scholar] [CrossRef] [PubMed]
- Bruning, M.; Barsukov, I.; Franke, B.; Barbieri, S.; Volk, M.; Leopoldseder, S.; Ucurum, Z.; Mayans, O. The intracellular Ig fold: A robust protein scaffold for the engineering of molecular recognition. Protein Eng. Des. Sel. PEDS 2012, 25, 205–212. [Google Scholar] [CrossRef]
- Hill, C.J.; Fleming, J.R.; Mousavinejad, M.; Nicholson, R.; Tzokov, S.B.; Bullough, P.A.; Bogomolovas, J.; Morgan, M.R.; Mayans, O.; Murray, P. Self-Assembling Proteins as High-Performance Substrates for Embryonic Stem Cell Self-Renewal. Adv. Mater. Deerfield Beach Fla 2019, 31, e1807521. [Google Scholar] [CrossRef]
- Halaby, D.M.; Poupon, A.; Mornon, J. The immunoglobulin fold family: Sequence analysis and 3D structure comparisons. Protein Eng. 1999, 12, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. RGD and other recognition sequences for integrins. Annu. Rev. Cell Dev. Biol. 1996, 12, 697–715. [Google Scholar] [CrossRef] [PubMed]
- Hersel, U.; Dahmen, C.; Kessler, H. RGD modified polymers: Biomaterials for stimulated cell adhesion and beyond. Biomaterials 2003, 24, 4385–4415. [Google Scholar] [CrossRef]
- Bellis, S.L. Advantages of RGD peptides for directing cell association with biomaterials. Biomaterials 2011, 32, 4205–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapp, T.G.; Rechenmacher, F.; Neubauer, S.; Maltsev, O.V.; Cavalcanti-Adam, E.A.; Zarka, R.; Reuning, U.; Notni, J.; Wester, H.-J.; Mas-Moruno, C.; et al. A Comprehensive Evaluation of the Activity and Selectivity Profile of Ligands for RGD-binding Integrins. Sci. Rep. 2017, 7, 39805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iozzo, R.V. Matrix proteoglycans: From molecular design to cellular function. Annu. Rev. Biochem. 1998, 67, 609–652. [Google Scholar] [CrossRef] [PubMed]
- Weber, I.T.; Harrison, R.W.; Iozzo, R.V. Model structure of decorin and implications for collagen fibrillogenesis. J. Biol. Chem. 1996, 271, 31767–31770. [Google Scholar] [CrossRef] [PubMed]
- Salinas, C.N.; Anseth, K.S. Decorin moieties tethered into PEG networks induce chondrogenesis of human mesenchymal stem cells. J. Biomed. Mater. Res. A 2009, 90, 456–464. [Google Scholar] [CrossRef]
- Potts, J.R.; Campbell, I.D. Structure and function of fibronectin modules. Matrix Biol. J. Int. Soc. Matrix Biol. 1996, 15, 313–320. [Google Scholar] [CrossRef]
- Hill, C.J.; Murray, P.; Mayans, O. Fusion Proteins Assembling into Scaffolds and Promoting Stem Cell Renewal 2019. Unpublished work.
- Lam, J.; Segura, T. The modulation of MSC integrin expression by RGD presentation. Biomaterials 2013, 34, 3938–3947. [Google Scholar] [CrossRef] [Green Version]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Pytela, R.; Pierschbacher, M.D.; Ruoslahti, E. A 125/115-kDa cell surface receptor specific for vitronectin interacts with the arginine-glycine-aspartic acid adhesion sequence derived from fibronectin. Proc. Natl. Acad. Sci. USA 1985, 82, 5766–5770. [Google Scholar] [CrossRef] [PubMed]
- Aota, S.; Nomizu, M.; Yamada, K.M. The short amino acid sequence Pro-His-Ser-Arg-Asn in human fibronectin enhances cell-adhesive function. J. Biol. Chem. 1994, 269, 24756–24761. [Google Scholar] [PubMed]
- Mas-Moruno, C.; Fraioli, R.; Rechenmacher, F.; Neubauer, S.; Kapp, T.G.; Kessler, H. αvβ3- or α5β1-Integrin-Selective Peptidomimetics for Surface Coating. Angew. Chem. Int. Ed. 2016, 55, 7048–7067. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Byron, A.; Humphries, M.J. Integrin ligands at a glance. J. Cell Sci. 2006, 119, 3901–3903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.; Castelletto, V.; Jones, R.R.; Connon, C.J.; Hamley, I.W. Hydrogelation of self-assembling RGD-based peptides. Soft Matter 2011, 7, 1326–1333. [Google Scholar] [CrossRef]
- Loo, Y.; Lakshmanan, A.; Ni, M.; Toh, L.L.; Wang, S.; Hauser, C.A.E. Peptide Bioink: Self-Assembling Nanofibrous Scaffolds for Three-Dimensional Organotypic Cultures. Nano Lett. 2015, 15, 6919–6925. [Google Scholar] [CrossRef] [PubMed]
- Schaub, N.J.; Le Beux, C.; Miao, J.; Linhardt, R.J.; Alauzun, J.G.; Laurencin, D.; Gilbert, R.J. The Effect of Surface Modification of Aligned Poly-L-Lactic Acid Electrospun Fibers on Fiber Degradation and Neurite Extension. PLoS ONE 2015, 10, e0136780. [Google Scholar] [CrossRef]
- Klinkhammer, K.; Bockelmann, J.; Simitzis, C.; Brook, G.A.; Grafahrend, D.; Groll, J.; Möller, M.; Mey, J.; Klee, D. Functionalization of electrospun fibers of poly(epsilon-caprolactone) with star shaped NCO-poly(ethylene glycol)-stat-poly(propylene glycol) for neuronal cell guidance. J. Mater. Sci. Mater. Med. 2010, 21, 2637–2651. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nesterenko, Y.; Hill, C.J.; Fleming, J.R.; Murray, P.; Mayans, O. The ZT Biopolymer: A Self-Assembling Protein Scaffold for Stem Cell Applications. Int. J. Mol. Sci. 2019, 20, 4299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174299
Nesterenko Y, Hill CJ, Fleming JR, Murray P, Mayans O. The ZT Biopolymer: A Self-Assembling Protein Scaffold for Stem Cell Applications. International Journal of Molecular Sciences. 2019; 20(17):4299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174299
Chicago/Turabian StyleNesterenko, Yevheniia, Christopher J. Hill, Jennifer R. Fleming, Patricia Murray, and Olga Mayans. 2019. "The ZT Biopolymer: A Self-Assembling Protein Scaffold for Stem Cell Applications" International Journal of Molecular Sciences 20, no. 17: 4299. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174299