Expression and Distribution of the Auxin Response Factors in Sorghum bicolor During Development and Temperature Stress


Abstract
:1. Introduction
2. Results
2.1. Identification of SbARF Genes
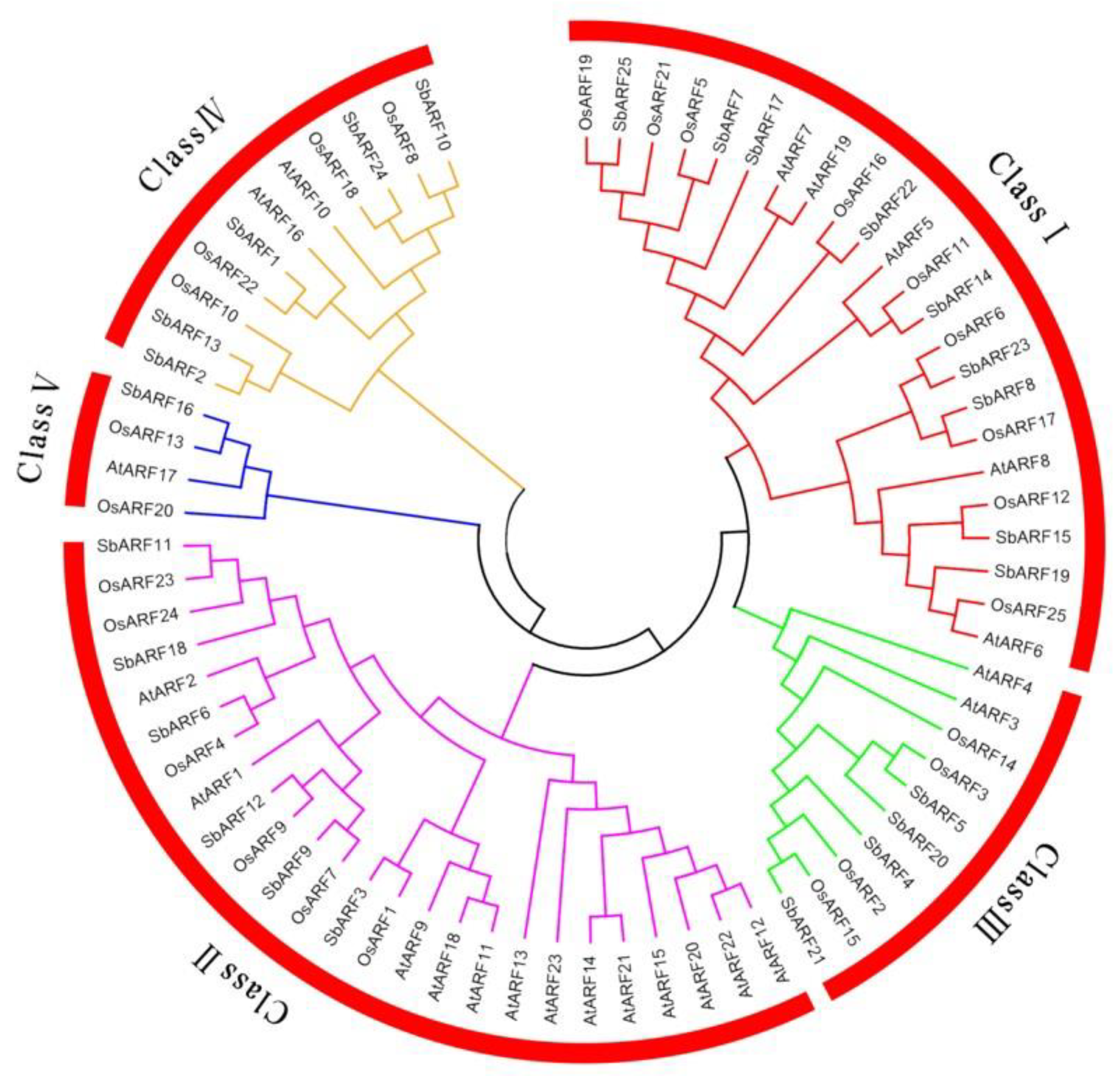
2.2. Phylogenetic Analysis and Classification of SbARF Genes
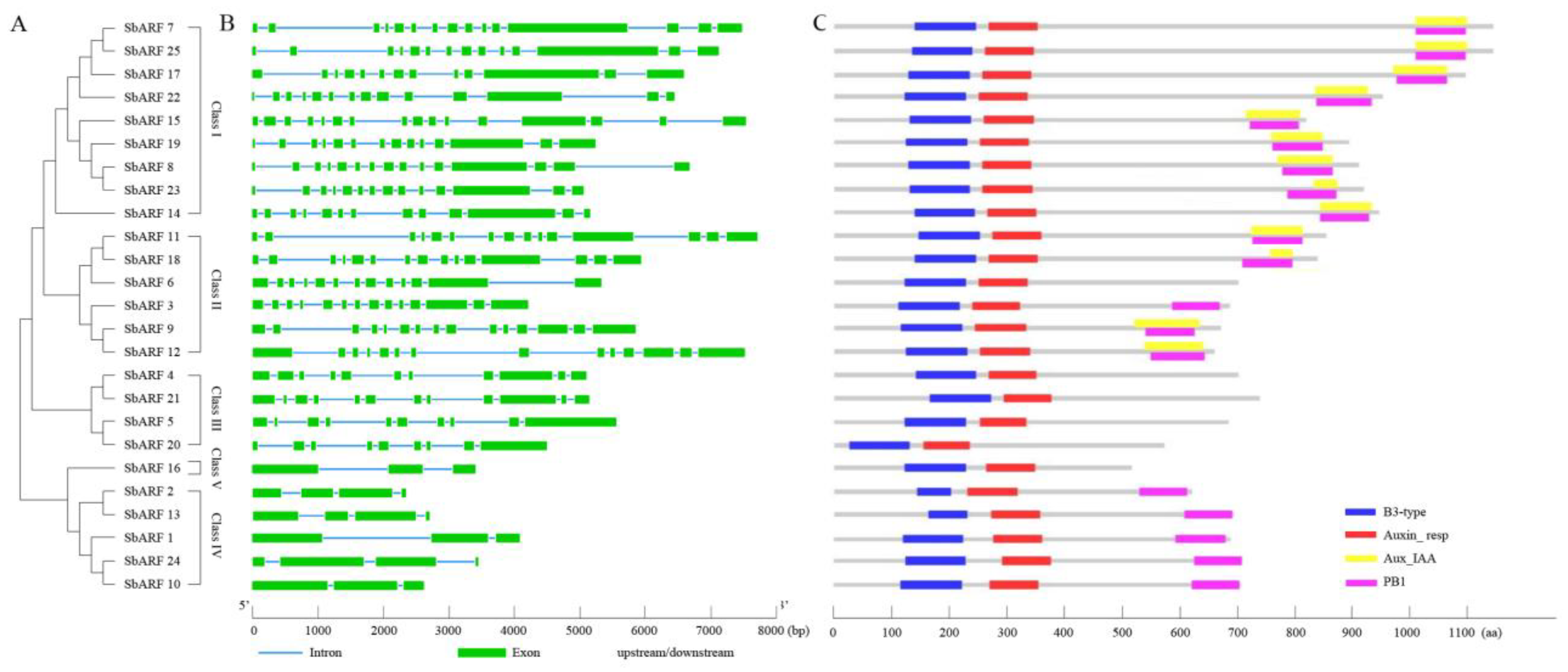
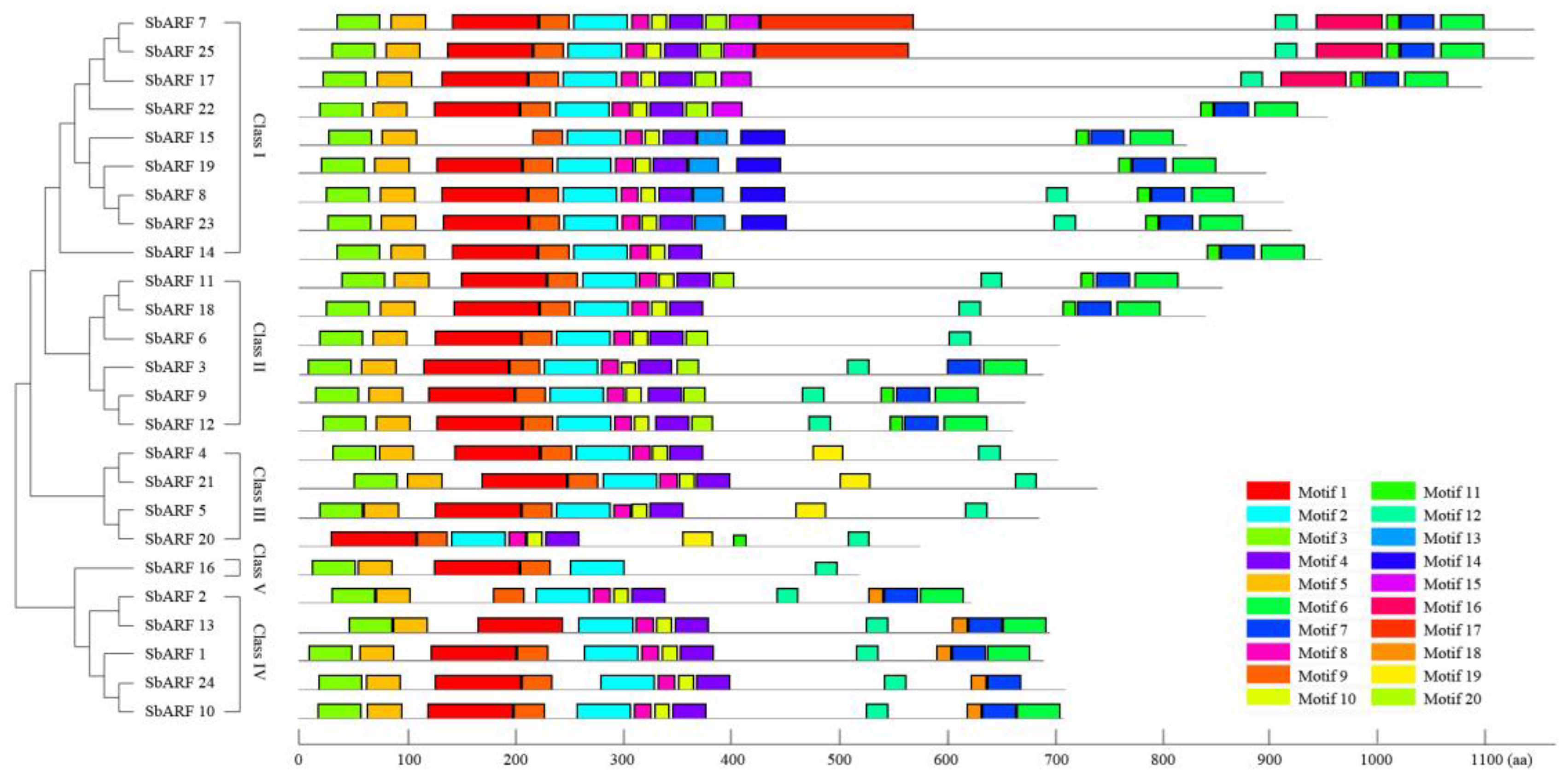
2.3. Gene Structure and Protein Structure of SbARF Genes
2.4. Expression of SbARF Genes in Different Organs and Tissues
2.5. Expression of SbARF Genes in Response to Temperature Stress
2.6. Distribution Patterns of the SbARF Genes under Temperature Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Phylogenetic Analysis of SbARF Genes
4.2. Gene Structure and Protein Structure Analysis
4.3. Plant Materials and Treatment Methods
4.4. Quantitative RT-PCR Analysis
4.5. Paraffin Section of Leaves and in Situ Hybridization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ARF | Auxin response factor |
| qRT-PCR | Reverse transcription-quantitative PCR |
| AuxREs | Auxin-response elements |
| DBD | N-terminal B3-type DNA binding domain |
| AD | Activation domain |
| RD | Repression domain |
| CTD | C-terminal domains |
| Aux/IAA | Auxin/indole-3-acetic acid |
| MW | Molecular weight |
| PI | Isoelectric point |
References
- Guilfoyle, T.J.; Hagen, G. Auxin response factors. Curr. Opin. Plant Biol. 2007, 10, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Liscum, E.; Reed, J.W. Genetics of Aux/IAA and ARF action in plant growth and development. Plant Mol. Biol. 2002, 49, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Harter, K.; Theologis, A. Protein-protein interactions among the Aux/IAA proteins. Proc. Natl. Acad. Sci. USA 1997, 94, 11786–11791. [Google Scholar] [CrossRef] [PubMed]
- Piya, S.; Shrestha, S.K.; Binder, B.; Stewart, C.N.; Hewezi, T. Protein-protein interaction and gene co-expression maps of ARFs and Aux/IAAs in Arabidopsis. Front. Plant Sci. 2014, 5, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. ARF1, a transcription factor that binds to auxin response elements. Science 1997, 276, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.K.; Pei, K.M.; Fu, Y.P.; Sun, Z.X.; Li, S.J.; Liu, H.Q.; Tang, K.; Han, B.; Tao, Y.Z. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene 2007, 394, 13–24. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, H.Y.; Chen, W.J.; Qian, Y.X.; Ma, Q.; Cheng, B.J.; Zhu, S.W. Genome-wide analysis of the auxin response factor (ARF) gene family in maize (Zea mays). Plant Growth Regul 2011, 63, 225–234. [Google Scholar] [CrossRef]
- Die, J.V.; Gil, J.; Millan, T. Genome-wide identification of the auxin response factor gene family in Cicer arietinum. BMC Genom. 2018, 19, 301. [Google Scholar] [CrossRef]
- Liu, M.Y.; Ma, Z.T.; Wang, A.H.; Zheng, T.R.; Huang, L.; Sun, W.J.; Zhang, Y.J.; Jin, W.Q.; Zhan, J.Y.; Cai, Y.T.; et al. Genome-Wide Investigation of the Auxin Response Factor Gene Family in Tartary Buckwheat (Fagopyrum tataricum). Int. J. Mol. Sci. 2018, 19, 3526. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Zeng, Z.H.; Chen, C.J.; Li, C.Q.; Xia, R.; Li, J.G. Genome-wide characterization of the auxin response factor (ARF) gene family of litchi (Litchi chinensis Sonn.): Phylogenetic analysis, miRNA regulation and expression changes during fruit abscission. Peerj 2019, 7, e6677. [Google Scholar] [CrossRef]
- Ellis, C.M.; Nagpal, P.; Young, J.C.; Hagen, G.; Guilfoyle, T.J.; Reed, J.W. AUXIN RESPONSE FACTOR1 and AUXIN RESPONSE FACTOR2 regulate senescence and floral organ abscission in Arabidopsis thaliana. Development 2005, 132, 4563–4574. [Google Scholar] [CrossRef] [PubMed]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hua, D.P.; He, J.N.; Duan, Y.; Chen, Z.Z.; Hong, X.H.; Gong, Z.Z. Auxin Response Factor2 (ARF2) and its regulated homeodomain gene HB33 mediate abscisic acid response in Arabidopsis. PLoS Genet. 2011, 7, e1002172. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.S.; Wang, Z.B.; Yao, S.Q.; Liu, A.Z. The ARF2-ANT-COR15A gene cascade regulates ABA-signaling-mediated resistance of large seeds to drought in Arabidopsis. J. Cell Sci. 2015, 128, 3922–3932. [Google Scholar] [CrossRef] [PubMed]
- Promchuea, S.; Zhu, Y.J.; Chen, Z.Z.; Zhang, J.; Gong, Z.Z. ARF2 coordinates with PLETHORAs and PINs to orchestrate ABA-mediated root meristem activity in Arabidopsis. J. Integr. Plant Biol. 2017, 59, 30–43. [Google Scholar] [CrossRef]
- Nemhauser, J.L.; Feldman, L.J.; Zambryski, P.C. Auxin and ETTIN in Arabidopsis gynoecium morphogenesis. Development 2000, 127, 3877–3888. [Google Scholar] [PubMed]
- Chitwood, D.H.; Nogueira, F.T.S.; Howell, M.D.; Montgomery, T.A.; Carrington, J.C.; Timmermans, M.C.P. Pattern formation via small RNA mobility. Gene. Dev. 2009, 23, 549–554. [Google Scholar] [CrossRef] [Green Version]
- Tantikanjana, T.; Nasrallah, J.B. Non-cell-autonomous regulation of crucifer self-incompatibility by Auxin Response Factor ARF3. Proc. Natl. Acad. Sci. USA 2012, 109, 19468–19473. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Wang, L.; Sun, W.; Zhang, Y.; Zhou, C.; Su, Y.H.; Li, W.; Sun, T.T.; Zhao, X.Y.; Li, X.G.; et al. Pattern of Auxin and Cytokinin Responses for Shoot Meristem Induction Results from the Regulation of Cytokinin Biosynthesis by AUXIN RESPONSE FACTOR3. Plant Physiol 2013, 161, 240–251. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, K.; Guo, L.; Liu, X.; Zhang, Z. AUXIN RESPONSE FACTOR3 plays distinct role during early flower development. Plant Signal. Behav. 2018, 13, e1467690. [Google Scholar] [CrossRef]
- Zhang, K.; Wang, R.Z.; Zi, H.L.; Li, Y.P.; Cao, X.W.; Li, D.M.; Guo, L.; Tong, J.H.; Pan, Y.Y.; Jiao, Y.L.; et al. AUXIN RESPONSE FACTOR3 regulates floral meristem determinacy by repressing cytokinin biosynthesis and signaling. Plant Cell 2018, 30, 324–346. [Google Scholar] [CrossRef] [PubMed]
- Hardtke, C.S.; Berleth, T. The Arabidopsis gene MONOPTEROS encodes a transcription factor mediating embryo axis formation and vascular development. EMBO J. 1998, 17, 1405–1411. [Google Scholar] [CrossRef] [PubMed]
- Krogan, N.T.; Ckurshumova, W.; Marcos, D.; Caragea, A.E.; Berleth, T. Deletion of MP/ARF5 domains III and IV reveals a requirement for Aux/IAA regulation in Arabidopsis leaf vascular patterning. New Phytol. 2012, 194, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.N.; Miao, L.M.; Huo, R.X.; Song, X.Y.; Johnson, C.; Kong, L.J.; Sundaresan, V.; Yu, X.L. ARF2-ARF4 and ARF5 are essential for female and male gametophyte development in Arabidopsis. Plant Cell Physiol. 2018, 59, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; He, S.Z.; Zhai, H.; Li, R.J.; Zhao, N.; Liu, Q.C. A sweetpotato auxin response factor gene (IbARF5) is involved in carotenoid biosynthesis and salt and drought tolerance in transgenic Arabidopsis. Front. Plant Sci. 2018, 9, 1307. [Google Scholar] [CrossRef] [PubMed]
- Harper, R.M.; Stowe-Evans, E.L.; Luesse, D.R.; Muto, H.; Tatematsu, K.; Watahiki, M.K.; Yamamoto, K.; Liscum, E. The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell 2000, 12, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Goetz, M.; Vivian-Smith, A.; Johnson, S.D.; Koltunow, A.M. AUXIN RESPONSE FACTOR8 is a negative regulator of fruit initiation in Arabidopsis. Plant Cell 2006, 18, 1873–1886. [Google Scholar] [CrossRef]
- Tian, C.; Muto, H.; Higuchi, K.; Matamura, T.; Tatematsu, K.; Koshiba, T.; Yamamoto, K.T. Disruption and overexpression of auxin response factor 8 gene of Arabidopsis affect hypocotyl elongation and root growth habit, indicating its possible involvement in auxin homeostasis in light condition. Plant J. 2004, 40, 333–343. [Google Scholar] [CrossRef]
- Waller, F.; Furuya, M.; Nick, P. OsARF1, an auxin response factor from rice, is auxin-regulated and classifies as a primary auxin responsive gene. Plant Mol. Biol. 2002, 50, 415–425. [Google Scholar] [CrossRef]
- Attia, K.A.; Abdelkhalik, A.F.; Ammar, M.H.; Wei, C.; Yang, J.; Lightfoot, D.A.; El-Sayed, W.M.; El-Shemy, H.A. Antisense Phenotypes Reveal a Functional Expression of OsARF1, an Auxin Response Factor, in Transgenic Rice. Curr. Issues Mol. Biol 2009, 11, I29–I34. [Google Scholar]
- Ding, Y.H.; Ma, Y.Z.; Liu, N.A.; Xu, J.; Hu, Q.; Li, Y.Y.; Wu, Y.L.; Xie, S.; Zhu, L.F.; Min, L.; et al. microRNAs involved in auxin signalling modulate male sterility under high-temperature stress in cotton (Gossypium hirsutum). Plant J. 2017, 91, 977–994. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Liu, H.B.; Xiong, L.Z. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mace, E.S.; Tai, S.S.; Gilding, E.K.; Li, Y.H.; Prentis, P.J.; Bian, L.L.; Campbell, B.C.; Hu, W.S.; Innes, D.J.; Han, X.L.; et al. Whole-genome sequencing reveals untapped genetic potential in Africa’s indigenous cereal crop sorghum. Nat. Commun. 2013, 4, 2320. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.K.; Bai, Y.H.; Shen, C.J.; Wu, Y.R.; Zhang, S.N.; Jiang, D.A.; Guilfoyle, T.J.; Chen, M.; Qi, Y.H. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Li, S.B.; Xie, Z.Z.; Hu, C.G.; Zhang, J.Z. A Review of Auxin Response Factors (ARFs) in Plants. Front. Plant Sci. 2016, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Roosjen, M.; Paque, S.; Weijers, D. Auxin Response Factors: Output control in auxin biology. J. Exp. Bot. 2018, 69, 179–188. [Google Scholar] [CrossRef]
- Qi, Y.H.; Wang, S.K.; Shen, C.J.; Zhang, S.N.; Chen, Y.; Xu, Y.X.; Liu, Y.; Wu, Y.R.; Jiang, D.A. OsARF12, a transcription activator on auxin response gene, regulates root elongation and affects iron accumulation in rice (Oryza sativa). New Phytol. 2012, 193, 109–120. [Google Scholar] [CrossRef]
- Wang, S.K.; Zhang, S.N.; Sun, C.D.; Xu, Y.X.; Chen, Y.; Yu, C.L.; Qian, Q.; Jiang, D.A.; Qi, Y.H. Auxin response factor (OsARF12), a novel regulator for phosphate homeostasis in rice (Oryza sativa). New Phytol. 2014, 201, 91–103. [Google Scholar] [CrossRef]
- Shen, C.J.; Wang, S.K.; Zhang, S.N.; Xu, Y.X.; Qian, Q.; Qi, Y.H.; Jiang, D.A. OsARF16, a transcription factor, is required for auxin and phosphate starvation response in rice (Oryza sativa L.). Plant Cell Env. 2013, 36, 607–620. [Google Scholar] [CrossRef]
- Adamowski, M.; Friml, J. PIN-Dependent Auxin Transport: Action, Regulation, and Evolution. Plant Cell 2015, 27, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Bennett, T.; Hines, G.; van Rongen, M.; Waldie, T.; Sawchuk, M.G.; Scarpella, E.; Ljung, K.; Leyser, O. Connective Auxin Transport in the Shoot Facilitates Communication between Shoot Apices. PLoS Biol. 2016, 14, e1002446. [Google Scholar] [CrossRef] [PubMed]
- Kohlen, W.; Ng, J.L.P.; Deinum, E.E.; Mathesius, U. Auxin transport, metabolism, and signalling during nodule initiation: Indeterminate and determinate nodules. J. Exp. Bot. 2018, 69, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Scarpella, E.; Marcos, D.; Friml, J.; Berleth, T. Control of leaf vascular patterning by polar auxin transport. Gene Dev. 2006, 20, 1015–1027. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Gribskov, M. Combining evidence using p-values: Application to sequence homology searches. Bioinformatics 1998, 14, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.P.; Cao, X.L.; Shi, S.D.; Ma, Y.L.; Wang, K.; Liu, S.J.; Chen, D.; Chen, Q.; Ma, H.L. Genome-wide survey of Aux/IAA gene family members in potato (Solanum tuberosum): Identification, expression analysis, and evaluation of their roles in tuber development. Biochem. Biophys. Res. Commun. 2016, 471, 320–327. [Google Scholar] [CrossRef]
- Deng, Y.T.; Zou, W.X.; Li, G.; Zhao, J. TRANSLOCASE OF THE INNER MEMBRANE9 and 10 Are Essential for Maintaining Mitochondrial Function during Early Embryo Cell and Endosperm Free Nucleus Divisions in Arabidopsis. Plant Physiol. 2014, 166, 853–868. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SbARF gene | Sequence ID | Chr.Location | Strand Direction | ORF Length (bp) | Transcript Length (bp) | No. Exons | No. Coding Exons | UniProt | Protein Length (aa) | MW (Da) | PI | Domains |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SbARF1 | Sb01g019130.1 | Chr1:20265083–20269209 | Reverse | 2070 | 2354 | 3 | 3 | C5WYD5 | 689 | 75,404.92 | 6.9 | DBD, MR, PB1 |
| SbARF2 | Sb02g032210.1 | Chr2:66973093–66975465 | Reverse | 1869 | 1869 | 4 | 4 | C5X7P6 | 622 | 68,290.21 | 8.7 | DBD, MR, PB1 |
| SbARF3 | Sb03g000530.1 | Chr3:375293–379543 | Reverse | 2067 | 2712 | 14 | 14 | C5XJJ7 | 688 | 76,895.05 | 6.4 | DBD, MR, PB1 |
| SbARF4 | Sb03g030740.1 | Chr3:58980057–58985203 | Reverse | 2109 | 2418 | 11 | 10 | C5XG18 | 702 | 77,003.66 | 6.4 | DBD, MR |
| SbARF5 | Sb03g034850.1 | Chr3:62997617–63003214 | Reverse | 2058 | 2584 | 10 | 10 | ─ | 685 | 74,377.98 | 7.2 | DBD, MR |
| SbARF6 | Sb03g044630.1 | Chr3:71975563–71980937 | Forward | 2115 | 2729 | 13 | 13 | C5XH00 | 704 | 78,702.14 | 7.4 | DBD, MR |
| SbARF7 | Sb04g003240.1 | Chr4:3072716–3080254 | Reverse | 3432 | 3836 | 15 | 14 | C5XUJ9 | 1143 | 127,041.26 | 6.3 | DBD, MR, CTD, PB1 |
| SbARF8 | Sb04g004430.1 | Chr4:4219215–4225954 | Forward | 2736 | 3126 | 15 | 14 | C5XVH8 | 911 | 100,417.56 | 5.7 | DBD, MR, CTD, PB1 |
| SbARF9 | Sb04g022830.1 | Chr4:52419119–52425028 | Reverse | 2019 | 2809 | 15 | 14 | C5XUU5 | 672 | 74,930.18 | 6.2 | DBD, MR, CTD, PB1 |
| SbARF10 | Sb04g026610.1 | Chr4:56508838..56511492 | Forward | 2127 | 2487 | 3 | 3 | C5XXU7 | 708 | 76,087.88 | 7.1 | DBD, MR, PB1 |
| SbARF11 | Sb05g019540.1 | Chr5:47874380–47882162 | Reverse | 2568 | 3071 | 15 | 14 | ─ | 855 | 93,782.34 | 6.5 | DBD, MR, CTD, PB1 |
| SbARF12 | Sb06g017490.1 | Chr6:46853929–46861508 | Forward | 1986 | 3151 | 14 | 14 | C5Y8U9 | 661 | 73,201.76 | 6.1 | DBD, MR, CTD, PB1 |
| SbARF13 | Sb06g022810.1 | Chr6:52026442–52029170 | Forward | 2088 | 2088 | 4 | 4 | C5YCE3 | 695 | 75,065.79 | 8.2 | DBD, MR, PB1 |
| SbARF14 | Sb06g031900.1 | Chr6:60209330–60214539 | Reverse | 2841 | 2841 | 13 | 13 | C5Y9Z0 | 946 | 103,749.51 | 6.2 | DBD, MR, CTD, PB1 |
| SbARF15 | Sb06g032500.1 | Chr6:60654734–60662333 | Forward | 2466 | 3084 | 16 | 14 | C5YA53 | 821 | 90,757.87 | 6.5 | DBD, MR, CTD, PB1 |
| SbARF16 | Sb06g033970.1 | Chr6:61857871–61861319 | Reverse | 1557 | 1928 | 3 | 2 | C5YB36 | 518 | 56,284.76 | 6 | DBD, MR |
| SbARF17 | Sb07g027080.1 | Chr7:62218509–62225164 | Reverse | 3288 | 3716 | 13 | 13 | C5YIB6 | 1095 | 121,248.57 | 6.5 | DBD, MR, CTD, PB1 |
| SbARF18 | Sb08g014320.1 | Chr8:37871039–37877027 | Reverse | 2520 | 3050 | 15 | 14 | C5YNM8 | 839 | 92,454.03 | 6.7 | DBD, MR, CTD, PB1 |
| SbARF19 | Sb08g021460.1 | Chr8:53122868–53128155 | Reverse | 2688 | 3063 | 14 | 14 | C5YRZ9 | 895 | 98,609.5 | 5.9 | DBD, MR, CTD, PB1 |
| SbARF20 | Sb09g025500.1 | Chr9:54931364–54935901 | Forward | 1728 | 2002 | 9 | 8 | C5Z0X8 | 575 | 63,169.94 | 7.5 | DBD, MR |
| SbARF21 | Sb09g028450.1 | Chr9:57360258–57365448 | Forward | 2220 | 2483 | 12 | 11 | C5YVJ4 | 739 | 80,854.09 | 7.7 | DBD, MR |
| SbARF22 | Sb10g006440.1 | Chr10:5930430–5936935 | Forward | 2859 | 2859 | 14 | 14 | C5Z600 | 952 | 105,068.55 | 6.4 | DBD, MR, CTD, PB1 |
| SbARF23 | Sb10g027220.1 | Chr10:56924176–56929286 | Reverse | 2760 | 2760 | 14 | 14 | C5Z7U5 | 919 | 101,692.05 | 6.4 | DBD, MR, CTD, PB1 |
| SbARF24 | Sb10g027790.1 | Chr10:57623853–57627338 | Forward | 2130 | 2493 | 4 | 3 | C5Z8A5 | 709 | 76,700.16 | 7.7 | DBD, MR, PB1 |
| SbARF25 | Sb10g029130.1 | Chr10:58946685–58953873 | Forward | 3432 | 3574 | 14 | 14 | C5Z981 | 1143 | 126,297.21 | 6.6 | DBD, MR, CTD, PB1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, D.; Wang, W.; Wu, Y.; Xie, H.; Zhao, L.; Zeng, Q.; Zhan, Y. Expression and Distribution of the Auxin Response Factors in Sorghum bicolor During Development and Temperature Stress. Int. J. Mol. Sci. 2019, 20, 4816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194816
Chen D, Wang W, Wu Y, Xie H, Zhao L, Zeng Q, Zhan Y. Expression and Distribution of the Auxin Response Factors in Sorghum bicolor During Development and Temperature Stress. International Journal of Molecular Sciences. 2019; 20(19):4816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194816
Chicago/Turabian StyleChen, Dan, Weian Wang, Yaqin Wu, Hui Xie, Linfei Zhao, Qi Zeng, and Yonghua Zhan. 2019. "Expression and Distribution of the Auxin Response Factors in Sorghum bicolor During Development and Temperature Stress" International Journal of Molecular Sciences 20, no. 19: 4816. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20194816