Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Wheat

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of CLAVATA3/EMBRYO SURROUNDING (CLE) Peptide-Encoding Genes in Wheat
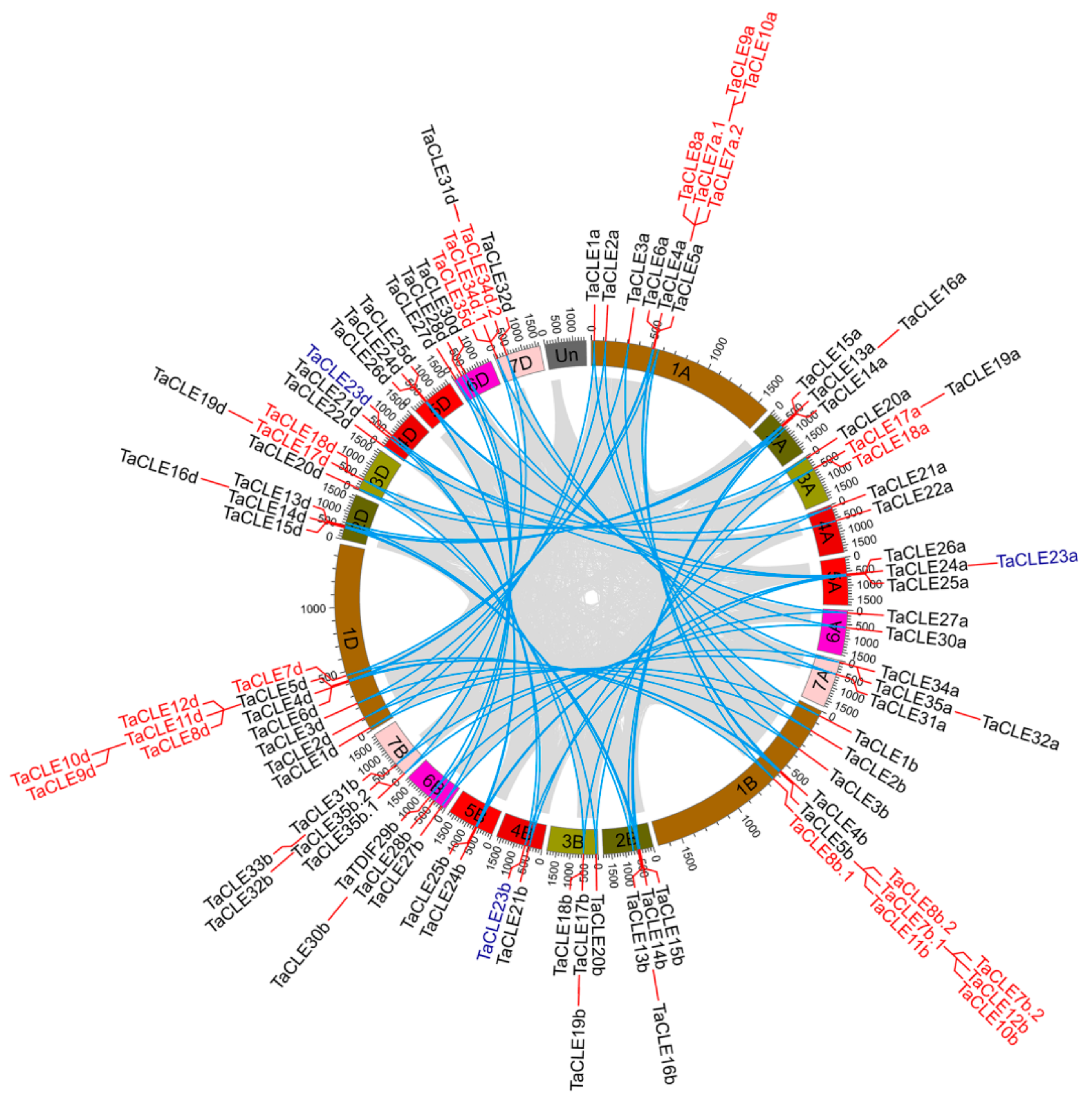
2.2. Gene Duplication Events in the TaCLE Gene Family
2.3. Structure Analysis of the TaCLE Gene Family
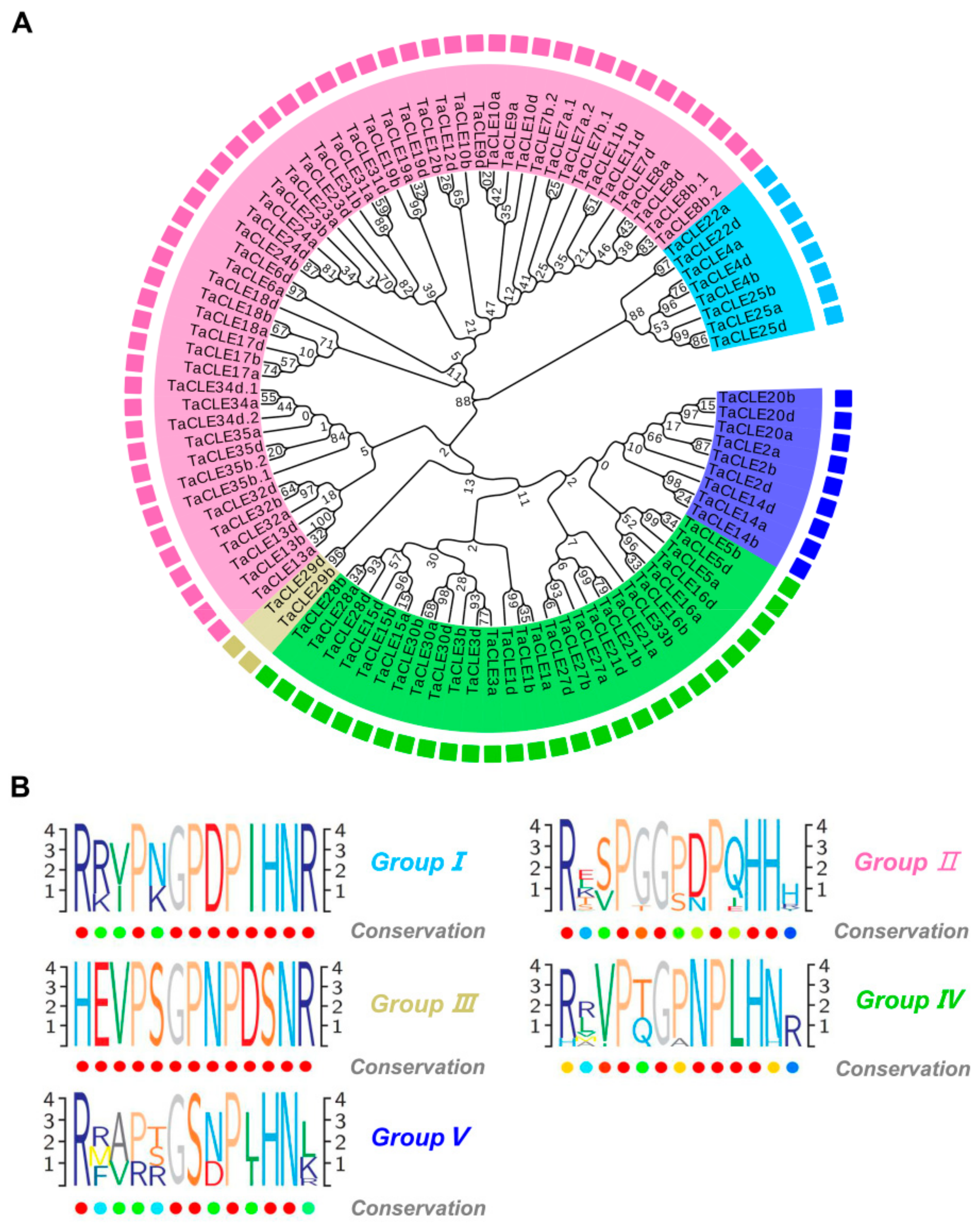
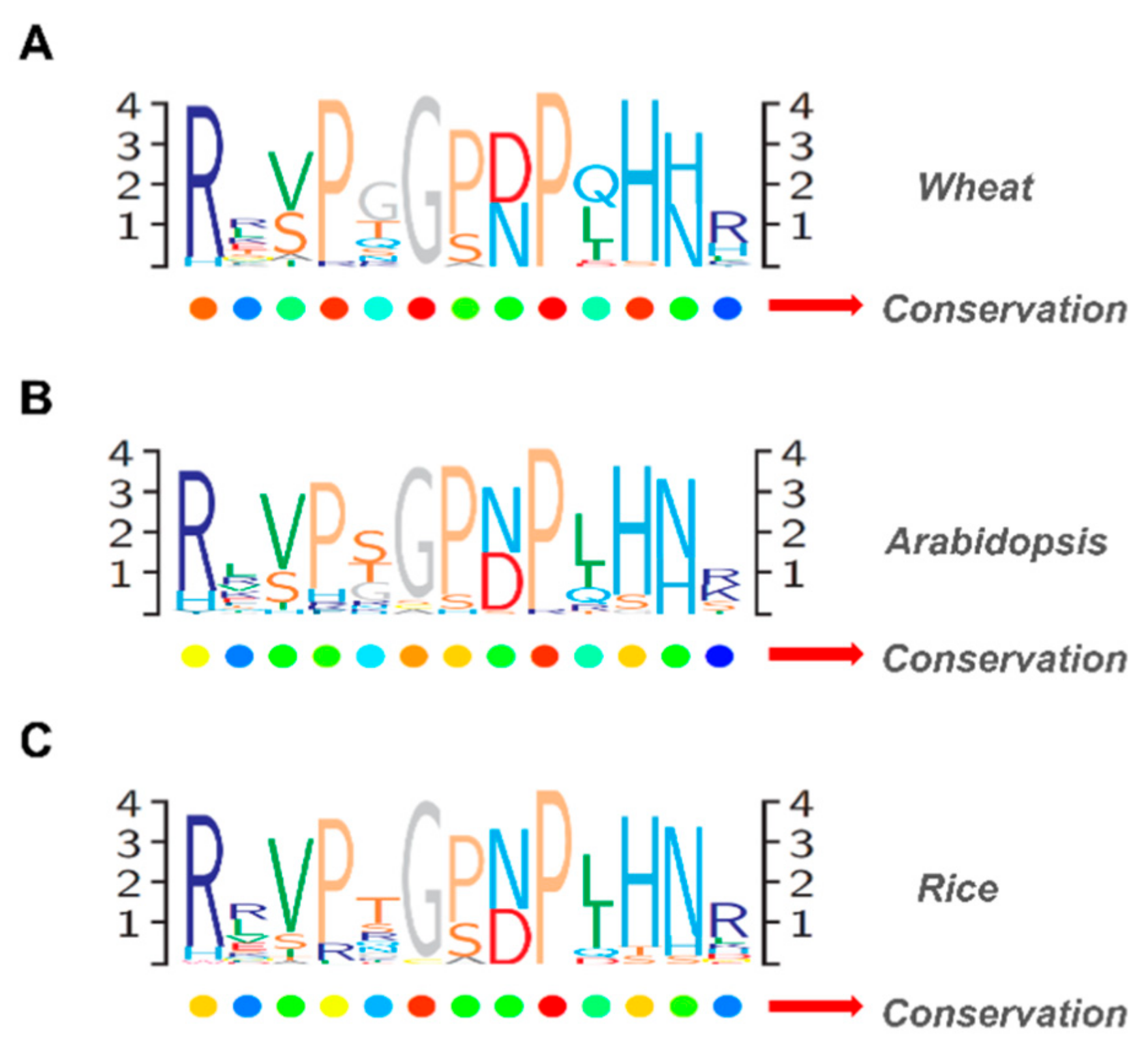
2.4. Categorization and Functional Predictions of Wheat CLAVATA3/EMBRYO SURROUNDING (CLE) Peptides Based on the Analysis of the Phylogenetic Tree and Expression Patterns
2.5. Multi-CLAVATA3/EMBRYO SURROUNDING Peptide-Encoding Genes of Wheat
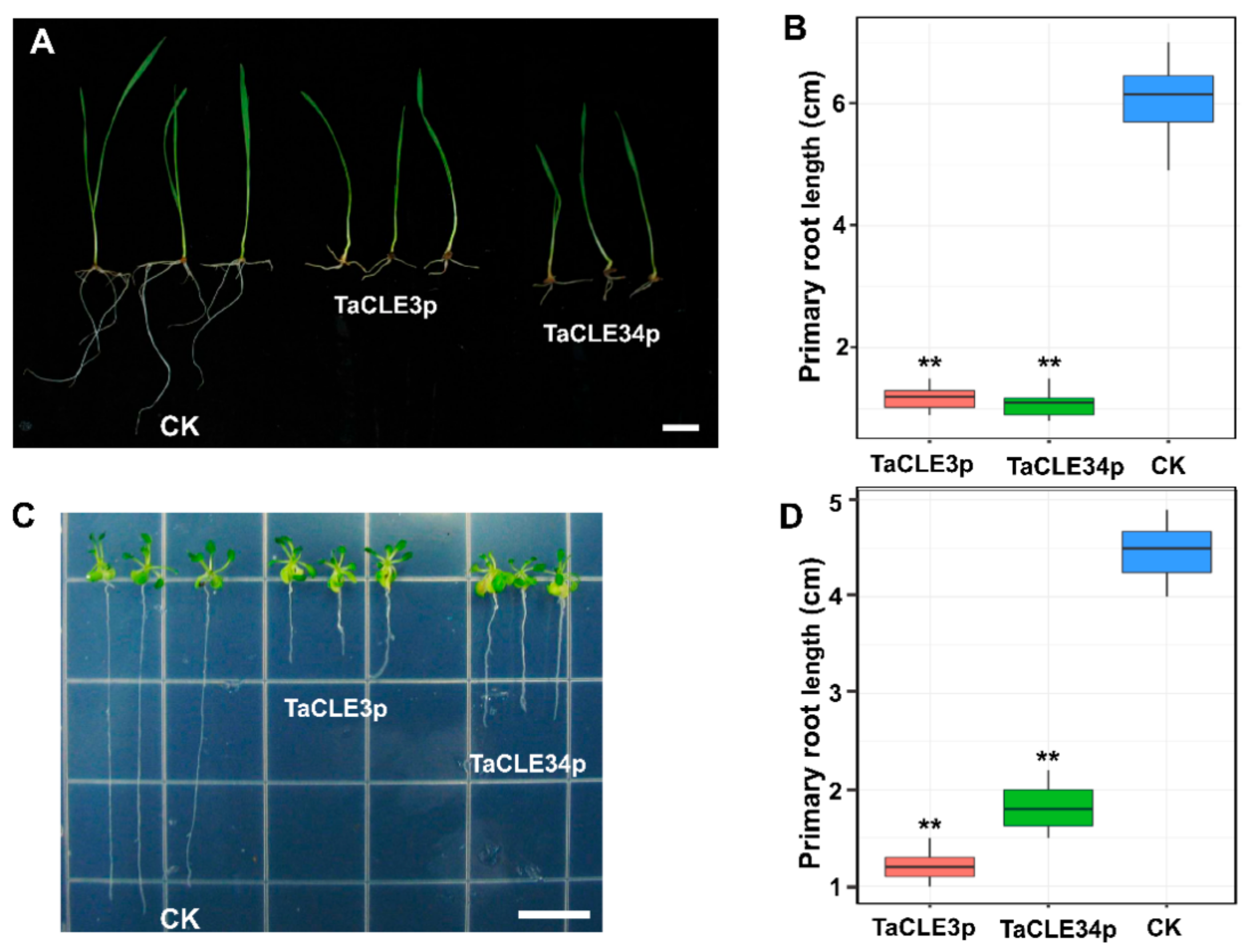
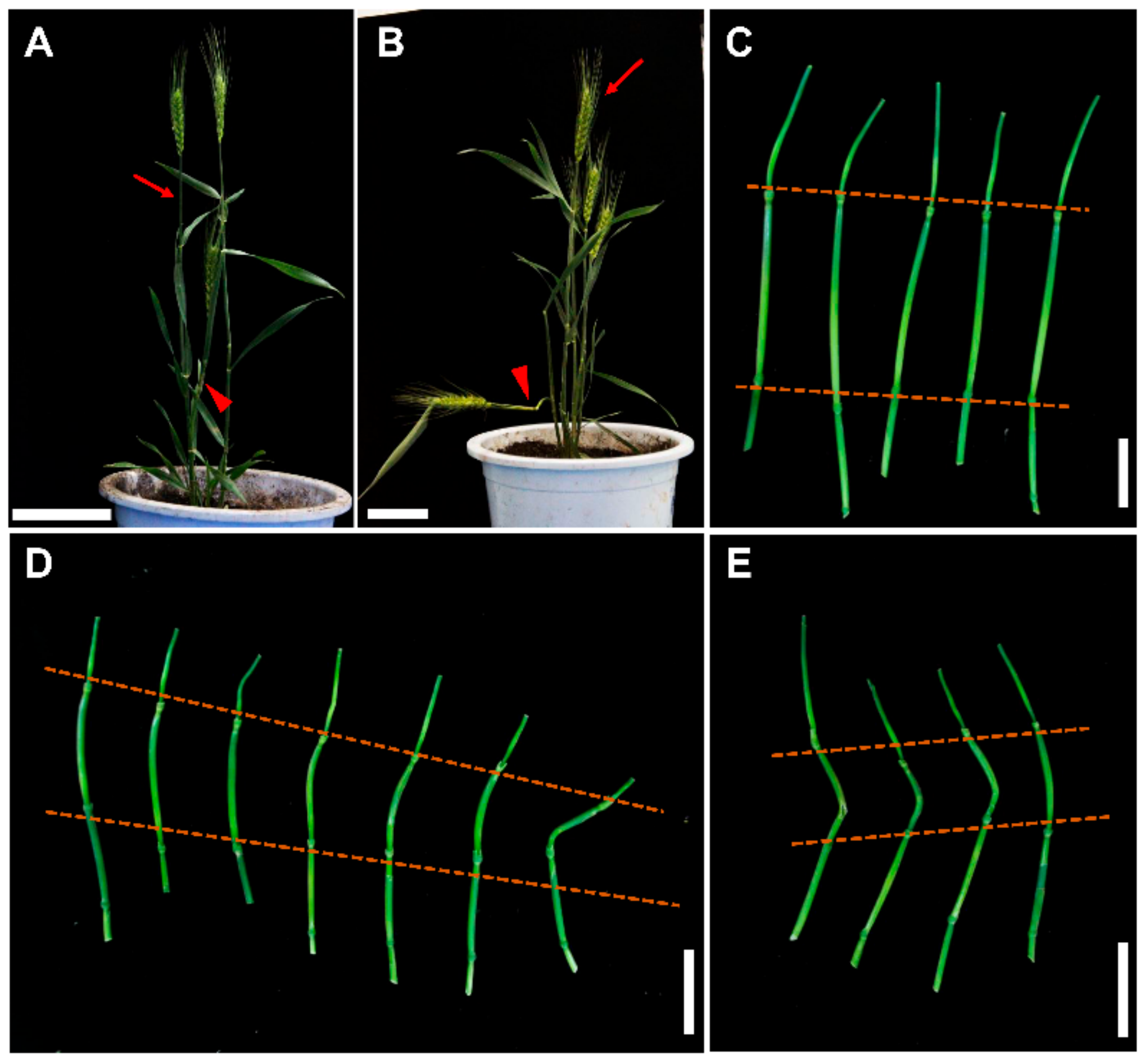
2.6. TaCLE3p and TaCLE34p Affect Wheat Root and Stem Development
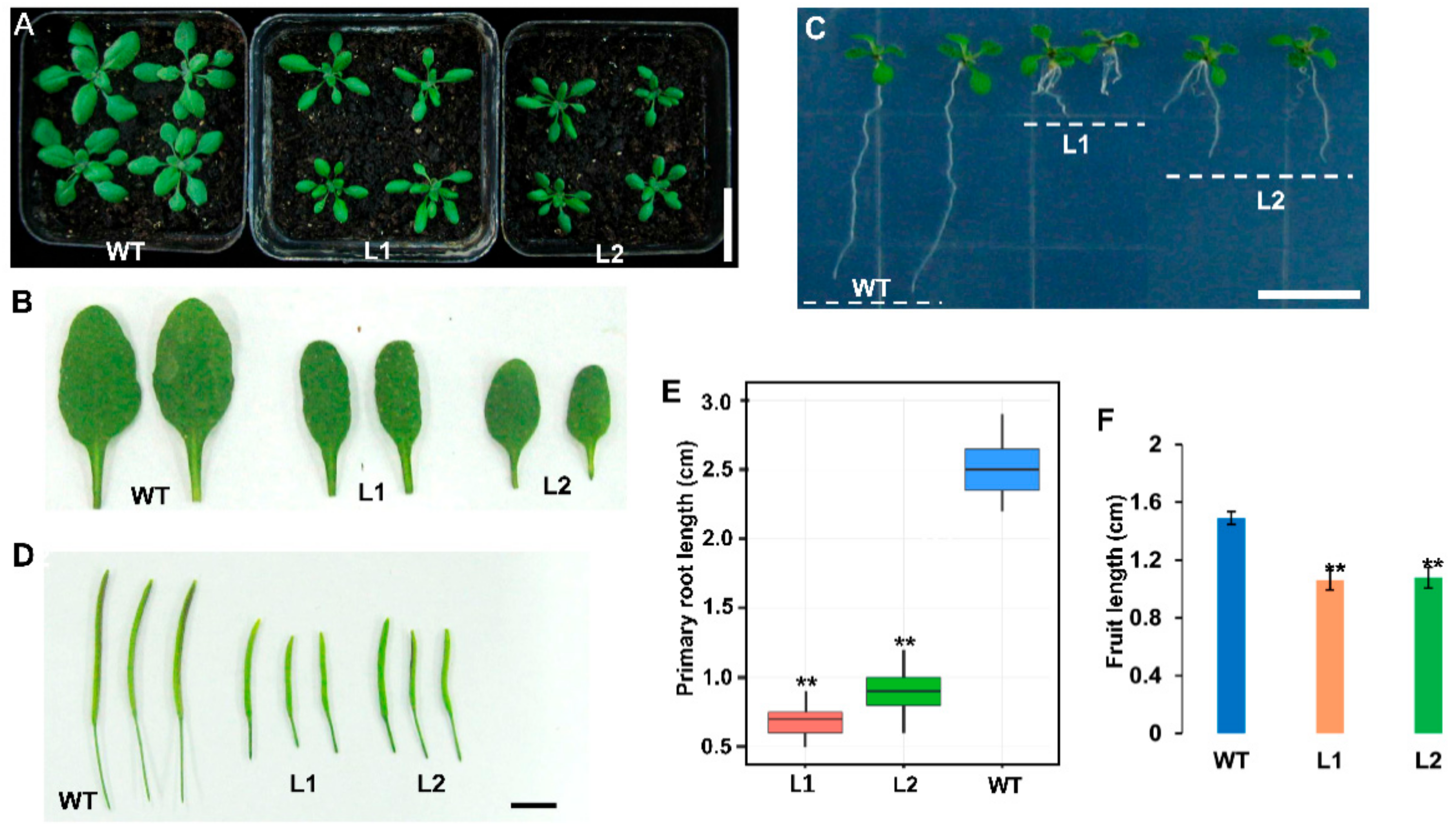
2.7. Overexpression Analysis of TaCLE3d in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization Analysis of TaCLE Genes and Gene Expression
4.2. Chromosomal Locations and Gene Duplication
4.3. Gene Structure and Protein Conserved Motifs Analysis
4.4. In Planta Peptide Treatment of Wheat Roots and Stems
4.5. Vector Constructs and Plant Transformation, Phenotypic Observation.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pearce, G.; Strydom, D.; Johnson, S.; Ryan, C.A. A polypeptide from tomato leaves induces wound-inducible proteinase inhibitor proteins. Science 1991, 253, 895–897. [Google Scholar] [PubMed]
- Lindsey, K.; Casson, S.; Chilley, P. Peptides: New signalling molecules in plants. Trends Plant Sci. 2002, 7, 78–83. [Google Scholar]
- Matsubayashi, Y. Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant Biol. 2014, 65, 385–413. [Google Scholar] [PubMed]
- Ferguson, B.J.; Mathesius, U. Phytohormone regulation of legume-rhizobia interactions. J. Chem. Ecol. 2014, 40, 770–790. [Google Scholar] [PubMed]
- Grienenberger, E.; Fletcher, J.C. Polypeptide signaling molecules in plant development. Curr. Opin. Plant Biol. 2015, 23, 8–14. [Google Scholar] [PubMed]
- Hastwell, A.H.; Gresshoff, P.M.; Ferguson, B.J. The structure and activity of nodulation-suppressing cle peptide hormones of legumes. Funct. Plant Biol. 2014, 42, 229–238. [Google Scholar]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Signaling of cell fate decisions by clavata3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar] [PubMed]
- Opsahl Ferstad, H.G.; Le, D.E.; Dumas, C.; Rogowsky, P.M.; Le, D.E. Zmesr, a novel endosperm-specific gene expressed in a restricted region around the maize embryo. Plant J. 2010, 12, 235–246. [Google Scholar]
- Cock, J.M.; Mccormick, S. A large family of genes that share homology with clavata3. Plant Physiol. 2001, 126, 939–942. [Google Scholar]
- Hastwell, A.H.; Gresshoff, P.M.; Ferguson, B.J. Genome-wide annotation and characterization of clavata/esr (cle) peptide hormones of soybean (Glycine max) and common bean (Phaseolus vulgaris), and their orthologues of Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 5271. [Google Scholar]
- Kinoshita, A.; Nakamura, Y.; Sasaki, E.; Kyozuka, J.; Fukuda, H.; Sawa, S. Gain-of-function phenotypes of chemically synthetic clavata3/esr-related (cle) peptides in Arabidopsis thaliana and Oryza sativa. Plant Cell Physiol. 2007, 48, 1821–1825. [Google Scholar] [PubMed]
- Oelkers, K.; Goffard, N.; Weiller, G.F.; Gresshoff, P.M.; Mathesius, U.; Frickey, T. Bioinformatic analysis of the cle signaling peptide family. BMC Plant Biol. 2008, 9, 17. [Google Scholar]
- Strabala, T.J.; O’Donnell, P.J.; Anne-Marie, S.; Charles, A.D.; E Jane, M.; Natalie, N.; Nieuwenhuizen, N.J.; Quinn, B.D.; Foote, H.C.C.; Hudson, K.R. Gain-of-function phenotypes of many clavata3/esr genes, including four new family members, correlate with tandem variations in the conserved clavata3/esr domain. Plant Physiol. 2006, 140, 1331–1344. [Google Scholar] [PubMed]
- Miyawaki, K.; Tabata, R.; Sawa, S. Evolutionarily conserved cle peptide signaling in plant development, symbiosis, and parasitism. Curr. Opin. Plant Biol. 2013, 16, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, M.; Shinohara, H.; Sakagami, Y.; Matsubayashi, Y. Arabidopsis clv3 peptide directly binds clv1 ectodomain. Science 2008, 319, 294. [Google Scholar] [PubMed]
- Müller, R.; Bleckmann, A.; Simon, R. The receptor kinase coryne of Arabidopsis transmits the stem cell–limiting signal clavata3 independently of clavata1. Plant Cell 2008, 20, 934–946. [Google Scholar] [CrossRef]
- Lenhard, M.; Laux, T. Stem cell homeostasis in the Arabidopsis shoot meristem is regulated by intercellular movement of clavata3 and its sequestration by clavata1. Development 2003, 130, 3163–3173. [Google Scholar] [PubMed]
- Kinoshita, A.; Betsuyaku, S.; Osakabe, Y.; Mizuno, S.; Nagawa, S.; Stahl, Y.; Simon, R.; Yamaguchi-Shinozaki, K.; Fukuda, H.; Sawa, S. Rpk2 is an essential receptor-like kinase that transmits the clv3 signal in Arabidopsis. Development 2010, 137, 3911–3920. [Google Scholar] [PubMed]
- Yingfang, Z.; Yuqing, W.; Ruili, L.; Xiufen, S.; Qinli, W.; Shanjin, H.; Bo, J.J.; Chun-Ming, L.; Jinxing, L. Analysis of interactions among the clavata3 receptors reveals a direct interaction between clavata2 and coryne in Arabidopsis. Plant J. 2010, 61, 223–233. [Google Scholar]
- Clark, S.E.; Williams, R.W.; Meyerowitz, E.M. The clavata1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 1997, 89, 575–585. [Google Scholar]
- Jeong, S.; Trotochaud, A.E.; Clark, S.E. The Arabidopsis clavata2 gene encodes a receptor-like protein required for the stability of the clavata1 receptor-like kinase. Plant Cell 1999, 11, 1925–1933. [Google Scholar] [PubMed]
- Bleckmann, A.; Weidtkamp-Peters, S.; Seidel, C.A.M.; Simon, R. Stem cell signaling in Arabidopsis requires crn to localize clv2 to the plasma membrane. Plant Physiol. 2010, 152, 166–176. [Google Scholar] [PubMed]
- Endo, S.; Betsuyaku, S.; Fukuda, H. Endogenous peptide ligand–receptor systems for diverse signaling networks in plants. Curr. Opin. Plant Biol. 2014, 21, 140–146. [Google Scholar] [PubMed]
- Depuydt, S.; Rodriguez-Villalon, A.; Santuari, L.; Wyser-Rmili, C.; Ragni, L.; Hardtke, C.S. Suppression of Arabidopsis protophloem differentiation and root meristem growth by cle45 requires the receptor-like kinase bam3. Proc. Natl. Acad. Sci. USA 2013, 110, 7074–7079. [Google Scholar] [PubMed]
- Fiers, M.; Golemiec, E.; Xu, J.; van der Geest, L.; Heidstra, R.; Stiekema, W.; Liu, C.-M. The 14–amino acid clv3, cle19, and cle40 peptides trigger consumption of the root meristem in Arabidopsis through a clavata2-dependent pathway. Plant Cell 2005, 17, 2542–2553. [Google Scholar] [PubMed]
- Scheible, W.-R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of Arabidopsis in response to nitrogen. Plant Physiol. 2004, 136, 2483–2499. [Google Scholar] [CrossRef] [PubMed]
- Araya, T.; Miyamoto, M.; Wibowo, J.; Suzuki, A.; Takahashi, H. Cle-clavata peptide-receptor signaling module regulates the expansion of plant root system in a nitrogen-dependent manner. Proc. Natl. Acad. Sci. USA 2014, 111, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Fiume, E.; Fletcher, J.C. Regulation of Arabidopsis embryo and endosperm development by the polypeptide signaling molecule cle8. Plant Cell 2012, 24, 1000–1012. [Google Scholar] [PubMed]
- Satoshi, E.; Hidefumi, S.; Yoshikatsu, M.; Hiroo, F. A novel pollen-pistil interaction conferring high-temperature tolerance during reproduction via cle45 signaling. Curr. Biol. 2013, 23, 1670–1676. [Google Scholar]
- Wang, S.; Lu, J.; Song, X.F.; Ren, S.C.; You, C.; Xu, J.; Liu, C.M.; Ma, H.; Chang, F. Cytological and transcriptomic analyses reveal important roles of cle19 in pollen exine formation. Plant Physiol. 2017, 175, 1186–1202. [Google Scholar]
- Xu, T.T.; Ren, S.C.; Song, X.F.; Liu, C.M. Cle19 expressed in the embryo regulates both cotyledon establishment and endosperm development in Arabidopsis. J. Exp. Bot. 2015, 66, 5217–5227. [Google Scholar] [PubMed]
- Yuki, H.; Hidefumi, S.; Yuki, K.; Asuka, I.; Ikuko, N.; Mari, O.; Shinichiro, S.; Kyoko, O.I.; Yoshikatsu, M.; Hiroo, F. Non-cell-autonomous control of vascular stem cell fate by a cle peptide/receptor system. Proc. Natl. Acad. Sci. USA 2008, 105, 15208–15213. [Google Scholar]
- Yasuko, I.; Ikuko, N.; Hiroyasu, M.; Kuninori, I.; Shinichiro, S.; Naoshi, D.; Hiroo, F. Dodeca-cle peptides as suppressors of plant stem cell differentiation. Science 2006, 313, 842–845. [Google Scholar]
- Yuki, H.; Yuki, K.; Hiroo, F. Tdif peptide signaling regulates vascular stem cell proliferation via the wox4 homeobox gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar]
- Strabala, T.J.; Phillips, L.; West, M.; Stanbra, L. Bioinformatic and phylogenetic analysis of the clavata3/embryo-surrounding region (cle) and the cle-like signal peptide genes in the pinophyta. BMC Plant Biol. 2014, 14, 47. [Google Scholar] [PubMed]
- Yu, Z.; Yang, S.; Song, Y.; Wang, J. Genome-wide characterization, expression and functional analysis of clv3/esr gene family in tomato. BMC Genom. 2014, 15, 827. [Google Scholar]
- Takuya, S.; Taiyo, T.; Masaru, F.; Nobuhiro, T.; Hidemi, K.; Hiro-Yuki, H. Conservation and diversification of meristem maintenance mechanism in Oryza sativa: Function of the floral organ number2 gene. Plant Cell Physiol. 2006, 47, 1591–1602. [Google Scholar]
- Takuya, S.; Makoto, S.; Motoyuki, A.; Masahiro, M.; Yasuo, N.; Hiro-Yuki, H. The gene floral organ number1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis clavata1. Development 2004, 131, 5649–5657. [Google Scholar]
- Suzaki, T.; Ohneda, M.; Toriba, T.; Yoshida, A.; Hirano, H.Y. Fon2 spare1 redundantly regulates floral meristem maintenance with floral organ number2 in rice. PLoS Genet. 2009, 5, e1000693. [Google Scholar]
- Ohmori, Y.; Tanaka, W.; Kojima, M.; Sakakibara, H.; Hirano, H.-Y. Wuschel-related homeobox4 is involved in meristem maintenance and is negatively regulated by the cle gene fcp1 in rice. Plant Cell 2013, 25, 229–241. [Google Scholar] [CrossRef]
- Yu, J.; Wang, J.; Lin, W.; Li, S.; Li, H.; Zhou, J.; Ni, P.; Dong, W.; Hu, S.; Zeng, C.; et al. The genomes of Oryza sativa: A history of duplications. PLoS Biol. 2005, 3, e38. [Google Scholar]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar]
- Tombuloglu, H.; Filiz, E.; Aydın, M.; Koc, I. Genome-wide identification and expression analysis of sulphate transporter (sultr) genes under sulfur deficiency in Brachypodium distachyon. J. Plant Biochem. Biotechnol. 2017, 26, 263–273. [Google Scholar] [CrossRef]
- Whitford, R.; Fernandez, A.; De Groodt, R.; Ortega, E.; Hilson, P. Plant cle peptides from two distinct functional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA 2008, 105, 18625–18630. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235. [Google Scholar] [PubMed]
- Pallakies, H.; Simon, R. The cle40 and crn/clv2 signaling pathways antagonistically control root meristem growth in Arabidopsis. Mol. Plant 2014, 7, 1619–1636. [Google Scholar] [PubMed]
- Qian, P.; Song, W.; Yokoo, T.; Minobe, A.; Wang, G.; Ishida, T.; Sawa, S.; Chai, J.; Kakimoto, T. The cle9/10 secretory peptide regulates stomatal and vascular development through distinct receptors. Nat. Plants 2018, 4, 1071–1081. [Google Scholar] [PubMed]
- Meng, L.; Feldman, L.J. Cle14/cle20 peptides may interact with clavata2/coryne receptor-like kinases to irreversibly inhibit cell division in the root meristem of Arabidopsis. Planta 2010, 232, 1061–1074. [Google Scholar] [PubMed]
- Gutiérrez-Alanís, D.; Yong-Villalobos, L.; Jiménez-Sandoval, P.; Alatorre-Cobos, F.; Oropeza-Aburto, A.; Mora-Macías, J.; Sánchez-Rodríguez, F.; Cruz-Ramírez, A.; Herrera-Estrella, L. Phosphate starvation-dependent iron mobilization induces cle14 expression to trigger root meristem differentiation through clv2/pepr2 signaling. Dev. Cell 2017, 41, 555–570. [Google Scholar] [CrossRef]
- Gregory, E.F.; Dao, T.Q.; Alexander, M.A.; Miller, M.J.; Fletcher, J.C. The signaling peptide-encoding genes cle16, cle17 and cle27 are dispensable for Arabidopsis shoot apical meristem activity. PLoS ONE 2018, 13, e0202595. [Google Scholar]
- Racolta, A.; Nodine, M.D.; Davies, K.; Lee, C.; Rowe, S.; Velazco, Y.; Wellington, R.; Tax, F.E. A common pathway of root growth control and response to cle peptides through two receptor kinases in Arabidopsis. Genetics 2017, 208, 300148. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.L.; Ishida, T.; Sawa, S. Cle peptides and their signaling pathways in plant development. J. Exp. Bot. 2016, 67, 4813–4826. [Google Scholar] [CrossRef]
- Song, X.F.; Yu, D.L.; Xu, T.T.; Ren, S.C.; Guo, P.; Liu, C.M. Contributions of individual amino acid residues to the endogenous clv3 function in shoot apical meristem maintenance in Arabidopsis. Mol. Plant 2012, 5, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Beitz, E. Texshade: Shading and labeling of multiple sequence alignments using latex2 epsilon. Bioinformatics 2000, 16, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the smart protein domain annotation resource. Nucleic Acids Res. 2017, 46, D493–D496. [Google Scholar] [CrossRef] [PubMed]
- Czyzewicz, N.; Shi, C.L.; Vu, L.D.; Van De Cotte, B.; Hodgman, C.; Butenko, M.A.; De Smet, I. Modulation of Arabidopsis and monocot root architecture by clavata3/embryo surrounding region 26 peptide. J. Exp. Bot. 2015, 66, 5229–5243. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium—Mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Liu, D.; Xia, Y.; Li, Z.; Niu, N.; Ma, S.; Wang, J.; Song, Y.; Zhang, G. Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Wheat. Int. J. Mol. Sci. 2019, 20, 4319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174319
Li Z, Liu D, Xia Y, Li Z, Niu N, Ma S, Wang J, Song Y, Zhang G. Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Wheat. International Journal of Molecular Sciences. 2019; 20(17):4319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174319
Chicago/Turabian StyleLi, Zheng, Dan Liu, Yu Xia, Ziliang Li, Na Niu, Shoucai Ma, Junwei Wang, Yulong Song, and Gaisheng Zhang. 2019. "Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION (CLE) Gene Family in Wheat" International Journal of Molecular Sciences 20, no. 17: 4319. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20174319